
Mutation Associated with Orange Fruit Color Increases Concentrations of β-Carotene in a Sweet Pepper Variety (Capsicum annuum L.)


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Biochemical Evaluation of Certain Carotenoid Levels through a High-Performance Liquid Chromatography (HPLC) Method
2.2.1. Chemicals
2.2.2. Sample Extraction
2.2.3. Saponification
2.2.4. Chromatographic Separation
2.2.5. Preparation of Standard Carotenoid Solutions and Calibration
2.2.6. Analysis of the Variation
2.3. Molecular Characterization of β-Carotene Hydroxylases
2.3.1. DNA and RNA Isolation
2.3.2. Primers
2.3.3. Gene Cloning and Sequencing
2.3.4. Gene-Specific PCR
2.3.5. RT-PCR and cDNA Synthesis
2.3.6. Gene Cloning
2.3.7. DNA Electrophoresis, Ethidium Bromide Staining and Gel Documentation
3. Results
3.1. Biochemical Evaluation of Certain Carotenoid Levels through a High-Performance Liquid Chromatography
3.2. Molecular Characterization of the Mutation, Leading to Orange-Colored Fruits
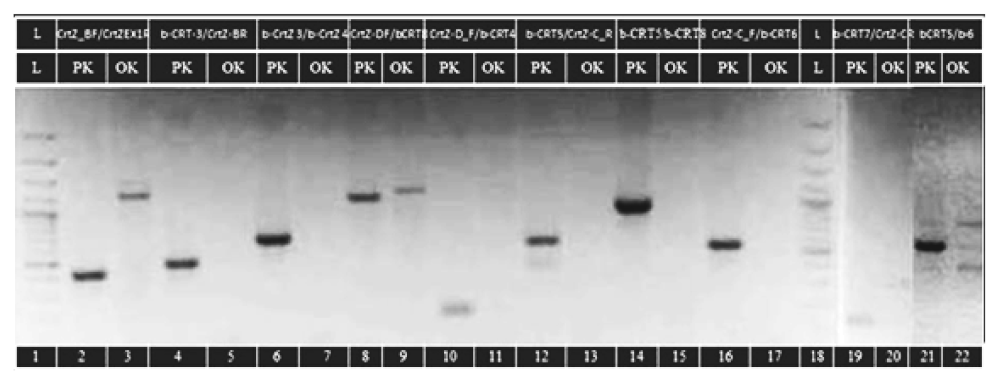
3.2.1. PCR amplifications of CrtZchr03 and CrtZchr06 gene
3.2.2. Cloning and Sequencing of Both β-Carotene Hydroxylase Genes
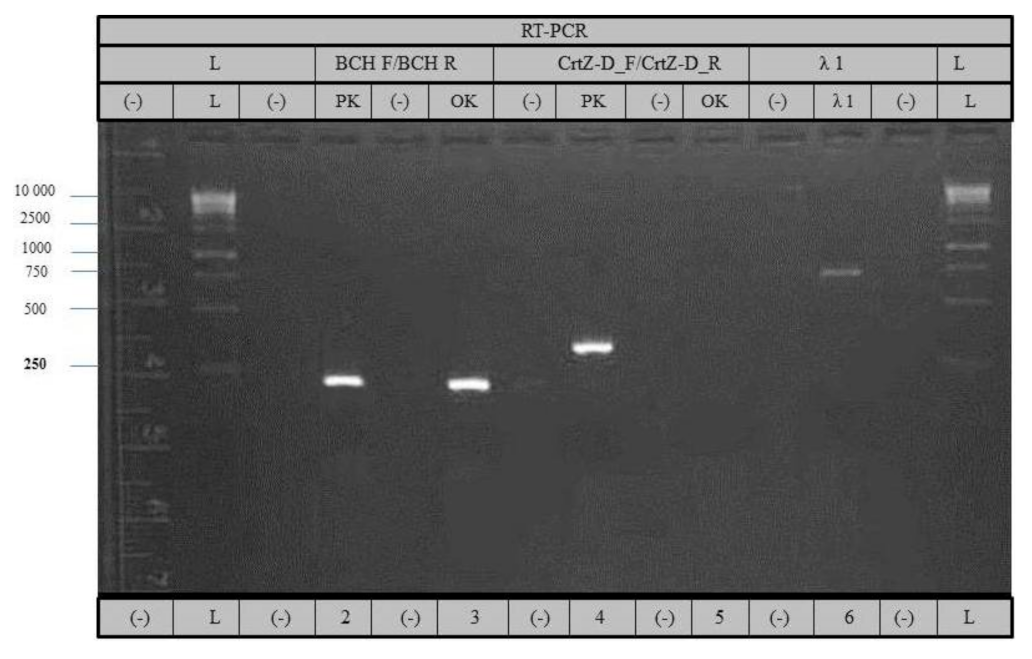
3.2.3. RT-PCR Analysis of Both (CrtZch03 and CrtZch06) β-Carotene Hydroxylase Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| AV | average value |
| BCX | β-crypthoxanthin |
| BHT | butyl hydroxytoluene |
| CCS | capsanthin/capsorubin syntase |
| CrtZchr0 | β-carotene hydroxylase on chromosomes 3 |
| CrtZchr0 | β-carotene hydroxylase on chromosomes 6 |
| HPLC | high performance liquid chromatography |
| I | Initial |
| Lcyb | lycopene-β-cyclase |
| M | mutant |
| NILs | near isogenic lines |
| PCR | polymerase chain reaction |
| PSY | phytoene synthase |
| RILs | recombinant inbred lines |
| TEA | triethylamine |
| Zea | zeaxanthin |
References
- Hurtado-Hernandez, H.; Smith, P.G. Inheritance of mature fruit color in Capsicum annuum L. J. Hered. 1985, 76, 211–213. [Google Scholar] [CrossRef]
- Kraft, K.H.; Brown, C.H.; Nabhan, G.P.; Luedeling, E.; Ruiz, J.D.J.L.; D’Eeckenbrugge, G.C.; Hijmans, R.J.; Gepts, P. Multiple lines of evidence for the origin of domesticated chili pepper, Capsicum annuum, in Mexico. Proc. Natl. Acad. Sci. USA 2014, 111, 6165–6170. [Google Scholar] [CrossRef]
- Bharath, S.M.; Cilas, C.; Umaharan, P. Fruit trait variation in a Caribbean germplasm collection of aromatic hot peppers (Capsicum chinense Jacq.). Hort. Sci. 2013, 48, 531–538. [Google Scholar] [CrossRef]
- Naegele, R.P.; Mitchell, J.; Hausbeck, M.K. Genetic diversity, population structure, and heritability of fruit traits in Capsicum annuum . PLoS ONE 2016, 11, e0156969. [Google Scholar] [CrossRef]
- Tomlekova, N.B. Induced mutagenesis for crop improvement in Bulgaria. Plant Mutat. Rep. 2010, 2, 1–32, invited paper. ISSN: 1011-260X. Available online: http://www.iaea.org/inis/collection/NCLCollectionStore/_Public/42/080/42080077.pdf?r=1 (accessed on 11 December 2020).
