
Application and Possible Mechanism of Microbial Fermentation and Enzyme Catalysis in Regulation of Food Flavour

 ,
,  ,
,  , , and
, , and
Abstract
1. Introduction
2. Application of Microbial Fermentation in Regulation of Food Flavor
2.1. Applications of Different Microorganisms
2.1.1. Bacteria
2.1.2. Yeast
2.1.3. Other Fungi
2.1.4. Mixed Fermentation
2.2. Possible Regulatory Mechanisms of Microorganisms on Food Flavor
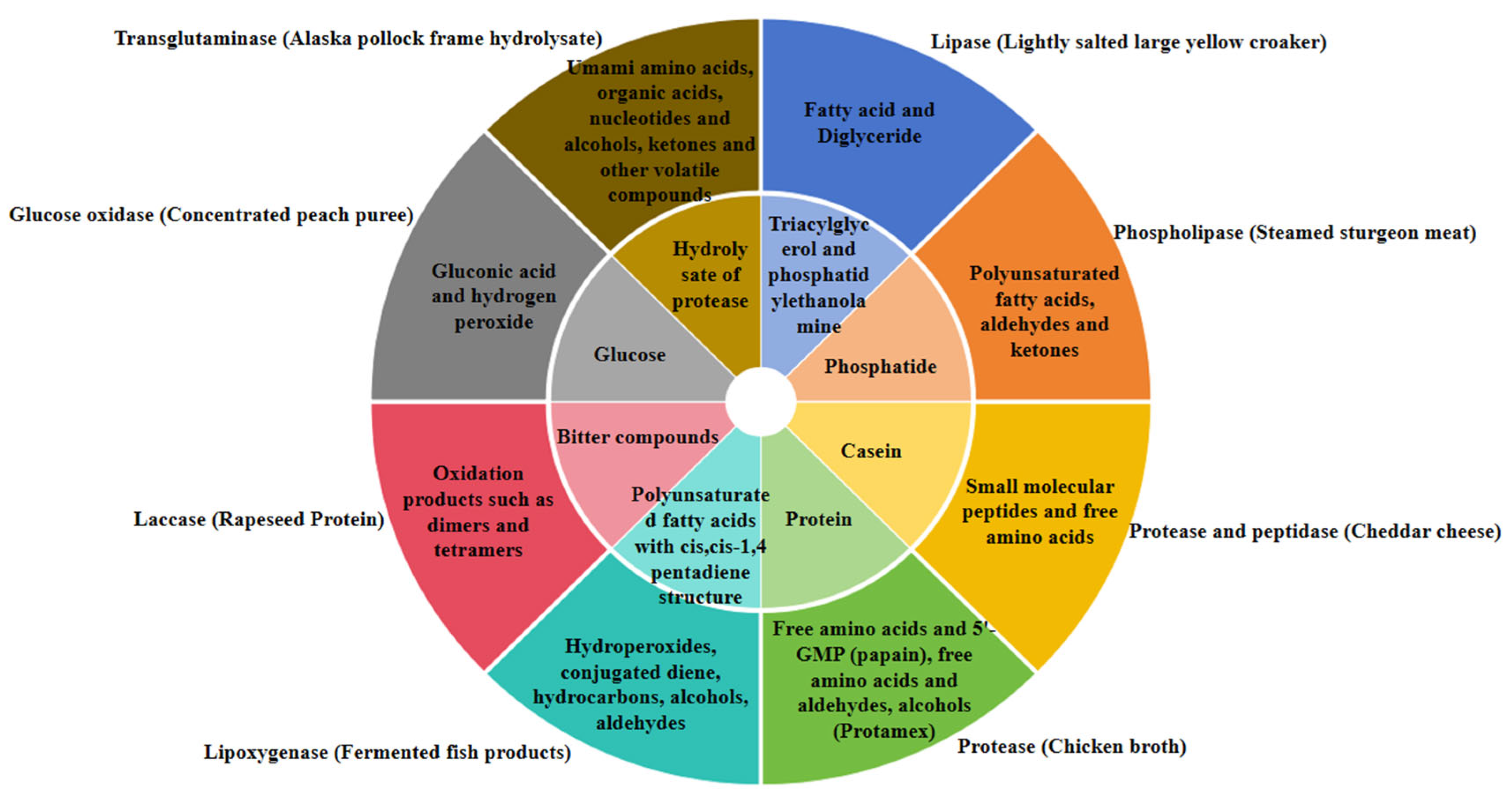
3. Application of Enzyme Catalysis in Regulation of Food Flavor
3.1. Application of Different Enzymes
3.1.1. Oxidoreductases (EC 1)
3.1.2. Transferases (EC 2)
3.1.3. Hydrolases (EC 3)
3.2. Enzyme Catalytic Reaction Schemes
3.2.1. Possible Catalytic Reaction Schemes by Oxidoreductases (EC 1)
3.2.2. Possible Catalytic Reaction Schemes by Transferases (EC 2)
3.2.3. Possible Catalytic Reaction Schemes by Hydrolases (EC 3)
4. Conclusions
- (1)
- Microorganisms are essential for the improvement of food flavor through fermentation. Therefore, the isolation and identification of flavor-producing microbial strains from traditional fermented food can be a potential way to enrich the microbes used in food. The construction of engineered strains for the recombinant expression of flavor-producing enzymes in food-safe microorganisms is also an alternative method.
- (2)
- Since the compounds in fermented food are complex, the identification of those compounds essential for food flavor is beneficial for quality control during food fermentation. The relationship between fermentation conditions and food flavor should also be explored in order to optimize standard protocol in food factories.
- (3)
- The enzymatic transformation of food to improve flavor is highly efficient. However, some flavor compounds are still extracted from plants or chemically synthesized. It is necessary to explore or design novel enzymes for the production of new natural flavor agents or low-cost and environmentally friendly synthesized flavors.
- (4)
- For the industrial application of enzymes in food flavor regulation, it is important to investigate food-safe, low-cost immobilization supports with good catalytic performance. The design of enzymatic reactors for food flavor improvement should also be explored.
- (5)
- Since an increasing number of flavor compounds produced by microorganisms and enzymes have been identified and their functions revealed, a database of flavor compounds, functions, producers, and transformers should be established; moreover, new flavor compounds could be designed with the assistance of AI, providing sufficient learning and training has been conducted. Further, potential pathways for the synthesis of new flavor compounds using microbial and/or enzymatic methods may also be identified by AI models based on the database.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tahir, H.E.; Zou, X.B.; Huang, X.W.; Shi, J.Y.; Mariod, A.A. Discrimination of honeys using colorimetric sensor arrays, sensory analysis and gas chromatography techniques. Food Chem. 2016, 206, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, T.-T.; Guo, R.-R.; Ye, Q.; Zhao, H.-L.; Huang, X.-H. The regulation of key flavor of traditional fermented food by microbial metabolism: A review. Food Chem. X 2023, 19, 100871. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.G.; Jiao, T.H.; Adade, S.; Zhen, W.; Qin, O.Y.; Chen, Q.S. Digital twin for predicting and controlling food fermentation: A case study of kombucha fermentation. J. Food Eng. 2025, 393, 112467. [Google Scholar] [CrossRef]
- Aregbe, A.Y.; Mubeen, B.; Xiong, Y.Q.; Ma, Y.K. Fermentation with Lactobacillus strains, Acetobacter pasteurianus, and Torulaspora delbrueckii D1-3 improves nutritional quality and volatile profile of sea buckthorn-based cereal beverage. Food Res. Int. 2025, 201, 115547. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, T.; Li, H.; Chen, T.; Liu, X. Control of beany flavor from soybean protein raw material in plant-based meat analog processing. Foods 2023, 12, 923. [Google Scholar] [CrossRef]
- Chiba, H.; Takahashi, N.; Sasaki, R. Enzymatic improvement of food flavor II. Removal of beany flavor from soybean products by aldehyde dehydrogenase. Agric. Biol. Chem. 1979, 43, 1883–1889. [Google Scholar]
- Redrejo-Rodriguez, M.; Tejeda-Cano, A.; del Carmen Pinto, M.; Macías, P. Lipoxygenase inhibition by flavonoids: Semiempirical study of the structure–activity relation. J. Mol. Struct. THEOCHEM 2004, 674, 121–124. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, G.; Zhu, G. The imitation and creation of a mango flavor. Food Sci. Technol. 2022, 42, e34622. [Google Scholar] [CrossRef]
- Yu, G.; Zhu, G.; Zheng, X. A hami melon flavor creation. Food Sci. Technol. 2022, 42, e95221. [Google Scholar] [CrossRef]
- Tchabo, W.; Ma, Y.K.; Kwaw, E.; Zhang, H.N.; Xiao, L.L.; Tahir, H.E. Aroma profile and sensory characteristics of a sulfur dioxide-free mulberry (Morus nigra) wine subjected to non-thermal accelerating aging techniques. Food Chem. 2017, 232, 89–97. [Google Scholar] [CrossRef]
- Gao, X.L.; Zhang, J.K.; Liu, E.M.; Yang, M.Q.; Chen, S.; Hu, F.; Ma, H.L.; Liu, Z.; Yu, X.T. Enhancing the taste of raw soy sauce using low intensity ultrasound treatment during moromi fermentation. Food Chem. 2019, 298, 124928. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.L.; Liu, E.M.; Zhang, J.K.; Yang, M.Q.; Chen, S.; Liu, Z.; Ma, H.L.; Hu, F. Effects of sonication during moromi fermentation on antioxidant activities of compounds in raw soy sauce. Lwt-Food Sci. Technol. 2019, 116, 108605. [Google Scholar] [CrossRef]
- Wang, X.W.; Lan, Y.B.; Yang, B.; Yang, J.; Ma, J.J.; Cheng, M.; Xia, X.D.; Xu, W.M.; Wang, D.Y.; Zou, Y. Effect of low-frequency ultrasound pretreatment on taste substances in chicken liver by fermentation. Lwt-Food Sci. Technol. 2025, 218, 117458. [Google Scholar] [CrossRef]
- Zheng, H.; Li, L.; Huang, C.; Liu, S.; Chen, X.; Wang, X.; Hu, P. Evaluation of ultrasound-assisted tomato sour soup marination on beef: Insights into physicochemical, sensory, microstructural, and flavour characteristics. Ultrason. Sonochemistry 2024, 110, 107028. [Google Scholar] [CrossRef]
- Zeng, X.; Li, Y.; Li, P.; Zhao, J.; Li, X.; Wang, X.; Liu, B.; Ni, L.; Li, H.; Xi, Y.; et al. Encapsulation of roast beef flavor by soy protein isolate/chitosan complex Pickering emulsions to improve its releasing properties during the processing of plant-based meat analogues. Food Chem. 2024, 450, 139313. [Google Scholar] [CrossRef]
