
Antimicrobial and Antibiofilm Effect of ε-Polylysine against Salmonella Enteritidis, Listeria monocytogenes, and Escherichia coli in Tryptic Soy Broth and Chicken Juice

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Growth Conditions, and Preparation of Experimental Group
2.2. Preparation of Chicken Juice
2.3. Minimum Inhibitory Concentration (MIC) of ε-PL
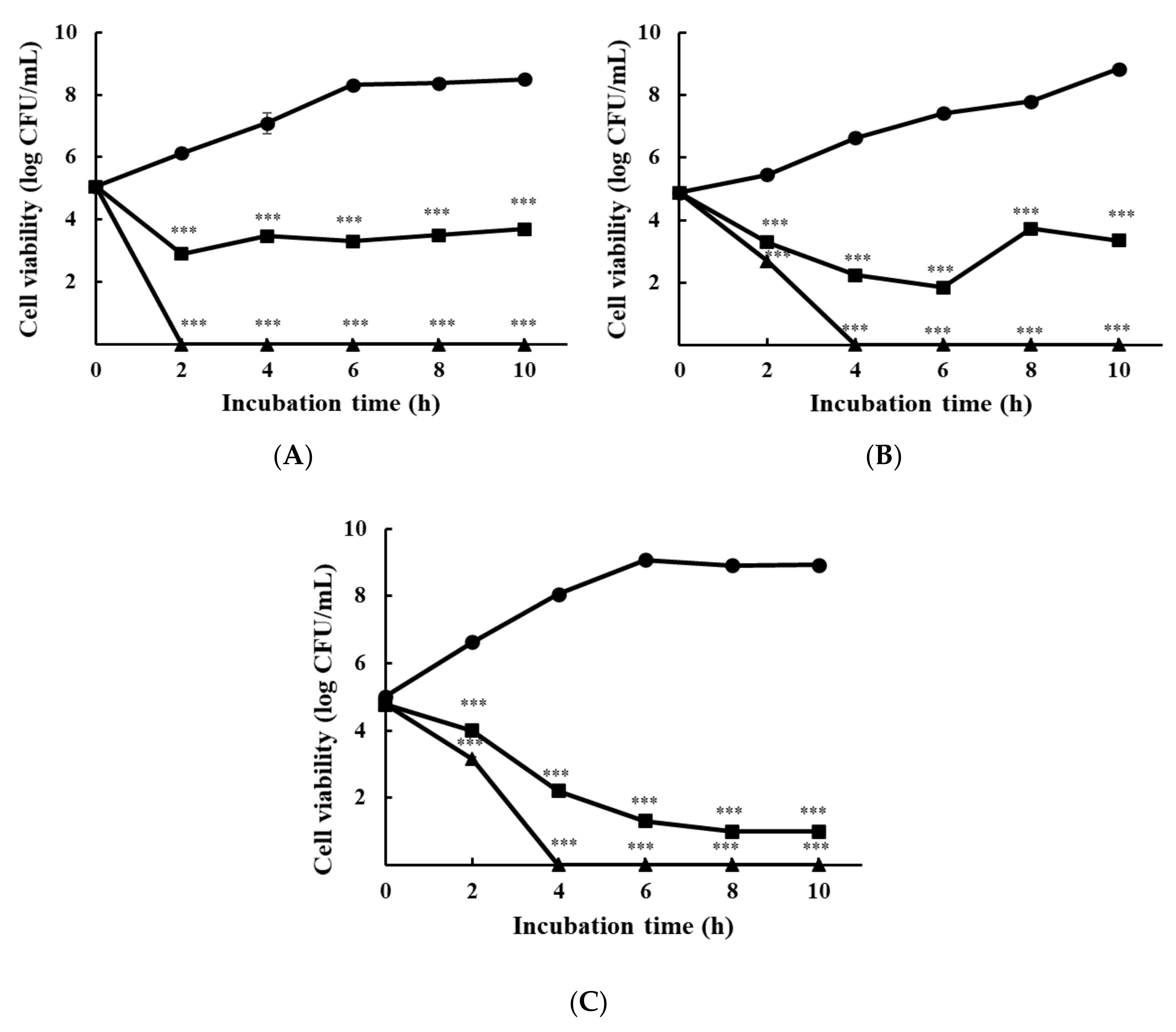
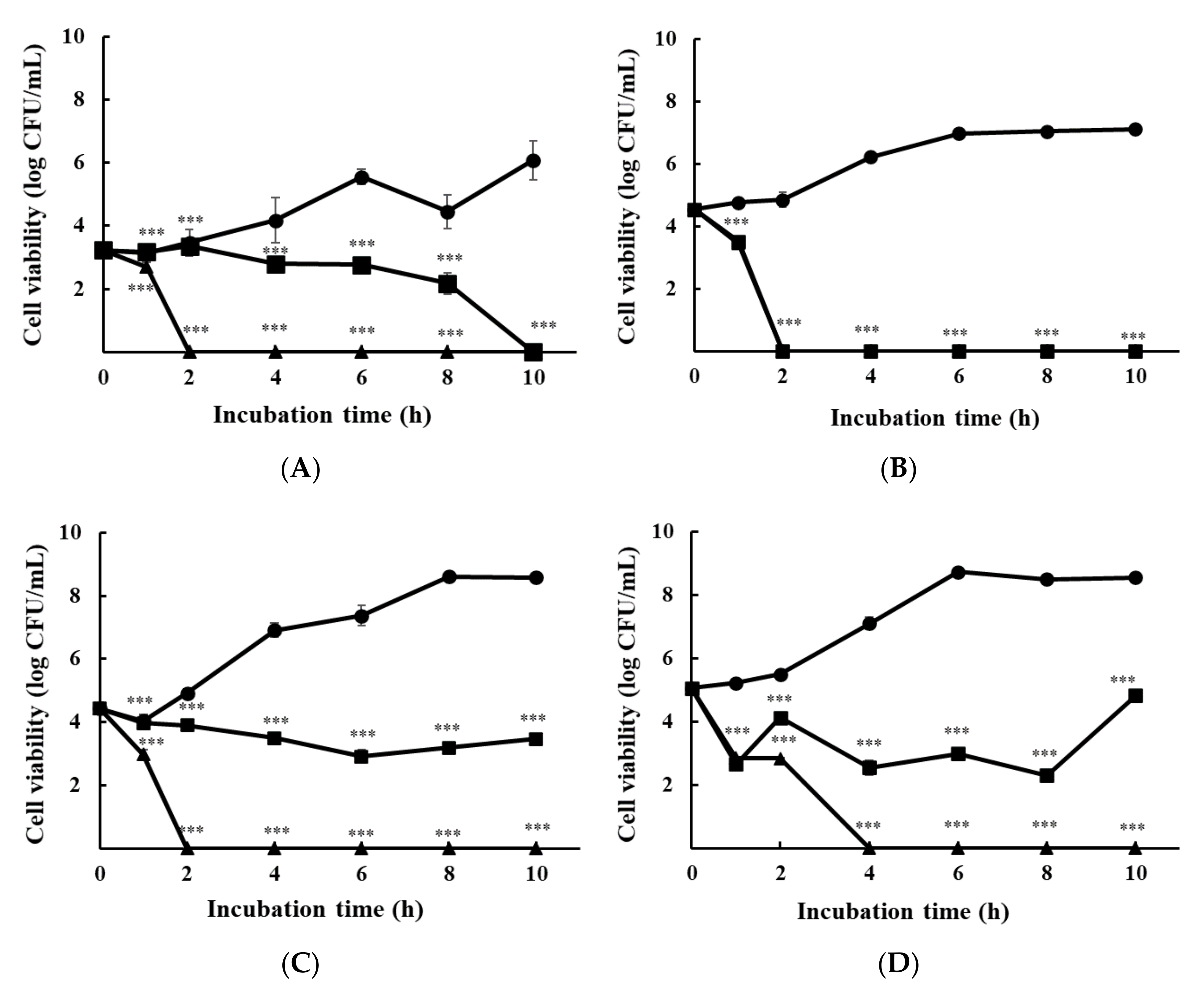
2.4. Time-Kill Assay
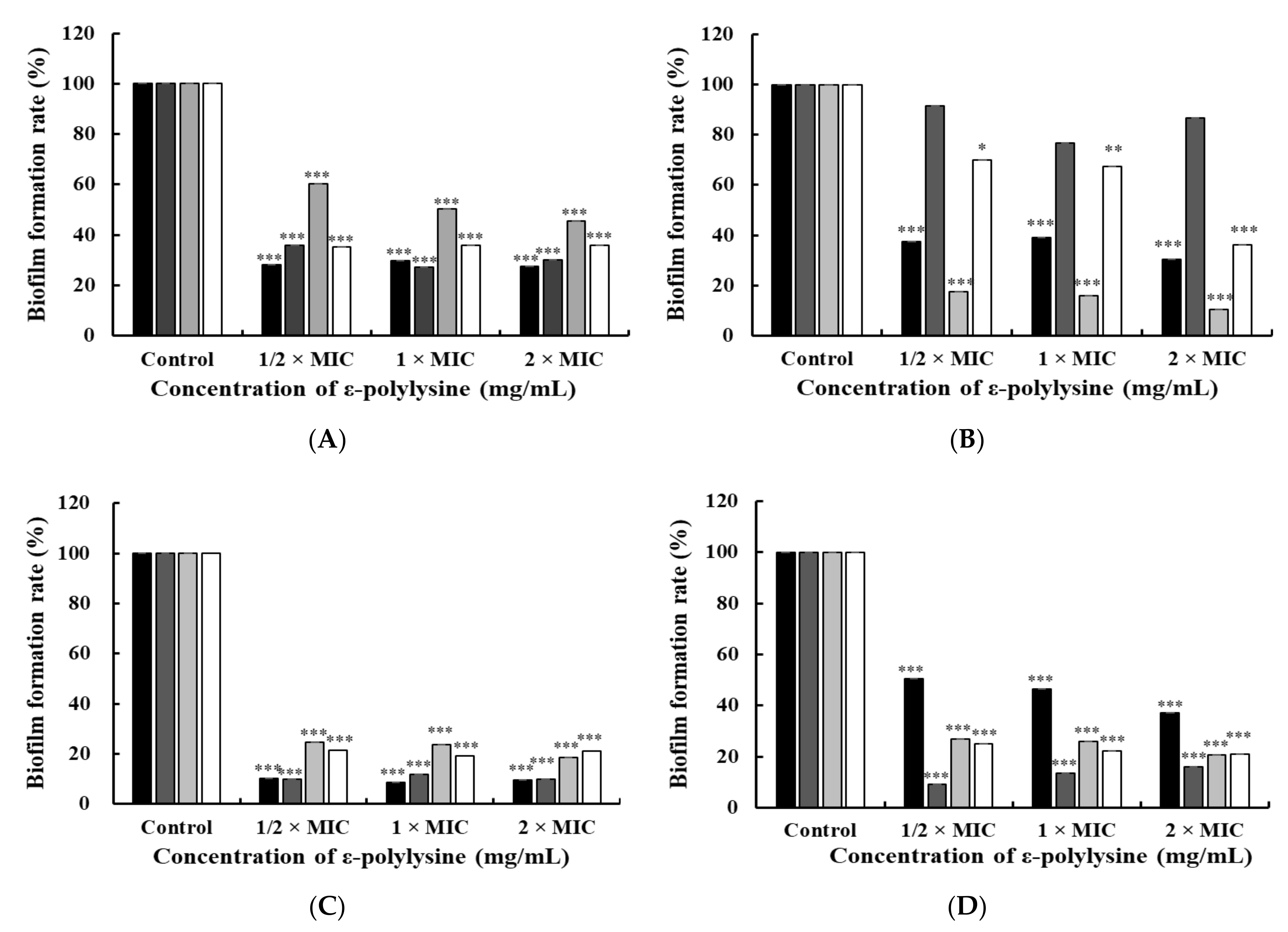
2.5. Biofilm Assay
2.6. Hydrophobicity and Auto-Aggregation
2.7. Total EPS Production Rate
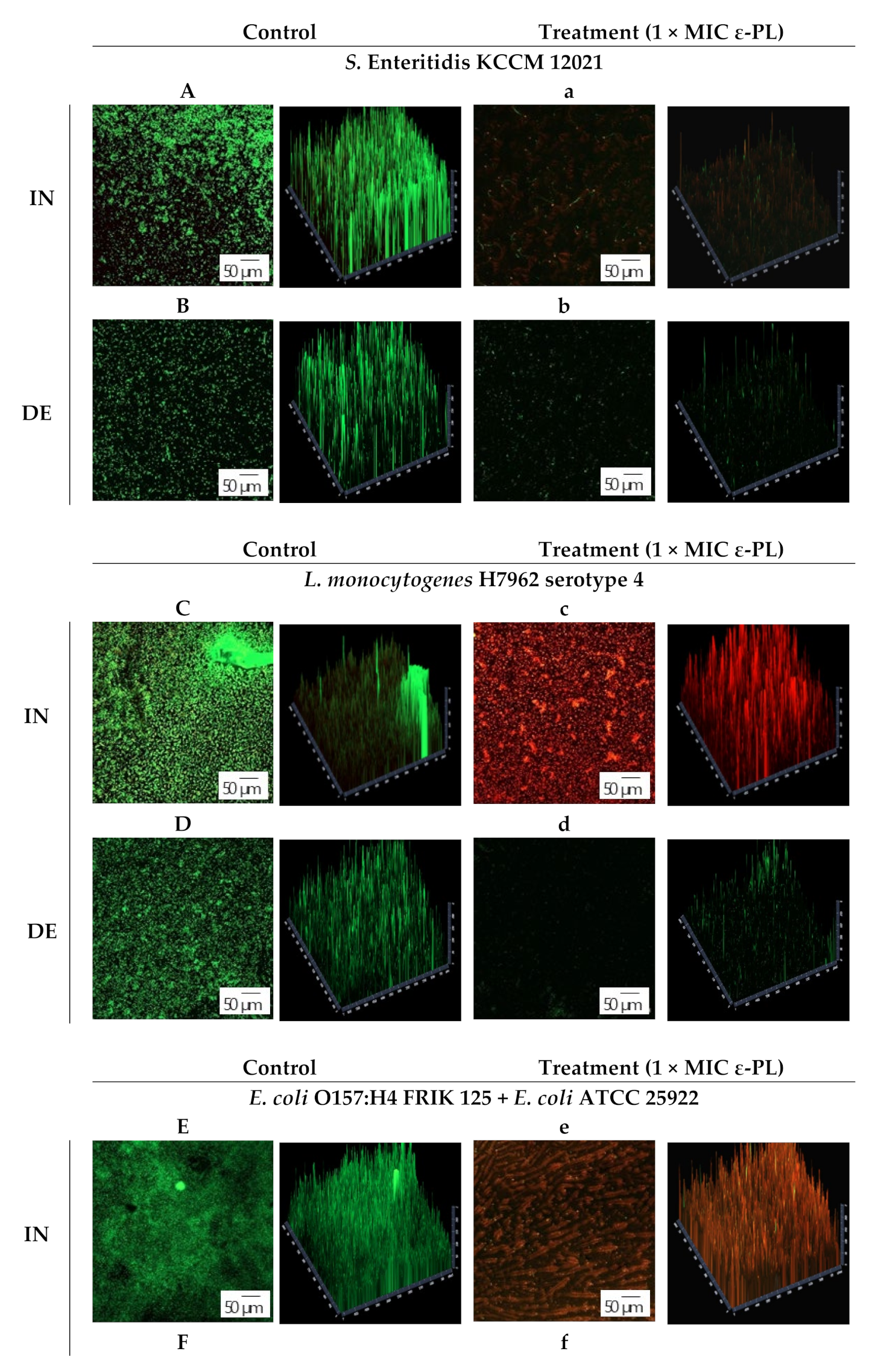
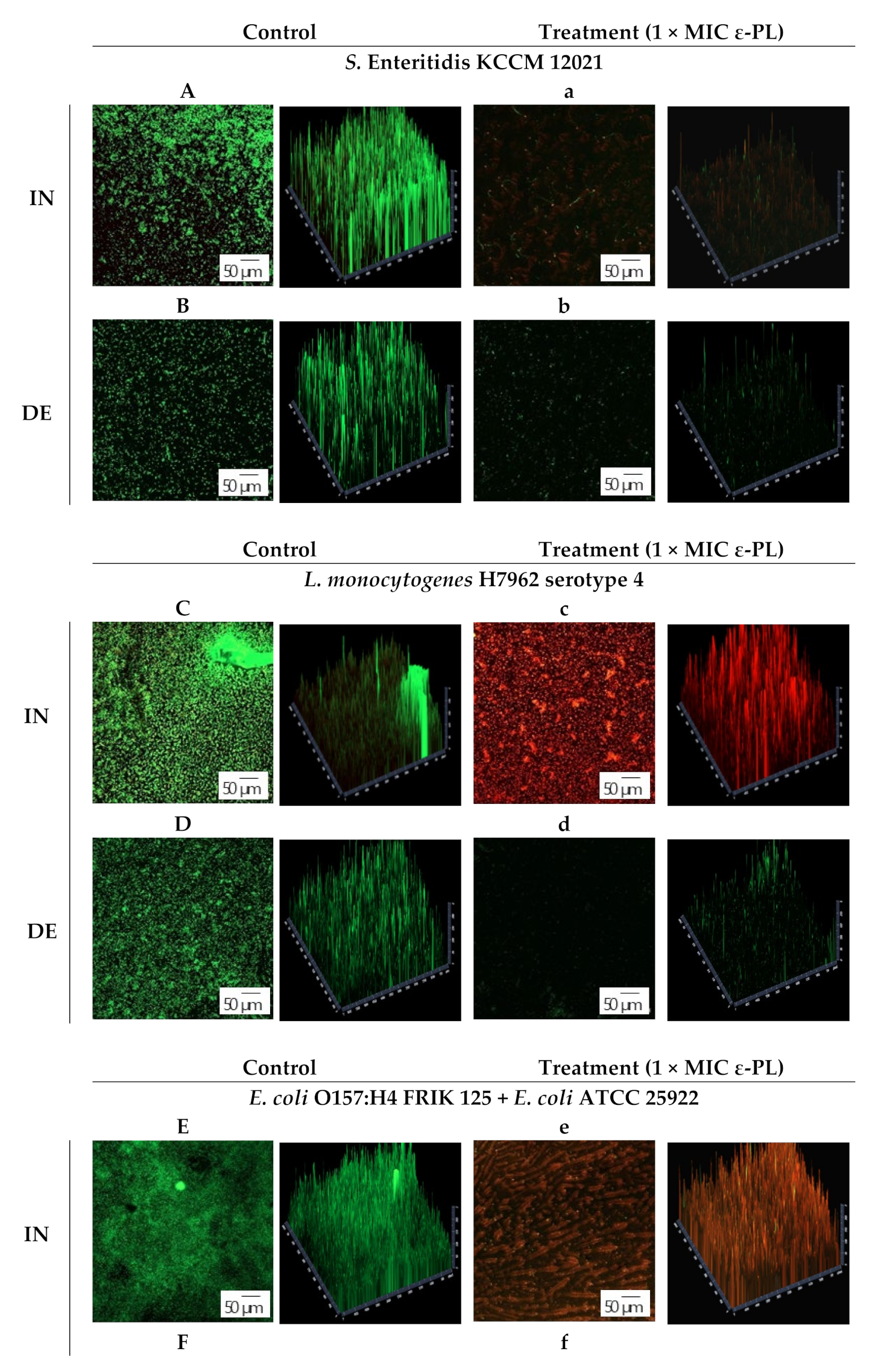
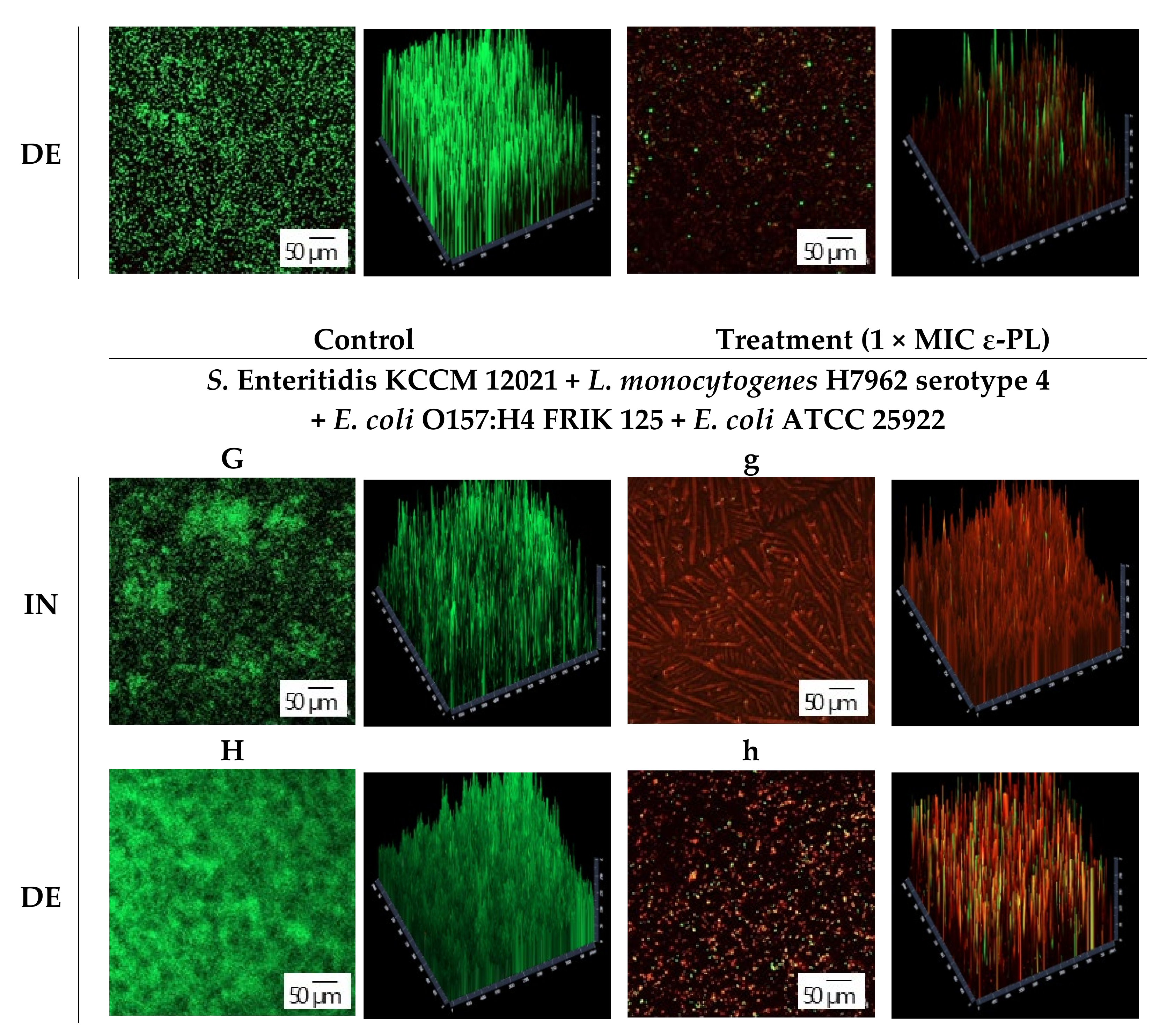
2.8. Confocal Laser Scanning Microscopy (CLSM)
2.9. Statistical Analysis
3. Results and Discussion
3.1. Antimicrobial Activity
3.1.1. Minimum Inhibitory Concentration
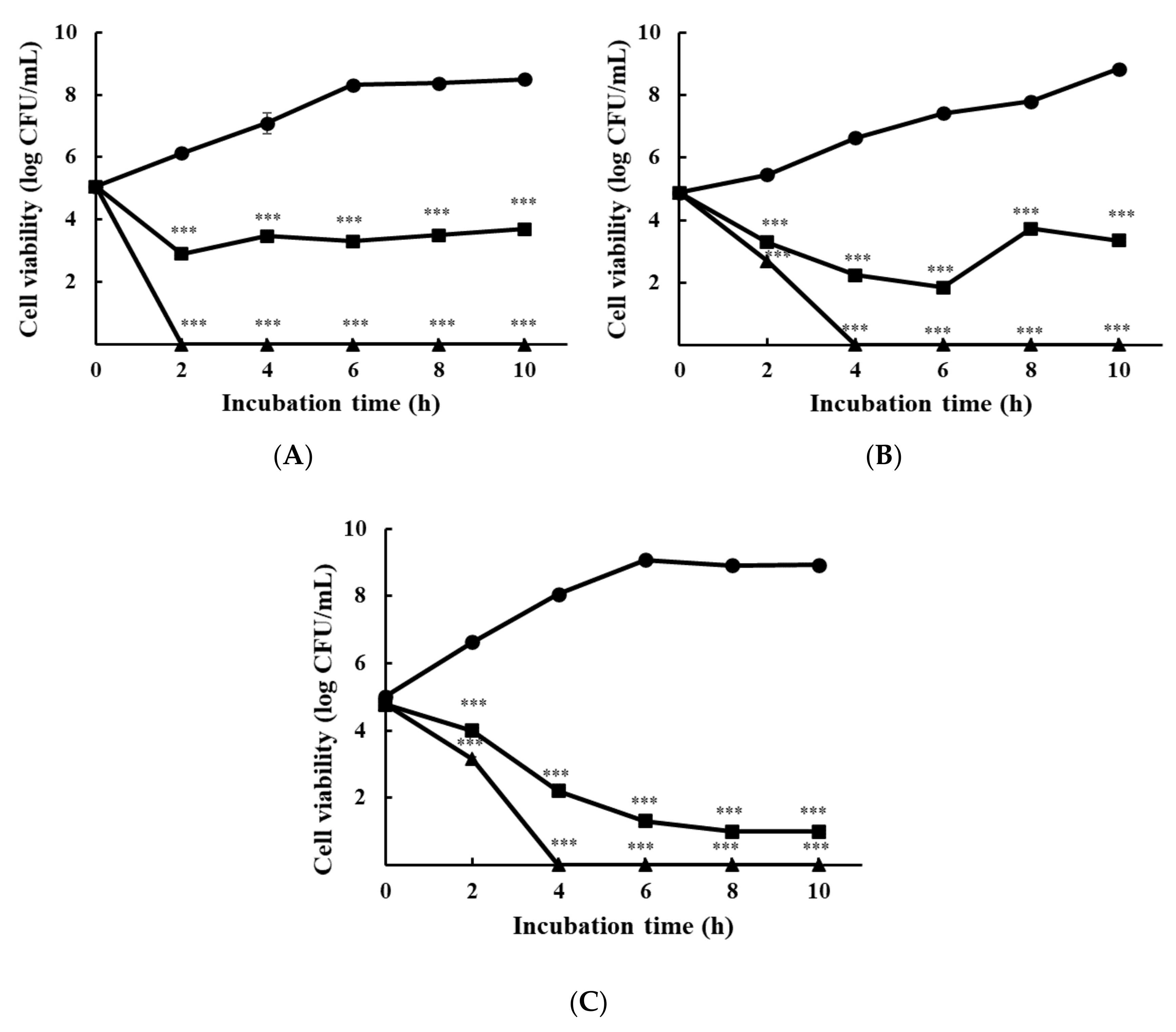
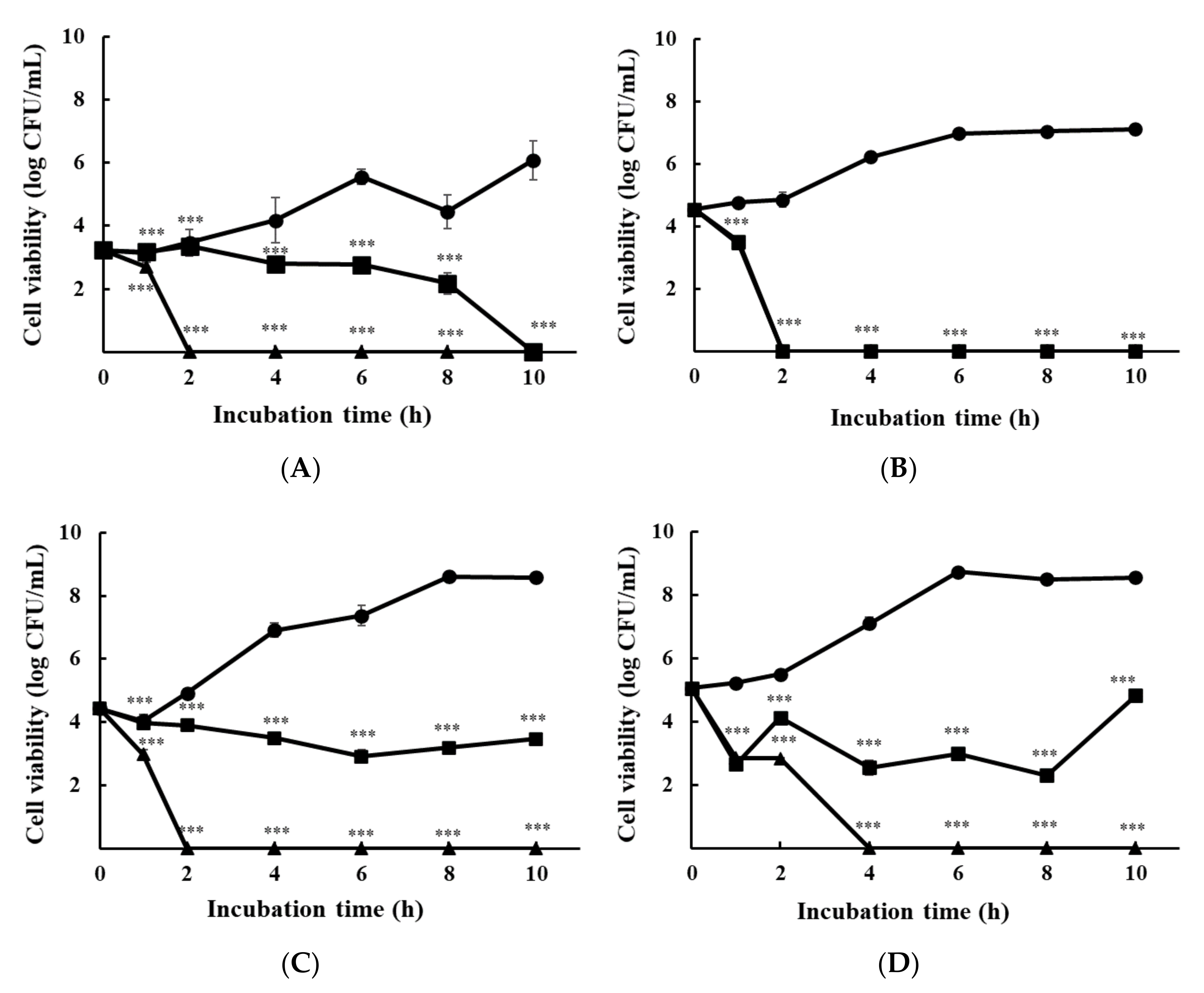
3.1.2. Time-Kill Analysis
3.2. Antibiofilm Effect
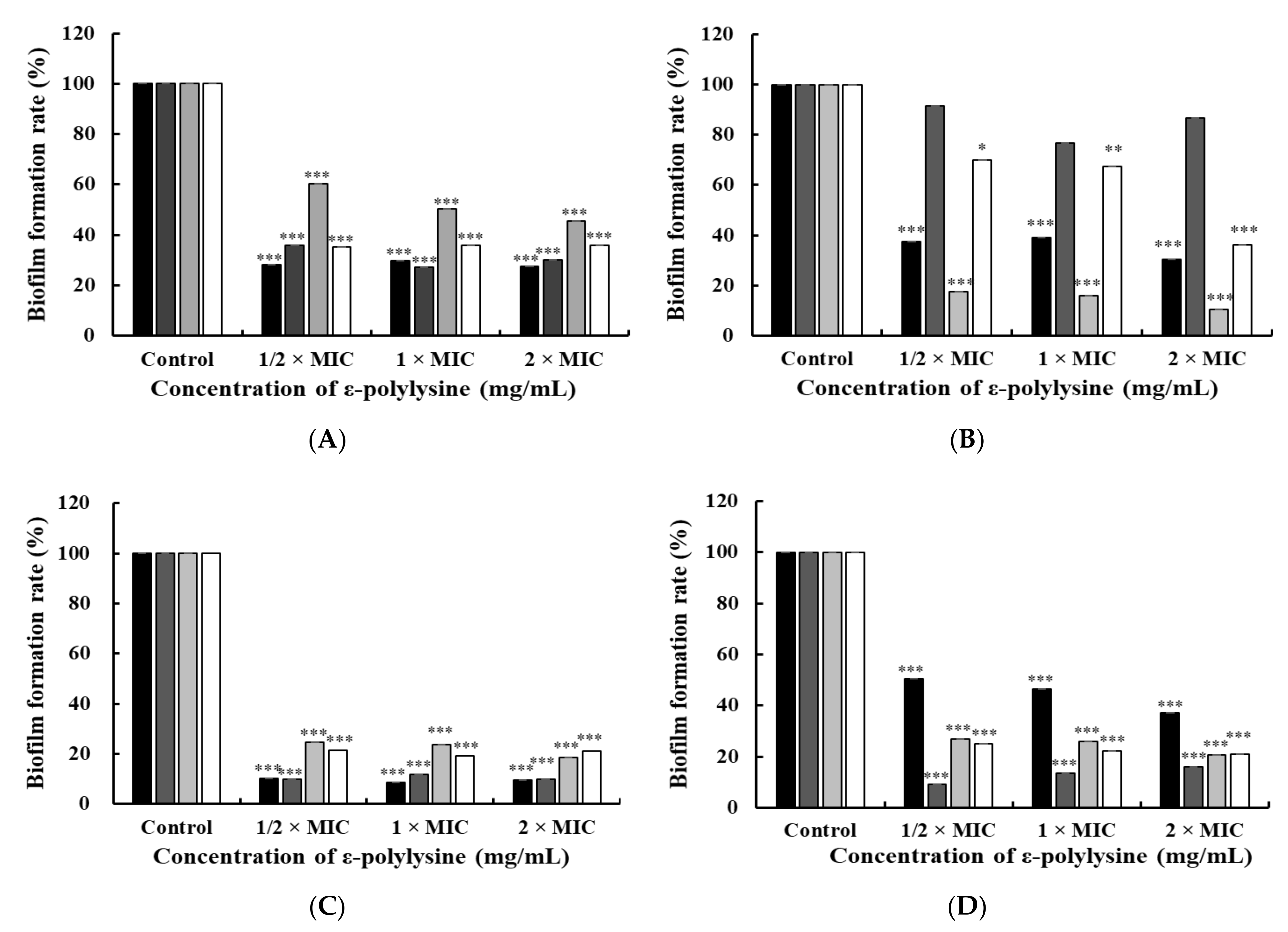
3.2.1. Crystal Violet Staining Assay
3.2.2. Bacterial Surface Properties and EPS Production
3.3. CLSM Observation
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Foodborne Germs and Illnesses. Available online: https://www.cdc.gov/foodsafety/foodborne-germs.html (accessed on 18 March 2020).
- Nerín, C.; Aznar, M.; Carrizo, D. Food contamination during food process. Trends Food Sci. Technol. 2016, 48, 63–68. [Google Scholar] [CrossRef]
- Food Safety. Available online: https://www.who.int/news-room/fact-sheets/detail/food-safety (accessed on 30 April 2020).
