
Seasonal Variations in the Composition and Physicochemical Characteristics of Sheep and Goat Milks



Abstract
:1. Introduction
2. Materials and Methods
2.1. Milk Sampling
2.2. Macronutrient and Mineral Analysis
2.3. Fatty Acid Composition and Melting Profile of Milk Fat
2.4. Protein Composition
2.5. Physicochemical Properties and Heat-Induced Changes
2.6. Statistical Analysis
3. Results and Discussion
3.1. Compositional and Physicochemical Properties of Sheep and Goat Milks
3.1.1. Composition of Macronutrients and Minerals
3.1.2. Fatty Acid Composition and Melting Properties of Milk Fat
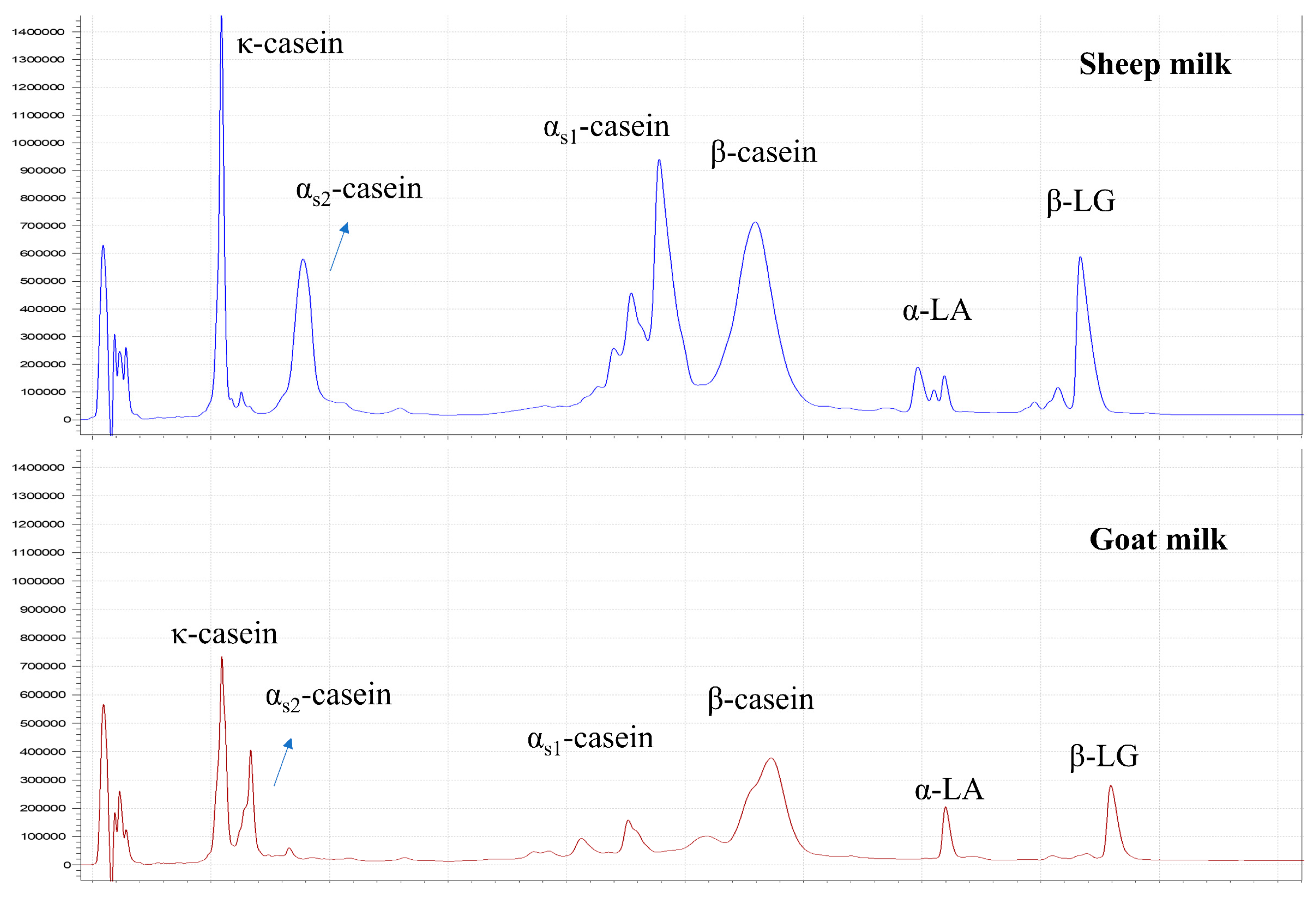
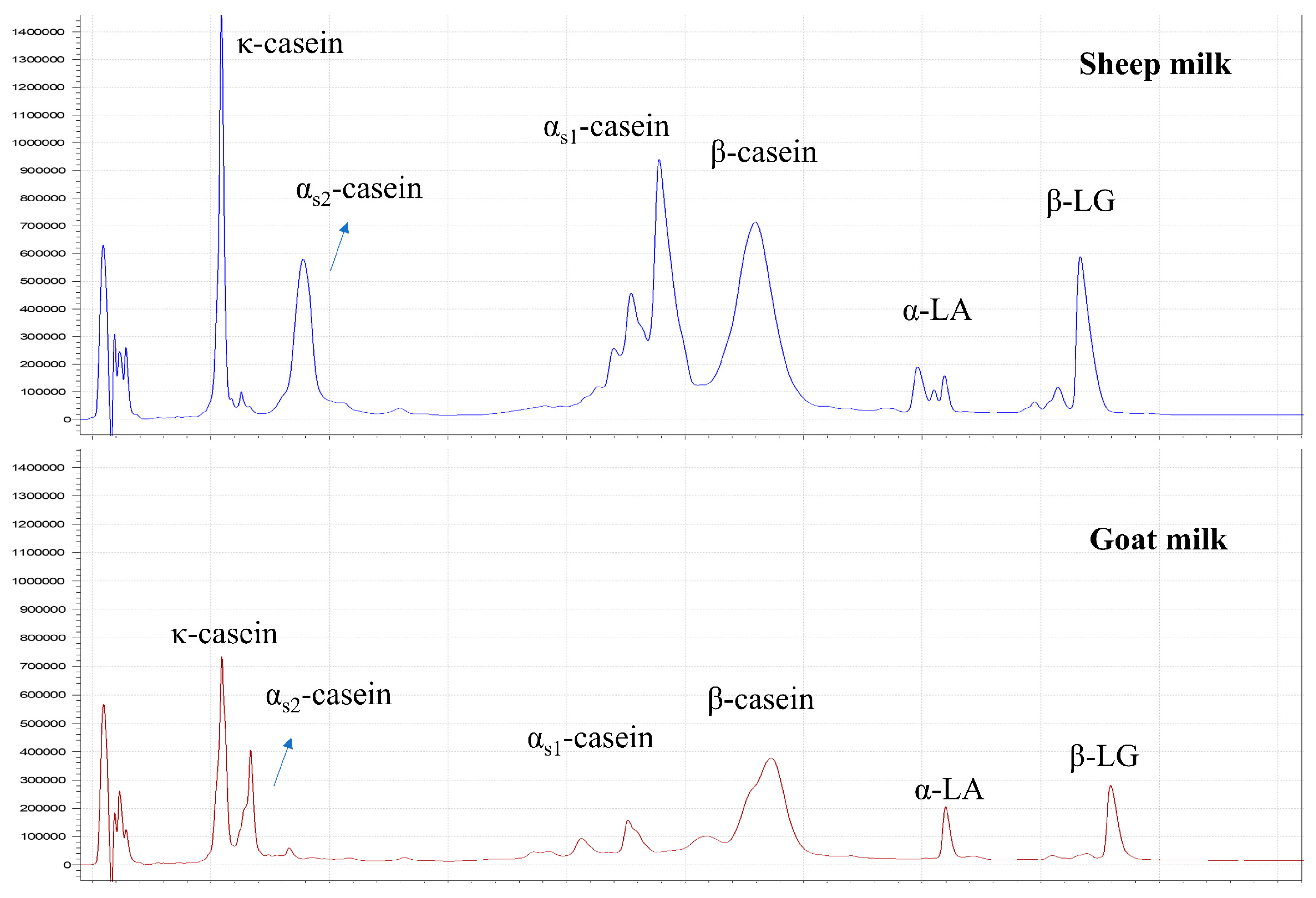
3.1.3. Protein Composition
3.1.4. Physicochemical Properties
3.2. Behaviour of Proteins and Physical Changes in Milk upon Heat Treatment
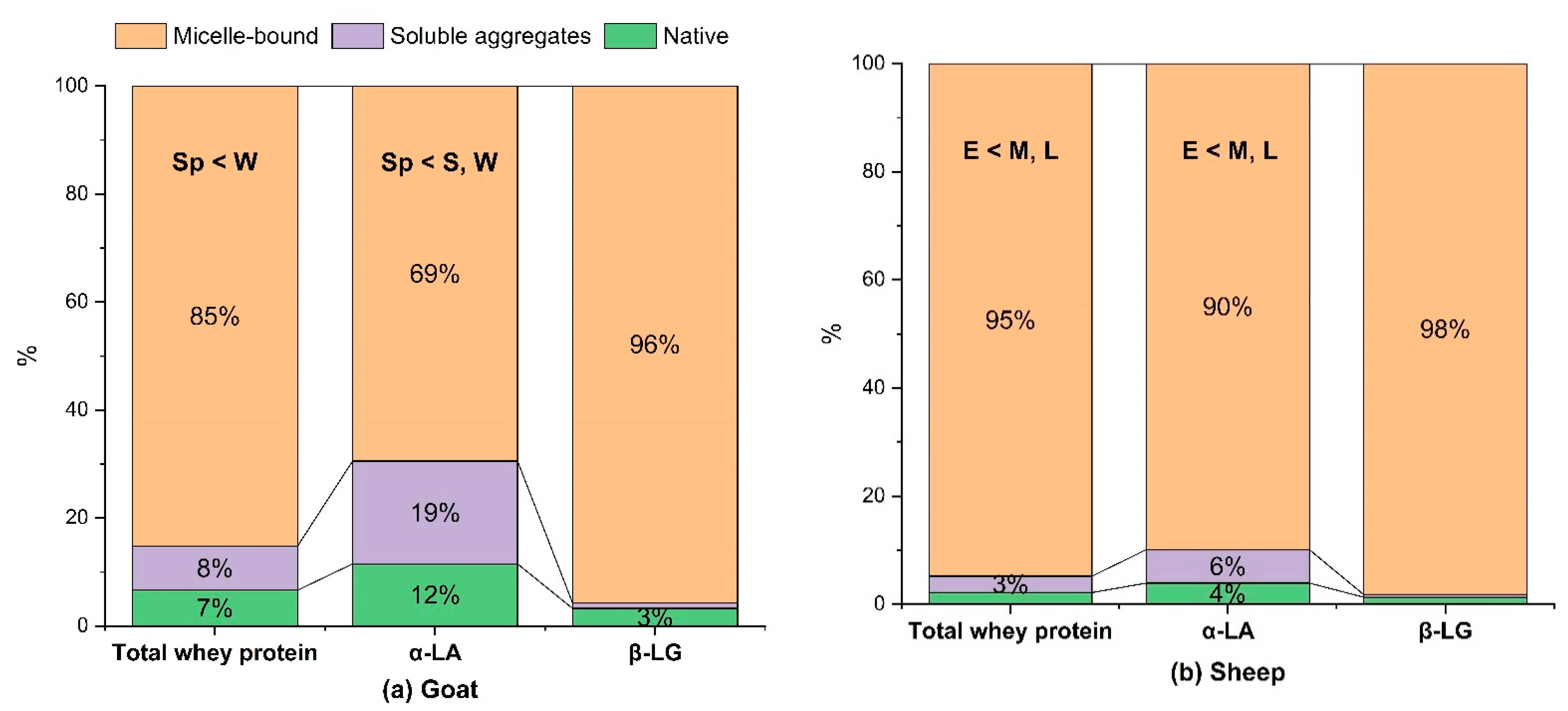
3.2.1. Whey Protein Denaturation and Association with the Casein Micelles
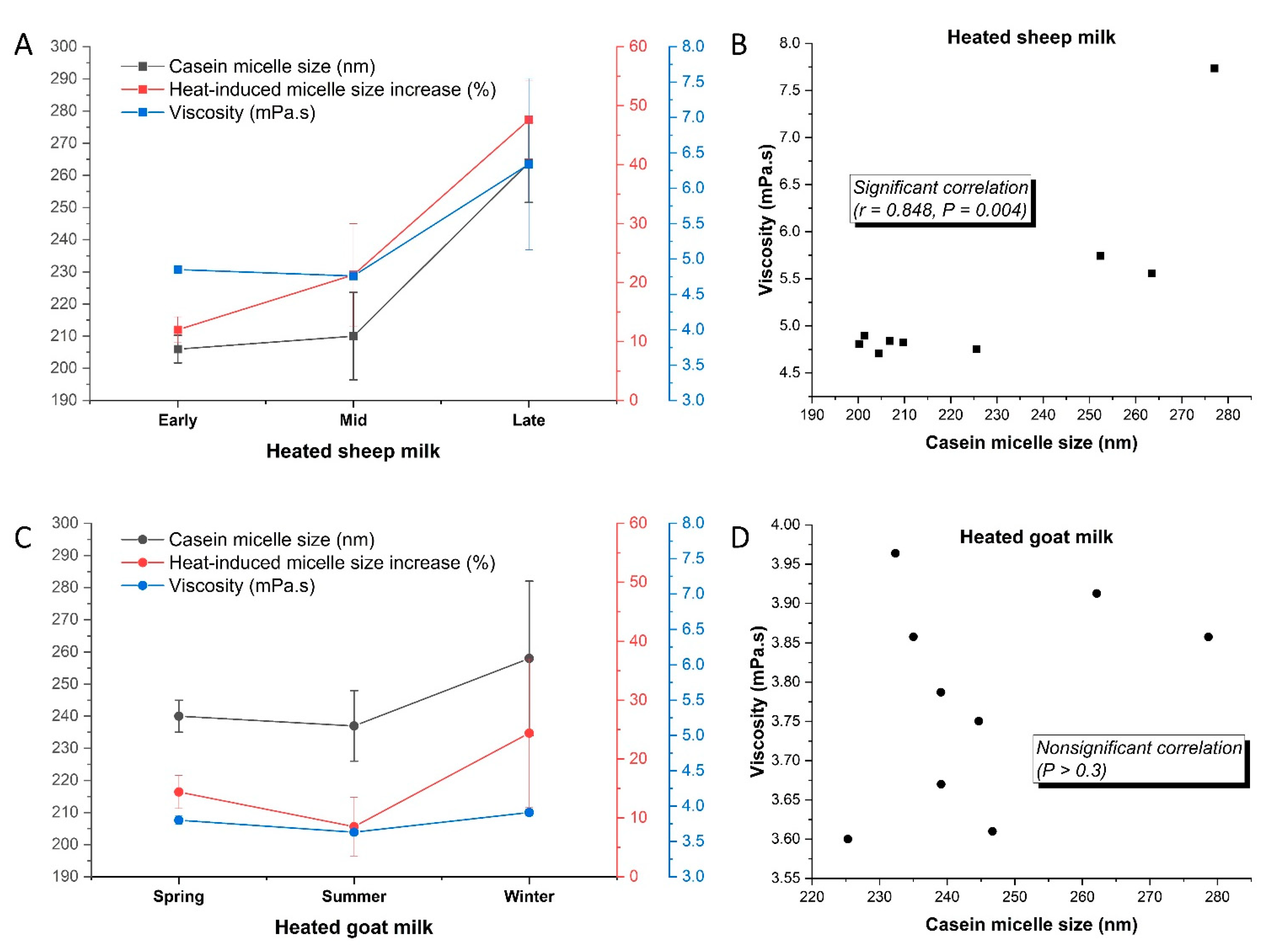
3.2.2. Casein Micelle Size and Milk Viscosity
3.2.3. Seasonal Variations in Heat-Induced Changes
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pulina, G.; Milán, M.J.; Lavín, M.P.; Theodoridis, A.; Morin, E.; Capote, J.; Thomas, D.L.; Francesconi, A.H.D.; Caja, G. Invited review: Current production trends, farm structures, and economics of the dairy sheep and goat sectors. J. Dairy Sci. 2018, 101, 6715–6729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.W.; Juárez, M.; Ramos, M.; Haenlein, G.F.W. Physico-chemical characteristics of goat and sheep milk. Small Rumin. Res. 2007, 68, 88–113. [Google Scholar] [CrossRef] [Green Version]
- Roy, D.; Ye, A.; Moughan, P.J.; Singh, H. Composition, structure, and digestive dynamics of milk from different species—A review. Front. Nutr. 2020, 7, 577759. [Google Scholar] [CrossRef] [PubMed]
- Raynal-Ljutovac, K.; Park, Y.W.; Gaucheron, F.; Bouhallab, S. Heat stability and enzymatic modifications of goat and sheep milk. Small Rumin. Res. 2007, 68, 207–220. [Google Scholar] [CrossRef]
- Heck, J.M.; van Valenberg, H.J.; Dijkstra, J.; van Hooijdonk, A.C. Seasonal variation in the Dutch bovine raw milk composition. J. Dairy Sci. 2009, 92, 4745–4755. [Google Scholar] [CrossRef]
- Li, S.; Ye, A.; Singh, H. Seasonal variations in composition, properties, and heat-induced changes in bovine milk in a seasonal calving system. J. Dairy Sci. 2019, 102, 7747–7759. [Google Scholar] [CrossRef]
- Mayer, H.K.; Fiechter, G. Physical and chemical characteristics of sheep and goat milk in Austria. Int. Dairy J. 2012, 24, 57–63. [Google Scholar] [CrossRef]
- Timlin, M.; Tobin, J.T.; Brodkorb, A.; Murphy, E.G.; Dillon, P.; Hennessy, D.; O’Donovan, M.; Pierce, K.M.; O’Callaghan, T.F. The impact of seasonality in pasture-based production systems on milk composition and functionality. Foods 2021, 10, 607. [Google Scholar] [CrossRef]
