Antioxidant Characterization and Biological Effects of Grape Pomace Extracts Supplementation in Caenorhabditis elegans

 ,
,  and
and
Abstract
1. Introduction
2. Material and Methods
2.1. Standards and Reagents
2.2. Samples
2.3. Analysis of Total Phenolic Compounds
2.4. In Vitro Antioxidant Assays
2.5. Assays with C. elegans
2.5.1. Strains and Maintenance Conditions
2.5.2. Stress Assays
2.5.3. Accumulation of Reactive Oxygen Species (ROS)
2.5.4. Lifespan Assay
2.6. Statistical Analysis
3. Results and Discussion
3.1. Chemical and Antioxidant Characterization of the GP Extracts
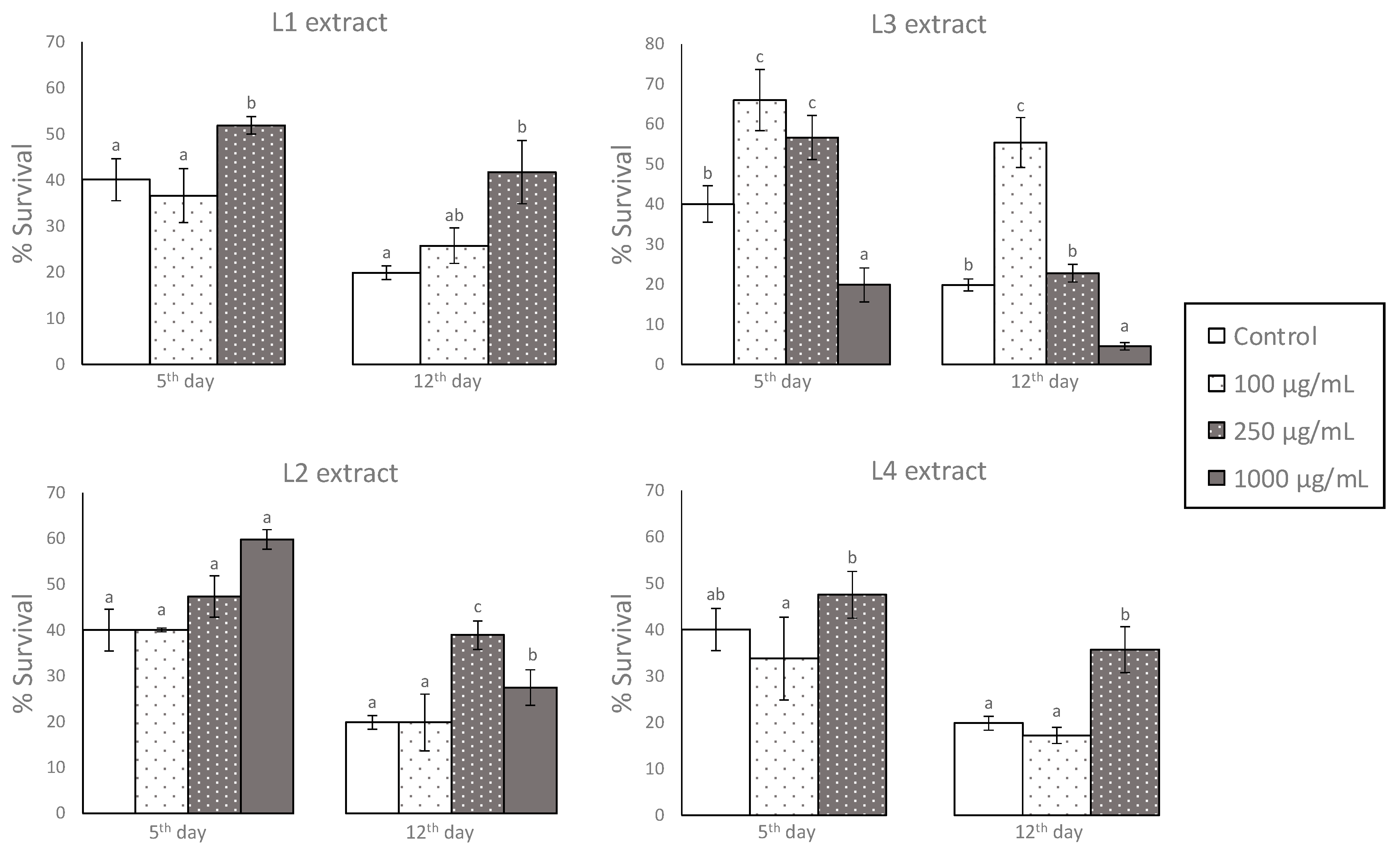
3.2. Effects of GP Extracts in the Resistance of C. elegans against Oxidative Damage
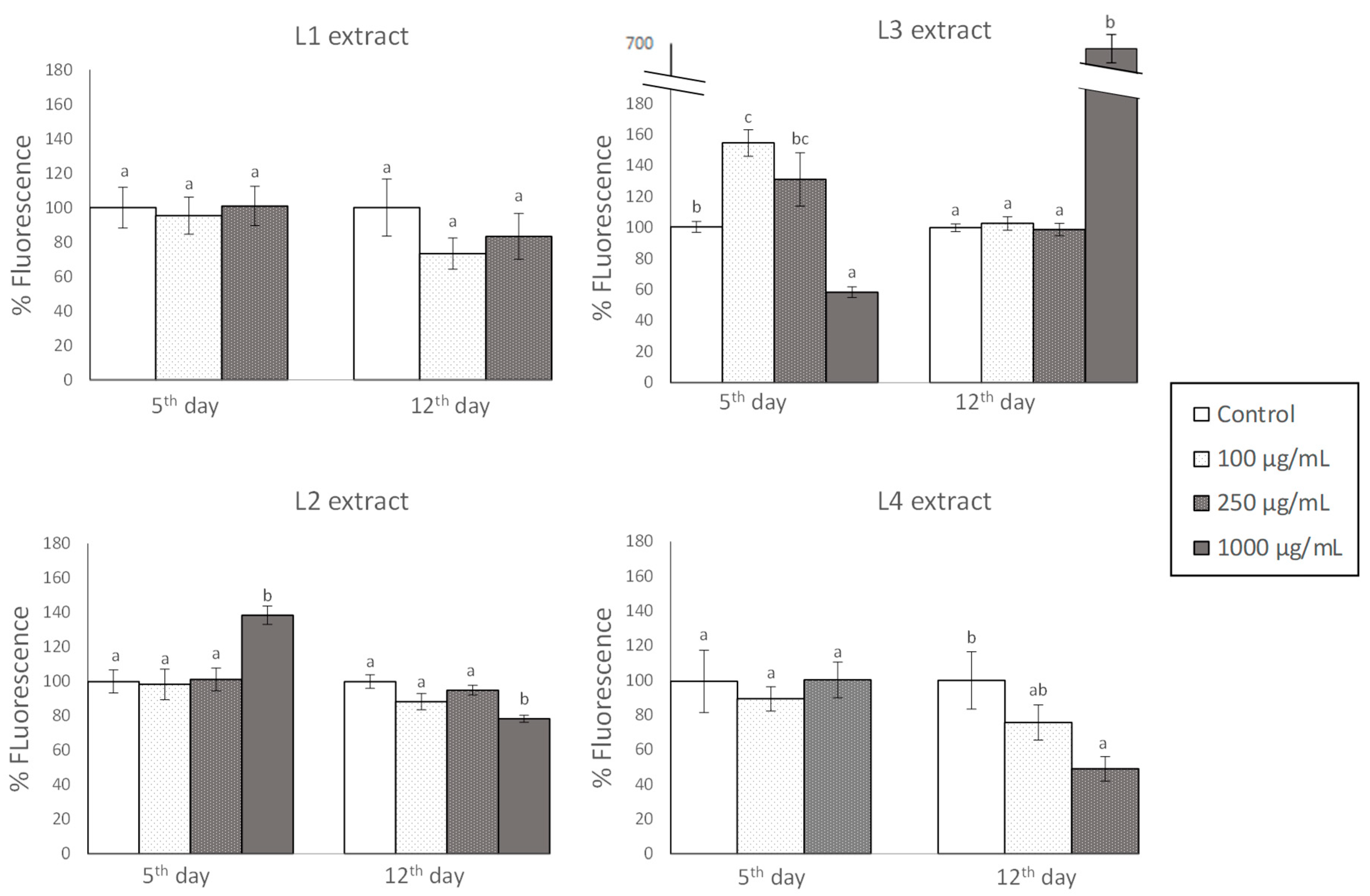
3.3. Effects of GP Extracts in the Accumulation of Reactive Oxygen Species (ROS) in C. elegans
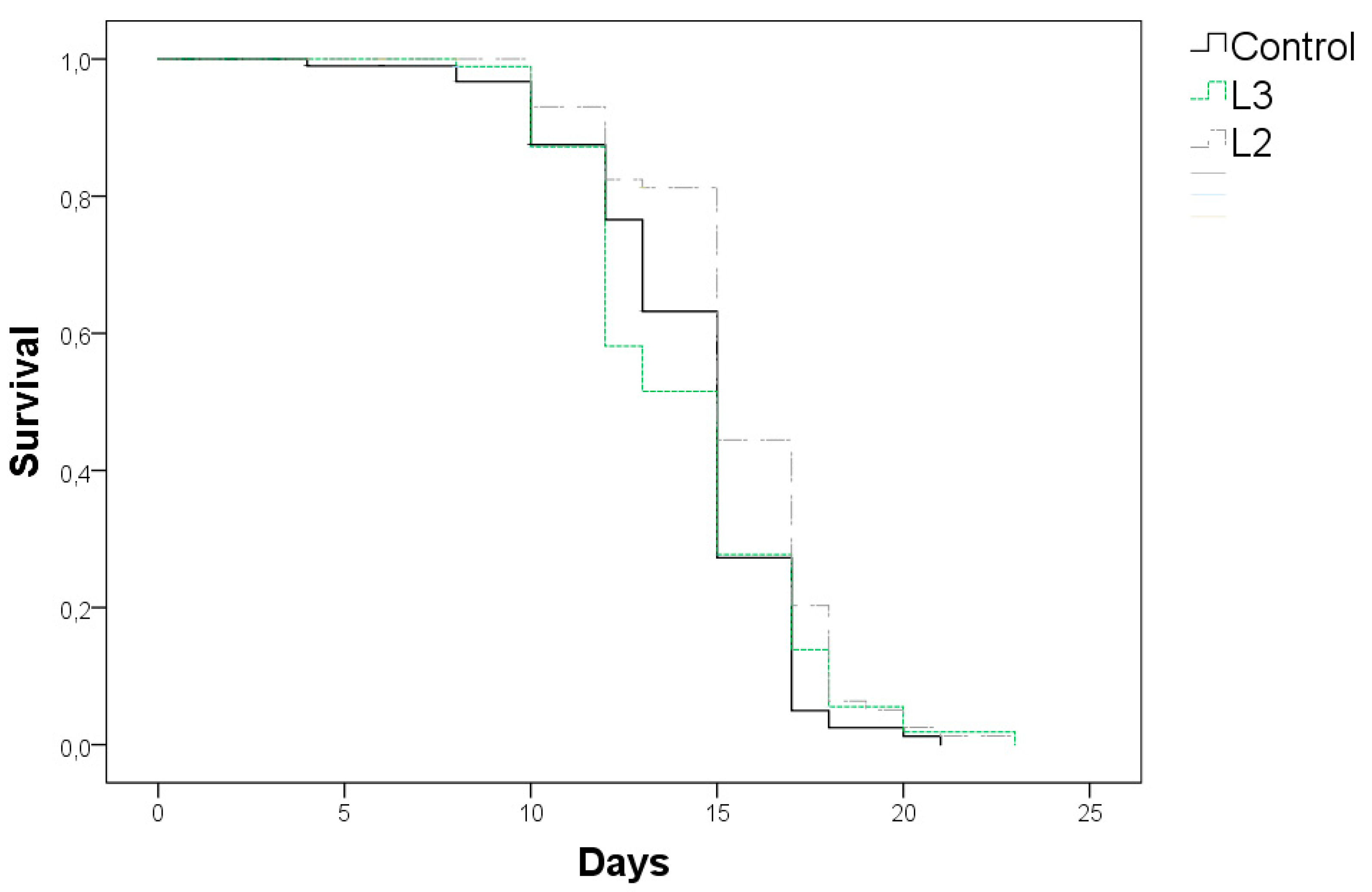
3.4. Effects of GP Extracts in C. elegans Lifespan
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhu, F.; Du, B.; Zheng, L.; Li, J. Advance on the bioactivity and potential applications of dietary fibre from grape pomace. Food Chem. 2015, 186, 207–212. [Google Scholar] [CrossRef] [PubMed]
- González-Paramás, A.M.; Esteban-Ruano, S.; Santos-Buelga, C.; De Pascual-Teresa, S.; Rivas-Gonzalo, J.-C. Flavanol Content and Antioxidant Activity in Winery Byproducts. J. Agric. Food Chem. 2004, 52, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, J.; Hekimi, S. When a theory of aging ages badly. Cell. Mol. Life Sci. 2010, 67, 1–8. [Google Scholar] [CrossRef]
- Asthana, J.; Mishra, B.N.; Pandey, R. Acacetin promotes healthy aging by altering stress response in Caenorhabditis elegans. Free Radic. Res. 2016, 50, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Queen, B.L.; Tollefsbol, T.O. Polyphenols and Aging. Curr. Aging Sci. 2010, 3, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Guarente, L.; Kenyon, C. Genetic pathways that regulate ageing in model organisms. Nature 2000, 408, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Braeckman, B. Insulin-like signaling, metabolism, stress resistance and aging in Caenorhabditis elegans. Mech. Ageing Dev. 2001, 122, 673–693. [Google Scholar] [CrossRef]