- Chicken and Food Poisoning. Available online: https://www.cdc.gov/foodsafety/chicken.html/ (accessed on 25 May 2020).
- McLauchlin, J.; Aird, H.; Amar, C.; Barker, C.; Dallman, T.; Elviss, N.; Jørgensen, F.; Willis, C. Listeria monocytogenes in cooked chicken: Detection of an outbreak in the United Kingdom (2016 to 2017) and analysis of L. monocytogenes from unrelated monitoring of foods (2013 to 2017). J. Food Protect. 2020, 83, 2041–2052. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Jiménez, D.; García-Meniño, I.; Fernández, J.; García, V.; Mora, A. Chicken and turkey meat: Consumer exposure to multidrug-resistant Enterobacteriaceae including mcr-carriers, uropathogenic E. coli and high-risk lineages such as ST131. Int. J. Food Microbiol. 2020, 331, 108750. [Google Scholar] [CrossRef] [PubMed]
- Chicken Meat Production Worldwide from 2012 to 2021 (in 1000 Metric Tons). Available online: https://www.statista.com/statistics/237637/production-of-poultry-meat-worldwide-since-1990/ (accessed on 20 April 2021).
- Projected Poultry Meat Consumption Worldwide from 2021 to 2030. Available online: https://www.statista.com/statistics/739951/poultry-meat-consumption-worldwide/ (accessed on 20 July 2021).
- Hu, W.S.; Nam, D.M.; Choi, J.Y.; Kim, J.S.; Koo, O.K. Anti-attachment, anti-biofilm, and antioxidant properties of Brassicaceae extracts on Escherichia coli O157:H7. Food Sci. Biotechnol. 2019, 28, 1881–1890. [Google Scholar] [CrossRef] [PubMed]
- Kim, U.; Kim, J.H.; Oh, S.W. Review of multi-species biofilm formation from foodborne pathogens multi-species biofilms and removal methodology. Crit. Rev. Food Sci. 2021, 1–11. [Google Scholar] [CrossRef]
