Apitherapy for Age-Related Skeletal Muscle Dysfunction (Sarcopenia): A Review on the Effects of Royal Jelly, Propolis, and Bee Pollen



Abstract
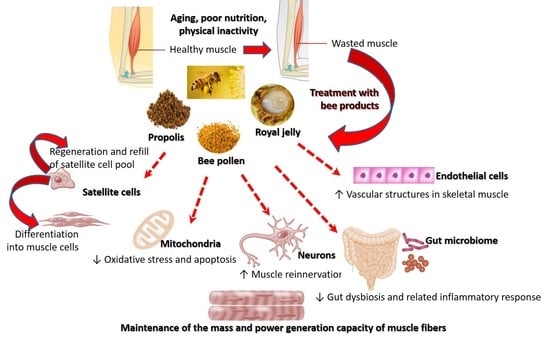
1. Introduction
2. Apitherapy as a Possible Complementary Treatment for Sarcopenia
2.1. Royal Jelly: Its Constituents, Biological, and Pharmacological Activities
2.2. Propolis: Its Constituents, Biological, and Pharmacological Activities
2.3. Bee Pollen: Its Constituents, Biological, and Pharmacological Activities
2.4. Safety Profile of Royal Jelly, Propolis, and Bee Pollen
3. Evidence of Anti-Sarcopenia Effects of Bee Products from Preclinical and Clinical Studies
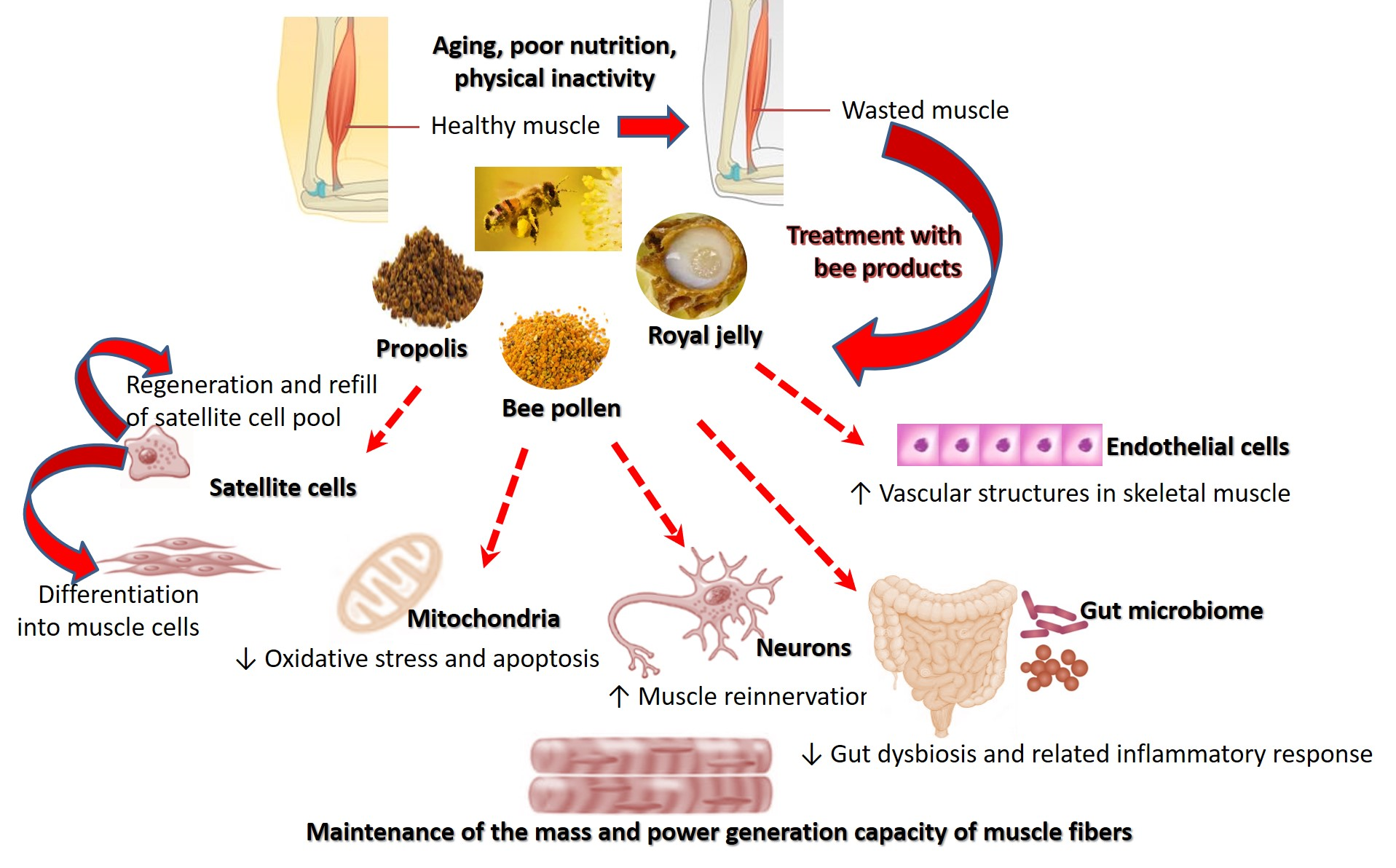
4. Mechanisms of Action of Royal Jelly, Bee Pollen, and Propolis in Sarcopenia
4.1. Modulating Inflammatory Responses in Skeletal Muscle
4.2. Counteracting Oxidative Stress in Skeletal Muscle
4.3. Metabolic Regulation
4.4. Enhancement of Muscle Protein Synthesis
4.5. Suppression of Catabolic Activity in Skeletal Muscle
4.6. Enhancement of Stem Cell Function
4.7. Counteracting Glycation Stress
4.8. Neuronal Regeneration
4.9. Improving Muscular Blood Supply
4.10. Improving the Composition of Gut Microbiome
5. Discussion
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 4eBP1/eIF4E | Eukaryotic translation initiation 4E-binding protein 1 |
| 10-HDA | trans-10-hydroxy-2-decenoic acid |
| AGEs | Advanced glycation end products |
| AKT | A serine/threonine nutrient sensing protein kinase |
| AMP | Adenosine monophosphate |
| AMPK | Adenosine monophosphate activated protein kinase |
| ARE | Antioxidant response element |
| ATF4 | Activating transcription factor-4 |
| ATP | Adenosine tri-phosphate |
| COX | Cyclooxygenase |
| eNOs | Endothelial nitric oxide synthase |
| ER | Endoplasmic reticulum |
| FOXO | Forkhead box O |
| GI | Gastrointestinal |
| GLUT4 | Glucose transporter 4 |
| HET | Heterogeneous head tilt |
| HFD | High fat diet |
| HIF | Hypoxia-inducible factor |
| HO-1 | Heme oxygenase 1 |
| HOMA-IR | Homeostatic model assessment of insulin resistance |
| IKK | IκB kinase |
| IGFs | Insulin-like growth factors |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| IR | Insulin receptor |
| LESM | Lower extremity skeletal muscle mass |
| MAPK | Mitogen-activated protein kinase |
| MDA | Malondialdehyde |
| mTOR | Mammalian target of rapamycin |
| MMP-12 | Metalloproteinase-12 |
| MCP-1 | Monocyte chemotactic protein 1 |
| MRJPs | Major royal jelly proteins |
| MyoD | Myogenic differentiation 1 |
| NF-kB | Nuclear factor kappa-B |
| NRF2 | Nuclear factor erythroid 2/Nuclear respiratory factor 2 |
| PGC-1α | Peroxisome proliferator-activated receptor gamma coactivator 1 alpha |
| PI3k | Phosphoinositide-3 kinase |
| PUFA | Polyunsaturated fatty acids |
| PKC | Protein kinase C |
| pRJ | Protease-treated royal jelly |
| QOL | Quality of life |
| RAGE | Receptor for Advanced Glycation End products |
| RCT | Randomized control trial |
| ROS | Reactive oxygen species |
| SDs | Standard deviations |
| S6K | P70 ribosomal proteins S6 kinase |
| SMART | Specific of Muscle Atrophy and Regulated by Transcription |
| TNF | Tumor necrosis factor |
| TNFR1 | Tumor necrosis factor receptor 1 |
| VEGF-A | Vascular endothelial growth factor A |
| VSMCs | Vascular smooth muscle cells |
References
- Miljkovic, N.; Lim, J.Y.; Miljkovic, I.; Frontera, W.R. Aging of skeletal muscle fibers. Ann. Rehabil. Med. 2015, 39, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Kunugi, H.; Ali, A.M. Royal Jelly and Its Components Promote Healthy Aging and Longevity: From Animal Models to Humans. Int. J. Mol. Sci. 2019, 20, 4662. [Google Scholar] [CrossRef] [PubMed]
- Welch, A.A.; Hayhoe, R.P.G.; Cameron, D. The relationships between sarcopenic skeletal muscle loss during ageing and macronutrient metabolism, obesity and onset of diabetes. Proc. Nutr. Soc. 2020, 79, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Keller, K. Sarcopenia. Wien. Med. Wochenschr. 2019, 169, 157–172. [Google Scholar] [CrossRef]
- Wilkinson, D.J.; Piasecki, M.; Atherton, P.J. The age-related loss of skeletal muscle mass and function: Measurement and physiology of muscle fibre atrophy and muscle fibre loss in humans. Ageing Res. Rev. 2018, 47, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Aran, L.; Bulli, G.; Curcio, F.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Sarcopenia: Assessment of disease burden and strategies to improve outcomes. Clin. Interv. Aging 2018, 13, 913–927. [Google Scholar] [CrossRef] [PubMed]
- Perkisas, S.; Vandewoude, M. Where frailty meets diabetes. Diabetes Metab. Res. 2016, 32, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Vandewoude, M.F.; Alish, C.J.; Sauer, A.C.; Hegazi, R.A. Malnutrition-sarcopenia syndrome: Is this the future of nutrition screening and assessment for older adults? J. Aging Res. 2012, 2012, 651570. [Google Scholar] [CrossRef]
- O’Toole, P.W.; Shiels, P.G. The role of the microbiota in sedentary lifestyle disorders and ageing: Lessons from the animal kingdom. J. Intern. Med. 2020, 287, 271–282. [Google Scholar] [CrossRef]
- Bleau, C.; Karelis, A.D.; St-Pierre, D.H.; Lamontagne, L. Crosstalk between intestinal microbiota, adipose tissue and skeletal muscle as an early event in systemic low-grade inflammation and the development of obesity and diabetes. Diabetes Metab. Res. Rev. 2015, 31, 545–561. [Google Scholar] [CrossRef]
- Zheng, C.; Huang, W.Y.; Sheridan, S.; Sit, C.H.; Chen, X.K.; Wong, S.H. COVID-19 Pandemic Brings a Sedentary Lifestyle in Young Adults: A Cross-Sectional and Longitudinal Study. Int. J. Environ. Res. Public Health 2020, 17, 6035. [Google Scholar] [CrossRef] [PubMed]
- Langhammer, B.; Sagbakken, M.; Kvaal, K.; Ulstein, I.; Nåden, D.; Rognstad, M.K. Music Therapy and Physical Activity to Ease Anxiety, Restlessness, Irritability, and Aggression in Individuals With Dementia With Signs of Frontotemporal Lobe Degeneration. J. Psychosoc. Nurs. Ment. Health Serv. 2019, 57, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Zhang, Y.; Zhang, D. Sedentary behaviour and the risk of depression: A meta-analysis. Br. J. Sports Med. 2014, 49, 1–6. [Google Scholar] [CrossRef]
- Bhasker, A.G.; Greve, J.W. Are Patients Suffering from Severe Obesity Getting a Raw Deal During COVID-19 Pandemic? Obes. Surg. 2020, 1–2. [Google Scholar] [CrossRef]
- Mori, H.; Kuroda, A.; Ishizu, M.; Ohishi, M.; Takashi, Y.; Otsuka, Y.; Taniguchi, S.; Tamaki, M.; Kurahashi, K.; Yoshida, S.; et al. Association of accumulated advanced glycation end-products with a high prevalence of sarcopenia and dynapenia in patients with type 2 diabetes. J. Diabetes Investig. 2019, 10, 1332–1340. [Google Scholar] [CrossRef]
- Gensous, N.; Bacalini, M.G.; Franceschi, C.; Meskers, C.G.M.; Maier, A.B.; Garagnani, P. Age-Related DNA Methylation Changes: Potential Impact on Skeletal Muscle Aging in Humans. Front. Physiol. 2019, 10, 996. [Google Scholar] [CrossRef]
- Rong, S.; Wang, L.; Peng, Z.; Liao, Y.; Li, D.; Yang, X.; Nuessler, A.K.; Liu, L.; Bao, W.; Yang, W. The mechanisms and treatments for sarcopenia: Could exosomes be a perspective research strategy in the future? J. Cachexia Sarcopenia Muscle 2020, 11, 348–365. [Google Scholar] [CrossRef] [PubMed]
- Laviano, A.; Koverech, A.; Zanetti, M. Nutrition support in the time of SARS-CoV-2 (COVID-19). Nutrition 2020, 74, 110834. [Google Scholar] [CrossRef] [PubMed]
- Rooks, D.; Roubenoff, R. Development of Pharmacotherapies for the Treatment of Sarcopenia. J. Frailty Aging 2019, 8, 120–130. [Google Scholar] [CrossRef]
- Morley, J.E. Treatment of sarcopenia: The road to the future. J. Cachexia Sarcopenia Muscle 2018, 9, 1196–1199. [Google Scholar] [CrossRef]
- Hardee, J.P.; Lynch, G.S. Current pharmacotherapies for sarcopenia. Expert Opin. Pharmacother. 2019, 20, 1645–1657. [Google Scholar] [CrossRef] [PubMed]
- Consitt, L.A.; Clark, B.C. The Vicious Cycle of Myostatin Signaling in Sarcopenic Obesity: Myostatin Role in Skeletal Muscle Growth, Insulin Signaling and Implications for Clinical Trials. J. Frailty Aging 2018, 7, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Vatic, M.; von Haehling, S.; Ebner, N. Inflammatory biomarkers of frailty. Exp. Gerontol. 2020, 133, 110858. [Google Scholar] [CrossRef] [PubMed]