- Sevi, A.; Albenzio, M.; Marino, R.; Santillo, A.; Muscio, A. Effects of lambing season and stage of lactation on ewe milk quality. Small Rumin. Res. 2004, 51, 251–259. [Google Scholar] [CrossRef]
- Garzón, A.; Figueroa, A.; Caballero-Villalobos, J.; Angón, E.; Arias, R.; Perea, J.M. Derivation of multivariate indices of milk composition, coagulation properties, and curd yield in Manchega dairy sheep. J. Dairy Sci. 2021, 104, 8618–8629. [Google Scholar] [CrossRef]
- de la Fuente, L.F.; Barbosa, E.; Carriedo, J.A.; Gonzalo, C.; Arenas, R.; Fresno, J.M.; Primitivo, F.S. Factors influencing variation of fatty acid content in ovine milk. J. Dairy Sci. 2009, 92, 3791–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.R.; Dixon, P.H.; Park, Y.W.; Gilmore, J.A.; Kindstedt, P.S. Seasonal changes in the chemical composition of commingled goat milk. J. Dairy Sci. 2001, 84, E79–E83. [Google Scholar] [CrossRef]
- Auldist, M.J.; Walsh, B.J.; Thomson, N.A. Seasonal and lactational influences on bovine milk composition in New Zealand. J. Dairy Res. 1998, 65, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ye, A.; Singh, H. Seasonal variations in the characteristics of milk fat and the whipping properties of cream. Int. Dairy J. 2022, 127, 105288. [Google Scholar] [CrossRef]
- Chen, B.; Grandison, A.S.; Lewis, M.J. Comparison of heat stability of goat milk subjected to ultra-high temperature and in-container sterilization. J. Dairy Sci. 2012, 95, 1057–1063. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Ye, A.; Dave, A.; Fraser, K.; Singh, H. Kinetics of heat-induced interactions among whey proteins and casein micelles in sheep skim milk and aggregation of the casein micelles. J. Dairy Sci. 2022, 105, 3871–3882. [Google Scholar] [CrossRef]
- Nguyen, H.T.H.; Afsar, S.; Day, L. Differences in the microstructure and rheological properties of low-fat yoghurts from goat, sheep and cow milk. Food Res. Int. 2018, 108, 423–429. [Google Scholar] [CrossRef]
- Miocinovic, J.; Miloradovic, Z.; Josipovic, M.; Nedeljkovic, A.; Radovanovic, M.; Pudja, P. Rheological and textural properties of goat and cow milk set type yoghurts. Int. Dairy J. 2016, 58, 43–45. [Google Scholar] [CrossRef]
- Li, S.; Pan, Z.; Ye, A.; Cui, J.; Dave, A.; Singh, H. Structural and rheological properties of the clots formed by ruminant milks during dynamic in vitro gastric digestion: Effects of processing and species. Food Hydrocoll. 2022, 126, 107465. [Google Scholar] [CrossRef]
- Li, S.; Ye, A.; Pan, Z.; Cui, J.; Dave, A.; Singh, H. Dynamic in vitro gastric digestion behavior of goat milk: Effects of homogenization and heat treatments. J. Dairy Sci. 2022, 105, 965–980. [Google Scholar] [CrossRef]