- Bos, R.; Van der Mei, H.C.; Busscher, H.J. Physico-chemistry of initial microbial adhesive interactions—its mechanisms and methods for study. FEMS Microbiol. Rev. 1999, 23, 179–230. [Google Scholar] [CrossRef]
- Shen, C.; Islam, M.T.; Masuda, Y.; Honjoh, K.I.; Miyamoto, T. Transcriptional changes involved in inhibition of biofilm formation by ε-polylysine in Salmonella Typhimurium. Appl. Microbiol. Biotechnol. 2020, 104, 5427–5436. [Google Scholar] [CrossRef]
- Joshi, R.V.; Gunawan, C.; Mann, R. We are one: Multispecies metabolism of a biofilm consortium and their treatment strategies. Front. Microbiol. 2021, 12, 635432. [Google Scholar] [CrossRef]
- Burmølle, M.; Ren, D.; Bjarnsholt, T.; Sørensen, S.J. Interactions in multispecies biofilms: Do they actually matter? Trends Microbiol. 2014, 22, 84–91. [Google Scholar] [CrossRef]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Dodd, A.; Swanevelder, D.; Zhou, N.; Brady, D.; Hallsworth, J.E.; Rumbold, K. Streptomyces albulus yields ε-poly-l-lysine and other products from salt contaminated glycerol waste. J. Ind. Microbiol. Biotechnol. 2018, 45, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.C.; Singh, A.; Pandey, A.K.; Mishra, A. Review on production and medical applications of ε-polylysine. Biochem. Eng. J. 2012, 65, 70–81. [Google Scholar] [CrossRef]
- Wang, Y.; Hong, X.; Liu, J.; Zhu, J.; Chen, J. Interactions between fish isolates Pseudomonas fluorescens and Staphylococcus aureus in dual-species biofilms and sensitivity to carvacrol. Food Microbiol. 2020, 91, 103506. [Google Scholar] [CrossRef] [PubMed]