- Rocha, R.A.R.; Ribeiro, M.N.; Silva, G.A.; Rocha, L.C.R.; Pinheiro, A.C.M.; Nunes, C.A.; Carneiro, J.d.D.S. Temporal profile of flavor enhancers MAG, MSG, GMP, and IMP, and their ability to enhance salty taste, in different reductions of sodium chloride. J. Food Sci. 2020, 85, 1565–1575. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Liu, M.T.; Zhao, Y.S.; Zhu, Y.; Bai, J.; Fan, S.T.; Zhu, L.; Song, C.; Xiao, X. Recent Developments in Fermented Cereals on Nutritional Constituents and Potential Health Benefits. Foods 2022, 11, 2243. [Google Scholar] [CrossRef]
- Chen, Q.S.; Hu, W.W.; Su, J.; Li, H.H.; Ouyang, Q.; Zhao, J.W. Nondestructively sensing of total viable count (TVC) in chicken using an artificial olfaction system based colorimetric sensor array. J. Food Eng. 2016, 168, 259–266. [Google Scholar] [CrossRef]
- Liu, S.; Zhao, L.; Li, M.; Zhu, Y.; Liang, D.; Ma, Y.; Sun, L.; Zhao, G.; Tu, Q. Probiotic Bacillus as fermentation agents: Status, potential insights, and future perspectives. Food Chem. X 2024, 22, 101465. [Google Scholar] [CrossRef]
- Ye, Q.; Tan, J.; He, X.; Wang, C.; Liu, X.; Li, C.; Fang, Z.; Chen, H.; Liu, Y.; Chen, S. Effect of lipase and lipoxygenase on lipid metabolism and the formation of main volatile flavour compounds in fermented fish products: A review. Int. J. Food Sci. Technol. 2024, 59, 1248–1259. [Google Scholar] [CrossRef]
- Gu, Y.G.; Bai, J.; Zhang, J.Y.; Zhao, Y.S.; Pan, R.R.; Dong, Y.; Cui, H.L.; Xiao, X. Transcriptomics reveals the anti-obesity mechanism of Lactobacillus plantarum fermented barley extract. Food Res. Int. 2022, 157, 111285. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.S.; Lu, F.; Liu, D.D.; Zhao, X.X.; Chao, J.P.; Wang, Y.C.; Luan, Y.; Ma, H.L. The process of solid-state fermentation of soybean meal: Antimicrobial activity, fermentation heat generation and nitrogen solubility index. J. Sci. Food Agric. 2024, 104, 3228–3234. [Google Scholar] [CrossRef] [PubMed]
- Kwaw, E.; Ma, Y.K.; Tchabo, W.; Apaliya, M.T.; Xiao, L.L.; Wu, M. Effect of lactic acid fermentation on the phytochemical, volatile profile and sensory attributes of mulberry juice. J. Food Nutr. Res. 2017, 56, 305–317. [Google Scholar]
- Zhang, L.; Yu, X.J.; Yagoub, A.E.; Xia, G.H.; Zhou, C.S. Effect of vacuum impregnation assisted probiotics fermentation suspension on shelf life quality of freshly cut lotus root. Food Chem. 2022, 381, 132281. [Google Scholar] [CrossRef]
- Chen, C.; Zhao, S.; Hao, G.; Yu, H.; Tian, H.; Zhao, G. Role of lactic acid bacteria on the yogurt flavour: A review. Int. J. Food Prop. 2017, 20, S316–S330. [Google Scholar] [CrossRef]
- Murtaza, M.A.; Ur-Rehman, S.; Anjum, F.M.; Huma, N.; Hafiz, I. Cheddar Cheese Ripening and Flavor Characterization: A Review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1309–1321. [Google Scholar] [CrossRef]
- Yin, X.; Huang, S.; Xiong, T.; Peng, F. Effects of Lactobacillus plantarum NCU1546 fermentation on active components, in vitro uric acid lowering activity and flavor of Lilium. Food Biosci. 2025, 65. [Google Scholar] [CrossRef]
- Bai, J.; Pan, B.B.; Luo, W.; Yang, Z.H.; Zhu, L.; Cheng, Z.C.; Zhao, Y.S.; Zhang, J.Y.; Zhu, Y.; Xiao, X. Role of sulfatase LPMS from Lactiplantibacillus plantarum dy-1 in releasing bound phenolic acids of barley bran dietary fiber. J. Food Sci. 2025, 90, e70062. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Xiao, X.; Dong, Y.; Xu, T.; Wu, F. Dietary supplementation with Lactobacillus plantarum dy-1 fermented barley suppresses body weight gain in high-fat diet-induced obese rats. J. Sci. Food Agric. 2016, 96, 4907–4917. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Xiao, X.; Dong, Y.; Shi, L.N.; Xu, T.; Wu, F. The anti-obesity effect of fermented barley extracts with Lactobacillus plantarum dy-1 and Saccharomyces cerevisiae in diet-induced obese rats. Food Funct. 2017, 8, 1132–1143. [Google Scholar] [CrossRef]
- Huang, J.; Wang, L.; Li, X.; Zhang, Y.; Du, T.; Xiong, S.; Yuan, L.; Xiong, T.; Liu, G. Fermentation of asparagus juice using selected probiotic Lactiplantibacillus plantarum JGS49 isolated from pickled asparagus: Microbiological, physicochemical and flavour characteristics. Int. J. Food Sci. Technol. 2024, 59, 7113–7124. [Google Scholar] [CrossRef]
- Liu, Z.; Xiong, S.; Hardie, W.J.; Huang, X.; Liu, Q.; Yang, J.; Li, J.; Liu, X.; Xiao, M.; Xiong, T.; et al. Transcriptomics and metabolomics reveal effects of Lactiplantibacillus plantarum NCU116 fermentation on carrot flavor. Food Biosci. 2025, 64, 105898. [Google Scholar] [CrossRef]
- Wang, S.; Li, C.; Xu, Q.; Wang, Y.; Wang, S.; Zou, Y.; Yang, Z.; Yuan, L. Addition of lactic acid bacteria modulates microbial community and promotes the flavor profiles of Kombucha. Food Biosci. 2024, 60, 104340. [Google Scholar] [CrossRef]
- Xia, X.D.; Dai, Y.Q.; Wu, H.; Liu, X.L.; Wang, Y.; Yin, L.Q.; Wang, Z.; Li, X.N.; Zhou, J.Z. Kombucha fermentation enhances the health-promoting properties of soymilk beverage. J. Funct. Foods 2019, 62, 103549. [Google Scholar] [CrossRef]
- Khazi, M.I.; Liaqat, F.; Liu, X.R.; Yan, Y.L.; Zhu, D.C. Fermentation, functional analysis, and biological activities of turmeric kombucha. J. Sci. Food Agric. 2024, 104, 759–768. [Google Scholar] [CrossRef]
- Ding, Y.; Yang, Q.; Wang, M.; He, W.; Dai, W.; Tang, X.; Zheng, R.; Zhou, X. Flavor and nutritional characteristics of surimi product with Lactiplantibacillus plantarum as a reinforcing starter culture. Food Biosci. 2024, 62, 105229. [Google Scholar] [CrossRef]
- Xu, B.; Mi, T.; Ma, S.; Yi, X.; Huang, P.; Huang, P.; Wu, C. Insight into the autochthonous lactic acid bacteria as starter culture for improving the quality of Sichuan radish paocai: Changes in microbial diversity and metabolic profiles. Int. J. Food Microbiol. 2024, 425, 110877. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhou, Q.; Li, D.; Wu, Y.; Zhong, K.; Gao, H. Effects of enhanced fermentation with Lactiplantibacillus plantarum WWPC on physicochemical characteristics and flavor profiles of radish paocai and dried-fermented radish. Food Biosci. 2024, 59, 103941. [Google Scholar] [CrossRef]
- Wei, W.; Yang, S.J.; Yang, F.; Hu, X.Y.; Wang, Y.; Guo, W.J.; Yang, B.Y.; Xiao, X.; Zhu, L. Cold Plasma Controls Nitrite Hazards by Modulating Microbial Communities in Pickled Radish. Foods 2023, 12, 2550. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, X.; Xin, L.; Xu, S.; Meng, X. Effect of rapid fermentation on the quality of northeastern sauerkraut analyzed based on HSSPME-GC-MS and LC-MS. Lwt-Food Sci. Technol. 2024, 198, 116005. [Google Scholar] [CrossRef]
- Chen, Z.; Gao, H.; Wu, W.; Chen, H.; Fang, X.; Han, Y.; Mu, H. Effects of fermentation with different microbial species on the umami taste of Shiitake mushroom (Lentinus edodes). Lwt-Food Sci. Technol. 2021, 141, 110889. [Google Scholar] [CrossRef]