- Davison, P.A.; Hunter, C.N.; Horton, P. Overexpression of beta-carotene hydroxylase enhances stress tolerance in Arabidopsis. Nature 2002, 418, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Pogson, B.; McDonald, K.A.; Truong, M.; Britton, G.; DellaPenna, D. Arabidopsis carotenoid mutants demonstrate that lutein is not essential for photosynthesis in higher plants. Plant Cell 1996, 8, 1627–1639. [Google Scholar] [PubMed]
- Tian, L.; Magallanes-Lundback, M.; Musetti, V.; DellaPenna, D. Functional analysis of beta- and epsilon-ring carotenoid hydroxylases in Arabidopsis. Plant Cell 2003, 15, 1320–1332. [Google Scholar] [CrossRef] [PubMed]
- Bramley, P.M. Regulation of carotenoid formation during tomato fruit ripening and development. J. Exp. Bot. 2002, 53, 2107–2113. [Google Scholar] [CrossRef] [PubMed]
- Kalloo, G. Genetic improvement of tomato. Monogr. Theor. Appl. Gen. 1990, 14, 358. [Google Scholar]
- Kimura, M.; Rodriguez-Amaya, D.B. Carotenoid composition of hydrophobic leafy vegetables. J. Agric. Food Chem. 2003, 51, 2603–2607. [Google Scholar] [CrossRef]
- Goodwin, T.W. Plant Pigments; Academic Press: London, UK, 1988; p. 583. [Google Scholar]
- Daskalov, S. Mutation breeding in pepper. In Mutation Breeding Review Joint FAO/IAEA Division of Isotope and Radiation Applications of Atomic Energy for Food and Agricultural Development; IAEA: Vienna, Austria, 1986; Volume 22, pp. 1–26. [Google Scholar]
- Daskalov, S. Studies on Mutagenesis and Heterosis in Pepper (Capsicum annuum L.). Ph.D. Thesis, Institute of Genetics “D. Kostov”–BAS, Sofia, Bulgaria, 1987. (In Bulgarian). [Google Scholar]
- Tian, S.L.; Li, L.; Chai, W.G.; Shah, S.N.; Gong, Z.H. Effects of silencing key genes in the capsanthin biosynthetic pathway on fruit colour of detached pepper fruits. BMC Plant Biol. 2014, 14, 314. [Google Scholar] [CrossRef]
- Thorup, T.A.; Tanyolac, B.; Livingstone, K.D.; Popovsky, S.; Paran, I.; Jahn, M. Candidate gene analysis of organ pigmentation loci in the Solanaceae. Proc. Natl. Acad. Sci. USA 2000, 97, 11192–11197. [Google Scholar] [CrossRef]
- Lang, Y.-Q.; Yanagawa, S.; Sasanuma, T.; Sasakuma, T. Orange fruit colour in Capsicum due to deletion of capsanthin-capsorubin synthesis gene. Breed. Sci. 2004, 54, 33–39. [Google Scholar] [CrossRef]
- Huh, J.H.; Kang, B.C.; Nahm, S.H.; Kim, S.; Ha, K.S.; Lee, M.H.; Kim, B.D. A candidate gene approach identified phytoene synthase as the locus for mature fruit colour in red pepper (Capsicum spp.). Theor. Appl. Genet. 2001, 102, 524–530. [Google Scholar] [CrossRef]
- Pola, W.; Sugaya, S.; Photchanachai, S. Influence of postharvest temperatures on carotenoid biosynthesis and phytochemicals in mature green chili (Capsicum annuum L.). Antioxidants 2020, 9, 203. [Google Scholar] [CrossRef]
- Daskalov, S. Experimental mutagenesis and mutation breeding in pepper. Capsicum Newsl. 1991, 10, 13–20. [Google Scholar]