- Pontes, E.K.U.; Melo, H.M.; Nogueira, J.W.A.; Firmino, N.C.S.; Carvalho, M.G.; Catunda, F.E.A., Jr.; Cavalcante, T.T.A. Antibiofilm activity of the essential oil of citronella (Cymbopogon nardus) and its major component, geraniol, on the bacterial biofilms of Staphylococcus aureus. Food Sci. Biotechnol. 2019, 28, 633–639. [Google Scholar] [CrossRef]
- Bae, W.Y.; Kim, H.Y.; Kim, K.T.; Paik, H.D. Inhibitory effects of Inula britannica extract fermented by Lactobacillus plantarum KCCM 11613P on coagulase activity and growth of Staphylococcus aureus including methicillin-resistant strains. J. Food Biochem. 2019, 43, e12785. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.H.; Song, Y.J.; Yu, H.S.; Lee, N.K.; Paik, H.D. Investigating the antimicrobial and antibiofilm effects of cinnamaldehyde against Campylobacter spp. using cell surface characteristics. J. Food Sci. 2020, 85, 157–164. [Google Scholar] [CrossRef]
- Yang, S.J.; Lee, J.E.; Lim, S.M.; Kim, Y.J.; Lee, N.K.; Paik, H.D. Antioxidant and immune-enhancing effects of probiotic Lactobacillus plantarum 200655 isolated from kimchi. Food Sci. Biotechnol. 2019, 28, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Yu, H.H.; Song, Y.J.; Park, Y.J.; Lee, N.K.; Paik, H.D. Anti-biofilm effect of the cell-free supernatant of probiotic Saccharomyces cerevisiae against Listeria monocytogenes. Food Control 2021, 121, 107667. [Google Scholar] [CrossRef]
- Song, Y.J.; Yu, H.H.; Kim, Y.J.; Lee, N.-K.; Paik, H.D. Anti-Biofilm Activity of Grapefruit Seed Extract against Staphylococcus aureus and Escherichia coli. J. Microbiol. Biotechnol. 2019, 29, 1177–1183. [Google Scholar] [CrossRef]