- Surco-Laos, F.; Dueñas, M.; González-Manzano, S.; Cabello, J.; Santos-Buelga, C.; González-Paramás, A.M.; Duenas-Paton, M. Influence of catechins and their methylated metabolites on lifespan and resistance to oxidative and thermal stress of Caenorhabditis elegans and epicatechin uptake. Food Res. Int. 2012, 46, 514–521. [Google Scholar] [CrossRef]
- Upadhyay, A.; Chompoo, J.; Taira, N.; Fukuta, M.; Tawata, S. Significant Longevity-Extending Effects of Alpinia zerumbet Leaf Extract on the Life Span of Caenorhabditis elegans. Biosci. Biotechnol. Biochem. 2013, 77, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Dinh, J.; Angeloni, J.T.; Pederson, D.B.; Wang, X.; Cao, M.; Dong, Y. Cranberry Extract Standardized for Proanthocyanidins Promotes the Immune Response of Caenorhabditis elegans to Vibrio cholerae through the p38 MAPK Pathway and HSF-1. PLoS ONE 2014, 9, e103290. [Google Scholar] [CrossRef]
- Jara-Palacios, M.J.; Escudero-Gilete, M.L.; Heredia, F.J.; González-Manzano, S.; Hernanz, D.; Duenas-Paton, M.; González-Paramás, A.M.; Santos-Buelga, C. Study of Zalema Grape Pomace: Phenolic Composition and Biological Effects in Caenorhabditis elegans. J. Agric. Food Chem. 2013, 61, 5114–5121. [Google Scholar] [CrossRef] [PubMed]
- Cueva, C.; Guadarrama, A.; Bartolomé, B.; Gil-Sánchez, I.; Ayuda-Durán, B.; González-Manzano, S.; Santos-Buelga, C.; Sanz-Buenhombre, M.; Martín-Cabrejas, M.A.; Moreno-Arribas, M.V. Chemical characterization and in vitro colonic fermentation of grape pomace extracts. J. Sci. Food Agric. 2017, 97, 3433–3444. [Google Scholar]
- Gomes, A.D.C.; Figueiredo, C.M.; Silva, L.P.; Peixoto, E.T.D.M. Antioxidant activity evaluation and determination of total phenols and flavonoids in Humulus lupulus. Planta Med. 2016, 82, S1–S381. [Google Scholar]
