
Carotenoids in Milk and the Potential for Dairy Based Functional Foods


Abstract
1. Introduction
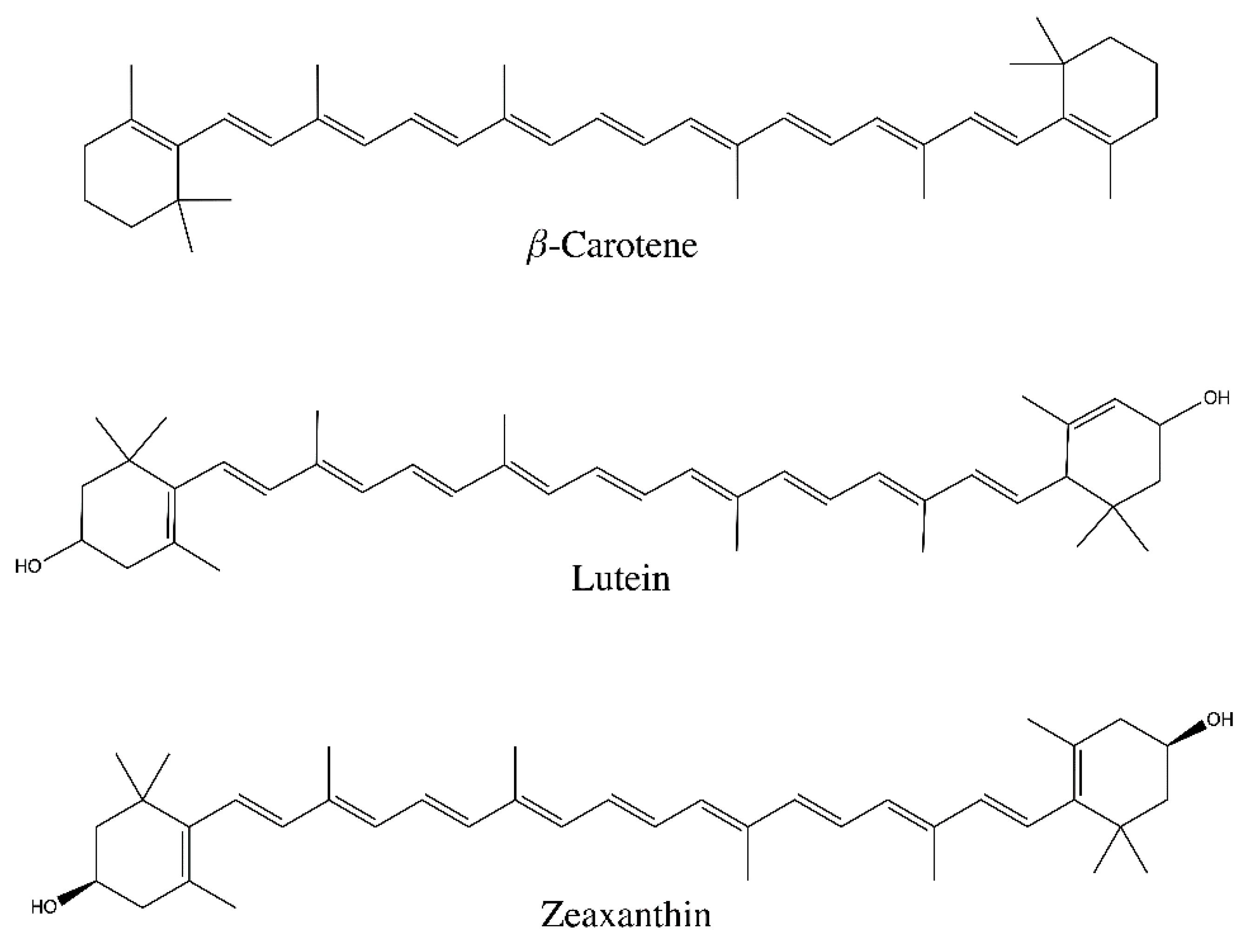
2. Carotenoids
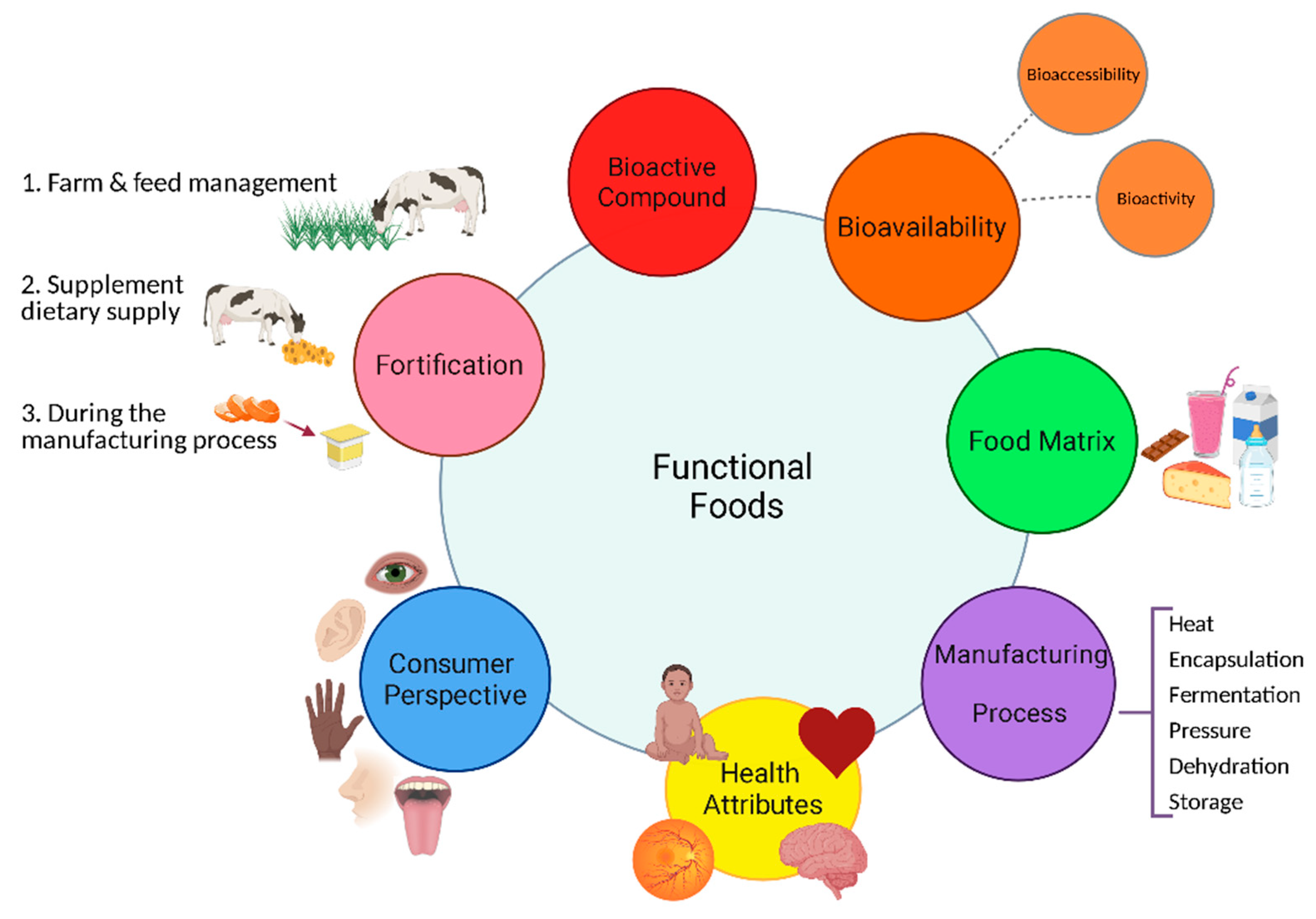
3. Functional Foods
3.1. Digestion, Absorption and Metabolism of Carotenoids
3.2. Dietary Factors Impacting Bioavailability
4. Bovine Milk
4.1. Carotenoids in Bovine Milk
4.2. The Health-Promoting and Sensorial Effects of Carotenoids in Bovine Milk
4.3. Factors Influencing the Carotenoid Content of Bovine Milk
4.3.1. Dietary Sources for Bovine Carotenoid Consumption
4.3.2. Feed Management
4.3.3. Grassland Management
4.3.4. Microbial Rumen Degradation
4.3.5. Saturation Phenomenon in the Carotenoids Transfer from Plasma to Milk
5. Dairy as a Functional Food
5.1. Bioaccessibility
5.2. Stability
5.3. Potential for Encapsulation in Dairy
5.4. Agri-Food Waste—A Future Source for Dairy Fortification?
5.5. The Effect of Processing on the Carotenoid Content and Bioaccessibility of Dairy Foods
6. Carotenoids in Human Milk and Infant Formula
6.1. Carotenoid Content of Human Milk
6.2. Carotenoid Content of Infant Formula
6.3. Fortified Infant Formula
6.3.1. Bioaccessibility and Bioavailability of Fortified Infant Formula
6.3.2. Safety and Efficacy
7. Future Considerations
Author Contributions
Funding
Conflicts of Interest
References
- O’Callaghan, T.F.; Sugrue, I.; Hill, C.; Ross, R.P.; Stanton, C. Nutritional aspects of raw milk: A beneficial or hazardous food choice. In Raw Milk; Elsevier: Amsterdam, The Netherlands, 2019; pp. 127–148. [Google Scholar]
- Mehta, B.M. Chemical composition of milk and milk products. In Handbook of Food Chemistry; Springer: Berlin/Heidelberg, Germany, 2015; pp. 511–553. [Google Scholar]
- Alothman, M.; Hogan, S.A.; Hennessy, D.; Dillon, P.; Kilcawley, K.N.; O’Donovan, M.; Tobin, J.; Fenelon, M.A.; O’Callaghan, T.F. The “grass-fed” milk story: Understanding the impact of pasture feeding on the composition and quality of bovine milk. Foods 2019, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- Agabriel, C.; Cornu, A.; Journal, C.; Sibra, C.; Grolier, P.; Martin, B. Tanker milk variability according to farm feeding practices: Vitamins A and E, carotenoids, color, and terpenoids. J. Dairy Sci. 2007, 90, 4884–4896. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, P.S.; Li, B.; Vachali, P.P.; Gorusupudi, A.; Shyam, R.; Henriksen, B.S.; Nolan, J.M. Lutein, zeaxanthin, and meso-zeaxanthin: The basic and clinical science underlying carotenoid-based nutritional interventions against ocular disease. Prog. Retin. Eye Res. 2016, 50, 34–66. [Google Scholar] [CrossRef] [PubMed]
- Feeney, J.; Finucane, C.; Savva, G.M.; Cronin, H.; Beatty, S.; Nolan, J.M.; Kenny, R.A. Low macular pigment optical density is associated with lower cognitive performance in a large, population-based sample of older adults. Neurobiol. Aging 2013, 34, 2449–2456. [Google Scholar] [CrossRef]
- Olson, J.A. Provitamin A function of carotenoids: The conversion of β-carotene into vitamin A. J. Nutr. 1989, 119, 105–108. [Google Scholar] [CrossRef]
- Hasler, C.M. Functional foods: Benefits, concerns and challenges—a position paper from the American Council on Science and Health. J. Nutr. 2002, 132, 3772–3781. [Google Scholar] [CrossRef]