- Pan, Z.; Ye, A.; Li, S.; Dave, A.; Fraser, K.; Singh, H. Dynamic in vitro gastric digestion of sheep milk: Influence of homogenization and heat treatment. Foods 2021, 10, 1938. [Google Scholar] [CrossRef] [PubMed]
- Fecher, P.A.; Goldmann, I.; Nagengast, A. Determination of iodine in food samples by inductively coupled plasma mass spectrometry after alkaline extraction. J. Anal. At. Spectrom. 1998, 13, 977–982. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for Examination of Water and Wastewater, 22nd ed.; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- Hara, A.; Radin, N.S. Lipid extraction of tissues. Anal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Loveday, S.M.; Fraser, K.; Luo, D.; Weeks, M.; Cakebread, J.A. A multivariate snapshot of New Zealand milk seasonality in individual cows. Int. Dairy J. 2021, 114, 104940. [Google Scholar] [CrossRef]
- Bobe, G.; Beitz, D.C.; Freeman, A.E.; Lindberg, G.L. Separation and quantification of bovine milk proteins by reversed-phase high-performance liquid chromatography. J. Agric. Food Chem. 1998, 46, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ye, A.; Singh, H. Physicochemical changes and age gelation in stored UHT milk: Seasonal variations. Int. Dairy J. 2021, 118, 105028. [Google Scholar] [CrossRef]
- Gaucheron, F. The minerals of milk. Reprod. Nutr. Dev. 2005, 45, 473–483. [Google Scholar] [CrossRef]
- Raynal-Ljutovac, K.; Lagriffoul, G.; Paccard, P.; Guillet, I.; Chilliard, Y. Composition of goat and sheep milk products: An update. Small Rumin. Res. 2008, 79, 57–72. [Google Scholar] [CrossRef]
- De La Fuente, M.A.; Olano, A.; Juárez, M. Distribution of calcium, magnesium, phosphorus, zinc, manganese, copper and iron between the soluble and colloidal phases of ewe’s and goat’s milk. Lait 1997, 77, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Lewis, M. The measurement and significance of ionic calcium in milk—A review. Int. J. Dairy Technol. 2011, 64, 1–13. [Google Scholar] [CrossRef]
- Kljajevic, N.V.; Tomasevic, I.B.; Miloradovic, Z.N.; Nedeljkovic, A.; Miocinovic, J.B.; Jovanovic, S.T. Seasonal variations of Saanen goat milk composition and the impact of climatic conditions. J. Food Sci. Technol. 2018, 55, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Couvreur, S.; Hurtaud, C.; Lopez, C.; Delaby, L.; Peyraud, J.L. The linear relationship between the proportion of fresh grass in the cow diet, milk fatty acid composition, and butter properties. J. Dairy Sci. 2006, 89, 1956–1969. [Google Scholar] [CrossRef]
- Tsiplakou, E.; Flemetakis, E.; Kalloniati, C.; Papadomichelakis, G.; Katinakis, P.; Zervas, G. Sheep and goats differences in cla and fatty acids milk fat content in relation with MRNA stearoyl-Coa desaturase and lipogenic genes expression in their mammary gland. J. Dairy Res. 2009, 76, 392–401. [Google Scholar] [CrossRef]
