

Storage Stability of a Multifunctional Fermented Blend Based on Sacha Inchi (Plukenetia volubilis) Oil Press Cake and Yacon (Smallanthus sonchifolius) Flour: Physicochemical Properties, Bioactivity, and Prebiotic–Probiotic Potential
 , , , and
, , , and
Abstract
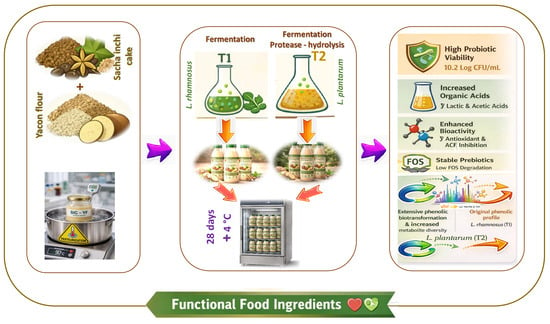
1. Introduction
2. Materials and Methods
2.1. Chemicals and Raw Materials
2.2. Experimental Setup
2.2.1. Inoculum and Fermentation Preparation
2.2.2. Stability During Storage
2.3. Analytical Determinations
2.3.1. Determination of pH and Titratable Acidity
2.3.2. Viable Cell Count
2.3.3. Analysis of Lactic and Acetic Acids
2.3.4. Crude and Soluble Protein Analysis
2.3.5. Glucose, Fructose, Sucrose, and Fructooligosaccharide (FOS) Analysis
2.3.6. Determination of Soluble Solids, Total Solids, and Dietary Fiber
2.3.7. In Vitro Antioxidant Capacity
2.3.8. Antihypertensive Activity
2.3.9. Determination of Total Phenolic Content and Phenolic Profile
2.4. Statistical Analysis
3. Results
3.1. Acidification of the Medium and Cell Viability During Storage
3.2. Impact of Storage on Lactic Acid and Acetic Acid Contents
3.3. Changes in Soluble and Crude Protein During Storage
3.4. Changes in Fructooligosaccharides, Sugars, Soluble and Total Solids, and Fiber During Storage
3.5. Evaluation of the Stability of Phenolic Compounds During Storage and Identification of Phenolic Compounds
Comparative Phenolic Profile of the SIC–YF Mixture Before and After Fermentation
3.6. Stability of Antioxidant and Antihypertensive Properties Throughout Storage
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Essa, M.M.; Bishir, M.; Bhat, A.; Chidambaram, S.B.; Al-Balushi, B.; Hamdan, H.; Govindarajan, N.; Freidland, R.P.; Qoronfleh, M.W. Functional foods and their impact on health. J. Food Sci. Technol. 2023, 60, 820–834. [Google Scholar] [CrossRef]
- Hidalgo-Fuentes, B.; de Jesús-José, E.; Cabrera-Hidalgo, A.d.J.; Sandoval-Castilla, O.; Espinosa-Solares, T.; González-Reza, R.M.; Zambrano-Zaragoza, M.L.; Liceaga, A.M.; Aguilar-Toalá, J.E. Plant-based fermented beverages: Nutritional composition, sensory properties, and health benefits. Foods 2024, 13, 844. [Google Scholar] [CrossRef]
- Zhao, Y.S.; Eweys, A.S.; Zhang, J.Y.; Zhu, Y.; Bai, J.; Darwesh, O.M.; Zhang, H.-B.; Xiao, X. Fermentation affects antioxidant activity of plant-based food materials through release and production of bioactive components. Antioxidants 2021, 10, 2004. [Google Scholar] [CrossRef] [PubMed]
- Adebo, O.A.; Medina-Meza, I.G. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [PubMed]
- Sathe, S.K.; Kshirsagar, H.H.; Sharma, G.M. Solubilization, fractionation, and electrophoretic characterization of Inca peanut (Plukenetia volubilis L.) proteins. Plant Foods Hum. Nutr. 2012, 67, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Torres Sánchez, E.G.; Hernández-Ledesma, B.; Gutiérrez, L.F. Sacha inchi oil press-cake: Physicochemical characteristics, food-related applications and biological activity. Food Rev. Int. 2021, 39, 148–159. [Google Scholar] [CrossRef]