- Benzie, I.F.; Strain, J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Bruskov, V.I. Heat-induced formation of reactive oxygen species and 8-oxoguanine, a biomarker of damage to DNA. Nucleic Acids Res. 2002, 30, 1354–1363. [Google Scholar] [CrossRef]
- Wang, H.; A Joseph, J. Quantifying cellular oxidative stress by dichlorofluorescein assay using microplate reader. Free Radic. Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef]
- Ruberto, G.; Renda, A.; Daquino, C.; Amico, V.; Spatafora, C.; Tringali, C.; De Tommasi, N. Polyphenol constituents and antioxidant activity of grape pomace extracts from five Sicilian red grape cultivars. Food Chem. 2007, 100, 203–210. [Google Scholar] [CrossRef]
- Muzolf, M.; Szymusiak, H.; Gliszczyńska-Świgło, A.; Rietjens, I.M.; Tyrakowska, B. pH-Dependent Radical Scavenging Capacity of Green Tea Catechins. J. Agric. Food Chem. 2008, 56, 816–823. [Google Scholar] [CrossRef]
- Hogan, S.; Zhang, L.; Li, J.; Zoecklein, B.; Zhou, K. Antioxidant properties and bioactive components of Norton (Vitis aestivalis) and Cabernet Franc (Vitis vinifera) wine grapes. LWT Food Sci. Technol. 2009, 42, 1269–1274. [Google Scholar] [CrossRef]
- Rockenbach, I.I.; Rodrigues, E.; Gonzaga, L.V.; Caliari, V.; Genovese, M.I.; Gonçalves, A.E.D.S.S.; Fett, R. Phenolic compounds content and antioxidant activity in pomace from selected red grapes (Vitis vinifera L. and Vitis labrusca L.) widely produced in Brazil. Food Chem. 2011, 127, 174–179. [Google Scholar] [CrossRef]
- Xiong, L.-G.; Yu, P.-H.; Gong, Y.-S.; Huang, J.-A.; Li, J.; Xiong, Z.; Zhang, J.-W.; Liu, Z.-H.; Chen, J.-H. Black Tea Increased Survival of Caenorhabditis elegans under Stress. J. Agric. Food Chem. 2014, 62, 11163–11169. [Google Scholar] [CrossRef]
- Martorell, P.; Forment, J.V.; De Llanos, R.; Montón, F.; Llopis, S.; González, N.; Genovés, S.; Cienfuegos, E.; Monzó, H.; Ramón, D. Use of Saccharomyces cerevisiae and Caenorhabditis elegans as Model Organisms To Study the Effect of Cocoa Polyphenols in the Resistance to Oxidative Stress. J. Agric. Food Chem. 2011, 59, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, K.; Chakrabarti, S.; Steinberg, C.E.W.; Saul, N.; Stürzenbaum, S.R.; Menzel, R. Hormetins, antioxidants and prooxidants: Defining quercetin-, caffeic acid- and rosmarinic acid-mediated life extension in C. elegans. Biogerontology 2011, 12, 329–347. [Google Scholar] [CrossRef]