- Grosicki, G.J.; Fielding, R.A.; Lustgarten, M.S. Gut Microbiota Contribute to Age-Related Changes in Skeletal Muscle Size, Composition, and Function: Biological Basis for a Gut-Muscle Axis. Calcif. Tissue Int. 2018, 102, 433–442. [Google Scholar] [CrossRef]
- Latham, C.M.; Wagner, A.L.; Urschel, K.L. Effects of dietary amino acid supplementation on measures of whole-body and muscle protein metabolism in aged horses. J. Anim. Physiol. Anim. Nutr. 2019, 103, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Tsintzas, K.; Jones, R.; Pabla, P.; Mallinson, J.; Barrett, D.A.; Kim, D.H.; Cooper, S.; Davies, A.; Taylor, T.; Gaffney, C.; et al. Effect of acute and short-term dietary fat ingestion on postprandial skeletal muscle protein synthesis rates in middle-aged, overweight and obese men. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E417–E429. [Google Scholar] [CrossRef]
- Kieliszek, M.; Piwowarek, K.; Kot, A.M.; Błażejak, S.; Chlebowska-Śmigiel, A.; Wolska, I. Pollen and bee bread as new health-oriented products: A review. Trends Food Sci. Technol. 2018, 71, 170–180. [Google Scholar] [CrossRef]
- Kocot, J.; Kielczykowska, M.; Luchowska-Kocot, D.; Kurzepa, J.; Musik, I. Antioxidant Potential of Propolis, Bee Pollen, and Royal Jelly: Possible Medical Application. Oxid. Med. Cell. Longev. 2018, 2018, 7074209. [Google Scholar] [CrossRef]
- Fratini, F.; Cilia, G.; Mancini, S.; Felicioli, A. Royal Jelly: An ancient remedy with remarkable antibacterial properties. Microbiol. Res. 2016, 192, 130–141. [Google Scholar] [CrossRef]
- Nitecka-Buchta, A.; Buchta, P.; Tabenska-Bosakowska, E.; Walczynska-Dragon, K.; Baron, S. Myorelaxant effect of bee venom topical skin application in patients with RDC/TMD Ia and RDC/TMD Ib: A randomized, double blinded study. BioMed Res. Int. 2014, 2014, 296053. [Google Scholar] [CrossRef]
- Alvarez-Fischer, D.; Noelker, C.; Vulinovic, F.; Grunewald, A.; Chevarin, C.; Klein, C.; Oertel, W.H.; Hirsch, E.C.; Michel, P.P.; Hartmann, A. Bee venom and its component apamin as neuroprotective agents in a Parkinson disease mouse model. PLoS ONE 2013, 8, e61700. [Google Scholar] [CrossRef] [PubMed]
- Cornara, L.; Biagi, M.; Xiao, J.; Burlando, B. Therapeutic Properties of Bioactive Compounds from Different Honeybee Products. Front. Pharmacol. 2017, 8, 412. [Google Scholar] [CrossRef] [PubMed]
- Eşrefoğlu, M.; Gül, M.; Ateş, B.; Erdoğan, A. The effects of caffeic acid phenethyl ester and melatonin on age-related vascular remodeling and cardiac damage. Fundam. Clin. Pharmacol. 2011, 25, 580–590. [Google Scholar] [CrossRef] [PubMed]
- Rzepecka-Stojko, A.; Stojko, J.; Kurek-Górecka, A.; Górecki, M.; Kabała-Dzik, A.; Kubina, R.; Moździerz, A.; Buszman, E. Polyphenols from Bee Pollen: Structure, Absorption, Metabolism and Biological Activity. Molecules 2015, 20, 21732–21749. [Google Scholar] [CrossRef] [PubMed]
- Ebadi, P.; Fazeli, M. Anti-photoaging potential of propolis extract in UVB-irradiated human dermal fibroblasts through increasing the expression of FOXO3A and NGF genes. Biomed. Pharmacother. 2017, 95, 47–54. [Google Scholar] [CrossRef]
- Salles, J.; Cardinault, N.; Patrac, V.; Berry, A.; Giraudet, C.; Collin, M.L.; Chanet, A.; Tagliaferri, C.; Denis, P.; Pouyet, C.; et al. Bee pollen improves muscle protein and energy metabolism in malnourished old rats through interfering with the Mtor signaling pathway and mitochondrial activity. Nutrients 2014, 6, 5500–5516. [Google Scholar] [CrossRef]
- Boisard, S.; Shahali, Y.; Aumond, M.-C.; Derbré, S.; Blanchard, P.; Dadar, M.; Le Ray, A.-M.; Richomme, P. Anti-AGE activity of poplar-type propolis: Mechanism of action of main phenolic compounds. Int. J. Food Sci. 2020, 55, 453–460. [Google Scholar] [CrossRef]
- Egawa, T.; Ohno, Y.; Yokoyama, S.; Yokokawa, T.; Tsuda, S.; Goto, K.; Hayashi, T. The Protective Effect of Brazilian Propolis against Glycation Stress in Mouse Skeletal Muscle. Foods 2019, 8, 439. [Google Scholar] [CrossRef]
- Shaha, A.; Mizuguchi, H.; Kitamura, Y.; Fujino, H.; Yabumoto, M.; Takeda, N.; Fukui, H. Effect of Royal Jelly and Brazilian Green Propolis on the Signaling for Histamine H1 Receptor and Interleukin-9 Gene Expressions Responsible for the Pathogenesis of the Allergic Rhinitis. Biol. Pharm. Bull. 2018, 41, 1440–1447. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Kiesswetter, E.; Drey, M.; Sieber, C.C. Nutrition, frailty, and sarcopenia. Aging Clin. Exp. Res. 2017, 29, 43–48. [Google Scholar] [CrossRef]
- Perna, S.; Alalwan, T.A.; Al-Thawadi, S.; Negro, M.; Parimbelli, M.; Cerullo, G.; Gasparri, C.; Guerriero, F.; Infantino, V.; Diana, M.; et al. Evidence-Based Role of Nutrients and Antioxidants for Chronic Pain Management in Musculoskeletal Frailty and Sarcopenia in Aging. Geriatrics 2020, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Ticinesi, A.; Lauretani, F.; Milani, C.; Nouvenne, A.; Tana, C.; Del Rio, D.; Maggio, M.; Ventura, M.; Meschi, T. Aging Gut Microbiota at the Cross-Road between Nutrition, Physical Frailty, and Sarcopenia: Is There a Gut-Muscle Axis? Nutrients 2017, 9, 1303. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, A.N.K.G.; Nair, A.J.; Sugunan, V.S. A review on Royal Jelly proteins and peptides. J. Funct. Foods 2018, 44, 255–264. [Google Scholar] [CrossRef]
- Virgiliou, C.; Kanelis, D.; Pina, A.; Gika, H.; Tananaki, C.; Zotou, A.; Theodoridis, G. A targeted approach for studying the effect of sugar bee feeding on the metabolic profile of Royal Jelly. J. Chromatogr. A 2019, 460783. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Chen, X.; Tian, Y.; Wu, D.; Du, M.; Wang, S. Protection against oxidative stress and anti-aging effect in Drosophila of royal jelly-collagen peptide. Food Chem. Toxicol. 2020, 135, 110881. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Rababah, T.; Sakandar, H.A.; Imran, M.; Mustafa, N.; Alhamad, M.N.; Mhaidat, N.; Kubow, S.; Tranchant, C.; Al-Tawaha, A.R.; et al. Fermented Food-Derived Bioactive Compounds with Anticarcinogenic Properties: Fermented Royal Jelly as a Novel Source for Compounds with Health Benefits. In Anticancer plants: Properties and Application; Akhtar, M., Swamy, M., Eds.; Springer: Singapore, 2018. [Google Scholar]
- Xin, X.-X.; Chen, Y.; Chen, D.; Xiao, F.; Parnell, L.D.; Zhao, J.; Liu, L.; Ordovas, J.M.; Lai, C.-Q.; Shen, L.-R. Supplementation with Major Royal-Jelly Proteins Increases Lifespan, Feeding, and Fecundity in Drosophila. J. Agric. Food Chem. 2016, 64, 5803–5812. [Google Scholar] [CrossRef]
- Hossen, M.S.; Nahar, T.; Gan, S.H.; Khalil, M.I. Bioinformatics and Therapeutic Insights on Proteins in Royal Jelly. Curr. Proteom. 2019, 16, 84–101. [Google Scholar] [CrossRef]
- Xue, X.; Wu, L.; Wang, K. Chemical Composition of Royal Jelly. In Bee Products—Chemical and Biological Properties; Alvarez-Suarez, J.M., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 181–190. [Google Scholar] [CrossRef]
- Buttstedt, A.; Ihling, C.H.; Pietzsch, M.; Moritz, R.F.A. Royalactin is not a royal making of a queen. Nature 2016, 537, E10. [Google Scholar] [CrossRef]
- Ali, A.M.; Hendawy, A.O. Royal Jelly Acid, 10-Hydroxy-Trans-2-Decenoic Acid, for Psychiatric and Neurological Disorders: How helpful could it be?! Edelweiss J. Food Sci. Technol. 2019, 1, 1–4. [Google Scholar]
- Chen, Y.F.; Wang, K.; Zhang, Y.Z.; Zheng, Y.F.; Hu, F.L. In Vitro Anti-Inflammatory Effects of Three Fatty Acids from Royal Jelly. Mediat. Inflamm. 2016, 2016, 3583684. [Google Scholar] [CrossRef]
- Yang, Y.C.; Chou, W.M.; Widowati, D.A.; Lin, I.P.; Peng, C.C. 10-hydroxy-2-decenoic acid of royal jelly exhibits bactericide and anti-inflammatory activity in human colon cancer cells. BMC Complement. Altern. Med. 2018, 18, 202. [Google Scholar] [CrossRef] [PubMed]