- Chirinos, R.; Pedreschi, R.; Campos, D. Enzyme-assisted hydrolysates from sacha inchi (Plukenetia volubilis) protein with in vitro antioxidant and antihypertensive properties. J. Food Process. Preserv. 2020, 44, e14969. [Google Scholar] [CrossRef]
- Chirinos, R.; Figueroa-Merma, A.; Valente de Oliveira, T.; Guzmán, F.; Aguilar-Galvez, A.; Carpentier, S.; Pedreschi, R.; Campos, D. Bioactive Peptides from Sacha Inchi (Plukenetia volubilis) Cake: Extraction Techniques and Their Antioxidant, Antihypertensive, Hypoglycemic, and Iron-Chelating Potential through In Vitro and In Silico Analysis. Appl. Food Res. 2026, 6, 101612. [Google Scholar] [CrossRef]
- Campos, D.; Valerio, K.; Carrasco, M.P.; Aguilar-Galvez, A.; Chirinos, R.; Pedreschi, R. Symbiotic and multifunctional properties of a fermented blend based on sacha inchi press cake (Plukenetia volubilis) and yacon (Smallanthus sonchifolius). Food Biosci. 2025, 74, 107886. [Google Scholar] [CrossRef]
- Campos, D.; Betalleluz-Pallardel, I.; Chirinos, R.; Aguilar-Galvez, A.; Noratto, G.; Pedreschi, R. Prebiotic effects of yacon (Smallanthus sonchifolius Poepp. & Endl.), a source of fructooligosaccharides and phenolic compounds with antioxidant activity. Food Chem. 2012, 135, 1592–1599. [Google Scholar] [CrossRef]
- Kumar, A.; Saranyadevi, S.; Thirumalaisamy, S.K.; Dapana Durage, T.T.; Jaiswal, S.G.; Kavitake, D.; Wei, S. Phenolic acids in fermented foods: Microbial biotransformation, antioxidant mechanisms, and functional health implications. Front. Mol. Biosci. 2025, 12, 1678673. [Google Scholar] [CrossRef]
- Alexandre, E.M.; Aguiar, N.F.; Voss, G.B.; Pintado, M.E. Properties of fermented beverages from food wastes/by-products. Beverages 2023, 9, 45. [Google Scholar] [CrossRef]
- Campos, D.; Aguilar-Galvez, A.; Pedreschi, R. Stability of fructooligosaccharides, sugars and colour of yacon (Smallanthus sonchifolius) roots during blanching and drying. Int. J. Food Sci. Technol. 2016, 51, 1177–1185. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; Horwitz, W., Latimer, G.W., Eds.; AOAC International: Gaithersburg, MD, USA, 2007. [Google Scholar]
- Baba, A.S.; Najarian, A.; Shori, A.B.; Lit, K.W.; Keng, G.A. In vitro inhibition of key enzymes related to diabetes and hypertension in Lycium barbarum yogurt. Arab. J. Sci. Eng. 2014, 39, 5355–5362. [Google Scholar] [CrossRef]
- Aguilar-Galvez, A.; Guillermo, S.; Dubois-Dauphin, R.; Campos, D.; Thonart, P. The influence of growth conditions on enterocin-like production by Enterococcus faecium CWBI-B1430 and Enterococcus mundtii CWBI-B1431 isolates from artisanal Peruvian cheeses. Ann. Microbiol. 2011, 61, 955–964. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Jaime, L.; Martín-Cabrejas, M.A.; Mollá, E.; López-Andréu, F.J.; Esteban, R.M. Effect of storage on fructan and fructooligosaccharide of onion (Allium cepa L.). J. Agric. Food Chem. 2001, 49, 982–988. [Google Scholar] [CrossRef]