- Dueñas, M.; Surco-Laos, F.; González-Manzano, S.; González-Paramás, A.M.; Gómez-Orte, E.; Cabello, J.; Santos-Buelga, C. Deglycosylation is a key step in biotransformation and lifespan effects of quercetin-3-O-glucoside in Caenorhabditis elegans. Pharmacol. Res. 2013, 76, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Saul, N.; Pietsch, K.; Stürzenbaum, S.R.; Menzel, R.; Steinberg, C.E.W. Diversity of polyphenol action in Caenorhabditis elegans: Between toxicity and longevity. J. Nat. Prod. 2011, 74, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Are polyphenols antioxidants or pro-oxidants? What do we learn from cell culture and in vivo studies? Arch. Biochem. Biophys. 2008, 476, 107–112. [Google Scholar] [CrossRef]
- Tang, S.T.; Halliwell, B. Medicinal plants and antioxidants: What do we learn from cell culture and Caenorhabditis elegans studies? Biochem. Biophys. Res. Commun. 2010, 394, 1–5. [Google Scholar] [CrossRef]
- Okamoto, T.; Kobayashi, R.; Natsume, M.; Nakazato, K. Habitual cocoa intake reduces arterial stiffness in postmenopausal women regardless of intake frequency: A randomized parallel-group study. CIA 2016, 11, 1645–1652. [Google Scholar] [CrossRef]
- Howe, P.R.C.; Davison, K. Potential Implications of Dose and Diet for the Effects of Cocoa Flavanols on Cardiometabolic Function. J. Agric. Food Chem. 2015, 63, 9942–9947. [Google Scholar]
- Saul, N.; Pietsch, K.; Menzel, R.; Sturzenbaum, S.R.; Steinberg, C.E. Catechin induced longevity in C. elegans: From key regulator genes to disposable soma. Mech. Ageing Dev. 2009, 130, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Abbas, S.; Wink, M. Green Tea Extract Induces the Resistance of Caenorhabditis elegans against Oxidative Stress. Antioxidants 2014, 3, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Surco-Laos, F.; Cabello, J.; Gómez-Orte, E.; González-Manzano, S.; González-Paramás, A.M.; Santos-Buelga, C.; Duenas-Paton, M. Effects of O-methylated metabolites of quercetin on oxidative stress, thermotolerance, lifespan and bioavailability on Caenorhabditis elegans. Food Funct. 2011, 2, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Ayuda-Durán, B.; González-Manzano, S.; Miranda-Vizuete, A.; Dueñas, M.; Santos-Buelga, C.; González-Paramás, A.M. Epicatechin modulates stress-resistance in C. elegans via insulin/IGF-1 signaling pathway. PLoS ONE 2019, 14, e0199483. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.-G.; Chen, Y.-J.; Tong, J.-W.; Gong, Y.-S.; Huang, J.-A.; Liu, Z.-H. Epigallocatechin-3-gallate promotes healthy lifespan through mitohormesis during early-to-mid adulthood in Caenorhabditis elegans. Redox Biol. 2018, 14, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Schulz, T.J.; Zarse, K.; Voigt, A.; Urban, N.; Birringer, M.; Ristow, M. Glucose Restriction Extends Caenorhabditis elegans Life Span by Inducing Mitochondrial Respiration and Increasing Oxidative Stress. Cell Metab. 2007, 6, 280–293. [Google Scholar] [CrossRef] [PubMed]
- Gems, D.; Partridge, L. Stress-Response Hormesis and Aging: “That which Does Not Kill Us Makes Us Stronger”. Cell Metab. 2008, 7, 200–203. [Google Scholar] [CrossRef]
- Fu, X.; Tang, Y.; Dickinson, B.C.; Chang, C.J.; Chang, Z. An oxidative fluctuation hypothesis of aging generated by imaging H2O2 levels in live Caenorhabditis elegans with altered lifespans. Biochem. Biophys. Res. Commun. 2015, 458, 896–900. [Google Scholar] [CrossRef]
- Desjardins, D.; Yee, C.; Bernard, K.; Khaki, A.; Breton, L.; Cacho-Valadez, B.; Liu, J.-L.; Wang, Y.; Hekimi, S. Antioxidants reveal an inverted U-shaped dose-response relationship between reactive oxygen species levels and the rate of aging in Caenorhabditis elegans. Aging Cell 2016, 16, 104–112. [Google Scholar] [CrossRef]