- Weiser, M.J.; Grimshaw, V.; Wynalda, K.M.; Mohajeri, M.H.; Butt, C.M. Long-Term Administration of Queen Bee Acid (QBA) to Rodents Reduces Anxiety-Like Behavior, Promotes Neuronal Health and Improves Body Composition. Nutrients 2017, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Hattori, N.; Nomoto, H.; Fukumitsu, H.; Mishima, S.; Furukawa, S. Royal jelly-induced neurite outgrowth from rat pheochromocytoma PC12 cells requires integrin signal independent of activation of extracellular signalregulated kinases. Biomed. Res. 2007, 28, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.C.; Sun, H.T.; Lin, I.P.; Kuo, P.C.; Li, J.C. The functional property of royal jelly 10-hydroxy-2-decenoic acid as a melanogenesis inhibitor. BMC Complement. Altern. Med. 2017, 17, 392. [Google Scholar] [CrossRef]
- Zheng, J.; Lai, W.; Zhu, G.; Wan, M.; Chen, J.; Tai, Y.; Lu, C. 10-Hydroxy-2-decenoic acid prevents ultraviolet A-induced damage and matrix metalloproteinases expression in human dermal fibroblasts. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 1269–1277. [Google Scholar] [CrossRef]
- Wang, J.G.; Ruan, J.; Li, C.Y.; Wang, J.M.; Li, Y.; Zhai, W.T.; Zhang, W.; Ye, H.; Shen, N.H.; Lei, K.F.; et al. Connective tissue growth factor, a regulator related with 10-hydroxy-2-decenoic acid down-regulate MMPs in rheumatoid arthritis. Rheumatol. Int. 2012, 32, 2791–2799. [Google Scholar] [CrossRef]
- Meng, G.; Wang, H.; Pei, Y.; Li, Y.; Wu, H.; Song, Y.; Guo, Q.; Guo, H.; Fukushima, S.; Tatefuji, T.; et al. Effects of protease-treated royal jelly on muscle strength in elderly nursing home residents: A randomized, double-blind, placebo-controlled, dose-response study. Sci. Rep. 2017, 7, 11416. [Google Scholar] [CrossRef]
- Wessler, I.; Gartner, H.A.; Michel-Schmidt, R.; Brochhausen, C.; Schmitz, L.; Anspach, L.; Grunewald, B.; Kirkpatrick, C.J. Honeybees Produce Millimolar Concentrations of Non-Neuronal Acetylcholine for Breeding: Possible Adverse Effects of Neonicotinoids. PLoS ONE 2016, 11, e0156886. [Google Scholar] [CrossRef]
- Ahmad, S.; Campos, M.G.; Fratini, F.; Altaye, S.Z.; Li, J. New Insights into the Biological and Pharmaceutical Properties of Royal Jelly. Int. J. Mol. Sci. 2020, 21, 382. [Google Scholar] [CrossRef]
- Ramadan, M.F.; Al-Ghamdi, A. Bioactive compounds and health-promoting properties of royal jelly: A review. J. Funct. Foods 2012, 4, 39–52. [Google Scholar] [CrossRef]
- Pan, Y.; Xu, J.; Jin, P.; Yang, Q.; Zhu, K.; You, M.; Chen, M.; Hu, F. Royal Jelly Ameliorates Behavioral Deficits, Cholinergic System Deficiency, and Autonomic Nervous Dysfunction in Ovariectomized Cholesterol-Fed Rabbits. Molecules 2019, 24, 1149. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.M.; Kunugi, H. Apitherapy for Parkinson’s disease: A focus on the effects of propolis and royal jelly. Oxid. Med. Cell. Longev. 2020. accepted. [Google Scholar]
- Ueda, T.; Inden, M.; Shirai, K.; Sekine, S.I.; Masaki, Y.; Kurita, H.; Ichihara, K.; Inuzuka, T.; Hozumi, I. The effects of Brazilian green propolis that contains flavonols against mutant copper-zinc superoxide dismutase-mediated toxicity. Sci. Rep. 2017, 7, 2882. [Google Scholar] [CrossRef] [PubMed]
- Rufatto, L.C.; dos Santos, D.A.; Marinho, F.; Henriques, J.A.P.; Roesch Ely, M.; Moura, S. Red propolis: Chemical composition and pharmacological activity. Asian Pac. J. Trop. Biomed. 2017, 7, 591–598. [Google Scholar] [CrossRef]
- Anjum, S.I.; Ullah, A.; Khan, K.A.; Attaullah, M.; Khan, H.; Ali, H.; Bashir, M.A.; Tahir, M.; Ansari, M.J.; Ghramh, H.A.; et al. Composition and functional properties of propolis (bee glue): A review. Saudi J. Biol. Sci. 2019, 26, 1695–1703. [Google Scholar] [CrossRef]
- Pobiega, K.; Kraśniewska, K.; Gniewosz, M. Application of propolis in antimicrobial and antioxidative protection of food quality—A review. Trends Food Sci. Technol. 2019, 83, 53–62. [Google Scholar] [CrossRef]
- Genovese, S.; Ashida, H.; Yamashita, Y.; Nakgano, T.; Ikeda, M.; Daishi, S.; Epifano, F.; Taddeo, V.A.; Fiorito, S. The interaction of auraptene and other oxyprenylated phenylpropanoids with glucose transporter type 4. Phytomedicine 2017, 32, 74–79. [Google Scholar] [CrossRef]
- Ueda, M.; Hayashibara, K.; Ashida, H. Propolis extract promotes translocation of glucose transporter 4 and glucose uptake through both PI3K- and AMPK-dependent pathways in skeletal muscle. Biofactors 2013, 39, 457–466. [Google Scholar] [CrossRef]
- Sung, S.H.; Choi, G.H.; Lee, N.W.; Shin, B.C. External Use of Propolis for Oral, Skin, and Genital Diseases: A Systematic Review and Meta-Analysis. Evid. Based Complement. Alternat. Med. 2017, 2017, 8025752. [Google Scholar] [CrossRef]
- Goes, A.T.R.; Jesse, C.R.; Antunes, M.S.; Lobo Ladd, F.V.; Lobo Ladd, A.A.B.; Luchese, C.; Paroul, N.; Boeira, S.P. Protective role of chrysin on 6-hydroxydopamine-induced neurodegeneration a mouse model of Parkinson’s disease: Involvement of neuroinflammation and neurotrophins. Chem. Biol. Interact. 2018, 279, 111–120. [Google Scholar] [CrossRef]
- Havermann, S.; Chovolou, Y.; Humpf, H.U.; Watjen, W. Caffeic acid phenethylester increases stress resistance and enhances lifespan in Caenorhabditis elegans by modulation of the insulin-like DAF-16 signalling pathway. PLoS ONE 2014, 9, e100256. [Google Scholar] [CrossRef] [PubMed]
- Zaitone, S.A.; Ahmed, E.; Elsherbiny, N.M.; Mehanna, E.T.; El-Kherbetawy, M.K.; ElSayed, M.H.; Alshareef, D.M.; Moustafa, Y.M. Caffeic acid improves locomotor activity and lessens inflammatory burden in a mouse model of rotenone-induced nigral neurodegeneration: Relevance to Parkinson’s disease therapy. Pharmacol. Rep. 2019, 71, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Noelker, C.; Bacher, M.; Gocke, P.; Wei, X.; Klockgether, T.; Du, Y.; Dodel, R. The flavanoide caffeic acid phenethyl ester blocks 6-hydroxydopamine-induced neurotoxicity. Neurosci. Lett. 2005, 383, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Escriche, I.; Juan-Borrás, M. Standardizing the analysis of phenolic profile in propolis. Food Res. Int. 2018, 106, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Y.; Zhao, L.; Cui, Q.; Wang, Y.; Du, G. Pinocembrin attenuates MPP(+)-induced neurotoxicity by the induction of heme oxygenase-1 through ERK1/2 pathway. Neurosci. Lett. 2016, 612, 104–109. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, J.; Miao, Y.; Cui, Q.; Zhao, W.; Zhang, J.; Wang, H. Pinocembrin protects SH-SY5Y cells against MPP+-induced neurotoxicity through the mitochondrial apoptotic pathway. J. Mol. Neurosci. 2014, 53, 537–545. [Google Scholar] [CrossRef]
- Chi, Y.; Luo, L.; Cui, M.; Hao, Y.; Liu, T.; Huang, X.; Guo, X. Chemical Composition and Antioxidant Activity of Essential Oil of Chinese Propolis. Chem. Biodivers. 2020, 17, e1900489. [Google Scholar] [CrossRef]
- El-Guendouz, S.; Lyoussi, B.; Miguel, M.G. Insight on Propolis from Mediterranean Countries: Chemical Composition, Biological Activities and Application Fields. Chem. Biodivers. 2019, 16, e1900094. [Google Scholar] [CrossRef]
- Hochheim, S.; Pacassa Borges, P.; Boeder, A.M.; Scharf, D.R.; Simionatto, E.L.; Yamanaka, C.N.; Alberton, M.D.; Guedes, A.; de Cordova, C.M.M. A Bioguided Approach for the Screening of Antibacterial Compounds Isolated From the Hydroalcoholic Extract of the Native Brazilian Bee’s Propolis Using Mollicutes as a Model. Front. Microbiol. 2020, 11, 558. [Google Scholar] [CrossRef]
- Rivero-Cruz, J.F.; Granados-Pineda, J.; Pedraza-Chaverri, J.; Pérez-Rojas, J.M.; Kumar-Passari, A.; Diaz-Ruiz, G.; Rivero-Cruz, B.E. Phytochemical Constituents, Antioxidant, Cytotoxic, and Antimicrobial Activities of the Ethanolic Extract of Mexican Brown Propolis. Antioxidants 2020, 9, 70. [Google Scholar] [CrossRef]
- Campos, M.G.R.; Bogdanov, S.; de Almeida-Muradian, L.B.; Szczesna, T.; Mancebo, Y.; Frigerio, C.; Ferreira, F. Pollen composition and standardisation of analytical methods. J. Apic. Res. 2008, 47, 154–161. [Google Scholar] [CrossRef]
- Denisow, B.; Denisow-Pietrzyk, M. Biological and therapeutic properties of bee pollen: A review. J. Sci. Food Agric. 2016, 96, 4303–4309. [Google Scholar] [CrossRef] [PubMed]
- Themelis, T.; Gotti, R.; Orlandini, S.; Gatti, R. Quantitative amino acids profile of monofloral bee pollens by microwave hydrolysis and fluorimetric high performance liquid chromatography. J. Pharm. Biomed. Anal. 2019, 173, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Nanda, V. Composition and functionality of bee pollen: A review. Trends Food Sci. Technol. 2020, 98, 82–106. [Google Scholar] [CrossRef]