- Pedreschi, R.; Campos, D.; Noratto, G.; Chirinos, R.; Cisneros-Zevallos, L. Andean yacon root (Smallanthus sonchifolius Poepp. & Endl.) fructooligosaccharides as a potential novel source of prebiotics. J. Agric. Food Chem. 2003, 51, 5278–5284. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 20th ed.; AOAC International: Gaithersburg, MD, USA, 2016. [Google Scholar]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Wu, H.; Rui, X.; Li, W.; Xiao, Y.; Zhou, J.; Dong, M. Whole-grain oats (Avena sativa L.) as a carrier of lactic acid bacteria and a source of ACE-inhibitory peptides through solid-state fermentation. Food Funct. 2018, 9, 2270–2281. [Google Scholar] [CrossRef] [PubMed]
- Chirinos, R.; Cerna, E.; Pedreschi, R.; Calsin, M.; Aguilar-Galvez, A.; Campos, D. Multifunctional in vitro bioactive properties: Antioxidant, antidiabetic and antihypertensive properties of protein hydrolysates from tarwi (Lupinus mutabilis Sweet) obtained by enzymatic biotransformation. Cereal Chem. 2021, 98, 423–433. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Campos, D.; García, D.; Aguilar, A.; Chirinos, R.; Pedreschi, R. Comparison of conventional and ultrasound-assisted extractions of polyphenols from Inca muña (Clinopodium bolivianum) and their characterization using UPLC–PDA-ESI–Q/TOF–MSn technique. J. Food Process. Preserv. 2022, 46, e16310. [Google Scholar] [CrossRef]
- Shori, A.B.; Aljohani, G.S.; Al-Zahrani, A.J.; Al-Sulbi, O.S.; Baba, A.S. Viability of probiotics and antioxidant activity of cashew milk-based yogurt fermented with selected strains of probiotic Lactobacillus spp. LWT 2022, 153, 112482. [Google Scholar] [CrossRef]
- Coda, R.; Lanera, A.; Trani, A.; Gobbetti, M.; Di Cagno, R. Yogurt-like beverages made of a mixture of cereals, soy and grape must: Microbiology, texture, nutritional and sensory properties. Int. J. Food Microbiol. 2012, 155, 120–127. [Google Scholar] [CrossRef]
- Grasso, N.; Alonso-Miravalles, L.; O’Mahony, J.A. Composition, physicochemical and sensorial properties of commercial plant-based yogurts. Foods 2020, 9, 252. [Google Scholar] [CrossRef]
- Annunziata, G.; Arnone, A.; Ciampaglia, R.; Tenore, G.C.; Novellino, E. Fermentation of foods and beverages as a tool for increasing availability of bioactive compounds: Focus on short-chain fatty acids. Foods 2020, 9, 999. [Google Scholar] [CrossRef]
- Salmerón, I.; Thomas, K.; Pandiella, S.S. Effect of potentially probiotic lactic acid bacteria on the physicochemical composition and acceptance of fermented cereal beverages. J. Funct. Foods 2015, 15, 106–115. [Google Scholar] [CrossRef]