- Van Raamsdonk, J.M.; Hekimi, S. Superoxide dismutase is dispensable for normal animal lifespan. Proc. Natl. Acad. Sci. USA 2012, 109, 5785–5790. [Google Scholar] [CrossRef]
- Noë, A.; Hekimi, S.; Wang, Y. Mitochondrial ROS and the Effectors of the Intrinsic Apoptotic Pathway in Aging Cells: The Discerning Killers. Front. Genet. 2016, 7, 1882. [Google Scholar]
- Benzie, I.F.; Choi, S.-W. Antioxidants in Food. In Advances in Food and Nutrition Research; Elsevier BV: Amsterdam, The Netherlands, 2014; Volume 71, pp. 1–53. [Google Scholar]
- Wang, H.; Li, D.; Hu, Z.; Zhao, S.; Zheng, Z. Protective Effects of Green Tea Polyphenol Against Renal Injury Through ROS-Mediated JNK-MAPK Pathway in Lead Exposed Rats. Mol. Cells 2016, 39, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Bartholome, A.; Kampkötter, A.; Tanner, S.; Sies, H.; Klotz, L.-O. Epigallocatechin gallate-induced modulation of FoxO signaling in mammalian cells and C. elegans: FoxO stimulation is masked via PI3K/Akt activation by hydrogen peroxide formed in cell culture. Arch. Biochem. Biophys. 2010, 501, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Sunagawa, T.; Kanda, T.; Tagashira, M.; Sami, M.; Shimizu, T.; Shirasawa, T. Procyanidins from Apples (Malus pumila Mill.) Extend the Lifespan of Caenorhabditis elegans. Planta Med. 2010, 77, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Grünz, G.; Haas, K.; Soukup, S.; Klingenspor, M.; Kulling, S.E.; Daniel, H.; Spanier, B. Structural features and bioavailability of four flavonoids and their implications for lifespan-extending and antioxidant actions in C. elegans. Mech. Ageing Dev. 2012, 133, 1–10. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Polyphenols (mg Gallic Acid Equivalents/g dw) | |
|---|---|
| L1 | 37.4 |
| L2 | 26.8 |
| L3 | 71.3 |
| L4 | 26.8 |
| ABTS (µmol TE/g Dry Extract) | FRAP (µmol TE/g Dry Extract) | |
|---|---|---|
| L1 | 420 ± 10 b | 220 ± 20 b |
| L2 | 400 ± 20 b | 170 ± 20 b |
| L3 | 840 ± 20 a | 530 ± 60 a |
| L4 | 300 ± 50 b | 110 ± 10 b |
| Polyphenols per Plate | ||||
|---|---|---|---|---|