- Chen, Z.; Fang, X.; Wu, W.; Chen, H.; Han, Y.; Yang, H.; Gao, H. Effects of fermentation with Lactiplantibacillus plantarum GDM1.191 on the umami compounds in shiitake mushrooms (Lentinus edodes). Food Chem. 2021, 364, 130398. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.R.; Ding, X.N.; Dai, C.H.; Ma, H.L. Changes in the structure and dissociation of soybean protein isolate induced by ultrasound-assisted acid pretreatment. Food Chem. 2017, 232, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Yi, M.; Sun, J.; Zhang, T.; Wen, R.; Li, C.; Reshetnik, E.I.; Gribanova, S.L.; Liu, L.; Zhang, G. Physicochemical properties and volatile profile of mung bean flour fermented by Lacticaseibacillus casei and Lactococcus lactis. Lwt-Food Sci. Technol. 2022, 163, 113565. [Google Scholar] [CrossRef]
- Meng, J.; Wang, J.-L.; Hao, Y.-P.; Zhu, M.-X.; Wang, J. Effects of Lactobacillus fermentum GD01 fermentation on the nutritional components and flavor substances of three kinds of bean milk. Lwt-Food Sci. Technol. 2023, 184, 115006. [Google Scholar] [CrossRef]
- Shangguan, L.; Liu, Z.; Xu, L.; Yang, Q.; Zhang, X.; Yao, L.; Li, P.; Chen, X.; Dai, J. Effect of Corynebacterium glutamicum Fermentation on the Volatile Flavors of the Enzymatic Hydrolysate of Soybean Protein Isolate. Foods 2024, 13, 2591. [Google Scholar] [CrossRef]
- Spaccasassi, A.; Utz, F.; Dunkel, A.; Borner, R.A.; Ye, L.; De Franceschi, F.; Bogicevic, B.; Glabasnia, A.; Hofmann, T.; Dawid, C. Screening of a Microbial Culture Collection: Empowering Selection of Starters for Enhanced Sensory Attributes of Pea-Protein-Based Beverages. J. Agric. Food Chem. 2024, 72, 15890–15905. [Google Scholar] [CrossRef]
- Xue, Y.; Chen, J.; Wang, L.; Wang, Y.; Xu, F. Exploring the flavor changes in mung bean flour through Lactobacillus fermentation: Insights from volatile compounds and non-targeted metabolomics analysis. J. Sci. Food Agric. 2024, 104, 7238–7248. [Google Scholar] [CrossRef]
- Yi, C.; Li, Y.; Zhu, H.; Liu, Y.; Quan, K. Effect of Lactobacillus plantarum fermentation on the volatile flavors of mung beans. Lwt-Food Sci. Technol. 2021, 146, 111434. [Google Scholar] [CrossRef]
- Qian, J.Y.; Zhou, C.S.; Ma, H.L.; Li, S.J.; Yagoub, A.A.; Abdualrahman, M.A.Y. Proteomics Analyses and Morphological Structure of Bacillus subtilis Inactivated by Pulsed Magnetic Field. Food Biophys. 2016, 11, 436–445. [Google Scholar] [CrossRef]
- Qian, J.Y.; Zhou, C.S.; Ma, H.L.; Li, S.J.; Yagoub, A.A.; Abdualrahman, M.A.Y. Biological Effect and Inactivation Mechanism of Bacillus subtilis Exposed to Pulsed Magnetic Field: Morphology, Membrane Permeability and Intracellular Contents. Food Biophys. 2016, 11, 429–435. [Google Scholar] [CrossRef]
- Dai, C.H.; Ma, H.L.; He, R.H.; Huang, L.R.; Zhu, S.Y.; Ding, Q.Z.; Luo, L. Improvement of nutritional value and bioactivity of soybean meal by solid-state fermentation with Bacillus subtilis. Lwt-Food Sci. Technol. 2017, 86, 1–7. [Google Scholar] [CrossRef]
- Tuly, J.A.; Ma, H.L.; Zabed, H.M.; Dong, Y.T.; Chen, G.; Guo, L.A.; Betchem, G.; Igbokwe, C.J. Exploring magnetic field treatment into solid-state fermentation of organic waste for improving structural and physiological properties of keratin peptides. Food Biosci. 2022, 49, 101872. [Google Scholar] [CrossRef]
- Hou, X.S.; Dai, C.H.; Tang, Y.X.; Xing, Z.; Mintah, B.K.; Dabbour, M.; Ding, Q.Z.; He, R.H.; Ma, H.L. Thermophilic solid-state fermentation of rapeseed meal and analysis of microbial community diversity. Lwt-Food Sci. Technol. 2019, 116, 108520. [Google Scholar] [CrossRef]
- Xing, Z.; Jiang, H.; He, R.H.; Mintah, B.K.; Dabbour, M.; Dai, C.H.; Sun, L.; Ma, H.L. Rapid detection model of Bacillus subtilis in solid-state fermentation of rapeseed meal. J. Food Saf. 2020, 40, e12754. [Google Scholar] [CrossRef]
- Zou, Y.; Zhong, Y.; Zhou, Q.; Jia, Z.; Chen, Q.; Xu, W.; Wu, Y.; Wei, S.; Zhong, K.; Gao, H. Effects of solid-state fermentation with Bacillus subtilis Y4 on the quality of Yibin Yacai. Food Biosci. 2023, 51, 102258. [Google Scholar] [CrossRef]
- Guo, Q.; Zabed, H.; Zhang, H.H.; Wang, X.; Yun, J.H.; Zhang, G.Y.; Yang, M.M.; Sun, W.J.; Qi, X.H. Optimization of fermentation medium for a newly isolated yeast strain (Zygosaccharomyces rouxii JM-C46) and evaluation of factors affecting biosynthesis of D-arabitol. Lwt-Food Sci. Technol. 2019, 99, 319–327. [Google Scholar] [CrossRef]
- Ma, J.; Ma, Y.H.; Zhang, H.Z.; Chen, Z.L.; Wen, B.; Wang, Y.; Huang, W.Y. The quality change of fig wine fermented by RV171 yeast during the six-month aging process. Lwt-Food Sci. Technol. 2022, 166, 113789. [Google Scholar] [CrossRef]
- Ma, D.; Yuan, L.; Mao, J.; Liu, T.; Zhao, Y.; Han, X.; Ji, Z.; Liu, S.; Mao, J. Optimizing Huangjiu fermentation for enhanced aroma: Insights into Saccharomyces cerevisiae jiangnan1# strain. J. Food Compos. Anal. 2025, 139, 107051. [Google Scholar] [CrossRef]
- Ouyang, Q.; Zhao, J.W.; Pan, W.X.; Chen, Q.S. Real-time monitoring of process parameters in rice wine fermentation by a portable spectral analytical system combined with multivariate analysis. Food Chem. 2016, 190, 135–141. [Google Scholar] [CrossRef]
- Li, Q.; Du, B.; Chen, X.; Zhao, Y.; Zhu, L.; Ma, H.; Sun, B.; Hao, J.; Li, X. Microbial community dynamics and spatial distribution of flavor compound metabolism during solid-state fermentation of Baijiu enhanced by Wickerhamomyces anomalus. Food Biosci. 2024, 59, 103909. [Google Scholar] [CrossRef]
- Bian, M.; Fang, Y.; Yang, S.; Yuan, T.; Zhou, X.; Xu, Q.; Jiang, D.; Han, B. Dynamic Changes in Physicochemical Properties, Amino Acid Content, and Flavour-Related Substances During Fortified Fermentation of Rice Wine with Saccharomycopsis fibuligera. J. Food Process. Preserv. 2024, 2024, 5581068. [Google Scholar] [CrossRef]
- Che, X.; Li, B.; Zeng, J.; Chen, Y.; Liu, J.; Zeng, W.; Cai, W.; Wan, Y.; Fu, G. Effect of co-fermentation of non-Saccharomyces yeasts with Saccharomyces cerevisiae on the quality and flavor of blueberry wine. Food Res. Int. 2024, 196, 115051. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Xiong, L.; Huang, Z.; Huang, R.; Tu, T.; Yu, S.; Wu, Q.; Bai, J.; Huang, Y. Enhancing the quality and flavor of pomelo wine through sequential fermentation with Schizosaccharomyces pombe and Saccharomyces cerevisiae: A non-targeted metabolomic analysis. Food Biosci. 2025, 63, 105585. [Google Scholar] [CrossRef]
- Liang, J.; Yuan, H.; Fei, Y.; Wang, H.; Qu, C.; Bai, W.; Liu, G. Effects of Saccharomyces cerevisiae and Cyberlindnera fabianii Inoculation on Rice-Flavor Baijiu Fermentation. Foods 2024, 13, 3175. [Google Scholar] [CrossRef]
- Liu, S.; Wang, R.; Zhang, Y.; Dai, Y.; Zhang, S.; Lin, X.; Ji, C.; Liang, H.; Chen, Y. Effect of co-fermentation of Rhodotorula mucilaginosa with Saccharomyces cerevisiae on the quality and flavor profile of cider. Food Biosci. 2025, 64, 105973. [Google Scholar] [CrossRef]