- Li, Q.-Q.; Wang, K.; Marcucci, M.C.; Sawaya, A.C.H.F.; Hu, L.; Xue, X.-F.; Wu, L.-M.; Hu, F.-L. Nutrient-rich bee pollen: A treasure trove of active natural metabolites. J. Funct. Foods 2018, 49, 472–484. [Google Scholar] [CrossRef]
- Zhang, D.; Shi, J.; Yang, X. Role of Lipid Metabolism in Plant Pollen Exine Development. Subcell. Biochem. 2016, 86, 315–337. [Google Scholar] [CrossRef] [PubMed]
- Diego-Taboada, A.; Beckett, S.T.; Atkin, S.L.; Mackenzie, G. Hollow pollen shells to enhance drug delivery. Pharmaceutics 2014, 6, 80–96. [Google Scholar] [CrossRef]
- Ketkar, S.; Rathore, A.; Kandhare, A.; Lohidasan, S.; Bodhankar, S.; Paradkar, A.; Mahadik, K. Alleviating exercise-induced muscular stress using neat and processed bee pollen: Oxidative markers, mitochondrial enzymes, and myostatin expression in rats. Integr. Med. Res. 2015, 4, 147–160. [Google Scholar] [CrossRef]
- Shen, X.; Liu, Y.; Luo, X.; Yang, Z. Advances in Biosynthesis, Pharmacology, and Pharmacokinetics of Pinocembrin, a Promising Natural Small-Molecule Drug. Molecules 2019, 24, 2323. [Google Scholar] [CrossRef]
- Guendouz, M.; Haddi, A.; Grar, H.; Kheroua, O.; Saidi, D.; Kaddouri, H. Preventive effects of royal jelly against anaphylactic response in a murine model of cow’s milk allergy. Pharm. Biol. 2017, 55, 2145–2152. [Google Scholar] [CrossRef]
- Cifuentes, L. Allergy to honeybee … not only stings. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, S.G.; Kim, I.S.; Lee, H.D. Standardization of the Manufacturing Process of Bee Venom Pharmacopuncture Containing Melittin as the Active Ingredient. Evid. Based Complement. Alternat. Med. 2018, 2018, 2353280. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.Z.; Zhang, C.P.; Wei, W.T.; Hu, F.L. The in vivo antiaging effect of enzymatic hydrolysate from royal jelly in d-galactose induced aging mouse. J. Chin. Inst. Food Sci. Technol. 2016, 16, 18–25. [Google Scholar]
- Niu, K.; Guo, H.; Guo, Y.; Ebihara, S.; Asada, M.; Ohrui, T.; Furukawa, K.; Ichinose, M.; Yanai, K.; Kudo, Y.; et al. Royal jelly prevents the progression of sarcopenia in aged mice in vivo and in vitro. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1482–1492. [Google Scholar] [CrossRef] [PubMed]
- Okumura, N.; Toda, T.; Ozawa, Y.; Watanabe, K.; Ikuta, T.; Tatefuji, T.; Hashimoto, K.; Shimizu, T. Royal Jelly Delays Motor Functional Impairment During Aging in Genetically Heterogeneous Male Mice. Nutrients 2018, 10, 1191. [Google Scholar] [CrossRef]
- Pyrzanowska, J.; Piechal, A.; Blecharz-Klin, K.; Graikou, K.; Widy-Tyszkiewicz, E.; Chinou, I. Chemical analysis of Greek royal jelly—Its influence of the long-term administration on spatial memory in aged rats. Planta Med. 2012, 78. [Google Scholar] [CrossRef]
- Pyrzanowska, J.; Piechal, A.; Blecharz-Klin, K.; Joniec-Maciejak, I.; Graikou, K.; Chinou, I.; Widy-Tyszkiewicz, E. Long-term administration of Greek Royal Jelly improves spatial memory and influences the concentration of brain neurotransmitters in naturally aged Wistar male rats. J. Ethnopharmacol. 2014, 155, 343–351. [Google Scholar] [CrossRef]
- Metwally Ibrahim, S.E.L.; Kosba, A.A. Royal jelly supplementation reduces skeletal muscle lipotoxicity and insulin resistance in aged obese rats. Pathophysiology 2018, 25, 307–315. [Google Scholar] [CrossRef]
- Shen, Y.C.; Yen, J.C.; Liou, K.T. Ameliorative effects of caffeic acid phenethyl ester on an eccentric exercise-induced skeletal muscle injury by down-regulating NF-kappab mediated inflammation. Pharmacology 2013, 91, 219–228. [Google Scholar] [CrossRef]
- Ozyurt, B.; Iraz, M.; Koca, K.; Ozyurt, H.; Sahin, S. Protective effects of caffeic acid phenethyl ester on skeletal muscle ischemia-reperfusion injury in rats. Mol. Cell. Biochem. 2006, 292, 197–203. [Google Scholar] [CrossRef]
- Ozyurt, H.; Ozyurt, B.; Koca, K.; Ozgocmen, S. Caffeic acid phenethyl ester (CAPE) protects rat skeletal muscle against ischemia-reperfusion-induced oxidative stress. Vascul. Pharmacol. 2007, 47, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.D.D.; Zadinelo, I.V.; Silva, L.; Zilli, R.L.; Barreiros, M.A.B.; Mauerwerk, M.T.; Meurer, F. Alcoholic extract of propolis in Nile tilapia post-larvae and fingerlings’ diets: Effects on production performance, body composition and intestinal histology. An. Acad. Bras. Cienc. 2019, 91, e20180297. [Google Scholar] [CrossRef] [PubMed]
- Santos, N.W.; Yoshimura, E.H.; Mareze-Costa, C.E.; Machado, E.; Agustinho, B.C.; Pereira, L.M.; Brito, M.N.; Brito, N.A.; Zeoula, L.M. Supplementation of cow milk naturally enriched in polyunsaturated fatty acids and polyphenols to growing rats. PLoS ONE 2017, 12, e0172909. [Google Scholar] [CrossRef] [PubMed]
- Takikawa, M.; Kumagai, A.; Hirata, H.; Soga, M.; Yamashita, Y.; Ueda, M.; Ashida, H.; Tsuda, T. 10-Hydroxy-2-decenoic acid, a unique medium-chain fatty acid, activates 5′-AMP-activated protein kinase in L6 myotubes and mice. Mol. Nutr. Food Res. 2013, 57, 1794–1802. [Google Scholar] [CrossRef]
- Edilova, M.I.; Abdul-Sater, A.A.; Watts, T.H. TRAF1 Signaling in Human Health and Disease. Front. Immunol. 2018, 9, 2969. [Google Scholar] [CrossRef]
- Washio, K.; Kobayashi, M.; Saito, N.; Amagasa, M.; Kitamura, H. Propolis Ethanol Extract Stimulates Cytokine and Chemokine Production through NF-kappaB Activation in C2C12 Myoblasts. Evid. Based Complement. Alternat. Med. 2015, 2015, 349751. [Google Scholar] [CrossRef]
- Kwon, T.D.; Lee, M.W.; Kim, K.H. The effect of exercise training and water extract from propolis intake on the antioxidant enzymes activity of skeletal muscle and liver in rat. J. Exerc. Nutr. Biochem. 2014, 18, 9–17. [Google Scholar] [CrossRef]
- Tanaka, M.; Kanazashi, M.; Maeshige, N.; Kondo, H.; Ishihara, A.; Fujino, H. Protective effects of Brazilian propolis supplementation on capillary regression in the soleus muscle of hindlimb-unloaded rats. J. Physiol. Sci. 2019, 69, 223–233. [Google Scholar] [CrossRef]
- Lee, E.S.; Uhm, K.O.; Lee, Y.M.; Han, M.; Lee, M.; Park, J.M.; Suh, P.G.; Park, S.H.; Kim, H.S. CAPE (caffeic acid phenethyl ester) stimulates glucose uptake through AMPK (AMP-activated protein kinase) activation in skeletal muscle cells. Biochem. Biophys. Res. Commun. 2007, 361, 854–858. [Google Scholar] [CrossRef]
- Takahashi, Y.; Hijikata, K.; Seike, K.; Nakano, S.; Banjo, M.; Sato, Y.; Takahashi, K.; Hatta, H. Effects of Royal Jelly Administration on Endurance Training-Induced Mitochondrial Adaptations in Skeletal Muscle. Nutrients 2018, 10, 1735. [Google Scholar] [CrossRef]
- Yuce, S.; Cemal Gokce, E.; Iskdemir, A.; Koc, E.R.; Cemil, D.B.; Gokce, A.; Sargon, M.F. An experimental comparison of the effects of propolis, curcumin, and methylprednisolone on crush injuries of the sciatic nerve. Ann. Plast. Surg. 2015, 74, 684–692. [Google Scholar] [CrossRef]
- Miko, A.; Poto, L.; Matrai, P.; Hegyi, P.; Furedi, N.; Garami, A.; Illes, A.; Solymar, M.; Vincze, A.; Balasko, M.; et al. Gender difference in the effects of interleukin-6 on grip strength—A systematic review and meta-analysis. BMC Geriatr. 2018, 18, 107. [Google Scholar] [CrossRef] [PubMed]
- Lima Cavendish, R.; de Souza Santos, J.; Belo Neto, R.; Oliveira Paixao, A.; Valeria Oliveira, J.; Divino de Araujo, E.; Berretta, E.S.A.A.; Maria Thomazzi, S.; Cordeiro Cardoso, J.; Zanardo Gomes, M. Antinociceptive and anti-inflammatory effects of Brazilian red propolis extract and formononetin in rodents. J. Ethnopharmacol. 2015, 173, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Roquetto, A.R.; Monteiro, N.E.S.; Moura, C.S.; Toreti, V.C.; de Pace, F.; Santos, A.D.; Park, Y.K.; Amaya-Farfan, J. Green propolis modulates gut microbiota, reduces endotoxemia and expression of TLR4 pathway in mice fed a high-fat diet. Food Res. Int. 2015, 76, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Buck, E.L.; Mizubuti, I.Y.; Alfieri, A.A.; Otonel, R.A.; Buck, L.Y.; Souza, F.P.; Prado-Calixto, O.P.; Poveda-Parra, A.R.; Alexandre Filho, L.; Lopera-Barrero, N.M. Effect of propolis ethanol extract on myostatin gene expression and muscle morphometry of Nile tilapia in net cages. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef]