- Daskalov, S.; Baralieva, D. Oranzheva kapiya: A new cv. in sweet pepper with increased beta-carotene /provitamin A/ content. Mutat. Breed. Newsl. 1992, 39, 9–10. [Google Scholar]
- Daskalov, S.; Chalukova, M.; Baralieva, D.; Lukarska, E. Biochemical investigations of an induced β-orange mutant in a sweet pepper (Capsicum annuum L.) and developing varieties with increased β-carotene content. In Proceedings of the 9th Meeting on Genetics and Breeding on Capsicum and Eggplant, Budapest, Hungary, 21–25 August 1995; pp. 124–127. [Google Scholar]
- Descriptors for Capsicum (Capsicum spp.). International Plant Genetic Resources Institute (IPBGR). 1996. Available online: https://bioversityinternational.org/fileadmin/_migrated/uploads/tx_news/Descriptors_for_capsicum__Capsicum_spp__345.pdf (accessed on 10 December 2020).
- Tomlekova, N.; Atanassova, B.; Marinova, D.; Baralieva, D.; Ribarova, F. Study on the variability of lycopene and β-carotene content in tomato (Lycopersicon esculentum Mill.). Acta Hortic. 2007, 729, 101–104. [Google Scholar] [CrossRef]
- Tomlekova, N.; Panayotov, N.; Kozgar, M.I.; Wani, M.R.; Serdaris, P.; Balacheva, E. Inducing and exploring new mutant donors of tomato. In Mutagenesis: Exploring Genetic Diversity in Crops; Tomlekova, N.B., Kozgar, M.I., Wani, M.R., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2014; pp. 285–308. ISBN 978-90-8686-244-3. [Google Scholar] [CrossRef]
- Hart, D.; Scott, K. Development and evaluation of an HPLC method for the analysis of carotenoids in foods, and the measurement of the carotenoid concentration of vegetables and fruits commonly consumed in the UK. Food Chem. 1995, 54, 101–111. [Google Scholar] [CrossRef]
- SPSS Inc. SPSS Statistics for Windows, Version 17.0; SPSS Inc.: Chicago, IL, USA, 2008. [Google Scholar]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucl. Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgensen, R.A.; Allard, R.W. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA 1984, 81, 8014–8018. [Google Scholar] [CrossRef]
- Cho, S.; Mitchell, A.; Regier, J.C.; Mitter, C.; Poole, R.W.; Friedlander, T.P.; Zhao, S. A highly conserved nuclear gene for low-level phylogenetics: Elongation factor-1 alpha recovers morphology-based tree for heliothine moths. Mol. Biol. Evol. 1995, 12, 650–656. [Google Scholar]
- Kozera, B.; Rapacz, M. Reference genes in real-time PCR. J. Appl. Genet. 2013, 54, 391–406. [Google Scholar] [CrossRef]
- Bouvier, F.; Keller, Y.; D’Harlingue, A.; Camara, B. Xanthophyll biosynthesis: Molecular and functional characterization of carotenoid hydroxylases from pepper fruits (Capsicum annuum L.). Biochim. Biophys. Acta 1998, 1391, 320–328. [Google Scholar] [CrossRef]
- Guzman, I.; Hamby, S.; Romero, J.; Bosland, P.W.; O’Connell, M.A. Variability of carotenoid biosynthesis in orange coloured Capsicum spp. Plant Sci. 2010, 179, 49–59. [Google Scholar] [CrossRef]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed]