- Howlett, J. Functional Foods: From Science to Health and Claims; ILSI Europe: Brussels, Belgium, 2008. [Google Scholar]
- Department of Health. Your Guide to Healthy Eating Using the Food Pyramid. Available online: https://www.hse.ie/eng/about/who/healthwellbeing/our-priority-programmes/heal/heal-docs/food-pyramid-poster-advice-version.pdf (accessed on 5 March 2021).
- Kopec, R.E.; Failla, M.L. Recent advances in the bioaccessibility and bioavailability of carotenoids and effects of other dietary lipophiles. J. Food Compos. Anal. 2018, 68, 16–30. [Google Scholar] [CrossRef]
- Granato, D.; Barba, F.J.; Bursać Kovačević, D.; Lorenzo, J.M.; Cruz, A.G.; Putnik, P. Functional foods: Product development, technological trends, efficacy testing, and safety. Annu. Rev. Food Sci. Technol. 2020, 11, 93–118. [Google Scholar] [CrossRef]
- Rubin, L.P.; Chan, G.M.; Barrett-Reis, B.M.; Fulton, A.B.; Hansen, R.; Ashmeade, T.; Oliver, J.; Mackey, A.; Dimmit, R.; Hartmann, E. Effect of carotenoid supplementation on plasma carotenoids, inflammation and visual development in preterm infants. J. Perinatol. 2012, 32, 418–424. [Google Scholar] [CrossRef]
- Bettler, J.; Zimmer, J.P.; Neuringer, M.; DeRusso, P.A. Serum lutein concentrations in healthy term infants fed human milk or infant formula with lutein. Eur. J. Nutr. 2010, 49, 45–51. [Google Scholar] [CrossRef]
- Yabuzaki, J. Carotenoids Database: Structures, chemical fingerprints and distribution among organisms. Database 2017, 2017, bax004. [Google Scholar] [CrossRef] [PubMed]
- Kalač, P. Carotenoids, ergosterol and tocopherols in fresh and preserved herbage and their transfer to bovine milk fat and adipose tissues: A review. J. Agrobiol. 2012, 29, 1–13. [Google Scholar] [CrossRef]
- Ucci, M.; Di Tomo, P.; Tritschler, F.; Cordone, V.G.; Lanuti, P.; Bologna, G.; Di Silvestre, S.; Di Pietro, N.; Pipino, C.; Mandatori, D. Anti-inflammatory role of carotenoids in endothelial cells derived from umbilical cord of women affected by gestational diabetes mellitus. Oxidative Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Terao, J.; Minami, Y.; Bando, N. Singlet molecular oxygen-quenching activity of carotenoids: Relevance to protection of the skin from photoaging. J. Clin. Biochem. Nutr. 2010, 48, 57–62. [Google Scholar] [CrossRef]
- Kawata, A.; Murakami, Y.; Suzuki, S.; Fujisawa, S. Anti-inflammatory activity of β-carotene, lycopene and tri-n-butylborane, a scavenger of reactive oxygen species. In Vivo 2018, 32, 255–264. [Google Scholar] [PubMed]
- Lin, K.-H.; Lin, K.-C.; Lu, W.-J.; Thomas, P.-A.; Jayakumar, T.; Sheu, J.-R. Astaxanthin, a carotenoid, stimulates immune responses by enhancing IFN-γ and IL-2 secretion in primary cultured lymphocytes in vitro and ex vivo. Int. J. Mol. Sci. 2016, 17, 44. [Google Scholar] [CrossRef] [PubMed]
- Toti, E.; Chen, C.-Y.O.; Palmery, M.; Villaño Valencia, D.; Peluso, I. Non-provitamin A and provitamin A carotenoids as immunomodulators: Recommended dietary allowance, therapeutic index, or personalized nutrition? Oxidative Med. Cell. Longev. 2018, 2018, 4637861. [Google Scholar] [CrossRef]
- Gurmu, F.; Hussein, S.; Laing, M. The potential of orange-fleshed sweet potato to prevent vitamin A deficiency in Africa. Int. J. Vitam. Nutr. Res. 2014, 84, 65–78. [Google Scholar] [CrossRef]