- Sriram, S.; Subramanian, S.; Sathiakumar, D.; Venkatesh, R.; Salerno, M.S.; McFarlane, C.D.; Kambadur, R.; Sharma, M. Modulation of reactive oxygen species in skeletal muscle by myostatin is mediated through NF-kappaB. Aging Cell 2011, 10, 931–948. [Google Scholar] [CrossRef]
- Watadani, R.; Kotoh, J.; Sasaki, D.; Someya, A.; Matsumoto, K.; Maeda, A. 10-Hydroxy-2-decenoic acid, a natural product, improves hyperglycemia and insulin resistance in obese/diabetic KK-Ay mice, but does not prevent obesity. J. Vet. Med. Sci. 2017, 79, 1596–1602. [Google Scholar] [CrossRef]
- D’Antona, G.; Ragni, M.; Cardile, A.; Tedesco, L.; Dossena, M.; Bruttini, F.; Caliaro, F.; Corsetti, G.; Bottinelli, R.; Carruba, M.O.; et al. Branched-chain amino acid supplementation promotes survival and supports cardiac and skeletal muscle mitochondrial biogenesis in middle-aged mice. Cell Metab. 2010, 12, 362–372. [Google Scholar] [CrossRef]
- Puigserver, P.; Spiegelman, B.M. Peroxisome proliferator-activated receptor-gamma coactivator 1 alpha (PGC-1 alpha): Transcriptional coactivator and metabolic regulator. Endocr. Rev. 2003, 24, 78–90. [Google Scholar] [CrossRef]
- Inoue, Y.; Hara, H.; Mitsugi, Y.; Yamaguchi, E.; Kamiya, T.; Itoh, A.; Adachi, T. 4-Hydroperoxy-2-decenoic acid ethyl ester protects against 6-hydroxydopamine-induced cell death via activation of Nrf2-ARE and eIF2α-ATF4 pathways. Neurochem. Int. 2018, 112, 288–296. [Google Scholar] [CrossRef]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat. Commun. 2016, 7, 11624. [Google Scholar] [CrossRef]
- Abdulla, H.; Smith, K.; Atherton, P.J.; Idris, I. Role of insulin in the regulation of human skeletal muscle protein synthesis and breakdown: A systematic review and meta-analysis. Diabetologia 2016, 59, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Guillet, C.; Boirie, Y. Insulin resistance: A contributing factor to age-related muscle mass loss? Diabetes Metab. 2005, 31, 5S20–5S26. [Google Scholar] [CrossRef]
- Tubbs, E.; Chanon, S.; Robert, M.; Bendridi, N.; Bidaux, G.; Chauvin, M.-A.; Ji-Cao, J.; Durand, C.; Gauvrit-Ramette, D.; Vidal, H.; et al. Disruption of Mitochondria-Associated Endoplasmic Reticulum Membrane (MAM) Integrity Contributes to Muscle Insulin Resistance in Mice and Humans. Diabetes 2018, 67, 636–650. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K. AMP-activated protein kinase (AMPK) controls the aging process via an integrated signaling network. Ageing Res. Rev. 2012, 11, 230–241. [Google Scholar] [CrossRef]
- Salminen, A.; Kauppinen, A.; Kaarniranta, K. AMPK activation inhibits the functions of myeloid-derived suppressor cells (MDSC): Impact on cancer and aging. J. Mol. Med. 2019, 97, 1049–1064. [Google Scholar] [CrossRef]
- You, M.; Miao, Z.; Tian, J.; Hu, F. Trans-10-hydroxy-2-decenoic acid protects against LPS-induced neuroinflammation through FOXO1-mediated activation of autophagy. Eur. J. Nutr. 2019, 59, 2875–2892. [Google Scholar] [CrossRef]
- Argiles, J.M.; Campos, N.; Lopez-Pedrosa, J.M.; Rueda, R.; Rodriguez-Manas, L. Skeletal Muscle Regulates Metabolism via Interorgan Crosstalk: Roles in Health and Disease. J. Am. Med. Direct. Assoc. 2016, 17, 789–796. [Google Scholar] [CrossRef]
- Landi, F.; Calvani, R.; Tosato, M.; Martone, A.M.; Ortolani, E.; Savera, G.; D’Angelo, E.; Sisto, A.; Marzetti, E. Protein Intake and Muscle Health in Old Age: From Biological Plausibility to Clinical Evidence. Nutrients 2016, 8, 295. [Google Scholar] [CrossRef]
- Chen, L.K.; Liu, L.K.; Woo, J.; Assantachai, P.; Auyeung, T.W.; Bahyah, K.S.; Chou, M.Y.; Chen, L.Y.; Hsu, P.S.; Krairit, O.; et al. Sarcopenia in Asia: Consensus report of the Asian Working Group for Sarcopenia. J. Am. Med. Direct. Assoc. 2014, 15, 95–101. [Google Scholar] [CrossRef]
- Walrand, S.; Gryson, C.; Salles, J.; Giraudet, C.; Migne, C.; Bonhomme, C.; Le Ruyet, P.; Boirie, Y. Fast-digestive protein supplement for ten days overcomes muscle anabolic resistance in healthy elderly men. Clin. Nutr. 2016, 35, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Boirie, Y.; Guillet, C. Fast digestive proteins and sarcopenia of aging. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Graber, T.G.; Borack, M.S.; Reidy, P.T.; Volpi, E.; Rasmussen, B.B. Essential amino acid ingestion alters expression of genes associated with amino acid sensing, transport, and mTORC1 regulation in human skeletal muscle. Nutr. Metab. 2017, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Ham, D.J.; Lynch, G.S.; Koopman, R. Amino acid sensing and activation of mechanistic target of rapamycin complex 1: Implications for skeletal muscle. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 67–73. [Google Scholar] [CrossRef]
- Moro, T.; Brightwell, C.R.; Deer, R.R.; Graber, T.G.; Galvan, E.; Fry, C.S.; Volpi, E.; Rasmussen, B.B. Muscle Protein Anabolic Resistance to Essential Amino Acids Does Not Occur in Healthy Older Adults Before or After Resistance Exercise Training. J. Nutr. 2018, 148, 900–909. [Google Scholar] [CrossRef]
- Osowska, S.; Duchemann, T.; Walrand, S.; Paillard, A.; Boirie, Y.; Cynober, L.; Moinard, C. Citrulline modulates muscle protein metabolism in old malnourished rats. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E582–E586. [Google Scholar] [CrossRef]
- Moro, T.; Ebert, S.M.; Adams, C.M.; Rasmussen, B.B. Amino Acid Sensing in Skeletal Muscle. Trends Endocrinol. Metab. 2016, 27, 796–806. [Google Scholar] [CrossRef]
- Zhu, Z.; Yang, C.; Iyaswamy, A.; Krishnamoorthi, S.; Sreenivasmurthy, S.G.; Liu, J.; Wang, Z.; Tong, B.C.; Song, J.; Lu, J.; et al. Balancing mTOR Signaling and Autophagy in the Treatment of Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 728. [Google Scholar] [CrossRef]
- Wang, X.; Cook, L.F.; Grasso, L.M.; Cao, M.; Dong, Y. Royal Jelly-Mediated Prolongevity and Stress Resistance in Caenorhabditis elegans Is Possibly Modulated by the Interplays of DAF-16, SIR-2.1, HCF-1, and 14-3-3 Proteins. J. Gerontol. A Biol. 2015, 70, 827–838. [Google Scholar] [CrossRef]
- Milan, G.; Romanello, V.; Pescatore, F.; Armani, A.; Paik, J.-H.; Frasson, L.; Seydel, A.; Zhao, J.; Abraham, R.; Goldberg, A.L.; et al. Regulation of autophagy and the ubiquitin–proteasome system by the FoxO transcriptional network during muscle atrophy. Nat. Commun. 2015, 6, 6670. [Google Scholar] [CrossRef]
- Chiche, A.; Le Roux, I.; von Joest, M.; Sakai, H.; Aguin, S.B.; Cazin, C.; Salam, R.; Fiette, L.; Alegria, O.; Flamant, P.; et al. Injury-Induced Senescence Enables In Vivo Reprogramming in Skeletal Muscle. Cell Stem Cell 2017, 20, 407–414.e404. [Google Scholar] [CrossRef]
- Sharman, M.J.; Verdile, G.; Kirubakaran, S.; Münch, G. Inflammation in Alzheimer’s Disease, and Prevention with Antioxidants and Phenolic Compounds—What Are the Most Promising Candidates? In Neurodegeneration and Alzheimer’s Disease: The Role of Diabetes, Genetics, Hormones, and Lifestyle; Martins, R.N., Brennan, C.S., Fernando, B., Brennan, M.A., Fuller, S.J., Eds.; JohnWiley & Sons Ltd.: Hoboken, NJ, USA, 2019; pp. 233–266. [Google Scholar] [CrossRef]
- MacLean, M.; Derk, J.; Ruiz, H.H.; Juranek, J.K.; Ramasamy, R.; Schmidt, A.M. The Receptor for Advanced Glycation End Products (RAGE) and DIAPH1: Implications for vascular and neuroinflammatory dysfunction in disorders of the central nervous system. Neurochem. Int. 2019, 126, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Chenet, A.L.; Duarte, A.R.; de Almeida, F.J.S.; Andrade, C.M.B.; de Oliveira, M.R. Carvacrol Depends on Heme Oxygenase-1 (HO-1) to Exert Antioxidant, Anti-inflammatory, and Mitochondria-Related Protection in the Human Neuroblastoma SH-SY5Y Cells Line Exposed to Hydrogen Peroxide. Neurochem. Res. 2019, 44, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Xu, J.; Chen, C.; Chen, F.; Jin, P.; Zhu, K.; Hu, C.W.; You, M.; Chen, M.; Hu, F. Royal Jelly Reduces Cholesterol Levels, Ameliorates Aβ Pathology and Enhances Neuronal Metabolic Activities in a Rabbit Model of Alzheimer’s Disease. Front. Aging Neurosci. 2018, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Bottini, N.; Gloria-Bottini, F.; Borgiani, P.; Antonacci, E.; Lucarelli, P.; Bottini, E. Type 2 diabetes and the genetics of signal transduction: A study of interaction between adenosine deaminase and acid phosphatase locus 1 polymorphisms. Metabolism 2004, 53, 995–1001. [Google Scholar] [CrossRef]