- Tomlekova, N.; Spasova-Apostolova, V.; Panchev, I. Caracterization of mutation affecting a region of the chromosome 3 in orange-coloured and beta-carotene-rich mutant registered as the variety Oranzheva kapiya. C. R. Acad. Bulg. Sci. 2016, 69, 145–150. [Google Scholar]
- Timina, O.; Timina, O.; Fedorov, S.; Tomlekova, N. Inheritance of pericarp colour and β-carotene content of sweet pepper. Vavilov J. Genet. Breed. 2011, 15, 585–594. [Google Scholar]
- Ha, J.-L.; Bae, J.-S.; Park, M.-K.; Kim, Y.-U.; Ha, S.-H.; Bae, J.-M.; Back, K.-W.; Lee, C.-H.; Lee, S.-W.; Ahn, M.-J. Quantitative analysis of carotenoids in carrot cultivars produced in Korea. J. Environ. Sci. Int. 2009, 18, 1135–1141. [Google Scholar] [CrossRef]
- Pacheco, S.; Peixoto, F.M.; Borguini, R.G.; Nascimento, L.D.S.D.M.D.; Bobeda, C.R.R.; Santiago, M.C.P.D.A.; Godoy, R.L.D.O. Microscale extraction method for HPLC carotenoid analysis in vegetable matrices. Sci. Agric. 2014, 71, 416–419. [Google Scholar] [CrossRef]
- Tomlekova, N. Use of mutant genes for exploring and altering carotenoid biosynthetic pathway of sweet pepper: The Bulgarian viewpoint. In Mutagenesis: Exploring Novel Genes and Pathways; Tomlekova, N.B., Kozgar, M.I., Wani, M.R., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2014; pp. 433–454. ISBN 978-90-8686-234-4. [Google Scholar] [CrossRef]
- Tomlekova, N.B.; White, P.J.; Thompson, J.A.; Penchev, E.A.; Nielen, S. Mutation increasing β-carotene concentrations does not adversely affect concentrations of essential mineral elements in pepper fruit. PLoS ONE 2017, 12, e0172180. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.X., Jr.; Gantt, E. Genes and enzymes of carotenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 557–583. [Google Scholar] [CrossRef] [PubMed]
- Gómez-García, M.; Ochoa-Alejo, N. Biochemistry and molecular biology of carotenoid biosynthesis in chili peppers (Capsicum spp.). Int. J. Mol. Sci. 2013, 14, 19025–19053. [Google Scholar] [CrossRef] [PubMed]
- Tomlekova, N.; Todorova, V.; Petkova, V.; Yancheva, S.; Nikolova, V.; Panchev, I.; Penchev, E. Creation and evaluation of induced mutants for pepper breeding programmes. In Proceedings of the FAO/IAEA International Symposium on Induced Mutations in Plants, Vienna, Austria, 12–15 August 2008; p. 187. [Google Scholar]
- Petrov, V.; Denev, I.; Draganov, M.; Timin, O.; Panchev, I.; Tomlekova, N. Molecular characterization of advanced mutants for early detection of high β-carotene concentration in pepper breeding programmes. C. R. Acad. Bulg. Sci. 2013, 66, 303–310. [Google Scholar]
- Borovsky, Y.; Tadmor, Y.; Bar, E.; Meir, A.; Lewinsohn, E.; Paran, I. Induced mutation in β-carotene hydroxylase results in accumulation of β-carotene and conversion of red to orange colour in pepper fruit. Theor. Appl. Genet. 2013, 126, 557–565. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Genetic Relationships Between Genotypes | Lutein | β-Carotene |