- Liu, G.; Ren, G.; Zhao, L.; Cheng, L.; Wang, C.; Sun, B. Antibacterial activity and mechanism of bifidocin A against Listeria monocytogenes. Food Control 2017, 73, 854–861. [Google Scholar] [CrossRef]
- Guard-Petter, J. The chicken, the egg and Salmonella enteritidis. Environ. Microbiol. 2001, 3, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Čabarkapa, I.; Čolović, R.; Đuragić, O.; Popović, S.; Kokić, B.; Milanov, D.; Pezo, L. Anti-biofilm activities of essential oils rich in carvacrol and thymol against Salmonella Enteritidis. Biofouling 2019, 35, 361–375. [Google Scholar] [CrossRef]
- Rodríguez-Campos, D.; Rodríguez-Melcón, C.; Alonso-Calleja, C.; Capita, R. Persistent Listeria monocytogenes isolates from a poultry-processing facility form more biofilm but do not have a greater resistance to disinfectants than sporadic strains. Pathogens 2019, 8, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crecencio, R.B.; Brisola, M.C.; Bitner, D.; Frigo, A.; Rampazzo, L.; Borges, K.A.; Furian, T.Q.; Salle, C.T.P.; Moraes, H.L.S.; Fari, G.A.; et al. Antimicrobial susceptibility, biofilm formation and genetic profiles of Escherichia coli isolated from retail chicken meat. Infect. Genet. Evol. 2020, 84, 104355. [Google Scholar] [CrossRef]
- Shao, Z.; Yang, Y.; Fang, S.; Li, Y.; Chen, J.; Meng, Y. Mechanism of the antimicrobial activity of whey protein-ε-polylysine complexes against Escherichia coli and its application in sauced duck products. Int. J. Food Microbiol. 2020, 328, 108663. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Liao, X.; Surendhiran, D.; Cui, H. Preparation of ε-polylysine/chitosan nanofibers for food packaging against Salmonella on chicken. Food Packag. Shelf Life 2018, 17, 131–141. [Google Scholar] [CrossRef]