- Barker, F.M.; Snodderly, D.M.; Johnson, E.J.; Schalch, W.; Koepcke, W.; Gerss, J.; Neuringer, M. Nutritional manipulation of primate retinas, V: Effects of lutein, zeaxanthin, and n–3 fatty acids on retinal sensitivity to blue-light–induced damage. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3934–3942. [Google Scholar] [CrossRef]
- Seddon, J.; Ajani, U.; Sperduto, R.; Hiller, R.; Blair, N.; Burton, T.; Farber, M.; Gragoudas, E.; Haller, J.; Miller, D. Dietary carotenoids, vitamin-A, vitamin-C, and vitamin-E, and advanced age-related macular degeneration. JAMA-J. Am. Med. Assoc. 1994, 272, 1413–1420. [Google Scholar] [CrossRef]
- Craft, N.; Haitema, T.; Garnett, K.; Fitch, K.; Dorey, C. Carotenoid, tocopherol, and retinol concentrations in elderly human brain. Exp. Anim. 2004, 21, 22. [Google Scholar]
- Johnson, E.J.; Vishwanathan, R.; Johnson, M.A.; Hausman, D.B.; Davey, A.; Scott, T.M.; Green, R.C.; Miller, L.S.; Gearing, M.; Woodard, J. Relationship between serum and brain carotenoids,-tocopherol, and retinol concentrations and cognitive performance in the oldest old from the Georgia Centenarian Study. J. Aging Res. 2013, 2013, 951786. [Google Scholar] [CrossRef] [PubMed]
- Vishwanathan, R.; Iannaccone, A.; Scott, T.M.; Kritchevsky, S.B.; Jennings, B.J.; Carboni, G.; Forma, G.; Satterfield, S.; Harris, T.; Johnson, K.C. Macular pigment optical density is related to cognitive function in older people. Age Ageing 2014, 43, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Nolan, J.M.; Loskutova, E.; Howard, A.N.; Moran, R.; Mulcahy, R.; Stack, J.; Bolger, M.; Dennison, J.; Akuffo, K.O.; Owens, N. Macular pigment, visual function, and macular disease among subjects with Alzheimer’s disease: An exploratory study. J. Alzheimer’s Dis. 2014, 42, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- Khachik, F. Distribution and metabolism of dietary carotenoids in humans as a criterion for development of nutritional supplements. Pure Appl. Chem. 2006, 78, 1551–1557. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B.; Kimura, M.; Godoy, H.T.; Amaya-Farfan, J. Updated Brazilian database on food carotenoids: Factors affecting carotenoid composition. J. Food Compos. Anal. 2008, 21, 445–463. [Google Scholar] [CrossRef]
- Biehler, E.; Alkerwi, A.A.; Hoffmann, L.; Krause, E.; Guillaume, M.; Lair, M.-L.; Bohn, T. Contribution of violaxanthin, neoxanthin, phytoene and phytofluene to total carotenoid intake: Assessment in Luxembourg. J. Food Compos. Anal. 2012, 25, 56–65. [Google Scholar] [CrossRef]
- Perry, A.; Rasmussen, H.; Johnson, E.J. Xanthophyll (lutein, zeaxanthin) content in fruits, vegetables and corn and egg products. J. Food Compos. Anal. 2009, 22, 9–15. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Böhm, V.; Borge, G.I.A.; Cano, M.P.; Fikselová, M.; Gruskiene, R.; Lavelli, V.; Loizzo, M.R.; Mandić, A.I.; Brahm, P.M. Carotenoids: Considerations for Their Use in Functional Foods, Nutraceuticals, Nutricosmetics, Supplements, Botanicals, and Novel Foods in the Context of Sustainability, Circular Economy, and Climate Change. Annu. Rev. Food Sci. Technol. 2021, 12, 433–460. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Mandić, A.I.; Bantis, F.; Böhm, V.; Borge, G.I.A.; Brnčić, M.; Bysted, A.; Cano, M.P.; Dias, M.G.; Elgersma, A. A comprehensive review on carotenoids in foods and feeds: Status quo, applications, patents, and research needs. Crit. Rev. Food Sci. Nutr. 2020, 2020, 1867959. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Mapelli-Brahm, P.; Stinco, C.M. The colourless carotenoids phytoene and phytofluene: From dietary sources to their usefulness for the functional foods and nutricosmetics industries. J. Food Compos. Anal. 2018, 67, 91–103. [Google Scholar] [CrossRef]