- Anagnostou, M.E.; Hepple, R.T. Mitochondrial Mechanisms of Neuromuscular Junction Degeneration with Aging. Cells 2020, 9, 197. [Google Scholar] [CrossRef]
- Sharma, B.; Dutt, V.; Kaur, N.; Mittal, A.; Dabur, R. Tinospora cordifolia protects from skeletal muscle atrophy by alleviating oxidative stress and inflammation induced by sciatic denervation. J. Ethnopharmacol. 2020, 254, 112720. [Google Scholar] [CrossRef]
- Siddharth, J.; Chakrabarti, A.; Pannerec, A.; Karaz, S.; Morin-Rivron, D.; Masoodi, M.; Feige, J.N.; Parkinson, S.J. Aging and sarcopenia associate with specific interactions between gut microbes, serum biomarkers and host physiology in rats. Aging 2017, 9, 1698–1720. [Google Scholar] [CrossRef]
- Bindels, L.B.; Delzenne, N.M. Muscle wasting: The gut microbiota as a new therapeutic target? Int. J. Biochem. Cell Biol. 2013, 45, 2186–2190. [Google Scholar] [CrossRef]
- Genin, E.C.; Madji Hounoum, B.; Bannwarth, S.; Fragaki, K.; Lacas-Gervais, S.; Mauri-Crouzet, A.; Lespinasse, F.; Neveu, J.; Ropert, B.; Auge, G.; et al. Mitochondrial defect in muscle precedes neuromuscular junction degeneration and motor neuron death in CHCHD10(S59L/+) mouse. Acta Neuropathol. 2019, 138, 123–145. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, J.; Shu, X.; Bai, L.; Xu, W.; Wang, A.; Chen, A.; Tu, W.Y.; Wang, J.; Zhang, K.; et al. Loss of mitochondrial protein CHCHD10 in skeletal muscle causes neuromuscular junction impairment. Hum. Mol. Genet. 2019, 29, 1784–1796. [Google Scholar] [CrossRef] [PubMed]
- Roos, T.U.; Heiss, E.H.; Schwaiberger, A.V.; Schachner, D.; Sroka, I.M.; Oberan, T.; Vollmar, A.M.; Dirsch, V.M. Caffeic acid phenethyl ester inhibits PDGF-induced proliferation of vascular smooth muscle cells via activation of p38 MAPK, HIF-1alpha, and heme oxygenase-1. J. Nat. Prod. 2011, 74, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Ticinesi, A.; Meschi, T.; Narici, M.V.; Lauretani, F.; Maggio, M. Muscle Ultrasound and Sarcopenia in Older Individuals: A Clinical Perspective. J. Am. Med. Direct. Assoc. 2017, 18, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, W.K.; Phillips, B.E.; Williams, J.P.; Rankin, D.; Smith, K.; Lund, J.N.; Atherton, P.J. Development of a new Sonovue contrast-enhanced ultrasound approach reveals temporal and age-related features of muscle microvascular responses to feeding. Physiol. Rep. 2013, 1, e00119. [Google Scholar] [CrossRef]
- Menzies, K.; Zaldivar-Jolissaint, J.F.; Auwerx, J. Sirtuins as Metabolic Modulators of Muscle Plasticity. In Sirtuins, Proteins and Cell Regulation; Houtkooper, R., Ed.; Springer: Dordrecht, The Netherlands, 2016; Volume 10. [Google Scholar]
- Chen, Y.M.; Wei, L.; Chiu, Y.S.; Hsu, Y.J.; Tsai, T.Y.; Wang, M.F.; Huang, C.C. Lactobacillus plantarum TWK10 Supplementation Improves Exercise Performance and Increases Muscle Mass in Mice. Nutrients 2016, 8, 205. [Google Scholar] [CrossRef]
- Bindels, L.B.; Beck, R.; Schakman, O.; Martin, J.C.; De Backer, F.; Sohet, F.M.; Dewulf, E.M.; Pachikian, B.D.; Neyrinck, A.M.; Thissen, J.P.; et al. Restoring specific lactobacilli levels decreases inflammation and muscle atrophy markers in an acute leukemia mouse model. PLoS ONE 2012, 7, e37971. [Google Scholar] [CrossRef]
- Ali, A.M.; Kunugi, H. Bee honey protects astrocytes against oxidative stress: A preliminary in vitro investigation. Neuropsychopharmacol. Rep. 2019, 39, 312–314. [Google Scholar] [CrossRef]
- Buigues, C.; Fernandez-Garrido, J.; Pruimboom, L.; Hoogland, A.J.; Navarro-Martinez, R.; Martinez-Martinez, M.; Verdejo, Y.; Mascaros, M.C.; Peris, C.; Cauli, O. Effect of a Prebiotic Formulation on Frailty Syndrome: A Randomized, Double-Blind Clinical Trial. Int. J. Mol. Sci. 2016, 17, 932. [Google Scholar] [CrossRef]
- Wang, D.; Ho, L.; Faith, J.; Ono, K.; Janle, E.M.; Lachcik, P.J.; Cooper, B.R.; Jannasch, A.H.; D’Arcy, B.R.; Williams, B.A.; et al. Role of intestinal microbiota in the generation of polyphenol-derived phenolic acid mediated attenuation of Alzheimer’s disease beta-amyloid oligomerization. Mol. Nutr. Food Res. 2015, 59, 1025–1040. [Google Scholar] [CrossRef]
- Picca, A.; Ponziani, F.R.; Calvani, R.; Marini, F.; Biancolillo, A.; Coelho-Junior, H.J.; Gervasoni, J.; Primiano, A.; Putignani, L.; Del Chierico, F.; et al. Gut Microbial, Inflammatory and Metabolic Signatures in Older People with Physical Frailty and Sarcopenia: Results from the BIOSPHERE Study. Nutrients 2019, 12, 65. [Google Scholar] [CrossRef]
- Hindupur, S.K.; González, A.; Hall, M.N. The opposing actions of target of rapamycin and AMP-activated protein kinase in cell growth control. Cold Spring Harb. Perspect. Biol. 2015, 7, a019141. [Google Scholar] [CrossRef] [PubMed]
- Jeejeebhoy, K.N. Malnutrition, fatigue, frailty, vulnerability, sarcopenia and cachexia: Overlap of clinical features. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Joksimović, A.; Stanković, D.; Joksimović, I.; Molnar, S.; Joksimović, S. Royal jelly as a supplement for young football players. Sport Sci. 2009, 2, 62–67. [Google Scholar]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef]
- Jeffery, I.B.; Lynch, D.B.; O’Toole, P.W. Composition and temporal stability of the gut microbiota in older persons. ISME J. 2016, 10, 170–182. [Google Scholar] [CrossRef]
- Fragala, M.S.; Alley, D.E.; Shardell, M.D.; Harris, T.B.; McLean, R.R.; Kiel, D.P.; Cawthon, P.M.; Dam, T.T.; Ferrucci, L.; Guralnik, J.M.; et al. Comparison of Handgrip and Leg Extension Strength in Predicting Slow Gait Speed in Older Adults. J. Am. Geriatr. Soc. 2016, 64, 144–150. [Google Scholar] [CrossRef]
- Solerte, S.B.; Gazzaruso, C.; Bonacasa, R.; Rondanelli, M.; Zamboni, M.; Basso, C.; Locatelli, E.; Schifino, N.; Giustina, A.; Fioravanti, M. Nutritional supplements with oral amino acid mixtures increases whole-body lean mass and insulin sensitivity in elderly subjects with sarcopenia. Am. J. Cardiol. 2008, 101, 69e–77e. [Google Scholar] [CrossRef]
- Ali, A.M.; Ahmed, A.H.; Smail, L. Psychological Climacteric Symptoms and Attitudes toward Menopause among Emirati Women. Int. J. Environ. Res. Public Health 2020, 17, 5028. [Google Scholar] [CrossRef]
- Landi, F.; Camprubi-Robles, M.; Bear, D.E.; Cederholm, T.; Malafarina, V.; Welch, A.A.; Cruz-Jentoft, A.J. Muscle loss: The new malnutrition challenge in clinical practice. Clin. Nutr. 2019, 38, 2113–2120. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Bee Products/Their Constituents | Animal Model/Cell Line | Experimental Design | Outcome Measures | Results and Possible Related Mechanisms | References |
|---|---|---|---|---|---|
| In vitro Royal jelly and pRJ (500 μg/mL: 1, 2, 3, and 5 d) In vivo Dietary royal jelly and pRJ (1% and 5% for 3 months) | In vitro SC isolated from aged mice In vivo 21-months old male C57BL/6 mice as a mouse model of sarcopenia | In vitro EG1: royal jelly treated SC EG2: pRJ treated SC CG: untreated SC In vivo EG1 and EG2: aged mice on 1% and 5% royal jelly, respectively EG3 and EG4: aged mice on 1% and 5% pRJ, respectively CG: untreated aged mice | In vitro SC proliferation and differentiation into myotubes and AKT signaling. In vivo No of SC, skeletal muscle weight, grip strength, regenerative capacity of injured muscle, and serum IGF-1. | In vitro pRJ enhanced SC proliferation rate and differentiation into myotubes through activation of AKT signaling. In vivo Royal jelly and pRJ significantly increased the number of SC, weight of skeletal muscle, grip strength, regenerative capacity of injured skeletal muscle, and IGF-1 serum level. | [96] |
| Intragastric 10-HDA (1.6 mM/kg body weight) | In vitro L6 myotubes obtained from Osaka biobank In vivo 7-weeks old male C57BL/6J mice | In vitro EG: 10-HDA CG1: AICAR (1 mM) CG2: DMSO (0.1%) In vivo EG: 10-HDA (1.6 mM) CG1: gum Arabic (5%) | Glucose uptake, AMPK signaling, and Glut4 translocation. | 10-HDA increased glucose uptake into L6 myotubes following AMPK activation and Glut4 translocation to the plasma membrane. AMPK activation was induced by the upstream kinase Ca²/calmodulin-dependent kinase β, independent of changes in AMP:ATP ratio and the liver kinase B1 pathway. | [106] |