- Gao, M.; Hu, J.; Wang, X.; Zhang, H.; Du, Z.; Ma, L.; Du, L.; Zhang, H.; Tian, X.; Yang, W. Effects of Pichia kluyveri on the flavor characteristics of wine by co-fermentation with Saccharomyces cerevisiae. Eur. Food Res. Technol. 2023, 249, 1449–1460. [Google Scholar] [CrossRef]
- Fu, B.; Wang, J.; Xiao, T.; Liu, Z.; Fu, C.; Zhou, M.; Liu, Z.; Li, W.; Hu, Y.; Wang, C.; et al. Co-culture relationship between Zygosaccharomyces rouxii and Candida versatilis and its effect on the flavour of soy sauce. Int. J. Food Sci. Technol. 2024, 59, 228–240. [Google Scholar] [CrossRef]
- Song, L.; Ma, F.; Chen, H.; Fei, Q.; Tao, G.; Wu, S.; Shi, D.; Deng, J.; Zhao, D.; Dong, X.; et al. Dynamic changes in flavor characteristics of black tea during solid-state fermentation with Eurotium cristatum. Food Chem. 2025, 465, 142028. [Google Scholar] [CrossRef]
- Yang, X.; Liu, Z.; Zhang, Y.; Zhao, S.; Yan, S.; Zhu, L.; Zhou, Q.; Chen, L. Effects of Fermentation with Eurotium cristatum on Sensory Properties and Flavor Compounds of Mulberry Leaf Tea. Foods 2024, 13, 2347. [Google Scholar] [CrossRef]
- Zhang, W.; Deng, Z.; Liu, T.; Liang, J.; Liu, J. Fermentation with edible mushroom mycelia improves flavor characteristics and techno-functionalities of soybean protein. Food Biosci. 2024, 59, 104123. [Google Scholar] [CrossRef]
- Wang, Z.; Gao, T.; He, Z.; Zeng, M.; Qin, F.; Chen, J. Reduction of off-flavor volatile compounds in okara by fermentation with four edible fungi. Lwt-Food Sci. Technol. 2022, 155, 112941. [Google Scholar] [CrossRef]
- Yu, J.; Yang, L.; Ma, Z.; Yu, G.; Xue, Y.; Xue, C. Exploring the effect of mold fermentation on the taste and flavor properties of traditional Japanese smoked-dried bonito (katsuobushi). Food Biosci. 2023, 56, 103366. [Google Scholar] [CrossRef]
- Huang, W.; Xu, H.N.; Pan, J.Y.; Dai, C.H.; Mintah, B.K.; Dabbour, M.; Zhou, R.; He, R.H.; Ma, H.L. Mixed-Strain Fermentation Conditions Screening of Polypeptides from Rapeseed Meal and the Microbial Diversity Analysis by High-Throughput Sequencing. Foods 2022, 11, 3285. [Google Scholar] [CrossRef]
- Min, Y.; Zhang, Q.; Liu, J.; Shang, Y.; Hou, Y.; Zhang, M.; Dai, J.; Li, Z.; Xiang, W.; Tang, J. Effects of synthetic microbial community fermentation on volatile flavor and quality characteristics of ginger pickle. Food Res. Int. 2025, 207, 116077. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhao, Y.; Wang, Y.; Wu, Y.; Chen, S. Improvement of the quality and safety of low-salt fish sauce by reconstruction of microbial community through cooperative fermentation of starters. Food Res. Int. 2025, 205, 115972. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Wang, Q.; Yang, Y.; Wen, A.; Zeng, H.; Liu, N.; Qin, L. Effects of enhanced fermentation with high-yielding strains of Tetramethylpyrazine on flavor quality of Douchiba. Food Chem. X 2025, 25, 102037. [Google Scholar] [CrossRef]
- Fernandez-Varela, R.; Hansen, A.H.; Svendsen, B.A.; Moghadam, E.G.; Bas, A.; Kracun, S.K.; Harle, O.; Poulsen, V.K. Harnessing Fermentation by Bacillus and Lactic Acid Bacteria for Enhanced Texture, Flavor, and Nutritional Value in Plant-Based Matrices. Ferment. 2024, 10, 411. [Google Scholar] [CrossRef]
- Luo, Y.-Y.; Guo, Y.; Hu, X.-Y.; Liu, W.-H.; Liu, B.-Q.; Yang, J.; Tu, Z.-C.; Huang, Y.-H. Flavor improvement of fermented soybean foods by co-fermentation with Bacillus velezensis and Lactiplantibacillus plantarum. Lwt Food Sci. Technol. 2023, 186, 115257. [Google Scholar] [CrossRef]
- Wang, M.; Wang, C.; Yang, C.; Peng, L.; Xie, Q.; Zheng, R.; Dai, Y.; Liu, S.; Peng, X. Effects of Lactobacillus plantarum C7 and Staphylococcus warneri S6 on flavor quality and bacterial diversity of fermented meat rice, a traditional Chinese food. Food Res. Int. 2021, 150, 110745. [Google Scholar] [CrossRef]
- Wang, J.; Hou, J.; Zhang, X.; Hu, J.; Yu, Z.; Zhu, Y. Improving the Flavor of Fermented Sausage by Increasing Its Bacterial Quality via Inoculation with Lactobacillus plantarum MSZ2 and Staphylococcus xylosus YCC3. Foods 2022, 11, 736. [Google Scholar] [CrossRef] [PubMed]
- Shan, K.; Yao, Y.; Wang, J.; Zhou, T.; Zeng, X.; Zhang, M.; Ke, W.; He, H.; Li, C. Effect of probiotic Bacillus cereus DM423 on the flavor formation of fermented sausage. Food Res. Int. 2023, 172, 113210. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-N.; Yao, Q.-B.; Zhang, H.-R.; Li, W.-Q.; Nie, J.; Liang, Q.-C.; Wang, L.-H.; Zeng, X.-A.; Huang, Y.-Y. Impact of different lactic acid bacteria on nitrite degradation and quality of fermented carrot. Int. J. Food Sci. Technol. 2024, 59, 6501–6512. [Google Scholar] [CrossRef]
- Peng, Q.; Li, J.; Fang, S.; Zhang, Y.; Zheng, H.; Meng, K.; Li, L.; Zhang, L.; Feng, X.; Chen, X.; et al. Enhancing the flavor quality of Chinese rice wine (Huangjiu) through inoculation with Lactobacillus plantarum and Leuconostoc mesenteroides. Lwt-Food Sci. Technol. 2024, 211, 116935. [Google Scholar] [CrossRef]
- Hu, Q.; Qi, Y.; Liu, C.; Chen, Q.; Mai, X.; Zhu, Z.; Ma, B. Effects of Lactobacillus plantarum cofermentation on the flavor and taste characteristics of mango vinegar. J. Food Meas. Charact. 2024, 18, 3744–3756. [Google Scholar] [CrossRef]
- Gu, J.; Li, S.; Shen, X.; Liang, Q.; Xu, T.; Shi, W. Effects of different fermenters on the quality and flavour of fermented mandarin fish (Siniperca chuatsi). Int. J. Food Sci. Technol. 2024, 59, 4992–5007. [Google Scholar] [CrossRef]
- Gao, R.C.; Zheng, Z.Y.; Zhou, J.; Tian, H.Y.; Yuan, L. Effects of mixed starter cultures and exogenous L-Lys on the physiochemical and sensory properties of rapid-fermented fish paste using longsnout catfish by-products. Lwt-Food Sci. Technol. 2019, 108, 21–30. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Zhao, F.; Shi, T.; Xiong, Z.Y.; Gao, R.C.; Yuan, L. Suanyu fermentation strains screening, process optimization and the effect of thermal processing methods on its flavor. Food Res. Int. 2023, 173, 113296. [Google Scholar] [CrossRef]
- Yuan, L.; Zhang, Q.Q.; Zhao, F.; Jin, W.G.; Shi, T.; Xiong, Z.Y.; Gao, R.C. Dynamic changes and correlation analysis of protease activities, texture and flavour compounds during fish fermentation (suanyu) using mixed culture (Enterococcus rivorum and Enterococcus lactis). Int. J. Food Sci. Technol. 2023, 58, 2312–2324. [Google Scholar] [CrossRef]
- Du, Y.; Shim, S.-M.; Wang, L.; Gao, X.; Fu, X. Impact of Monascus purpureus combined with Lactobacillus plantarum and Saccharomyces cerevisiae fermentation on nutritional and flavor characteristics of Pyropia yezoensis. Food Chem. 2025, 472, 142973. [Google Scholar] [CrossRef]
- Peng, Q.; Zhou, H.; Zheng, H.; Xie, G. Investigating the role of primary fungi in Huangjiu fermentation: Insights from flavor orientation and synthetic microbiomes. Food Microbiol. 2025, 129, 104765. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, S.; Liu, B.; Feng, S. Flavor characteristics of hulless barley wine fermented with mixed starters by molds and yeasts isolated from Jiuqu. Food Biosci. 2023, 52, 102349. [Google Scholar] [CrossRef]