- Kücükgöz, K.; Franczak, A.; Borysewicz, W.; Kamińska, K.; Salman, M.; Mosiej, W.; Trząskowska, M. Impact of lactic acid fermentation on organic acids and sugars of oat and buckwheat beverages. Fermentation 2024, 10, 373. [Google Scholar] [CrossRef]
- Cui, Q.; Sun, Y.; Zhou, Z.; Cheng, J.; Guo, M. Effects of enzymatic hydrolysis on physicochemical properties and solubility and bitterness of milk protein hydrolysates. Foods 2021, 10, 2462. [Google Scholar] [CrossRef]
- Yousefi, N.; Shokrollahi Yancheshmeh, B.; Gernaey, K.V. The potential of fermentation-based processing on protein modification: A review. Foods 2025, 14, 3461. [Google Scholar] [CrossRef]
- Deziderio, M.A.; de Souza, H.F.; Kamimura, E.S.; Petrus, R.R. Plant-based fermented beverages: Development and characterization. Foods 2023, 12, 4128. [Google Scholar] [CrossRef]
- Chun, S.C.; Tan, W.Q.; Choo, W.S. Effect of prebiotic supplementation on the production of fermented sugarcane juice with Lactiplantibacillus plantarum and Lacticaseibacillus rhamnosus. Bioact. Carbohydr. Diet. Fibre 2025, 34, 100500. [Google Scholar] [CrossRef]
- Fuhren, J.; Schwalbe, M.; Rösch, C.; Nijland, R.; Wels, M.; Schols, H.A.; Kleerebezem, M. Dietary inulin increases Lactiplantibacillus plantarum strain Lp900 persistence in rats depending on dietary calcium level. Appl. Environ. Microbiol. 2021, 87, e00122-21. [Google Scholar] [CrossRef]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104, S1–S63. [Google Scholar] [CrossRef]
- Cunningham, M.; Vinderola, G.; Charalampopoulos, D.; Lebeer, S.; Sanders, M.E.; Grimaldi, R. Applying probiotics and prebiotics in new delivery formats—Is the clinical evidence transferable? Trends Food Sci. Technol. 2021, 112, 495–506. [Google Scholar] [CrossRef]
- Luana, N.; Rossana, C.; Curiel, J.A.; Kaisa, P.; Marco, G.; Rizzello, C.G. Manufacture and characterization of a yogurt-like beverage made with oat flakes fermented by selected lactic acid bacteria. Int. J. Food Microbiol. 2014, 185, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Rai, D.; Pandey, R.K.; Maurya, A.K.; Rai, D.C.; Kumar, D.; Tiwari, M. Characterization and antioxidant property of cereal-enriched bio-yogurt. J. Pure Appl. Microbiol. 2016, 10, 3071–3078. [Google Scholar] [CrossRef]
- Takenaka, M.; Ono, H. Novel octulosonic acid derivatives in the composite Smallanthus sonchifolius. Tetrahedron Lett. 2003, 44, 999–1002. [Google Scholar] [CrossRef]
- Pacheco, M.T.; Escribano-Bailón, M.T.; Moreno, F.J.; Villamiel, M.; Dueñas, M. Determination by HPLC-DAD-ESI/MSn of phenolic compounds in Andean tubers grown in Ecuador. J. Food Compos. Anal. 2019, 84, 103258. [Google Scholar] [CrossRef]
- Cuellar, N.V.P.; Hern, I.P.; Rey, D.V.; Rodríguez, K.N.A. Sacha inchi (Plukenetia volubilis L.) husks and seed shells as a source of phenolic compounds with potential health benefits. Univ. Sci. 2024, 29, 275–291. [Google Scholar] [CrossRef]