- Clark, S.; Mora García, M.B. A 100-year review: Advances in goat milk research. J. Dairy Sci. 2017, 100, 10026–10044. [Google Scholar] [CrossRef] [PubMed]
- Wendorff, W.L.; Haenlein, G.F.W. Sheep milk—Composition and nutrition. In Handbook of Milk of Non-Bovine Mammals, 2nd ed.; Park, Y.W., Wendorff, W.L., Haenlein, G.F.W., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2017; pp. 210–221. [Google Scholar]
- Tudisco, R.; Grossi, M.; Calabrò, S.; Cutrignelli, M.I.; Musco, N.; Addi, L.; Infascelli, F. Influence of pasture on goat milk fatty acids and stearoyl-CoA desaturase expression in milk somatic cells. Small Rumin. Res. 2014, 122, 38–43. [Google Scholar] [CrossRef]
- Chilliard, Y.; Rouel, J.; Leroux, C. Goat’s alpha-s1 casein genotype influences its milk fatty acid composition and delta-9 desaturation ratios. Anim. Feed Sci. Technol. 2006, 131, 474–487. [Google Scholar] [CrossRef]
- Barłowska, J.; Szwajkowska, M.; Litwińczuk, Z.; Król, J. Nutritional value and technological suitability of milk from various animal species used for dairy production. Compr. Rev. Food Sci. Food Saf. 2011, 10, 291–302. [Google Scholar] [CrossRef]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, Rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Norris, G.E.; Gray, I.K.; Dolby, R.M. Seasonal variations in the composition and thermal properties of New Zealand milk fat: Ii. Thermal properties of milk fat and their relation to composition. J. Dairy Res. 1973, 40, 311–321. [Google Scholar] [CrossRef]
- Casoli, C.; Duranti, E.; Morbidini, L.; Panella, F.; Vizioli, V. Quantitative and compositional variations of Massese sheep milk by parity and stage of lactation. Small Rumin. Res. 1989, 2, 47–62. [Google Scholar] [CrossRef]
- Soyeurt, H.; Dehareng, F.; Mayeres, P.; Bertozzi, C.; Gengler, N. Variation of Δ9-desaturase activity in dairy cattle. J. Dairy Sci. 2008, 91, 3211–3224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mele, M.; Dal Zotto, R.; Cassandro, M.; Conte, G.; Serra, A.; Buccioni, A.; Bittante, G.; Secchiari, P. Genetic parameters for conjugated linoleic acid, selected milk fatty acids, and milk fatty acid unsaturation of Italian Holstein-Friesian cows. J. Dairy Sci. 2009, 92, 392–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siefarth, C.; Buettner, A. The aroma of goat milk: Seasonal effects and changes through heat treatment. J. Agric. Food Chem. 2014, 62, 11805–11817. [Google Scholar] [CrossRef] [PubMed]
- Lock, A.L.; Garnsworthy, P.C. Seasonal variation in milk conjugated linoleic acid and Δ9-desaturase activity in dairy cows. Livest. Prod. Sci. 2003, 79, 47–59. [Google Scholar] [CrossRef]
- Moioli, B.; Pilla, F.; Tripaldi, C. Detection of milk protein genetic polymorphisms in order to improve dairy traits in sheep and goats: A review. Small Rumin. Res. 1998, 27, 185–195. [Google Scholar] [CrossRef]