- Martínez-Ramos, A.R.; Ibarra-Sánchez, L.A.; Amaya-Llano, S.L.; Miller, M.J. Evaluation of combinations of nisin, lauric arginate, and ε-polylysine to control Listeria monocytogenes in queso fresco. J. Dairy Sci. 2020, 103, 11152–11162. [Google Scholar] [CrossRef]
- Tan, Z.; Shi, Y.; Xing, B.; Hou, Y.; Cui, J.; Jia, S. The antimicrobial effects and mechanism of ε-poly-lysine against Staphylococcus aureus. Bioresour. Bioprocess. 2019, 6, 1–10. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Vad, B.S.; Stenvang, M.; Otzen, D.E.; Meyer, R.L. The antimicrobial mechanism of action of epsilon-poly-l-lysine. Appl. Environ. Microbiol. 2014, 80, 7758–7770. [Google Scholar] [CrossRef] [Green Version]
- Ye, R.; Xu, H.; Wan, C.; Peng, S.; Wang, L.; Xu, H.; Aguilar, Z.P.; Xiong, Y.; Zeng, Z.; Wei, H. Antibacterial activity and mechanism of action of ε-poly-l-lysine. Biochem. Biophys. Res. Commun. 2013, 439, 148–153. [Google Scholar] [CrossRef]
- Lin, L.; Gu, Y.; Li, C.; Vittayapadung, S.; Cui, H. Antibacterial mechanism of ε-poly-lysine against Listeria monocytogenes and its application on cheese. Food Control 2018, 91, 76–84. [Google Scholar] [CrossRef]
- Wang, Y.; Samaranayake, L.P.; Dykes, G.A. Tea extracts modulate oral biofilm development by altering bacterial hydrophobicity and aggregation. Arch. Oral Biol. 2021, 122, 105032. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.; Luís, Â.; Oleastro, M.; Domingues, F.C. Antioxidant properties of coriander essential oil and linalool and their potential to control Campylobacter spp. Food Control 2016, 61, 115–122. [Google Scholar] [CrossRef]