- Chirinos, R.; Necochea, O.; Pedreschi, R.; Campos, D. Sacha inchi (Plukenetia volubilis L.) shell: An alternative source of phenolic compounds and antioxidants. Int. J. Food Sci. Technol. 2016, 51, 986–993. [Google Scholar] [CrossRef]
- Heng, X.; Chen, H.; Lu, C.; Feng, T.; Li, K.; Gao, E. Study on synergistic fermentation of bean dregs and soybean meal by multiple strains and proteases. LWT 2022, 154, 112626. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, X.; Xu, Y.; Liu, B.; Meng, X. Effect of lactic acid bacteria fermentation on biotransformation of phenolic compounds and bioactivities of hawthorn pulp based on metabolomics. Process Biochem. 2025, 153, 304–314. [Google Scholar] [CrossRef]
- Chen, X.; Yuan, M.; Wang, Y.; Zhou, Y.; Sun, X. Influence of fermentation with different lactic acid bacteria and in vitro digestion on the change of phenolic compounds in fermented kiwifruit pulps. Int. J. Food Sci. Technol. 2022, 57, 2670–2679. [Google Scholar] [CrossRef]
- Wongsa, P.; Yuenyongrattanakorn, K.; Pongvachirint, W.; Auntalarok, A. Improving anti-hypertensive properties of plant-based alternatives to yogurt fortified with rice protein hydrolysate. Heliyon 2022, 8, e11087. [Google Scholar] [CrossRef] [PubMed]
- Simsek, S.; El, S.N.; Kilinc, A.K.; Karakaya, S. Vegetable and fermented vegetable juices containing germinated seeds and sprouts of lentil and cowpea. Food Chem. 2014, 156, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Aluko, R.; Muir, E. Improved method for direct high-performance liquid chromatography assay of angiotensin-converting enzyme-catalyzed reactions. J. Chromatogr. A 2002, 950, 125–130. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | T1 (Day 0) | T1 (Day 28) | T2 (Day 0) | T2 (Day 28) |
|---|---|---|---|---|
| Soluble solids (°Brix) | 7.63 ± 0.18 a | 7.40 ± 0.16 a | 8.03 ± 0.17 a | 7.97 ± 0.11 a |
| Total solids (%) | 14.37 ± 0.15 a | 14.40 ± 0.41 a | 14.80 ± 1.11 a | 14.64 ± 1.25 a |
| Soluble fiber (%) | 0.87 ± 0.04 a | 0.89 ± 0.07 a | 0.87 ± 0.04 a | 0.99 ± 0.02 a |
| Insoluble fiber (%) | 2.61 ± 0.21 a | 2.35 ± 0.18 a | 2.45 ± 0.15 a | 2.60 ± 0.19 a |
| Total dietary fiber (%) | 3.39 ± 0.11 a | 3.37 ± 0.10 a | 3.32 ± 0.12 a | 3.59 ± 0.21 a |
| Peak N° | RT (min) | m/z [M−H]− | Error (ppm) | Formula | Fragments (Relative Abundance, %) | Tentative Identification a |
|---|---|---|---|---|---|---|
| SIC-YF mixture before inoculation | ||||||
| 1 | 8.559 | 487.1438 | −2.9 | C21H27O13 | 487.1438 (100), 443.1901 (81.09), 193.0495 (28.89), 178.0260 (2.29) | Feruloyl derivative |
| 2 | 8.696 | 487.1437 | −3.1 | C21H27O13 | 487.1437 (100), 193.0495(50.77), 443.1900 (4554), 178.0261 (7.00) | Feruloyl derivative |
| 3 | 9.405 | 353.0876 | 0.8 | C16H17O9 | 191.0557 (100), 353.0873 (20.97), 192.0591 (8.35), 293.1240 (5.54), 248.9849 (3.06) | 5-O-caffeoylquinic acid (chlorogenic acid) |
| 4 | 10.469 | 353.0873 | 0.0 | C16H17O9 | 173.0451 (100), 353.0873 (19.45), 307.1228 (12.19), 281.1389 (8.60), 347.1173 (4.66), 237.1493 (4.26) | 4-O-caffeoylquinic acid (cryptochlorogenic acid) |