- Ha, M.; Bekhit, A.E.D.; McConnell, M.; Mason, S.; Carne, A. Fractionation of whey proteins from red deer (Cervus elaphus) milk and comparison with whey proteins from cow, sheep and goat milks. Small Rumin. Res. 2014, 120, 125–134. [Google Scholar] [CrossRef]
- Claeys, W.L.; Verraes, C.; Cardoen, S.; De Block, J.; Huyghebaert, A.; Raes, K.; Dewettinck, K.; Herman, L. Consumption of raw or heated milk from different species: An evaluation of the nutritional and potential health benefits. Food Control 2014, 42, 188–201. [Google Scholar] [CrossRef]
- Selvaggi, M.; Laudadio, V.; Dario, C.; Tufarelli, V. Investigating the genetic polymorphism of sheep milk proteins: A useful tool for dairy production. J. Sci. Food Agric. 2014, 94, 3090–3099. [Google Scholar] [CrossRef]
- Pirisi, A.; Piredda, G.; Papoff, C.M.; Di Salvo, R.; Pintus, S.; Garro, G.; Ferranti, P.; Chianese, L. Effects of sheep αs1-casein CC, CD and DD Genotypes on milk composition and cheesemaking properties. J. Dairy Res. 1999, 66, 409–419. [Google Scholar] [CrossRef]
- Moatsou, G.; Hatzinaki, A.; Samolada, M.; Anifantakis, E. Major whey proteins in ovine and caprine acid wheys from indigenous Greek breeds. Int. Dairy J. 2005, 15, 123–131. [Google Scholar] [CrossRef]
- de la Vara, J.A.; Berruga, M.I.; Cappelli, J.; Landete-Castillejos, T.; Carmona, M.; Gallego, L.; Molina, A. Some aspects of the ethanol stability of red deer milk (Cervus elaphus hispanicus): A comparison with other dairy species. Int. Dairy J. 2018, 86, 103–109. [Google Scholar] [CrossRef]
- Dalgleish, D.G.; Corredig, M. The structure of the casein micelle of milk and its changes during processing. Annu. Rev. Food Sci. Technol. 2012, 3, 449–467. [Google Scholar] [CrossRef] [PubMed]
- de Kruif, C.G.; Huppertz, T. Casein micelles: Size distribution in milks from individual cows. J. Agric. Food Chem. 2012, 60, 4649–4655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bijl, E.; de Vries, R.; van Valenberg, H.; Huppertz, T.; Van Hooijdonk, T. Factors influencing casein micelle size in milk of individual cows: Genetic variants and glycosylation of κ-casein. Int. Dairy J. 2014, 34, 135–141. [Google Scholar] [CrossRef]
- Raynal, K.; Remeuf, F. The effect of heating on physicochemical and renneting properties of milk: A comparison between caprine, ovine and bovine milk. Int. Dairy J. 1998, 8, 695–706. [Google Scholar] [CrossRef]
- Pesic, M.B.; Barac, M.B.; Stanojevic, S.P.; Ristic, N.M.; Macej, O.D.; Vrvic, M.M. Heat induced casein-whey protein interactions at natural ph of milk: A comparison between caprine and bovine milk. Small Rumin. Res. 2012, 108, 77–86. [Google Scholar] [CrossRef]
- Anema, S.G.; Li, Y. Association of denatured whey proteins with casein micelles in heated reconstituted skim milk and its effect on casein micelle size. J. Dairy Res. 2003, 70, 73–83. [Google Scholar] [CrossRef]