| Intragastric royal jelly and pRJ (0.7 and 1.4 mg/kg body weight/d/90 d) | D-galactose induced mouse model of aging | EG1 and EG2: mice on 0.7 and 1.4 mg RJ, respectively EG3 and EG4: mice on 0.7 and 1.4 mg RJ hydrolysate, respectively CG: untreated mice | Antioxidant enzymes, body weight, muscular performance, memory, and learning. | Both doses of pRJ prevented age-related locomotor decline, preserved body weight, enhanced memory and learning, increased antioxidant enzyme activity, and inhibited the production of lipid peroxides. | [95] |
| Oral 10-HDA (12-60 mg/kg body weight/d/4 months) | Obese old rats and stressed mice as models of sarcopenia and depression (male and female Sprague-Dawley and 2-months old male and female BALB/c) | EG1 and EG2: aged obese rats on 10-HDA (12 or 24 mg/kg/d for 3.5 months) EG3 and EG4: stressed BALB/c mice on 10-HDA (30 or 60 mg/kg/d for 4 months) CG1 and CG2: untreated aged obese rats and untreated stressed BALB/c mice | Body weight, weight of abdominal adipose tissue, and muscle mass. | 10-HDA significantly increased weight gain and weight maintenance in aged rats/mice undergoing behavioral stress without any change in diet consumption. It also significantly decreased adipose tissue in female animals and increased muscle mass in male rodents compared with untreated controls. | [54] |
| Dietary royal jelly and pRJ + MF powder diet (0.05% or 0.5%) | 6-months old male HET mouse model of severe sarcopenia (background strains: BALB/c, C57BL/6, C3H, and DBA/2) | EG1 and EG2: HET mice on 0.05% and 0.5% royal jelly, respectively EG3 and EG4: HET mice on 0.05% and 0.5% pRJ, respectively CG1: untreated HET mice CG2: untreated young mice | No of blood cells and Pax7 SC, albumin, AST, ALT, T-CHO, TG, expression of muscle genes (MyoD, myogenein, myostatin) and catabolic genes (E3 ubiquitin ligases MuRF1, and atrogin-1). Behavioral tests: grip strength, wire hang, rotarod, and horizontal bar tests. | RJ and pRJ significantly delayed age-related impairment of motor functions, positively improved physical performance of treated mice in 4 types of tests (grip strength, wire hang, horizontal bar, and rotarod), lowered age-related muscular atrophy, increased No of Pax7 SC markers, and suppressed catabolic genes. | [97] |
| Intragastric royal jelly (100 mg/kg body weight/d/8 weeks) | 10-months old male Sprague-Dawley on HFD as a rat model of sarcopenic obesity | EG1: aged rats on royal jelly and HFD EG2: aged rats on royal jelly CG1: untreated young rats CG2: untreated aged rats CG3: untreated old rats on HFD | Serum levels of T-CHO, TG, HDL-c, LDL-c, insulin, HOMA-IR. Skeletal muscle TG levels. Serum and adipose tissue levels of TNFR1. Percentage of weight gain of the body, abdominal fat, and tibialis anterior and hind limb muscles. | Royal jelly significantly decreased insulin levels, HOMA-IR, TNFR1 in serum and adipose tissue, serum lipids, muscle TG levels, body weight gain and abdominal fat weight and significantly increased the weight of hind limb muscle in aged rats on HFD compared with aged mice on HFD only. | [100] |
| Oral royal jelly (1.0 mg/g body weight/d/3 weeks) | 6–7 weeks old ICR mice | EG: endurance exercise + royal jelly CG1: sedentary rats on royal jelly CG2: endurance exercise+ distilled water CG3: sedentary rats on distilled water | CS, β-HAD, and activities of AMPK and acetyl-CoA carboxylase in the soleus, plantaris, and tibialis anterior muscles. | Royal jelly increased CS and β-HAD maximal activities in the soleus muscle compared with all CGs but failed to affect these enzymes in the plantaris and tibialis anterior muscles of sedentary mice compared with CG2. Royal jelly effects in the soleus muscle were mediated by AMPK and acetyl-CoA carboxylase activity. | [106] |
| Gavage/oral lyophilized royal jelly (50 and 100 mg/kg body weight/d/8 weeks) | 18-months old (naturally aging) Wistar male rats as a model of aging | EG1 and EG2: aged rats on royal jelly 50 and 100 mg, respectively CG: aged rats on gavage solution of 0.9% NaCl. | Learning, spatial memory, and motor performance on Morris water maze. | Royal jelly improved learning, spatial memory, and motor performance e.g., increased the number of crossings, swimming speed, and swimming distance. | [98,99] |
| Oral pRJ (1.2 or 4.8 g/d over 1 year) | Institutionalized older adults (mean age: 78.5 ± 7.5 years, N = 199, N males = 99, N females = 95) | EG1 and EG2: pRJ (1.2 and 4.8 g/d), respectively CG: placebo | Handgrip strength, six-minute walk test, timed up and go test, and standing on one leg with eyes closed. | pRJ had no significant effect on handgrip strength, six-minute walk test, timed up and go test, and standing on one leg with eyes closed. | [59] |
| Brazilian propolis extract (100 μg/mL/4-12 h) | In vitro Differentiated myoblast C2C12 cells and RAW264 macrophages isolated from mice | EGs: propolis (100 μg/mL) CG1: ethanol (0.008%) CG2: DMSO (0.08%). CG3: IKK inhibitor (BMS-345541) | IL-6, LIF, CCL-2, CCL-5, CXCL-10, VEGF-A, COX2, MMP-12, migration of RAW264 macrophages, and activities of MAIL/IκBζ and NF-κB. | Propolis (at 8h) induced RAW264 macrophage migration, activated MAIL/IκBζ and NF-κB proteins p50 and p65, and increased levels of VEGF-A, COX-2, MMP-12, CCL-2, CCL-5, CCL-10, LIF, and IL-6. Propolis inhibited the production of IL-1β and TNF-α. | [108] |
| CAPE (1 and 10 μM/3 min-12 h) | In vitro Differentiated L6 myoblast cells isolated from rats | EGs: CAPE (1, 10 μM) CG1: insulin (100 nM) CG2: AICAR | 2-Deoxyglucose uptake, AMPK and AKT signaling. | CAPE (10 μM/1h) increased 2-Deoxyglucose uptake (same as insulin) and activated AMPK (same as AICAR, an AMPK activator). CAPE (10 μM/3 min) activated AKT in a PI3K-dependent manner. | [111] |
| Boropinic acid, 4-geranyloxyferulic acid, 7-isopentenyloxycoumarin, auraptene (0.1, 1, 10 μM), and raw Italian propolis (0.001–1 mg/mL) | In vitro Differentiated L6 myoblast cells isolated from rats | EGs: boropinic acid, 4-geranyloxyferulic acid, 7-isopentenyloxycoumarin, auraptene (0.1, 1, 10 μM), and propolis (0.001–1 mg/mL) CG1: insulin (0.1 μM) CG2: DMSO | GLUT4-mediated glucose uptake and GLUT4 translocation. | Propolis (1.0 and 1 mg/mL), 4-geranyloxyferulic acid, 7-isopentenyloxycoumarin, and auraptene significantly increased glucose uptake and GLUT4 translocation. | [69] |
| In vitro A single oral dose of Brazilian propolis extract (250 mg/kg body weight) In vivo Artepillin C, coumaric acid, and kaempferide (1–104 ng/mL for 15 min) | In vitro Differentiated L6 myoblast cells isolated from rats In vivo 5-weeks old male ICR mice | EGs: artepillin C, coumaric acid, and kaempferide (1, 10 μM), and propolis (1–104 ng/mL) CG1: insulin (100 nM) CG2: AICAR CG3: DMSO | 2-Deoxyglucose uptake, OGTT, maltase and sucrase-isomaltase activities in epithelial cells of the small intestinal, phosphorylation of AMPK, PI3K, AKT, AS160, IR, and GLUT4 translocation. | Polyphenols in propolis activated PI3K and AMPK signaling pathways and promoted GLUT4 translocation in L6 myotubes though only kaempferide increased glucose uptake. Propolis extract (In vitro, 1 μg) and In vivo significantly promoted the phosphorylation of IR, PI3K, and AMPK and increased GLUT4 translocation in rat skeletal muscle and subsequently decreased postprandial blood glucose levels. Propolis extract had no effect on α-glucosidase activity in the small intestine. | [70] |
| Gavage CAPE (5 and 10 mg/kg/d/5 d) | 6-7-weeks old male adult Wistar rats | EG: CAPE + eccentric exercise CG1: normal rats + propylene glycol in saline CG2: acute eccentric treadmill exercise | Serum creatine kinase levels, IL-1β, MCP-1, COX-2, iNOS, leukocyte infiltration, and the extent of muscle fiber damage (vacuolization and fragmentation). | CAPE decreased serum creatine kinase, protein nitrotyrosine, PARP activity, MDA, leukocyte infiltration, skeletal muscle cell fragmentation and vacuolization, muscle levels of COX2, iNOS, IL-1β, MCP-1, and p65NF-κB activity to levels in resting CG1 compared with CG2. | [101] |