| 5 | 10.686 | 179.0347 | 1.7 | C9H7O4 | 135.0451 (13.51) | Caffeic acid b |
| 6 | 15.342 | 533.0928 | −0.6 | C24H21O14 | 337.1494 (24.72), 343.1181 (2.08), 163.0391 (0.01) | p-Coumaroyl diglucoside |
| 7 | 16.234 | 957.2886 | 7.2 | C49H49O20 | 957.2886 (100), 193.0502 (14.03), 939.2772 (11.64), 487.1453 (11.33), 469.1348 (5.87), 940.2817 (5.24) | Ellagitannin |
| 8 | 18.785 | 559.1100 | 2.1 | C26H23O14 | 559.1100 (100), 161.0241 (12.33), 397.0772 (10.31), 235.0454 (10.0), 179.0347 (2.87), 203.0345 (1.52) | 4,5-di-O-caffeoyl-2,7-anhydro-d-glycero-beta-d-galacto-oct-2-ulopyranosonic acid |
| 9 | 19.289 | 593.1514 | 1.3 | C27H29O15 | 593.1514 (100), 285.0399 (42.83), 286.0433 (4.28), | Kaempferol-3-O-rutinoside |
| 10 | 20.284 | 577.1563 | 1.0 | C27H29O14 | 271.0607 (100), 269.0452 (88.59), 577.1563 (57.17), 173.0448 (53.73), 270.0485 (12.31), 335.0763 (10.94) | Rhoifolin (Apigenin-7-O-neohesperidoside) |
| 11 | 20.284 | 515.1198 | 1.6 | C25H23O12 | 515.1198 (100), 353.0862 (39.41), 179.0339 (29.58), 173.0446 (28.06), 191.0550 (20.75), 209.0291 (16.72), 335.0760 (10.70) | 3,4-di-O-caffeoylquinic acid |
| 12 | 20.352 | 579.1716 | 0.3 | C27H31O14 | 579.1716 (100), 271.0606 (95.89), 269.0451 (41.80), 577.1559 (31.88), 270.0485 (5.21), 151.0031 (1.10) | Naringin b |
| 13 | 20.49 | 515.1186 | −0.8 | C25H23O12 | 353.0867 (100), 191.0553 (30.05), 354.0899 (17.20), 375.0683 (14.21), 349.2007 (10.61) | Chlorogenic acid derivative |
| 14 | 20.776 | 607.1667 | 0.7 | C28H31O15 | 299.0554 (100), 607.1667 (79.93), 284.0318 (26.87), 503.2489 (18.58), 300.0587 (17.21), 183.0655 (2.63) | Diosmin |
| 15 | 21.027 | 609.1824 | 0.8 | C28H33O15 | 609.1824 (100), 301.0714 (88.36), 302.0747 (88.36), 303.0766 (2.37), 286.0482 (2.11), 325.0716 (1.77), 164.0112 (0.42) | Hesperidin b |
| 16 | 22.938 | 593.1881 | 1.9 | C28H33O14 | 285.0761 (100), 593.1881 (80), 286.0792 (17.46), 309.0761 (0.73), 287.0819 (0.28), 164.0110 (0.12) | Poncirin |
| Treatment T1 after 28 days of storage | ||||||
| 2 | 8.679 | 487.1453 | 0.5 | 487.1453 (100), 443.1917 (75.17), 193.0502 (61.34), 178.0268 (7.40) | Feruloyl derivative | |
| 3 | 9.405 | 353.0871 | −0.6 | C16H17O9 | 191.0555 (100), 353.0869 (27.69), 293.1235 (11.93), 192.0588 (8.80), 179.0343 (3.43) | 5-O-caffeoylquinic acid (chlorogenic acid) |
| 4 | 10.469 | 353.0878 | 1.4 | C16H17O9 | 173.0448 (100), 281.1381 (4.85), 307.1217 (4.45), 237.1485 (0.89) | 4-O-caffeoylquinic acid (cryptochlorogenic acid) |
| 5 | 10.721 | 179.0343 | −0.6 | C9H7O4 | 135.0446 (26.21) | Caffeic acid |
| 6 | 15.273 | 533.0931 | 0.6 | C24H21O14 | 533.0934 (100), 337.1500 (59.98), 399.2725 (29.28), 444.2467 (9.28), 163.0394 (8.63) | p-Coumaroyl diglucoside |
| 7 | 16.206 | 957.2873 | 5.8 | C49H49O20 | 957.2873 (100), 939.2767 (18.62), 193.0500 (9.52), 940.2794 (8.23), 487.1453 (6.84), 469.1342 (5.98), 899.2450 (4.70), 663.1923 (3.41) | Ellagitannin |