- Ollilainen, V.; Heinonen, M.; Linkola, E.; Varo, P.; Koivistoinen, P. Carotenoids and retinoids in Finnish foods: Dairy products and eggs. J. Dairy Sci. 1989, 72, 2257–2265. [Google Scholar] [CrossRef]
- Fernández-García, E.; Carvajal-Lérida, I.; Pérez-Gálvez, A. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res. 2009, 29, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Lakshminarayana, R.; Baskaran, V. Influence of olive oil on the bioavailability of carotenoids. Eur. J. Lipid Sci. Technol. 2013, 115, 1085–1093. [Google Scholar] [CrossRef]
- Xavier, A.A.O.; Mercadante, A.Z. The bioaccessibility of carotenoids impacts the design of functional foods. Curr. Opin. Food Sci. 2019, 26, 1–8. [Google Scholar] [CrossRef]
- Reboul, E. Absorption of vitamin A and carotenoids by the enterocyte: Focus on transport proteins. Nutrients 2013, 5, 3563–3581. [Google Scholar] [CrossRef]
- Reboul, E. Mechanisms of carotenoid intestinal absorption: Where do we stand? Nutrients 2019, 11, 838. [Google Scholar] [CrossRef]
- Borel, P.; Lietz, G.; Goncalves, A.; Szabo de Edelenyi, F.; Lecompte, S.; Curtis, P.; Goumidi, L.; Caslake, M.J.; Miles, E.A.; Packard, C. CD36 and SR-BI are involved in cellular uptake of provitamin A carotenoids by Caco-2 and HEK cells, and some of their genetic variants are associated with plasma concentrations of these micronutrients in humans. J. Nutr. 2013, 143, 448–456. [Google Scholar] [CrossRef]
- During, A.; Dawson, H.D.; Harrison, E.H. Carotenoid transport is decreased and expression of the lipid transporters SR-BI, NPC1L1, and ABCA1 is downregulated in Caco-2 cells treated with ezetimibe. J. Nutr. 2005, 135, 2305–2312. [Google Scholar] [CrossRef]
- During, A.; Doraiswamy, S.; Harrison, E.H. Xanthophylls are preferentially taken up compared with β-carotene by retinal cells via a SRBI-dependent mechanism. J. Lipid Res. 2008, 49, 1715–1724. [Google Scholar] [CrossRef]
- Sato, Y.; Suzuki, R.; Kobayashi, M.; Itagaki, S.; Hirano, T.; Noda, T.; Mizuno, S.; Sugawara, M.; Iseki, K. Involvement of cholesterol membrane transporter Niemann-Pick C1-like 1 in the intestinal absorption of lutein. J. Pharm. Pharm. Sci. 2012, 15, 256–264. [Google Scholar] [CrossRef]
- Moussa, M.; Gouranton, E.; Gleize, B.; Yazidi, C.E.; Niot, I.; Besnard, P.; Borel, P.; Landrier, J.F. CD36 is involved in lycopene and lutein uptake by adipocytes and adipose tissue cultures. Mol. Nutr. Food Res. 2011, 55, 578–584. [Google Scholar] [CrossRef]
- Reboul, E.; Abou, L.; Mikail, C.; Ghiringhelli, O.; André, M.; Portugal, H.; Jourdheuil-Rahmani, D.; Amiot, M.-J.; Lairon, D.; Borel, P. Lutein transport by Caco-2 TC-7 cells occurs partly by a facilitated process involving the scavenger receptor class B type I (SR-BI). Biochem. J. 2005, 387, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Mapelli-Brahm, P.; Desmarchelier, C.; Margier, M.; Reboul, E.; Meléndez Martínez, A.J.; Borel, P. Phytoene and phytofluene isolated from a tomato extract are readily incorporated in mixed micelles and absorbed by Caco-2 cells, as compared to lycopene, and SR-BI is involved in their cellular uptake. Mol. Nutr. Food Res. 2018, 62, 1800703. [Google Scholar] [CrossRef] [PubMed]