- Cao, C.; Song, X.; Mu, Y.; Sun, W.; Su, G. Co-culturing Limosilactobacillus fermentum and Pichia fermentans to ferment soybean protein hydrolysates: An effective flavor enhancement strategy. Innov. Food Sci. Emerg. Technol. 2025, 100, 103920. [Google Scholar] [CrossRef]
- Peng, X.; Liao, Y.; Ren, K.; Liu, Y.; Wang, M.; Yu, A.; Tian, T.; Liao, P.; Huang, Z.; Wang, H.; et al. Fermentation performance, nutrient composition, and flavor volatiles in soy milk after mixed culture fermentation. Process Biochem. 2022, 121, 286–297. [Google Scholar] [CrossRef]
- Qiu, Y.; Ye, S.; Huang, X.; Li, C.; Feng, Y.; Xia, M.; Yu, Q.; Zhang, X.; Wang, Q. Fermentation with a multi-strain to enhance the flavor of HongJun Tofu, a Chinese fermented okara food. Lwt-Food Sci. Technol. 2023, 189, 115495. [Google Scholar] [CrossRef]
- Wu, W.; Chen, T.; Zhao, M.; Feng, Y. Effect of co-inoculation of different halophilic bacteria and yeast on the flavor of fermented soy sauce. Food Biosci. 2023, 51, 102292. [Google Scholar] [CrossRef]
- Gao, X.L.; Liu, E.M.; Zhang, J.K.; Yang, L.X.; Huang, Q.R.; Chen, S.; Ma, H.L.; Ho, C.T.; Liao, L. Accelerating aroma formation of raw soy sauce using low intensity sonication. Food Chem. 2020, 329, 127118. [Google Scholar] [CrossRef]
- Gao, P.; Zhang, Z.; Jiang, Q.; Hu, X.; Zhang, X.; Yu, P.; Yang, F.; Liu, S.; Xia, W. Metabolomics ravels flavor compound formation and metabolite transformation in rapid fermentation of salt-free fish sauce from catfish frames induced by mixed microbial cultures. Food Chem. 2025, 463, 141246. [Google Scholar] [CrossRef]
- Fan, P.; Liang, X.; Fei, Y.; Zhao, W.; Liang, J.; Bai, W.; He, S. Effect of Mixed Strains on Microbial Community and Flavor Metabolites in Fermentation Process of Chi-Flavor Baijiu. Foods 2024, 13, 3497. [Google Scholar] [CrossRef]
- Tian, S.; Li, Y.; Du, G.; Liu, Q. Effect of cultures of synthetic starters on the flavour and ethyl carbamate content of rice wine. Int. J. Food Sci. Technol. 2024, 59, 5868–5876. [Google Scholar] [CrossRef]
- Zhao, X.; Xue, Y.; Tang, F.; Cai, W.; Hao, G.; Shan, C. Quality improvement of jujube wine through mixed fermentation with Saccharomyces cerevisiae and Bacillus licheniformis. Lwt Food Sci. Technol. 2022, 164, 113444. [Google Scholar] [CrossRef]
- Wang, J.; Yan, C.; Ma, C.; Huang, S.; Chang, X.; Li, Z.; Chen, X.; Li, X. Effects of two kinds of Bacillus on flavour formation of Baijiu solid-state fermentation with pure mixed bacteria. Int. J. Food Sci. Technol. 2023, 58, 1250–1262. [Google Scholar] [CrossRef]
- Fang, L.; Wang, W.; Dou, Z.; Chen, J.; Meng, Y.; Cai, L.; Li, Y. Effects of mixed fermentation of different lactic acid bacteria and yeast on phytic acid degradation and flavor compounds in sourdough. Lwt Food Sci. Technol. 2023, 174, 114438. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, F.; Ananta, E.; Muller, J.A.; Liang, Y.; Lee, Y.K.; Liu, S.-Q. Co-Inoculation of Latilactobacillus sakei with Pichia kluyveri or Saccharomyces boulardii Improves Flavour Compound Profiles of Salt-Free Fermented Wheat Gluten. Fermentation 2024, 10, 100492. [Google Scholar] [CrossRef]
- Wang, L.; Deng, K.W.; Zhang, Y. Isolation and screening of high-quality lactic acid bacteria and yeast strains in kefir grains and preparation of kefir compound fermentation starter. J. Food Process. Preserv. 2022, 46, e17073. [Google Scholar] [CrossRef]
- Li, Z.; Su, W.; Mu, Y.; Qi, Q.; Jiang, L. Effects of Pediococcus acidilactici and Rhizopus Oryzae on protein degradation and flavor formation in fermented mutton sausages. Lwt-Food Sci. Technol. 2024, 213, 117075. [Google Scholar] [CrossRef]
- Gong, Z.; Wu, Z.; Yang, Q.; Liu, J.; Jiao, P.; Tang, C. Influences of lactic acid bacteria strains on the flavor profiles, metabolites and quality characteristics of red yeast rice produced by solid-state fermentation. Food Res. Int. 2024, 197, 115172. [Google Scholar] [CrossRef]
- Li, X.; He, Y.; Xie, Y.; Zhang, L.; Li, J.; Liu, H. Effects of fermentation with Kefir grains on nutrient composition, flavor volatiles, and product physical stability of a hemp seed (Cannabis sativa L.) beverage. Lwt Food Sci. Technol. 2023, 183, 114934. [Google Scholar] [CrossRef]
- Rabelo, M.H.S.; Borem, F.M.; Alves, A.P.d.C.; Pieroni, R.S.; Santos, C.M.; Nakajima, M.; Sugino, R. Fermentation of coffee fruit with sequential inoculation of Lactiplantibacillus plantarum and Saccharomyces cerevisiae: Effects on volatile composition and sensory characteristics. Food Chem. 2024, 444, 138608. [Google Scholar] [CrossRef]
- Wang, L.; Zhong, H.; Liu, K.Y.; Guo, A.Z.; Qi, X.H.; Cai, M.H. The evaluation of kefir pure culture starter: Liquid-core capsule entrapping microorganisms isolated from kefir grains. Food Sci. Technol. Int. 2016, 22, 598–608. [Google Scholar] [CrossRef]
- Chen, X.J.; Zhou, Y.J.; Zhang, D. Engineering Corynebacterium crenatum for enhancing succinic acid production. J. Food Biochem. 2018, 42, 12645. [Google Scholar] [CrossRef]
- Liang, J.; Wang, Y.; Wang, T.; Chu, C.; Yi, J.; Liu, Z. Enhancing fermented vegetable flavor with Lactobacillus plantarum and Rhodotorula mucilaginosa. Food Res. Int. 2025, 200, 115500. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Jomaa, S.A.; Abd El-Wahed, A.; El-Seedi, H.R. The Many Faces of Kefir Fermented Dairy Products: Quality Characteristics, Flavour Chemistry, Nutritional Value, Health Benefits, and Safety. Nutrients 2020, 12, 346. [Google Scholar] [CrossRef]
- Jia, J.; Duan, J.; Bao, S.; Zhang, X.; Jia, X.; Ye, J.; Liu, Y.; Liu, X.; Duan, X. Metabolomic and proteomic profiling reveals the formation mechanism of volatile flavor in egg whites during fermentation by Streptococcus thermophilus. Food Chem. 2025, 466, 142219. [Google Scholar] [CrossRef]
- Zeng, Z.; Song, C.; Hu, X.; Zhu, X.; Li, Y.; Ren, J.; Wang, Y.; Yang, H.; Huang, X.; Zhao, M.; et al. Constituent-taste relationship of Kuding tea fermented by Aspergillus neoniger and Aspergillus cristatus: Unveiling taste characteristics through untargeted metabolomics. Food Biosci. 2024, 62, 105027. [Google Scholar] [CrossRef]
- Wang, F.; Zhu, L.; Hadiatullah, H.; Li, Z.; He, J.; Yao, Y.; Zhao, G. Enhancing flavour with non-Saccharomyces during vinegar alcoholisation: A mechanism study. Int. J. Food Sci. Technol. 2023, 58, 5001–5008. [Google Scholar] [CrossRef]