- Santana, I.L.; Goncalves, L.M.; Vasconcellos, A.A.; Silva, W.J.; Cury, J.A.; Cury, A.A.D.B. Dietary carbohydrates modulate Candida albicans biofilm development on the denture surface. PLoS ONE 2013, 8, e64645. [Google Scholar]
- Islam, M.T.; Oishi, A.; Machida, C.; Ogura, A.; Kin, S.; Honjoh, K.I.; Miyamoto, T. Combined effects of selected food additives on adhesion of various foodborne pathogens onto microtiter plate and cabbage leaves. Food Control 2014, 46, 233–241. [Google Scholar] [CrossRef]
- Choi, N.Y.; Bae, Y.M.; Lee, S.Y. Cell surface properties and biofilm formation of pathogenic bacteria. Food Sci. Biotechnol. 2015, 24, 2257–2264. [Google Scholar] [CrossRef]
- Misba, L.; Zaidi, S.; Khan, A.U. A comparison of antibacterial and antibiofilm efficacy of phenothiazinium dyes between Gram positive and Gram negative bacterial biofilm. Photodiagn. Photodyn. 2017, 18, 24–33. [Google Scholar] [CrossRef]
- Olszewska, M.A.; Gędas, A.; Simões, M. The effects of eugenol, trans-cinnamaldehyde, citronellol, and terpineol on Escherichia coli biofilm control as assessed by culture-dependent and -independent methods. Molecules 2020, 25, 2641. [Google Scholar] [CrossRef] [PubMed]
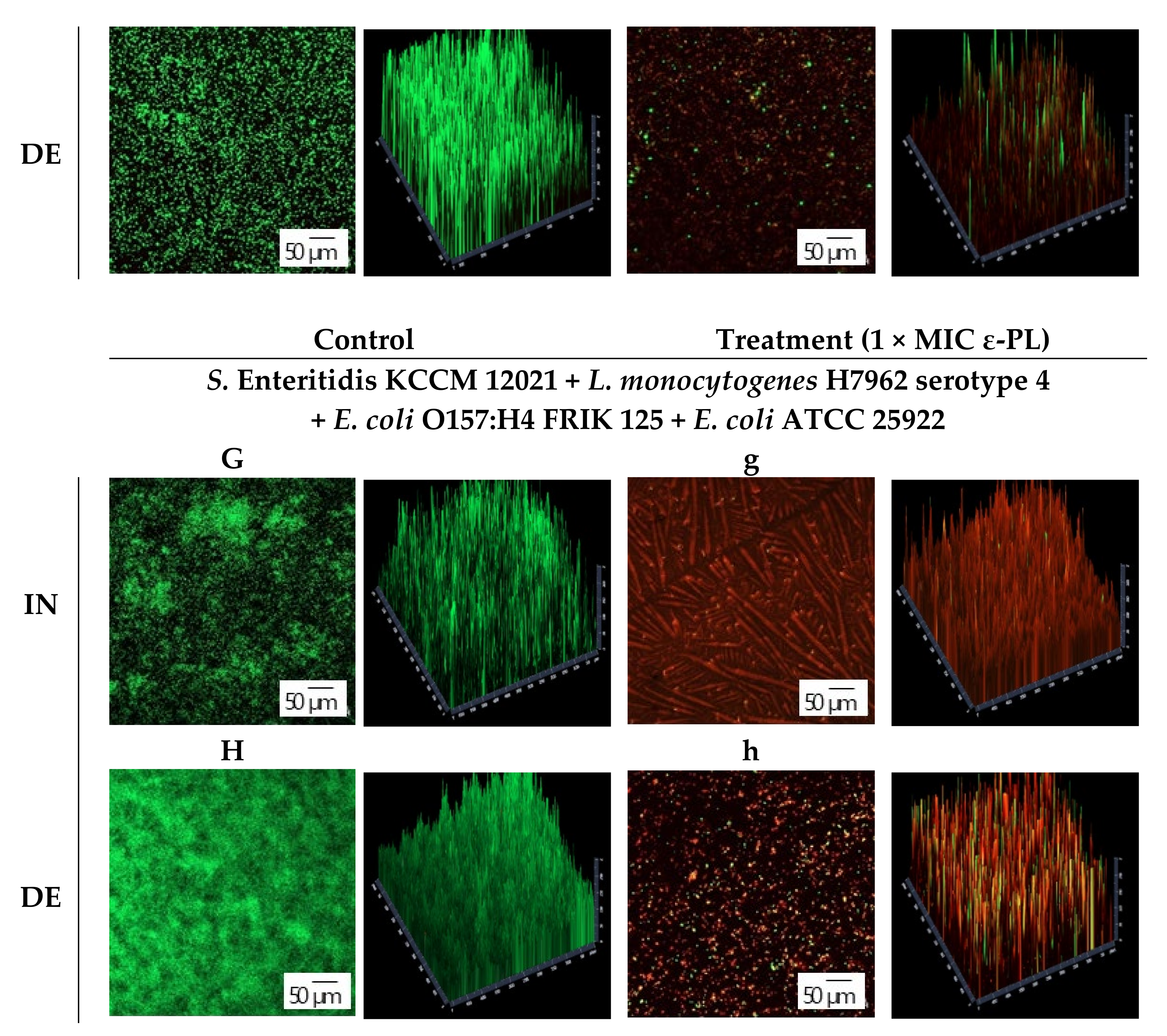

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial Number | Strains 1 | MIC Value (mg/mL) | |||
|---|---|---|---|---|---|