| 8 | 18.785 | 559.1091 | 0.5 | C26H23O14 | 559.1091 (100), 235.0453 (7.30), 161.0240 (5.94), 397.0769 (2.68), 293.0659 (1.08), 203.0343 (0.68), 236.0491 (0.51) | 4,5-di-O-caffeoyl-2,7-anhydro-d-glycero-beta-d-galacto-oct-2-ulopyranosonic acid |
| 9 | 19.243 | 593.1502 | −0.7 | C27H29O15 | 593.1502 (100), 285.0399 (45.33), 286.0437 (3.37), 401.1810 (2.55) | Kaempferol-3-O-rutinoside |
| 10 | 20.284 | 577.1564 | 1.2 | C27H30O14 | 271.0605 (100), 579.1713 (85.85), 269.0450 (76.66), 577.1559 (49.89), 580.1744 (22.64), 173.0449 (33.09) | Rhoifolin (Apigenin-7-O-neohesperidoside) |
| 20.284 | 515.1192 | 0.4 | C25H23O12 | 515.1190 (100), 173.0450 (21.69), 353.0872 (6.89), 335.0764 (6.67) | 3,4-di-O-dicaffeoylquinic acid | |
| 20.352 | 579.1716 | 0.3 | C27H31O14 | 271.0605 (100), 579.1716 (98.32), 269.0449 (40.20), 577.1560 (30.75), 559.1088 (23.21), 151.0031 (2.07) | Naringin | |
| 13 | 20.472 | 723.5016 | −4.2 | 353.0858 (100), 191.0548 (46.07), 354.0894 (18.93), 375.0676 (18.25), 569.0366 (12.06), 677.7020 (9.89) | Chlorogenic acid derivative | |
| 14 | 20.776 | 607.1642 | −3.5 | C28H31O15 | 299.0546 (100), 607.1641 (76.94), 284.0314 (26.91), 503.2477 (21.51), 300.0580 (18.50), 285.0354 (4.01) | Diosmin |
| 15 | 21.062 | 609.1808 | −1.8 | C28H33O15 | 301.0704 (100), 609.1802 (95.25), 325.0704 (3.59), 242.0574 (2.30), 164.0107 (0.91) | Hesperidin |
| 16 | 22.937 | 593.1876 | 1.0 | C28H33O14 | 285.0765 (100), 593.1876 (81.80), 286.0799 (17.47) | Poncirin |
| Treatment T2 after 28 days of storage | ||||||
| 1 | 8.559 | 702.3229 | 702.3229 (99.27), 675.3952 (32.26), 593.3419 (30.28), 487.1458 (14.37), 616.2614 (8.5), 466.1944 (0.18) | Unknown phenolic metabolite; absence of characteristic feruloyl fragment (m/z 193.0499) | ||
| 2 | 8.662 | 487.1458 | 487.1458 (100), 193.0503 (61.97), 357.2504 (35.62), 443.1924 (20.40), 178.0267 (7.57), 214.1922 (4.11) | Feruloyl derivative | ||
| 3 | 9.302 | 353.0873 | −0.3 | C16H17O9 | 191.0557 (100), 353.0872 (27.13), 192.0561 (9.01), 203.0823 (6.50) | 5-O-caffeoylquinic acid (chlorogenic acid) b |
| 4 | 10.515 | 353.0870 | −0.8 | C16H17O9 | 173.0449 (12.95), 353.0870 (4.86), 257.1612 (4.12), 361.0957 (2.71), 470.3339 (2.30) | 4-O-caffeoylquinic acid (cryptochlorogenic acid) |
| 5 | 10.652 | 179.0341 | −1.7 | C9H7O4 | 179.0341 (100), 135.0443 (30.16) | Caffeic acid b |
| 6 | 15.273 | 614.3306 | 614.3306 (100), 570.3616 (23.31), 444.2458 (18.92), 414.2356 (8.03), 213.1237 (6.45), 533.0938 (5.84) | Unidentified phenolic compound; fragmentation suggests presence of galloyl group (loss 170 Da) | ||
| 7 | 16.120 | 787.4675 | 787.4675 (100), 743.4410 (23.13), 474.2386 (11.99), 744.4439 (8.90), 745.4454 (2.07) | Unidentified high-molecular-weight phenolic metabolite; fragmentation not consistent with ellagitannins | ||