- Moussa, M.; Landrier, J.-F.; Reboul, E.; Ghiringhelli, O.; Coméra, C.; Collet, X.; Frohlich, K.; Bohm, V.; Borel, P. Lycopene absorption in human intestinal cells and in mice involves scavenger receptor class B type I but not Niemann-Pick C1-like 1. J. Nutr. 2008, 138, 1432–1436. [Google Scholar] [CrossRef] [PubMed]
- Harrison, E.H. Mechanisms involved in the intestinal absorption of dietary vitamin A and provitamin A carotenoids. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2012, 1821, 70–77. [Google Scholar] [CrossRef] [PubMed]
- . Shilpa, S.; Shwetha, H.J.; Raju, M.; Lakshminarayana, R. Factors affecting bioaccessibility and bio-efficacy of carotenoids. In Carotenoids: Properties, Processing and Applications; Elsevier: Amsterdam, The Netherlands, 2020; pp. 41–73. [Google Scholar]
- Brown, M.J.; Ferruzzi, M.G.; Nguyen, M.L.; Cooper, D.A.; Eldridge, A.L.; Schwartz, S.J.; White, W.S. Carotenoid bioavailability is higher from salads ingested with full-fat than with fat-reduced salad dressings as measured with electrochemical detection. Am. J. Clin. Nutr. 2004, 80, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Unlu, N.Z.; Bohn, T.; Clinton, S.K.; Schwartz, S.J. Carotenoid absorption from salad and salsa by humans is enhanced by the addition of avocado or avocado oil. J. Nutr. 2005, 135, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Early, R. Dairy products and milk-based food ingredients. In Natural Food Additives, Ingredients and Flavourings; Elsevier: Amsterdam, The Netherlands, 2012; pp. 417–445. [Google Scholar]
- Mashurabad, P.C.; Palika, R.; Jyrwa, Y.W.; Bhaskarachary, K.; Pullakhandam, R. Dietary fat composition, food matrix and relative polarity modulate the micellarization and intestinal uptake of carotenoids from vegetables and fruits. J. Food Sci. Technol. 2017, 54, 333–341. [Google Scholar] [CrossRef]
- Clark, R.M.; Yao, L.; She, L.; Furr, H.C. A comparison of lycopene and astaxanthin absorption from corn oil and olive oil emulsions. Lipids 2000, 35, 803–806. [Google Scholar] [CrossRef]
- Borel, P.; Tyssandier, V.; Mekki, N.; Grolier, P.; Rochette, Y.; Alexandre-Gouabau, M.C.; Lairon, D.; Azaıs-Braesco, V. Chylomicron β-carotene and retinyl palmitate responses are dramatically diminished when men ingest β-carotene with medium-chain rather than long-chain triglycerides. J. Nutr. 1998, 128, 1361–1367. [Google Scholar] [CrossRef]
- Yuan, X.; Liu, X.; McClements, D.J.; Cao, Y.; Xiao, H. Enhancement of phytochemical bioaccessibility from plant-based foods using excipient emulsions: Impact of lipid type on carotenoid solubilization from spinach. Food Funct. 2018, 9, 4352–4365. [Google Scholar] [CrossRef]
- Calderón, F.; Chauveau-Duriot, B.; Pradel, P.; Martin, B.; Graulet, B.; Doreau, M.; Nozière, P. Variations in carotenoids, vitamins A and E, and color in cow’s plasma and milk following a shift from hay diet to diets containing increasing levels of carotenoids and vitamin E. J. Dairy Sci. 2007, 90, 5651–5664. [Google Scholar] [CrossRef]