- Li, C.; Chen, S.; Huang, H.; Li, J.; Zhao, Y. Improvement mechanism of volatile flavor in fermented tilapia surimi by cooperative fermentation of Pediococcus acidilactici and Latilactobacillus sakei: Quantization of microbial contribution through influence of genus. Food Chem. 2024, 449, 139239. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Lu, Y.; Li, X.; Huang, J.; Dong, L.; Luo, J.; Tian, F.; Ni, Y. Omics analysis of key pathway in flavour formation and B vitamins synthesis during chickpea milk fermentation by Lactiplantibacillus plantarum. Food Chem. 2025, 463, 141083. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Tang, P.; Bai, J.; Zhou, Y.; Shen, T.; Wu, Z.; Li, X.; Tong, P.; Chen, H.; Yang, A. Anti-food allergic activity of soymilk fermented by Lactobacillus in vitro. J. Food Sci. Technol. Mysore 2024, 1–12. [Google Scholar] [CrossRef]
- Khatami, S.H.; Vakili, O.; Ahmadi, N.; Soltani Fard, E.; Mousavi, P.; Khalvati, B.; Maleksabet, A.; Savardashtaki, A.; Taheri-Anganeh, M.; Movahedpour, A. Glucose oxidase: Applications, sources, and recombinant production. Biotechnol. Appl. Biochem. 2022, 69, 939–950. [Google Scholar] [CrossRef]
- Luo, D.; Pan, X.; Zhang, W.; Bi, S.; Wu, J. Effect of glucose oxidase treatment on the aroma qualities and release of cooked off-odor components from heat-treated Hami melon juice. Food Chem. 2022, 371, 131166. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Chen, Q.; Gou, M.; Bi, J. The potential of glucosidase and glucose oxidase for aroma improvement in concentrated peach puree based on volatilomics and metabolomics. Food Chem. 2024, 450, 139375. [Google Scholar] [CrossRef] [PubMed]
- Botezatu, A.; Essary, A.; Bajec, M. Glucose Oxidase in Conjunction with Catalase—An Effective System of Wine pH Management in Red Wine. Am. J. Enol. Vitic. 2023, 74, 0740004. [Google Scholar] [CrossRef]
- Li, J.; Fu, J.; Ma, Y.; He, Y.; Fu, R.; Qayum, A.; Jiang, Z.; Wang, L. Low temperature extrusion promotes transglutaminase cross-linking of whey protein isolate and enhances its emulsifying properties and water holding capacity. Food Hydrocoll. 2022, 125, 107410. [Google Scholar] [CrossRef]
- Shan, C.; Wu, G.; Bu, Y.; Zhu, W.; Li, J.; Li, X. Effect of transglutaminase cross-linking on debittering of Alaska pollock frame hydrolysate. Int. J. Food Sci. Technol. 2024, 59, 438–449. [Google Scholar] [CrossRef]
- Zhang, Q.; Cheng, Z.; Wang, Y.; Zheng, S.; Wang, Y.; Fu, L. Combining Alcalase hydrolysis and transglutaminase-cross-linking improved bitterness and techno-functional properties of hypoallergenic soybean protein hydrolysates through structural modifications. Lwt Food Sci. Technol. 2021, 151, 112096. [Google Scholar] [CrossRef]
- Li, J.; Cui, H.; Xu, X.; Li, J.; Lu, M.; Yu, Y.; Song, H.; Zhu, D.; Liu, H. Effects of pectic fat mimetics and transglutaminase on the regularity of protein and fat degradation and flavour compounds in Cheddar cheese during ripening. Int. J. Food Sci. Technol. 2022, 57, 1291–1302. [Google Scholar] [CrossRef]
- Li, B.; Zhao, L.N.; Chen, H.J.; Sun, D.W.; Deng, B.X.; Li, J.W.; Liu, Y.F.; Wang, F. Inactivation of Lipase and Lipoxygenase of Wheat Germ with Temperature-Controlled Short Wave Infrared Radiation and Its Effect on Storage Stability and Quality of Wheat Germ Oil. PLoS ONE 2016, 11, e0167330. [Google Scholar] [CrossRef]
- Jin, J.; Ma, H.L.; Wang, W.W.; Luo, M.; Wang, B.; Qu, W.J.; He, R.H.; Owusu, J.; Li, Y.L. Effects and mechanism of ultrasound pretreatment on rapeseed protein enzymolysis. J. Sci. Food Agric. 2016, 96, 1159–1166. [Google Scholar] [CrossRef]
- Tang, S.W.; Dong, S.Y.; Chen, M.; Gao, R.C.; Chen, S.J.; Zhao, Y.H.; Liu, Z.Y.; Sun, B.W. Preparation of a fermentation solution of grass fish bones and its calcium bioavailability in rats. Food Funct. 2018, 9, 4135–4142. [Google Scholar] [CrossRef]
- Cui, H.Y.; Li, W.; Li, C.Z.; Lin, L. Intelligent release of cinnamon oil from engineered proteoliposome via stimulation of Bacillus cereus protease. Food Control 2016, 67, 68–74. [Google Scholar] [CrossRef]
- Gao, R.C.; Shi, T.; Liu, X.D.; Zhao, M.Q.; Cui, H.L.; Yuan, L. Purification and characterisation of a salt-stable protease fromthe halophilic archaeon Halogranum rubrum. J. Sci. Food Agric. 2017, 97, 1412–1419. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xu, J.; Liu, Q.; Xia, X.; Sun, F.; Kong, B. Effect of the protease from Staphylococcus carnosus on the proteolysis, quality characteristics, and flavor development of Harbin dry sausage. Meat Sci. 2022, 189, 108827. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Wang, H.; Liu, Q.; Xia, X.; Chen, Q.; Kong, B. Proteolysis and quality characteristics of Harbin dry sausages caused by the addition of Staphylococcus xylosus protease. Food Chem. 2023, 404, 134692. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Pei, Y.; Cheng, K.; Deng, Y.; Dong, X.; Fang, R.; Chu, B.; Wei, P.; Chen, Q.; Xiao, G. Production and evaluation of enzyme-modified cheese adding protease or lipase to improve quality properties. J. Biosci. Bioeng. 2023, 135, 389–394. [Google Scholar] [CrossRef]
- Su, L.; Yang, W.; Liu, S.; Yuan, C.; Huang, T.; Jia, R.; Wei, H. Effect of Neutral Protease on Freshness Quality of Shucked Pacific Oysters at Different Storage Conditions. Foods 2024, 13, 1273. [Google Scholar] [CrossRef]
- Weng, Z.; Sun, L.; Wang, F.; Sui, X.; Fang, Y.; Tang, X.; Shen, X. Assessment the flavor of soybean meal hydrolyzed with Alcalase enzyme under different hydrolysis conditions by E-nose, E-tongue and HS-SPME-GC-MS. Food Chem. -X 2021, 12, 100141. [Google Scholar] [CrossRef]
- Purbaningrum, K.; Hidayat, C.; Witasari, L.D.; Utami, T. Flavor Precursors and Volatile Compounds Improvement of Unfermented Cocoa Beans by Hydrolysis Using Bromelain. Foods 2023, 12, 820. [Google Scholar] [CrossRef]
- De la Hoz, A.; Caripan, P.; Moltedo, B.; Ferrada, N.; Contreras, R.A. A metabolomic approach of AI-driven enzymatic digestion of pumpkin seed flour for producing umami metabolites. Int. J. Gastron. Food Sci. 2025, 39, 101117. [Google Scholar] [CrossRef]
- Xu, D.; Gu, M.; Xi, J.; Chen, L.; Jin, Y.; Wu, F.; Xu, X.; Zhao, Q. Effect of a commercial peptidase on volatile composition and sensory properties of wheat bread crumb and crust. Lwt-Food Sci. Technol. 2023, 189, 115532. [Google Scholar] [CrossRef]
- Zhang, W.; Chan, J.X.; Lu, Y.; Liu, S.-Q. Pre-treatment of coconut kernels by proteases to modulate the flavour of coconut oil. Food Biosci. 2022, 48, 101736. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, J.; Zheng, X.; Zhang, Z.; Wang, Z.; Tan, X. Characterization of a Desiccation Stress Induced Lipase Gene from Brassica napus L. J. Agric. Sci. Technol. 2016, 18, 1129–1141. [Google Scholar]
- He, W.S.; Cu, D.D.; Zhang, Y.L.; Liu, Y.; Yin, J.; Chen, G.; Jia, C.S.; Feng, B. Highly Efficient Synthesis of Phytosterol Linolenate Catalyzed by Candida Rugosa Lipase through Transesterification. Food Sci. Technol. Res. 2017, 23, 525–533. [Google Scholar] [CrossRef]