|---|---|---|---|
| Pazardzhishka kapia 794 | Initial | 1.22 ± 0.21 * | 0.32 ± 0.14 * |
| Oranzheva kapia | Mutant | 0.74 ± 0.16 * | 0.71 ± 0.23 * |
| Mutation Event | Alteration in M | 1.65× decreased | 2.25× increased |
| Genotypes | α-Carotene | Lutein | β-Carotene | BCX | Zea |
|---|---|---|---|---|---|
| Pazardzhishka kapia 794 | 0.09 ± 0.04 * | 11.52 ± 3.76 * | 3.76 ± 0.90 * | 4.07 ± 1.66 * | 18.46 ± 3.92 * |
| Oranzheva kapia | 0.67 ± 0.21 * | 1.37 ± 0.45 * | 7.31 ± 0.98 * | 0.000 | 2.67 ± 0.52 * |
| Mutation Event | 7.49× higher | 8.39× lower | 1.94× higher | Not detected in M | 6.90× lower |
| Primers for CrtZchr03 Gene | Amplified Regions on Exons and Introns of the Gene | Position of Primers in the Genomic Sequence | Expected Fragments Length [bp] | Results from Amplifications of Genotypes | |
|---|---|---|---|---|---|
| Initial | Mutant | ||||
| CrtZ-B_F/ CrtZ_R EX1 | E′1 + E′1 | 7/438 | 432 | 432 | 1300 |
| b-CRT 3 F/ CrtZ-B_R | I′1 + E2 + I2 + E′3 | 483/970 | 488 | 488 | 500 |
| b-CRT 3 F/ b-CRT 4 R | I′1 + E2 + I2 + E3 + I3 + E′4 | 483/1155 | 673 | 673 | 750 |
| CrtZ-D_F/ b-CRT 8 R | E′3 + I3 + E4 + I4 + E5 + I5 + E6 + I6 + E′7 | 966/2088 | 1123 | 1123 | 1300 |
| CrtZ-D_F/ b-CRT 4 | E′3 + I3 + E′4 | 966/1155 | 190 | 190 | 200 |
| b-CRT 5 F/ CrtZ-C_R | I′3 + E′4 + I4 + E5 + I5 + E′6 | 1127/1784 | 658 | 658 | 900 |
| b-CRT 5 F/ b-CRT 8 R | I′3 + E′4 + I4 + E5 + I5 + E6 + I6 + E′7 | 1127/2088 | 962 | 962 | 1250 |
| CrtZ-C_F/ b-CRT 6 R | E′4 + I4 + E5 + I′5 | 1163/1710 | 548 | 548 | 350 |
| b-CRT 7 F/ CrtZ-C_R | I′5 + E′6 | 1602/1784 | 183 | 183 | 400 |
| b-CRT 5 F/ b-CRT6 R | I′3 + E4 + I4 + E5 + I′5 | 1127/1710 | 584 | 584 | 750, 580, 450, 300 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomlekova, N.; Spasova-Apostolova, V.; Pantchev, I.; Sarsu, F. Mutation Associated with Orange Fruit Color Increases Concentrations of β-Carotene in a Sweet Pepper Variety (Capsicum annuum L.). Foods 2021, 10, 1225. https://doi.org/10.3390/foods10061225
Tomlekova N, Spasova-Apostolova V, Pantchev I, Sarsu F. Mutation Associated with Orange Fruit Color Increases Concentrations of β-Carotene in a Sweet Pepper Variety (Capsicum annuum L.). Foods. 2021; 10(6):1225. https://doi.org/10.3390/foods10061225
Chicago/Turabian StyleTomlekova, Nasya, Velichka Spasova-Apostolova, Ivelin Pantchev, and Fatma Sarsu. 2021. "Mutation Associated with Orange Fruit Color Increases Concentrations of β-Carotene in a Sweet Pepper Variety (Capsicum annuum L.)" Foods 10, no. 6: 1225. https://doi.org/10.3390/foods10061225
APA StyleTomlekova, N., Spasova-Apostolova, V., Pantchev, I., & Sarsu, F. (2021). Mutation Associated with Orange Fruit Color Increases Concentrations of β-Carotene in a Sweet Pepper Variety (Capsicum annuum L.). Foods, 10(6), 1225. https://doi.org/10.3390/foods10061225