- Calderón, F.; Tornambé, G.; Martin, B.; Pradel, P.; Chauveau-Duriot, B.; Nozière, P. Effects of mountain grassland maturity stage and grazing management on carotenoids in sward and cow’s milk. Anim. Res. 2006, 55, 533–544. [Google Scholar] [CrossRef]
- Hulshof, P.J.; van Roekel-Jansen, T.; van de Bovenkamp, P.; West, C.E. Variation in retinol and carotenoid content of milk and milk products in The Netherlands. J. Food Compos. Anal. 2006, 19, 67–75. [Google Scholar] [CrossRef]
- Stergiadis, S.; Leifert, C.; Seal, C.; Eyre, M.; Larsen, M.K.; Slots, T.; Nielsen, J.; Butler, G. A 2-year study on milk quality from three pasture-based dairy systems of contrasting production intensities in Wales. J. Agric. Sci. 2015, 153, 708. [Google Scholar] [CrossRef]
- Michal, J.; Heirman, L.; Wong, T.; Chew, B.; Frigg, M.; Volker, L. Modulatory effects of dietary β-carotene on blood and mammary leukocyte function in periparturient dairy cows. J. Dairy Sci. 1994, 77, 1408–1421. [Google Scholar] [CrossRef]
- Ascarelli, I.; Edelman, Z.; Rosenberg, M.; Folman, Y. Effect of dietary carotene on fertility of high-yielding dairy cows. Anim. Sci. 1985, 40, 195–207. [Google Scholar] [CrossRef]
- Chew, B.; Hollen, L.; Hillers, J.; Herlugson, M. Relationship between vitamin A and β-carotene in blood plasma and milk and mastitis in Holsteins. J. Dairy Sci. 1982, 65, 2111–2118. [Google Scholar] [CrossRef]
- Prache, S.; Priolo, A.; Tournadre, H.; Jailler, R.; Dubroeucq, H.; Micol, D.; Martin, B. Traceability of grass-feeding by quantifying the signature of carotenoid pigments in herbivores meat, milk and cheese. In Proceedings of the 19th General Meeting of the European Grassland Federation, La Rochelle, France, 27–30 May 2002; pp. 592–593. [Google Scholar]
- Barrefors, P.; Granelli, K.; Appelqvist, L.-A.; Bjoerck, L. Chemical characterization of raw milk samples with and without oxidative off-flavor. J. Dairy Sci. 1995, 78, 2691–2699. [Google Scholar] [CrossRef]
- Fakourelis, N.; Lee, E.; Min, D. Effects of chlorophyll and β-carotene on the oxidation stability of olive oil. J. Food Sci. 1987, 52, 234–235. [Google Scholar] [CrossRef]
- Sattar, A.; Deman, J.; Alexander, J. Stability of edible oils and fats to fluorescent light irradiation. J. Am. Oil Chem. Soc. 1976, 53, 473–477. [Google Scholar] [CrossRef]
- Havemose, M.; Weisbjerg, M.R.; Bredie, W.L.; Nielsen, J. Influence of feeding different types of roughage on the oxidative stability of milk. Int. Dairy J. 2004, 14, 563–570. [Google Scholar] [CrossRef]
- Nozière, P.; Graulet, B.; Lucas, A.; Martin, B.; Grolier, P.; Doreau, M. Carotenoids for ruminants: From forages to dairy products. Anim. Feed Sci. Technol. 2006, 131, 418–450. [Google Scholar] [CrossRef]
- Morris, C.; Knight, T.; Newman, S.A.; Hickey, S.; Death, A.; O’Neill, K.; Ridland, M. Genetic studies of carotenoid concentration in the plasma and milk of New Zealand dairy cattle. N. Z. J. Agric. Res. 2002, 45, 27–33. [Google Scholar] [CrossRef]
- Nozière, P.; Grolier, P.; Durand, D.; Ferlay, A.; Pradel, P.; Martin, B. Variations in carotenoids, fat-soluble micronutrients, and color in cows’ plasma and milk following changes in forage and feeding level. J. Dairy Sci. 2006, 89, 2634–2648. [Google Scholar] [CrossRef]