- Dumpler, J.; Wohlschläger, H.; Kulozik, U. Dissociation and coagulation of caseins and whey proteins in concentrated skim milk heated by direct steam injection. Dairy Sci. Technol. 2017, 96, 807–826. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Liu, D.; Peng, X.; Jiang, Y.; Gao, Y.; Liu, X.; Regenstein, J.M.; Zhou, P. The heat stability of caprine and bovine micellar casein dispersions. Int. Dairy J. 2022, 131, 105373. [Google Scholar] [CrossRef]
- Singh, H. Heat stability of milk. Int. J. Dairy Technol. 2004, 57, 111–119. [Google Scholar] [CrossRef]
- Jeurnink, T.J.M.; De Kruif, K.G. Changes in milk on heating: Viscosity measurements. J. Dairy Res. 1993, 60, 139–150. [Google Scholar] [CrossRef]
- Lewis, T.B.; Nielsen, L.E. Viscosity of dispersed and aggregated suspensions of spheres. Trans. Soc. Rheol. 1968, 12, 421–443. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Goat | Sheep | |||||
|---|---|---|---|---|---|---|
| Mean ± SD | Min–Max | Seasonal Effect * | Mean ± SD | Min–Max | Seasonal Effect * | |
| Total solids (wt/wt%) | 11.90 ± 0.53 b | 11.22–12.41 | S < Sp, W | 17.46 ± 0.60 a | 16.91–18.55 | L > E, M |
| Protein (wt/wt%) | 3.19 ± 0.08 b | 3.09–3.30 | S < W | 5.72 ± 0.28 a | 5.45–6.26 | L > E, M |
| Fat (wt/wt%) | 3.79 ± 0.39 b | 3.30–4.27 | S < Sp, W | 5.99 ± 0.44 a | 5.43–6.70 | L > E, M |
| Lactose (wt/wt%) | 4.40 ± 0.05 b | 4.28–4.45 | NS | 4.76 ± 0.12 a | 4.58–4.85 | L < E, M |
| Ca (g/100 g) | 0.114 ± 0.005 b | 0.108–0.121 | W > Sp, S | 0.200 ± 0.007 a | 0.189–0.210 | NS |
| Soluble Ca (% in total Ca) | 29.7 ± 1.8 a | 27.3–33.3 | NS | 19.6 ± 0.8 b | 18.5–20.5 | NS |
| Ionic Ca (% in total Ca) | 10.8 ± 0.8 a | 10.0–12.6 | NS | 5.2 ± 0.4 b | 4.9–5.8 | NS |
| Magnesium (g/100 g) | 0.014 ± 0.001 b | 0.013–0.015 | NS | 0.018 ± 0.002 a | 0.016–0.021 | L > E, M |
| Soluble Mg (% in total Mg) | 69.2 ± 4.3 a | 64.8–78.8 | NS | 58.2 ± 1.3 b | 55.4–60.0 | NS |
| Phosphorus (g/100 g) | 0.100 ± 0.003 b | 0.095–0.104 | W > S | 0.160 ± 0.003 a | 0.156–0.165 | M < E, L |
| Soluble P (% in total P) | 49.7 ± 3.8 a | 45.6–57.6 | NS | 38.7 ± 2.1 b | 35.6–42.9 | NS |
| Potassium (g/100 g) | 0.203 ± 0.009 a | 0.195–0.220 | S > Sp, W | 0.132 ± 0.007 b | 0.119–0.138 | L < E, M |
| Sodium (g/100 g) | 0.037 ± 0.001 | 0.035–0.038 | NS | 0.042 ± 0.007 | 0.035–0.052 | L > E, M |
| Chloride (g/100 g) | 0.164 ± 0.008 a | 0.153–0.175 | S > Sp, W | 0.089 ± 0.012 b | 0.071–0.105 | L > M > E |
| Copper (mg/kg) | 0.103 ± 0.020 b | 0.064–0.129 | S < Sp, W | 0.171 ± 0.072 a | 0.099–0.290 | E > M, L |
| Iodine (mg/kg) | 0.251 ± 0.049 | 0.176–0.330 | S > W | 0.224 ± 0.042 | 0.153–0.270 | NS |
| Selenium (mg/kg) | 0.029 ± 0.004 b | 0.023–0.035 | NS | 0.036 ± 0.005 a | 0.030–0.044 | M > E, L |