| Salmonella Enteritidis KCCM 12021 | Listeria monocytogenes H7962 Serotype 4 | Escherichia coli O157:H4 FRIK 125 | Escherichia coli ATCC 25922 | ||
| 1 | + | − | − | − | 1.0 |
| 2 | − | + | − | − | 0.031 |
| 3 | − | − | + | + | 1.0 |
| 4 | + | + | + | + | 1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.-U.; Park, Y.J.; Yu, H.H.; Jung, S.-C.; Park, J.-H.; Lee, D.-H.; Lee, N.-K.; Paik, H.-D. Antimicrobial and Antibiofilm Effect of ε-Polylysine against Salmonella Enteritidis, Listeria monocytogenes, and Escherichia coli in Tryptic Soy Broth and Chicken Juice. Foods 2021, 10, 2211. https://doi.org/10.3390/foods10092211
Lee D-U, Park YJ, Yu HH, Jung S-C, Park J-H, Lee D-H, Lee N-K, Paik H-D. Antimicrobial and Antibiofilm Effect of ε-Polylysine against Salmonella Enteritidis, Listeria monocytogenes, and Escherichia coli in Tryptic Soy Broth and Chicken Juice. Foods. 2021; 10(9):2211. https://doi.org/10.3390/foods10092211
Chicago/Turabian StyleLee, Do-Un, Yeong Jin Park, Hwan Hee Yu, Suk-Chae Jung, Jung-Hee Park, Dae-Hee Lee, Na-Kyoung Lee, and Hyun-Dong Paik. 2021. "Antimicrobial and Antibiofilm Effect of ε-Polylysine against Salmonella Enteritidis, Listeria monocytogenes, and Escherichia coli in Tryptic Soy Broth and Chicken Juice" Foods 10, no. 9: 2211. https://doi.org/10.3390/foods10092211
APA StyleLee, D.-U., Park, Y. J., Yu, H. H., Jung, S.-C., Park, J.-H., Lee, D.-H., Lee, N.-K., & Paik, H.-D. (2021). Antimicrobial and Antibiofilm Effect of ε-Polylysine against Salmonella Enteritidis, Listeria monocytogenes, and Escherichia coli in Tryptic Soy Broth and Chicken Juice. Foods, 10(9), 2211. https://doi.org/10.3390/foods10092211