- Larsen, M.K.; Fretté, X.C.; Kristensen, T.; Eriksen, J.; Søegaard, K.; Nielsen, J.H. Fatty acid, tocopherol and carotenoid content in herbage and milk affected by sward composition and season of grazing. J. Sci. Food Agric. 2012, 92, 2891–2898. [Google Scholar] [CrossRef] [PubMed]
- Cardinault, N.; Doreau, M.; Poncet, C.; Nozière, P. Digestion and absorption of carotenoids in sheep given fresh red clover. Anim. Sci. 2006, 82, 49–55. [Google Scholar] [CrossRef]
- Livingston, A.; Smith, D.; Carnahan, H.; Knowles, R.; Nelson, J.; Kohler, G. Variation in the xanthophyll and carotene content of lucerne, clovers and grasses. J. Sci. Food Agric. 1968, 19, 632–636. [Google Scholar] [CrossRef]
- Elgersma, A.; Søegaard, K.; Jensen, S.K. Fatty acids, α-tocopherol, β-carotene, and lutein contents in forage legumes, forbs, and a grass–clover mixture. J. Agric. Food Chem. 2013, 61, 11913–11920. [Google Scholar] [CrossRef]
- Lindqvist, H.; Nadeau, E.; Jensen, S.K. Alpha-tocopherol and β-carotene in legume–grass mixtures as influenced by wilting, ensiling and type of silage additive. Grass Forage Sci. 2012, 67, 119–128. [Google Scholar] [CrossRef]
- Lv, R.; Elsabagh, M.; Obitsu, T.; Sugino, T.; Kurokawa, Y.; Kawamura, K. Effect of varying fermentation conditions with ensiling period and inoculum on photosynthetic pigments and phytol content in Italian ryegrass (Lolium multiflorum Lam.) silage. Anim. Sci. J. 2020, 91, e13309. [Google Scholar] [CrossRef]
- Antoszkiewicz, Z.; Fijalkowska, M.; Mazur-Kusnirek, M.; Przemieniecki, S.; Purwin, C. Effect of a harvest date and cutting height on the concentrations of carotenoids and tocopherols in Virginia fanpetals (Sida hermaphrodita) Herbage and Silage. J. Elem. 2019, 24, 1195–1202. [Google Scholar] [CrossRef]
- Bojović, B.M.; Stojanović, J. Chlorophyll and carotenoid content in wheat cultivars as a function of mineral nutrition. Arch. Biol. Sci. 2005, 57, 283–290. [Google Scholar] [CrossRef]
- Lv, R.; EL-Sabagh, M.; Obitsu, T.; Sugino, T.; Kurokawa, Y.; Kawamura, K. Effects of nitrogen fertilizer and harvesting stage on photosynthetic pigments and phytol contents of Italian ryegrass silage. Anim. Sci. J. 2017, 88, 1513–1522. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Anderson, M.; Walters, J.; Mahoney, A. Effects of processing methods and agronomic variables on carotene contents in forages and predicting carotene in alfalfa hay with near-infrared-reflectance spectroscopy. J. Dairy Sci. 1983, 66, 235–245. [Google Scholar] [CrossRef]
- Elgersma, A.; Søegaard, K.; Jensen, S.K. Interrelations between herbage yield, α-tocopherol, β-carotene, lutein, protein, and fiber in non-leguminous forbs, forage legumes, and a grass–clover mixture as affected by harvest date. J. Agric. Food Chem. 2015, 63, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.D.; Greer, D.H.; Laing, W.A.; McManus, M.T. Changes in photosynthetic efficiency and carotenoid composition in leaves of white clover at different developmental stages. Plant Physiol. Biochem. 2003, 41, 887–893. [Google Scholar] [CrossRef]
- O’Callaghan, T.F.; Vázquez-Fresno, R.; Serra-Cayuela, A.; Dong, E.; Mandal, R.; Hennessy, D.; McAuliffe, S.; Dillon, P.; Wishart, D.S.; Stanton, C. Pasture feeding changes the bovine rumen and milk metabolome. Metabolites 2018, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, T.F.; Hennessy, D.; McAuliffe, S.; Kilcawley, K.N.; O’Donovan, M.; Dillon, P.; Ross, R.P.; Stanton, C. Effect of pasture versus indoor feeding systems on raw milk composition and quality over an entire lactation. J. Dairy Sci. 2016, 99, 9424–9440. [Google Scholar] [CrossRef]