| Zinc (mg/kg) | 3.578 ± 0.286 b | 3.200–4.000 | S < Sp, W | 5.822 ± 0.307 a | 5.300–6.400 | E > L |
| Goat | Sheep | |||||
|---|---|---|---|---|---|---|
| Item | Mean ± SD | Min–Max | Seasonal Effect * | Mean ± SD | Min–Max | Seasonal Effect * |
| C4:0 | 2.62 ± 0.09 b | 2.45–2.74 | S < Sp, W | 3.44 ± 0.26 a | 3.00–3.78 | L < E, M |
| C6:0 | 2.47 ± 0.13 b | 2.30–2.68 | Sp > W | 2.76 ± 0.26 a | 2.32–3.08 | L < E, M |
| C8:0 | 2.73 ± 0.21 | 2.37–3.07 | NS | 2.81 ± 0.31 | 2.30–3.14 | L < E, M |
| C10:0 | 8.56 ± 0.91 | 6.79–9.58 | NS | 9.15 ± 0.74 | 8.07–9.96 | M > L |
| C10:1 | 0.20 ± 0.02 b | 0.17–0.23 | NS | 0.27 ± 0.04 a | 0.23–0.33 | L > E, M |
| C12:0 | 3.76 ± 0.63 b | 2.78–4.91 | NS | 5.33 ± 0.40 a | 4.51–5.76 | NS |
| C14:0 | 8.37 ± 0.99 b | 6.71–9.66 | S > W | 11.20 ± 1.07 a | 9.47–12.60 | L > M > E |
| iso C15 | 0.15 ± 0.03 b | 0.11–0.19 | S > Sp, W | 0.20 ± 0.01 a | 0.17–0.21 | E < M, L |
| anteiso C15 | 0.26 ± 0.04 b | 0.20–0.32 | S > Sp, W | 0.40 ± 0.03 a | 0.36–0.45 | M > E, L |
| C14:1 | 0.10 ± 0.01 b | 0.08–0.12 | NS | 0.19 ± 0.08 a | 0.11–0.33 | L > E, M |
| C15:0 | 0.66 ± 0.08 b | 0.57–0.77 | S > Sp, W | 1.09 ± 0.09 a | 0.95–1.23 | L > M > E |
| iso C16 | 0.19 ± 0.01 b | 0.17–0.21 | S > W | 0.23 ± 0.01 a | 0.21–0.25 | NS |
| C16:0 | 25.35 ± 1.51 | 22.83–28.20 | NS | 24.08 ± 1.28 | 22.39–26.24 | L > E, M |
| iso C17 | 0.33 ± 0.04 | 0.28–0.39 | Sp < S, W | 0.33 ± 0.02 | 0.29–0.36 | NS |
| C16:1 | 0.52 ± 0.04 b | 0.48–0.59 | W > Sp, S | 0.87 ± 0.16 a | 0.71–1.14 | L > E, M |
| anteiso C17 | 0.35 ± 0.04 | 0.31–0.43 | NS | 0.38 ± 0.01 | 0.35–0.40 | NS |
| C17:0 | 0.47 ± 0.04 b | 0.41–0.53 | S > Sp | 0.55 ± 0.05 a | 0.51–0.67 | NS |
| C17:1 | 0.19 ± 0.03 b | 0.15–0.22 | W > Sp | 0.23 ± 0.03 a | 0.21–0.29 | NS |
| C18:0 | 10.79 ± 1.05 a | 9.93–12.47 | W > Sp, S | 7.41 ± 0.47 b | 6.76–8.20 | NS |
| C18:1 t9 | 0.25 ± 0.04 | 0.21–0.30 | W > Sp, S | 0.22 ± 0.03 | 0.19–0.30 | NS |
| C18:1 t10 | 0.45 ± 0.09 b | 0.32–0.61 | Sp > S | 0.71 ± 0.23 a | 0.49–1.21 | NS |
| C18:1 t11 | 1.33 ± 0.22 b | 1.07–1.73 | W > S | 2.03 ± 0.56 a | 1.15–2.92 | E > L |
| C18:1 c9 | 21.64 ± 2.16 a | 19.09–25.60 | W > Sp, S | 15.04 ± 1.09 b | 13.89–17.45 | NS |
| C18:1 c11 | 0.33 ± 0.04 b | 0.27–0.41 | NS | 0.42 ± 0.02 a | 0.40–0.44 | NS |
| C18:2 n6 | 3.61 ± 0.59 a | 2.88–4.69 | Sp > S, W | 1.94 ± 0.24 b | 1.67–2.35 | E > M, L |
| C20:0 | 0.18 ± 0.01 a | 0.17–0.20 | W > Sp | 0.15 ± 0.02 b | 0.12–0.18 | L > E, M |
| C18:3 n3 | 0.42 ± 0.04 b | 0.37–0.51 | NS | 1.06 ± 0.08 a | 0.96–1.20 | E > M |
| CLA | 0.79 ± 0.10 b | 0.67–0.94 | W > Sp, S | 1.15 ± 0.20 a | 0.86–1.47 | NS |
| C20:4 n6 | 0.21 ± 0.02 a | 0.19–0.24 | NS | 0.16 ± 0.01 b | 0.15–0.18 | E > M, L |
| SFA | 67.2 ± 2.8 b | 62.9–71.1 | NS | 69.6 ± 1.5 a | 66.5–71.1 | E > M |