| Concentration of Extracts per Plate | L1 | L2 | L3 | L4 |
| 100 µg/mL | 3.74 µg GAE | 2.68 µg GAE | 7.13 µg GAE | 2.68 µg GAE |
| 250 µg/mL | 9.36 µg GAE | 6.70 µg GAE | 17.83 µg GAE | 6.70 µg GAE |
| 1000 µg/mL | - | 26.79 µg GAE | 71.30 µg GAE | - |
| Treatment | Mean (Days) a | p vs. Control (Log-Rank) | Maximun 10% (Days) b | p vs. Control (Anova) |
|---|---|---|---|---|
| Control | 14.3 ± 0.30 | 17.7 ± 1.26 | ||
| L3 100 µg/mL | 14.2 ± 0.35 | 0.96 | 19.1 ± 1.52 | 0.047 |
| L2 250 µg/mL | 15.6 ± 0.18 | 0.00 | 19.5 ± 1.41 | 0.007 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayuda-Durán, B.; González-Manzano, S.; Gil-Sánchez, I.; Moreno-Arribas, M.V.; Bartolomé, B.; Sanz-Buenhombre, M.; Guadarrama, A.; Santos-Buelga, C.; González-Paramás, A.M. Antioxidant Characterization and Biological Effects of Grape Pomace Extracts Supplementation in Caenorhabditis elegans. Foods 2019, 8, 75. https://doi.org/10.3390/foods8020075
Ayuda-Durán B, González-Manzano S, Gil-Sánchez I, Moreno-Arribas MV, Bartolomé B, Sanz-Buenhombre M, Guadarrama A, Santos-Buelga C, González-Paramás AM. Antioxidant Characterization and Biological Effects of Grape Pomace Extracts Supplementation in Caenorhabditis elegans. Foods. 2019; 8(2):75. https://doi.org/10.3390/foods8020075
Chicago/Turabian StyleAyuda-Durán, Begoña, Susana González-Manzano, Irene Gil-Sánchez, M. Victoria Moreno-Arribas, Begoña Bartolomé, Marisa Sanz-Buenhombre, Alberto Guadarrama, Celestino Santos-Buelga, and Ana M. González-Paramás. 2019. "Antioxidant Characterization and Biological Effects of Grape Pomace Extracts Supplementation in Caenorhabditis elegans" Foods 8, no. 2: 75. https://doi.org/10.3390/foods8020075
APA StyleAyuda-Durán, B., González-Manzano, S., Gil-Sánchez, I., Moreno-Arribas, M. V., Bartolomé, B., Sanz-Buenhombre, M., Guadarrama, A., Santos-Buelga, C., & González-Paramás, A. M. (2019). Antioxidant Characterization and Biological Effects of Grape Pomace Extracts Supplementation in Caenorhabditis elegans. Foods, 8(2), 75. https://doi.org/10.3390/foods8020075