- Chen, X.; Wang, J.; Stevenson, R.J.; Ang, X.; Peng, Y.; Quek, S.Y. Lipase-catalyzed modification of milk fat: A promising way to alter flavor notes of goat milk products. Lwt Food Sci. Technol. 2021, 145, 111286. [Google Scholar] [CrossRef]
- Guan, T.; Liu, B.; Wang, R.; Huang, Y.; Luo, J.; Li, Y. The enhanced fatty acids flavor release for low-fat cheeses by carrier immobilized lipases on O/W Pickering emulsions. Food Hydrocoll. 2021, 116, 106651. [Google Scholar] [CrossRef]
- Sun, R.; Chen, S.; Liu, Z.; Wu, J.; Su, L. Enzyme-Assisted Osmosis Process of Functional Sugar to Improve the Quality Characteristics of Cherry Tomatoes. Food Bioprocess Technol. 2024, 17, 3776–3789. [Google Scholar] [CrossRef]
- Jiang, G.; Li, B.; Ding, Z.; Zhu, J.; Li, S. Effect of cellulase on antioxidant activity and flavor of Rosa roxburghii Tratt. Food Chem. X 2024, 21, 101148. [Google Scholar] [CrossRef]
- Xiang, Y.; Chen, X.; Sun, H.; Zhan, Q.; Zhong, L.; Hu, Q.; Zhao, L. The critical roles of α-amylase and amyloglucosidase in improving the quality of black waxy corn beverages: Special attentions to the color and flavor. J. Cereal Sci. 2023, 110, 103625. [Google Scholar] [CrossRef]
- Nakamura, R.; Saito, M.; Maruyama, M.; Yamanaka, S.; Tanemura, T.; Watanabe, M.; Fukumori, F.; Hayashi, K. Reduction in the bitterness of protein hydrolysates by an aminopeptidase from Aspergillus oryzae. Food Sci. Technol. Res. 2023, 29, 71. [Google Scholar] [CrossRef]
- Sakai, K.; Okada, M.; Yamaguchi, S. Protein-glutaminase improves water-/oil-holding capacity and beany off-flavor profiles of plant-based meat analogs. PLoS ONE 2023, 18, e0294637. [Google Scholar] [CrossRef]
- Huang, W.R.; He, X.C.; Wu, J.C.; Ma, X.N.; Han, J.; Wang, L.; Wang, Y. The evaluation of deep eutectic solvents and ionic liquids as cosolvents system for improving cellulase properties. Ind. Crops Prod. 2023, 197, 116555. [Google Scholar] [CrossRef]
- Li, J.; Zhang, X.; Luo, S.; Wu, X.; Zhang, G.; Li, J.; Liu, X. Heterologous expression of protein-glutaminase in Escherichia coli Nissle 1917 for improved flavor properties of soy protein and its implications on high-moisture extrudates. Food Biosci. 2024, 57, 103591. [Google Scholar] [CrossRef]
- Zhao, J.; Ma, M.; Yan, X.; Zhang, G.; Xia, J.; Zeng, G.; Tian, W.; Bao, X.; Zeng, Z.; Yu, P.; et al. Expression and characterization of a novel lipase from Bacillus licheniformis NCU CS-5 for application in enhancing fatty acids flavor release for low-fat cheeses. Food Chem. 2022, 368, 130868. [Google Scholar] [CrossRef]
- Sakai, K.; Broches, N.; Okuda, K.; Okada, M.; Yamaguchi, S. Umami and saltiness enhancements of textured pea proteins by combining protease- and glutaminase-catalyzed reactions. Curr. Res. Food Sci. 2025, 10, 101022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, Y.; Gao, L.; Wang, J. Analysis of flavor changes in Huangshan floral mushroom hydrolysates obtained by different enzyme treatments. Food Chem. 2024, 443, 138554. [Google Scholar] [CrossRef]
- Tan, M.; Chen, C.; Cui, F.-J.; Ye, P.P.; Zhang, H.B.; Zhou, T.L.; Shi, J.-C.; Shu, X.-Q.; Chen, Z.-W. Flavor enhancement of strong fragrant rapeseed oils by enzymatic treatment. Ind. Crops Prod. 2024, 210, 118098. [Google Scholar] [CrossRef]
- Betchem, G.; Dabbour, M.; Tuly, J.A.; Billong, L.F.; Ma, H.L. Optimization of fermentation conditions to improve the functional and structural characteristics of rapeseed meal with a mutant Bacillus subtilis species. Ind. Crops Prod. 2023, 205, 117424. [Google Scholar] [CrossRef]
- Huang, R.; Zhang, F.; Zhou, H.; Yu, H.; Shen, L.; Jiang, J.; Qin, Y.; Liu, Y.; Song, Y. Characterization of Trichoderma reesei endoglucanase displayed on the Saccharomyces cerevisiae cell surface and its effect on wine flavor in combination with β-glucosidase. Process Biochem. 2023, 124, 140–149. [Google Scholar] [CrossRef]
- Qin, D.; Sheng, B.; Xu, S.; Ma, Q.; Xu, Z.; Liu, M.; Zhao, D. The deodorising and flavouring effect of enzymatic hydrolysis and glycation on boiled pig trotters. Czech J. Food Sci. 2024, 42, 447–455. [Google Scholar] [CrossRef]
- Li, C.; Tu, Z.; Liu, W.; Wu, C.; Hu, Y.; Wang, H. Flavor substances of low-valued red swamp crayfish (Procambarus clarkii) hydrolysates derived from double enzymatic systems. Food Res. Int. 2023, 165, 112461. [Google Scholar] [CrossRef]
- Xiao, N.; Zhang, Q.; Xu, H.; Zheng, C.; Yin, Y.; Liu, S.; Shi, W. Effect of Lactobacillus plantarum and flavourzyme on protein degradation and flavor development in grass carp during fermentation. Food Chem. X 2024, 22, 101439. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, J.; Li, L.; Wang, S.; Liu, Y.; Gao, M. Effect of enzymatic hydrolysis on volatile flavor compounds of Monascus -fermented tartary buckwheat based on headspace gas chromatography-ion mobility spectrometry. Food Res. Int. 2023, 163, 112180. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Z.; Lin, X.; Zhang, S.; Ji, C. Impact of Enzyme-Microbe Combined Fermentation on the Safety and Quality of Soy Paste Fermented with Grass Carp By-Products. Foods 2025, 14, 106. [Google Scholar] [CrossRef]
- Xiang, Q.; Xia, Y.; Chen, L.; Chen, M.; Wang, D.; Zhong, F. Flavor precursors and flavor compounds in Cheddar-flavored enzyme-modified cheese due to pre-enzymolysis combined with lactic acid bacteria fermentation. Food Biosci. 2023, 53, 102698. [Google Scholar] [CrossRef]
- Cui, H.Y.; Wu, J.; Li, C.Z.; Lin, L. Anti-listeria effects of chitosan-coated nisin-silica liposome on Cheddar cheese. J. Dairy Sci. 2016, 99, 8598–8606. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Zhang, B.; Han, Y.; Li, J.; Zhang, B.; Jiang, Y. Optimization of the quality of sea buckthorn juice by enzymatic digestion and inoculation sequence. Food Chem. 2025, 470, 142623. [Google Scholar] [CrossRef]
- Yan, Y.; Fan, X.; Hua, D.; Liu, X.; Li, C.; Li, Z.; Bai, M.; Zhang, Y.; Liu, H. Effects of different enzymatic hydrolysis techniques on volatile flavor compounds and nutritional metabolites of soybean meal yogurt. J. Sci. Food Agric. 2025, 105, 1312–1323. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, Z.; Wang, J.; Wang, X.-S.; Zhou, D.-Y.; Qin, L.; Huang, X.-H. Mechanism of differentiated and targeted catalysis in complex lipid system under lipase and lipoxygenase mediation. Food Chem. 2025, 469, 142503. [Google Scholar] [CrossRef]
- Spaccasassi, A.; Walser, C.; Nisov, A.; Sozer, N.; Frank, O.; Dawid, C.; Hofmann, T.F. Reducing the Bitterness of Rapeseed Protein: Integrating Enzymatic Treatment, Metabolomics, and Sensory Analysis to Elucidate Underlying Mechanisms. J. Agric. Food Chem. 2025, 73, 3657–3668. [Google Scholar] [CrossRef]
- Su, G.; Xie, Y.; Liu, R.; Cui, G.; Zhao, M.; Zhang, J. Effect of transglutaminase on taste characteristics of pea protein hydrolysates through altering the composition of amino acids and peptides. Food Biosci. 2023, 56, 103261. [Google Scholar] [CrossRef]