| Dietary propolis (0.1% over 20 weeks) | MGO-induced muscle wasting in male C57BL/6NCr mice (4-weeks old) | EG: propolis + MGO CG1: MGO only CG2: propolis only CG3: untreated mice | Weight of EDL and soleus muscles, soleus and EDL levels of AGEs, inflammation-related molecules, and activity of glyoxalase 1. | Propolis had no effect on MGO-induced loss of EDL muscle but tended to increase the weight of the soleus muscle regardless of MGO treatment. Propolis decreased muscular levels of AGEs, IL-1β, IL-6, TLR4 and enhanced the activity of glyoxalase 1. | [38] |
| Dietary crude propolis (0.2% over 2 or 5 weeks) | HFD-induced muscle wasting in male C57BL/6 mice (4-weeks old) | EG: propolis + HFD CG1: HFD only CG2: untreated mice | 16S rRNA of gut microbiota, serum levels of LPS, triacylglycerols and glucose, and skeletal muscle levels of inflammatory cytokine TLR4 expression. | Propolis (5 weeks) significantly decreased serum triacylglycerols, glucose, circulating LPS and down-regulated the expression TLR4 and inflammatory cytokine in muscle. It countereacted the effect of HFD on gut microbiota. | [116] |
| Oral propolis water extract (50 mg/kg body weight/d/6 weeks) | 6-weeks old Sprague-Dawley rats | EG: propolis + eccentric exercise CG1: only eccentric exercise CG2: no treatment | Blood levels of glucose and insulin, MDA, SOD, GPX, and CAT in the liver and in the tissue of the liver and the gastrocnemius muscle. | Serum levels of glucose and insulin were significantly lower in EG and CG1 than CG2. Glycogen level in skeletal muscle was higher in EG and CG1 than CG2. Skeletal muscle levels of MDA were lower in EG than CG1 and CG2. Liver levels of SOD as well as gastrocnemius muscle levels of SOD, GPX and CAT were higher in EG only. | [109] |
| Gavage naturally-enriched milk with PUFA and propolis polyphenols (PUFA/P-M: 5 mL/kg body weight /85 d) | 21-d old male Wistar rats | EG: HFD + PUFA/P-M CG1: HFD + water CG2: HFD + whole milk CG3: HFD + PUFA milk CG 4: standard chow + water N.B. All treatments were repeated in absence of HFD | Weight gain, mass of internal organs and the soleus and gastrocnemius muscles, and glucose tolerance. | Among all treatments in obese rats, only PUFA/P-m increased gastrocnemius muscle mass (tended to increase soleus muscle mass) and mesenteric fat and tended to lower LDL levels. It decreased the size of adipocytes compared with all groups except PUFA milk with no effect on body weight. | [105] |
| Dietary propolis 4% (105 d) | Nile tilapia in net cages (males only) | EG: propolis rich diet CG: propolis free diet | Muscle morphometry and myostatin gene expression. | Propolis diet had no effect on muscle growth or myostatin gene expression at 35, 70, and 105 d. | [117] |
| Dietary propolis (1, 2, 3 and 4 g/kg of feed/45 d) | Nile tilapia post-larvae and fingerlings in tanks | EG: propolis rich diet CG: propolis free diet | Final weight, total and standard body length, survival, body composition, and intestinal villus height. | Propolis supplementation had no effect on weight, total and standard length, survival, and intestinal villus height. However, 2.6 g propolis/kg of feed significantly improved body protein deposition and body condition factor—an estimate of future growth. | [104] |
| Intraperitoneal CAPE (10 μM/kg 1 h before ischemia reperfusion) | Adult male Wistar rats undergoing ischemia reperfusion | EG: ischemia reperfusion + CAPE CG1: ischemia reperfusion CG2: sham | Neutrophil infiltration, serum creatine kinase, serum and gastrocnemius muscle levels of protein carbonyl, xanthine oxidase, and adenosine deaminase. | CAPE reduced neutrophil infiltration and serum creatine kinase as well as protein carbonyl, xanthine oxidase, and adenosine deaminase levels in the blood and gastrocnemius muscle. | [102,103] |
| Gavage propolis (1 g/kg body weight/d/2 weeks) | Adult male Wistar rats with 2-week hind limb unloading (HU) | EG: HU rats + propolis CG1: normal rats + propolis CG2: normal rats CG3: untreated HU rats | Soleus muscle weight, FCSA, myofiber number, apoptosis of endothelial cells, capillary to muscle fiber ratio, capillary number, luminal diameter, and capillary volume, levels of ROS, SOD-1, anti-angiogenic factors, and pro-angiogenic factors. | Propolis had no effect on soleus muscle weight or FCSA. However, the relative soleus muscle-to-body weight and the capillary to muscle fiber ratio of the soleus muscle were significantly higher in EG than in CG3. Propolis decreased the number of apoptotic endothelial cells, improved levels of SOD-1, ROS, and VEGF leading to increased capillary number, luminal diameter, and capillary volume in the EG to the levels of CG1 and CG2, which were all significantly different from CG3. | [110] |
| Gavage propolis (200-mg/kg body weight/d/28 d) | Adult female Wistar rats undergoing crush injuries of the sciatic nerve | EG: propolis CG1: curcumin CG2: methylprednisolone CG3: sham rats CG4: untreated rats with sciatic nerve injury | Gastrocnemius muscle mass, motor function, nerve fiber myelination, and nerve conduction. | Propolis and curcumin significantly restored gastrocnemius muscle mass, improved walking, nerve fiber myelination, and motor conduction to the gastrocnemius muscle compared with CG4. | [113] |
| Dietary fresh monofloral bee pollen 5% or 10% (3 weeks) | Malnourished old male Wistar rats (22-month-old) | EG1 and EG2: refeeding diet + bee pollen 5% and 10%, respectively CG1: refeeding diet CG2: no treatment CG3: untreated normal weight rats | Body weight and composition, muscle mass, muscle protein synthesis rate, plasma cytokines, mitochondrial enzyme activity, and mTOR/p70S6kinase/4eBP1 signaling. | Bee pollen restored visceral and subcutaneous adipose tissues and increased plantaris and gastrocnemius muscle mass. 10% pollen restored the levels of cytokines to normal, boosted muscle protein synthesis, and increased complex IV activity while both 5% and 10% increased the activity CS and the phosphorylation of mTOR/p70S6kinase/4eBP1 signaling. | [36] |
| Oral crude and processed monofloral Indian mustard bee pollen (100, 200, and 300 mg/kg body weight/4 weeks) | Adult male Wistar rats and Swiss albino mice | EG1: neat bee pollen + acute eccentric swimming EG2: processed bee pollen + acute eccentric swimming CG1: no treatment CG2: bee pollen only CG3: acute eccentric swimming + vehicle gum acacia | Body weight, relative weight of the gastrocnemius muscle, SOD, GSH, MDA, NO, total protein content, lipid peroxidation, myostatin mRNA, β-actin, mitochondrial complex I, II, III, and IV enzyme activity. | Crude (300 mg/kg) and processed (200 and 300 mg/kg) bee pollen prevented myofiber fragmentation and restored body weight and the relative weight of the gastrocnemius muscle as well as mitochondrial complex-I, -II, -III, and -IV enzyme activity to normal (CG1 and CG2) compared with CG3. Both bee pollen treatments decreased MDA, NO, total protein content, lipid peroxidation, and myostatin and increased SOD and GSH in skeletal muscle. | [90] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.M.; Kunugi, H. Apitherapy for Age-Related Skeletal Muscle Dysfunction (Sarcopenia): A Review on the Effects of Royal Jelly, Propolis, and Bee Pollen. Foods 2020, 9, 1362. https://doi.org/10.3390/foods9101362
Ali AM, Kunugi H. Apitherapy for Age-Related Skeletal Muscle Dysfunction (Sarcopenia): A Review on the Effects of Royal Jelly, Propolis, and Bee Pollen. Foods. 2020; 9(10):1362. https://doi.org/10.3390/foods9101362
Chicago/Turabian StyleAli, Amira Mohammed, and Hiroshi Kunugi. 2020. "Apitherapy for Age-Related Skeletal Muscle Dysfunction (Sarcopenia): A Review on the Effects of Royal Jelly, Propolis, and Bee Pollen" Foods 9, no. 10: 1362. https://doi.org/10.3390/foods9101362
APA StyleAli, A. M., & Kunugi, H. (2020). Apitherapy for Age-Related Skeletal Muscle Dysfunction (Sarcopenia): A Review on the Effects of Royal Jelly, Propolis, and Bee Pollen. Foods, 9(10), 1362. https://doi.org/10.3390/foods9101362