| MUFA | 25.1 ± 2.4 a | 22.0–29.3 | W > Sp, S | 20.0 ± 0.9 b | 19.2–22.2 | NS |
| PUFA | 5.1 ± 0.6 | 4.4–6.3 | NS | 4.7 ± 0.4 | 4.3–5.3 | E > M, L |
| C10:1/C10:0 | 0.024 ± 0.001 b | 0.022–0.026 | NS | 0.030 ± 0.006 a | 0.025–0.041 | L > E, M |
| C14:1/C14:0 | 0.012 ± 0.001 b | 0.011–0.013 | NS | 0.016 ± 0.005 a | 0.012–0.026 | L > E, M |
| C16:1/C16:0 | 0.021 ± 0.002 b | 0.017–0.024 | W > Sp, S | 0.036 ± 0.005 a | 0.030–0.043 | L > E, M |
| C18:1 c9/C18:0 | 2.01 ± 0.02 | 1.92–2.13 | NS | 2.03 ± 0.15 | 1.84–2.29 | L > M |
| CLA/C18:1 t11 | 0.59 ± 0.01 | 0.54–0.65 | S > Sp, W | 0.58 ± 0.07 | 0.51–0.74 | L > E |
| LMF | 56.1 ± 3.0 b | 53.4–60.4 | W > Sp, S | 59.4 ± 1.7 a | 55.4–61.1 | NS |
| MMF | 28.6 ± 2.8 | 24.6–32.2 | W < Sp, S | 26.3 ± 2.0 | 23.6–30.1 | NS |
| HMF | 15.3 ± 0.7 | 14.5–16.6 | NS | 14.4 ± 1.3 | 12.7–13.7 | L > E, M |
| Goat | Sheep | |||
|---|---|---|---|---|
| Mean ± SD | Seasonal Effect * | Mean ± SD | Seasonal Effect * | |
| κ-Casein | 16.6 ± 0.7 | S > Sp > W | 11.2 ± 1.0 | E < M, L |
| Serum κ-casein (% of total κ-casein) | 22.5 ± 4.9 | Sp > S, W | 8.1 ± 1.8 | NS |
| αs1-Casein | 8.9 ± 0.4 | NS | 34.1 ± 1.1 | NS |
| αs2-Casein | 10.9 ± 0.4 | NS | 11.9 ± 1.0 | NS |
| β-Casein | 44.2 ± 0.8 | NS | 29.0 ± 0.5 | NS |
| Serum β-casein (% of total β-casein) | 16.0 ± 4.3 | NS | 6.7 ± 2.3 | E > M, L |
| α-Lactalbumin | 8.3 ± 0.4 | NS | 4.7 ± 0.5 | NS |
| β-Lactoglobulin | 11.1 ± 0.8 | W > S, Sp | 9.1 ± 0.5 | L > E, M |
| Goat | Sheep | |||
|---|---|---|---|---|
| Mean ± SD | Seasonal Effect * | Mean ± SD | Seasonal Effect * | |
| pH | 6.67 ± 0.03 | NS | 6.60 ± 0.05 | L < E, M |
| Fat globule size (µm, D[4,3]) | 4.00 ± 0.07 b | NS | 4.50 ± 0.12 a | E > L |
| Ethanol stability (%) | 48.6 ± 2.5 | Sp > S, W | 51.4 ± 2.8 | L < E, M |
| Casein micelle size (nm) | 212 ± 9 a | NS | 179 ± 5 b | E > L > M |
| Viscosity (mPa⋅s) | 3.75 ± 0.12 b | NS | 4.64 ± 0.24 a | L > E |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Delger, M.; Dave, A.; Singh, H.; Ye, A. Seasonal Variations in the Composition and Physicochemical Characteristics of Sheep and Goat Milks. Foods 2022, 11, 1737. https://doi.org/10.3390/foods11121737
Li S, Delger M, Dave A, Singh H, Ye A. Seasonal Variations in the Composition and Physicochemical Characteristics of Sheep and Goat Milks. Foods. 2022; 11(12):1737. https://doi.org/10.3390/foods11121737
Chicago/Turabian StyleLi, Siqi, Munkhzul Delger, Anant Dave, Harjinder Singh, and Aiqian Ye. 2022. "Seasonal Variations in the Composition and Physicochemical Characteristics of Sheep and Goat Milks" Foods 11, no. 12: 1737. https://doi.org/10.3390/foods11121737
APA StyleLi, S., Delger, M., Dave, A., Singh, H., & Ye, A. (2022). Seasonal Variations in the Composition and Physicochemical Characteristics of Sheep and Goat Milks. Foods, 11(12), 1737. https://doi.org/10.3390/foods11121737