- Xia, X.; Fu, Y.; Ma, L.; Zhu, H.; Yu, Y.; Dai, H.; Han, J.; Liu, X.; Liu, Z.; Zhang, Y. Protein Hydrolysates from Pleurotus geesteranus Modified by Bacillus amyloliquefaciens γ-Glutamyl Transpeptidase Exhibit a Remarkable Taste-Enhancing Effect. J. Agric. Food Chem. 2022, 70, 12143–12155. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, X.-H.; Wang, X.; Tian, H.; Wang, L.; Zhou, D.-Y.; Qin, L. Compensatory effect of lipase on the flavor of lightly-salted large yellow croaker: Integration of flavoromics and lipidomics. Food Biosci. 2024, 59, 103907. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, Y.; Bai, F.; Wang, J.; Gao, R.; Zhao, Y.; Xu, X. Contribution of phospholipase B to the formation of characteristic flavor in steamed sturgeon meat. Food Chem. X 2024, 22, 101391. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Peng, S.; Guo, H.; Yuan, X.; Li, K.; Ma, L.; Chen, F.; Hu, X.; Liao, X.; Ji, J. The effect of enzymatic hydrolysis on the flavor improvement of chicken broth by characterizing key taste and odor-active compounds. Food Biosci. 2024, 62, 105054. [Google Scholar] [CrossRef]
- Fan, X.; Zhao, Y.; Mao, W.; Zhang, H.; Li, M.; Luo, Y.; Zhou, H.; Shen, Q.; Quan, W.; Luo, J. Preparation of a novel enzyme-modified cheddar cheese: Molecular mechanism of cheese flavor compensation by synergistic action of cell-free extracts and enzyme systems. Food Chem. 2025, 467, 142281. [Google Scholar] [CrossRef]
- Chen, Y.; Ma, L.; Liu, Y.; Huo, J.; Gao, Y.; Dong, S.; Li, S. Study on the effect of enzymolysis combined fermentation on reducing the off-flavor of egg white powder. J. Sci. Food Agric. 2024, 104, 7864–7872. [Google Scholar] [CrossRef]
- Wang, L.; Gao, E.Y.; Hu, M.; Oladejo, A.; Gong, X.F.; Wang, J.Y.; Zhong, H. Isolation, identification and screening of high-quality yeast strains for the production of milk beer. Int. J. Dairy Technol. 2018, 71, 944–953. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Microbial Type | Name | Processing Object | Flavor Substances | Odor | Reference |
|---|---|---|---|---|---|
| Bacteria | L. planturum | Asparagus juice | 1-Octen-3-ol, b-damascenone, 1-Octen-3-one, and dimethyl trisulfide | Enhanced mushroom and floral flavor Reduced green flavor | [74] |
| L. planturum | Surimi product | Ethyl octanoate | Produced a sweet and pleasant brandy aroma | [36] | |
| L. planturum | Dried-fermented radish | Lactic acid, oxalic acid, glutamic acid (Glu), ocimenol, and trans-4-thujanol | Produced fruit, floral, and grassy flavors | [38] | |
| L. planturum | Shiitake mushrooms | 5′-nucleotides, cysteine, histidine, citric acid, and tartaric acid | Enhanced umami | [42] | |
| B. subtilis | Yibin Yacai | β-Myrcene, Germacrene D, γ-Elemene, ethyl Cinnamate, ethyl p-methoxycinnamate, and sweet amino acids | Enhanced fruit, herb, green, oil, wood, and aroma flavors | [57] | |
| Fungi | S. cerevisiae | Huangjiu | Acetate esters | Enhanced aroma | [60] |
| W. anomalus | Baijiu | Ethyl acetate | Enhanced the aftertaste, reduced dryness and astringency | [62] | |
| E. cristatum | Black tea | β-damascenone, 2-ethyl-5-methylpyrazine, acetophenone, and tetrahydro-α, α, 5-trimethyl-5-vinylfuran-2-methanol | Enhanced floral, honey, sweet flavor Reduced bitterness and astringency | [70] | |
| Mold | Smoked-dried bonito | 3-Methyl-1-butanol, 2,5-dimethylpyrazine, and α-pinene | Produced malt, nutty, roasted aromas | [74] |
| Category | Microorganism | Processing Object | Flavor Substances | Odor | Reference |
|---|---|---|---|---|---|
| Bacterial mixed fermentation | B. velezensis and L. plantarum | Fermented soybean foods | Acetoin and pyrazines | Enhanced flavors | [73] |
| L. plantarum and Staphylococcus warneri | Fermented meat rice | Ethyl hexanoate, β-eudesmol, nerolidol, ethyl caproate, and citronellal | Enhanced flavors | [80] | |
| Leuconostoc mesenteroides, Weissella cibaria, and L. plantarum | Ginger pickle | Eucalyptol, α-terpinene, 1-hexanol, 2,4-di-tert-butylphenol, methyl geranate, and (−)-lavender acetate | Produced floral, sweet, and sour fragrance | [74] | |
| Tetragenococcus muriaticus, B. subtilis, and Staphylococcus edaphicus | Low-salt fish sauce | 1-heptanol, 1-nonanol, 1-octanol, ethyl acetate, and 2-pentylfuran | Enhanced floral, mushroom, fruity, and grassy flavors Reduced pungent, sulfur, and fishy flavors | [75] | |
| Fungal mixed fermentation | M. purpureus and S. cerevisiae | Pyropia yezoensis | Glu, ethanol, and ethyl acetate | Enhanced alcohol, fruity, and sweet aromas Reduced fishy, grass, and seawater odors | [90] |
| Rhodotorula mucilaginosa and S.cerevisiae | Cider | Isoamyl acetate and phenethyl acetate | Enhanced fruity and floral notes | [66] | |
| Candida glabrata and S. cerevisiae | Blueberry wine | Isoamylol, 2-phenylethanol, isoamyl acetate, ethyl acetate, and ethyl laurate | Enhanced sweetness, fruitiness, and floral flavors | [63] | |
| Mixed bacteria and fungi fermentation | L. fermentum and P. fermentans | Soybean protein hydrolysates | Umami amino acids, lactic acid, acetic acid, and malic acid | Enhanced saltiness, umami, and sweetness Reduced bitterness | [93] |
| N. crassa, L. plantarum and Meyerozyma guilliermondii | HongJun tofu | Glu, aspartic acid, γ-aminobutyric acid, and 5′-nucleotides | Enhanced fruity, mushroom, floral, and nutty flavors Reduced bitterness | [95] | |
| Pichia anomala and L. plantarum | Chi-flavor baijiu | Phenylethyl alcohol, (E)-2-octenal, and diethyl succinate | Enhanced flavors | [99] | |
| S. cerevisiae and B. licheniformis | Jujube wine | Isoamyl acetate and phenethyl acetate | Enhanced flavors | [101] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Wang, M.; Xu, L.; Qian, J.; Xu, B.; Gao, X.; Ding, Z.; Cui, K. Application and Possible Mechanism of Microbial Fermentation and Enzyme Catalysis in Regulation of Food Flavour. Foods 2025, 14, 1909. https://doi.org/10.3390/foods14111909
Wang F, Wang M, Xu L, Qian J, Xu B, Gao X, Ding Z, Cui K. Application and Possible Mechanism of Microbial Fermentation and Enzyme Catalysis in Regulation of Food Flavour. Foods. 2025; 14(11):1909. https://doi.org/10.3390/foods14111909
Chicago/Turabian StyleWang, Feng, Mingtong Wang, Ling Xu, Jingya Qian, Baoguo Xu, Xianli Gao, Zhongyang Ding, and Kai Cui. 2025. "Application and Possible Mechanism of Microbial Fermentation and Enzyme Catalysis in Regulation of Food Flavour" Foods 14, no. 11: 1909. https://doi.org/10.3390/foods14111909
APA StyleWang, F., Wang, M., Xu, L., Qian, J., Xu, B., Gao, X., Ding, Z., & Cui, K. (2025). Application and Possible Mechanism of Microbial Fermentation and Enzyme Catalysis in Regulation of Food Flavour. Foods, 14(11), 1909. https://doi.org/10.3390/foods14111909