| 8 | 18.819 | 559.1084 | −0.7 | C26H23O14 | 559.1084 (100), 161.0237 (7.90), 235.0449 (6.76), 397.0762 (6.60), 494.2028 (1.80), 362.2075 (1.36) | 4,5-di-O-caffeoyl-2,7-anhydro-d-glycero-beta-d-galacto-oct-2-ulopyranosonic acid |
| 9 | 19.208 | 593.3406 | 593.3420 (100), 493.2658 (57.62), 462.2346 (46.80), 484.2391 (24.21), 540.2452 (19.07), 285.0395 (1.0) | Kaempferol-3-O-rutinoside | ||
| 11 | 20.284 | 515.1196 | 1.2 | C25H23O12 | 515.1194 (89.43), 413.2765 (40.34), 444.2249 (27.17), 173.0450 (12.97), 353.0875 (8.01), 335.0773 (4.65) | 3,4-di-O-Dicaffeoylquinic acid |
| 12 | 20.352 | 1175.694 | 1175.6935 (100), 579.1725 (17.37), 271.0612 (15.52), 559.1102 (12.24), 677.4984 (8.35), 833.4530 (5.15) | Putative naringin-based conjugate; MS/MS fragments at m/z 579 (naringin) and 271 (naringenin) | ||
| 13 | 20.490 | 801.3547 | 801.3542 (100), 717.3466 (22.30), 353.0879 (14.15), 617.4036 (13.53), 655.3162 (7.12), 191.0559 (3.01) | Chlorogenic acid derivative | ||
| 14 | 20.81 | 607.1675 | 2.0 | C28H31O15 | 486.2932 (100), 299.0558 (16.37), 284.0322 (4.50), 298.2491 (4.32), 456.2822 (4.23) | Diosmin |
| 15 | 21.061 | 609.1816 | −0.5 | C28H33O15 | 609.1813 (100), 301.0708 (92.10), 302.0743 (16.96), 303.0763 (2.35), 286.0477 (1.95), 325.0710 (1.48) | Hesperidin |
| 16 | 22.937 | 593.1873 | 0.5 | C28H33O14 | 285.0759 (100), 593.1865 (81.36), 286.0791 (17.32), 309.0755 (0.52), 327.0871 (0.31) | Poncirin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Campos, D.; Chirinos, R.; Aguilar-Galvez, A.; Carrasco, M.P.; Pedreschi, R. Storage Stability of a Multifunctional Fermented Blend Based on Sacha Inchi (Plukenetia volubilis) Oil Press Cake and Yacon (Smallanthus sonchifolius) Flour: Physicochemical Properties, Bioactivity, and Prebiotic–Probiotic Potential. Foods 2026, 15, 2131. https://doi.org/10.3390/foods15122131
Campos D, Chirinos R, Aguilar-Galvez A, Carrasco MP, Pedreschi R. Storage Stability of a Multifunctional Fermented Blend Based on Sacha Inchi (Plukenetia volubilis) Oil Press Cake and Yacon (Smallanthus sonchifolius) Flour: Physicochemical Properties, Bioactivity, and Prebiotic–Probiotic Potential. Foods. 2026; 15(12):2131. https://doi.org/10.3390/foods15122131
Chicago/Turabian StyleCampos, David, Rosana Chirinos, Ana Aguilar-Galvez, María P. Carrasco, and Romina Pedreschi. 2026. "Storage Stability of a Multifunctional Fermented Blend Based on Sacha Inchi (Plukenetia volubilis) Oil Press Cake and Yacon (Smallanthus sonchifolius) Flour: Physicochemical Properties, Bioactivity, and Prebiotic–Probiotic Potential" Foods 15, no. 12: 2131. https://doi.org/10.3390/foods15122131
APA StyleCampos, D., Chirinos, R., Aguilar-Galvez, A., Carrasco, M. P., & Pedreschi, R. (2026). Storage Stability of a Multifunctional Fermented Blend Based on Sacha Inchi (Plukenetia volubilis) Oil Press Cake and Yacon (Smallanthus sonchifolius) Flour: Physicochemical Properties, Bioactivity, and Prebiotic–Probiotic Potential. Foods, 15(12), 2131. https://doi.org/10.3390/foods15122131