- Dillon, P.; Roche, J.; Shalloo, L.; Horan, B. Optimising financial return from grazing in temperate pastures. In Proceedings of the Satellite Workshop of the XXth International Grassland Congress, Cork, Ireland, 26 June–1 July 2005; pp. 131–147. [Google Scholar]
- Larsen, M.K.; Kidmose, U.; Kristensen, T.; Beaumont, P.; Mortensen, G. Chemical composition and sensory quality of bovine milk as affected by type of forage and proportion of concentrate in the feed ration. J. Sci. Food Agric. 2013, 93, 93–99. [Google Scholar] [CrossRef]
- Mora, O.; Romano, J.L.; González, E.; Ruiz, F.J.; Shimada, A. In vitro and in situ disappearance of β-carotene and lutein from lucerne (Medicago sativa) hay in bovine and caprine ruminal fluids. J. Sci. Food Agric. 1999, 79, 273–276. [Google Scholar] [CrossRef]
- Fernandez, S.C.; Budowski, P.; Ascarelli, I.; Neumark, H.; Bondi, A. Pre-intestinal stability of beta-carotene in ruminants. Int. J. Vitam. Nutr. Research. Int. Z. Vitam. Ernahrungsforsch. J. Int. Vitaminol. Nutr. 1976, 46, 439–445. [Google Scholar]
- Hino, T.; Andoh, N.; Ohgi, H. Effects of β-carotene and α-tocopherol on rumen bacteria in the utilization of long-chain fatty acids and cellulose. J. Dairy Sci. 1993, 76, 600–605. [Google Scholar] [CrossRef]
- Yan, H.; Sun, L.; Zhao, G. Effect of β-carotene on selected indices of in vitro rumen fermentation in goats. J. Anim. Feed Sci. 2007, 16, 581–585. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Use of Lycopene as a food colour-Scientific Opinion of the Panel on Food additives, Flavourings, Processing Aids and Materials in Contact with Food. EFSA J. 2008, 6, 674. [Google Scholar] [CrossRef]
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS). Scientific Opinion on the re-evaluation of lutein (E 161b) as a food additive. EFSA J. 2010, 8, 1678. [Google Scholar] [CrossRef]
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS). Scientific Opinion on the re-evaluation of mixed carotenes (E 160a (i)) and beta-carotene (E 160a (ii)) as a food additive. EFSA J. 2012, 10, 2593. [Google Scholar]
- EEFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Statement on the safety of synthetic zeaxanthin as an ingredient in food supplements. EFSA J. 2012, 10, 2891. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J. An Overview of Carotenoids, Apocarotenoids, and Vitamin A in Agro-Food, Nutrition, Health, and Disease. Mol. Nutr. Food Res. 2019, 63, 1801045. [Google Scholar] [CrossRef]
- O’Callaghan, T.F.; Faulkner, H.; McAuliffe, S.; O’Sullivan, M.G.; Hennessy, D.; Dillon, P.; Kilcawley, K.N.; Stanton, C.; Ross, R.P. Quality characteristics, chemical composition, and sensory properties of butter from cows on pasture versus indoor feeding systems. J. Dairy Sci. 2016, 99, 9441–9460. [Google Scholar] [CrossRef]
- Xu, C.; Wang, H.; Yang, J.; Wang, J.; Duan, Z.; Wang, C.; Liu, J.; Lao, Y. Effects of feeding lutein on production performance, antioxidative status, and milk quality of high-yielding dairy cows. J. Dairy Sci. 2014, 97, 7144–7150. [Google Scholar] [CrossRef]
- Wang, C.; Liu, J.; Duan, B.; Lao, Y.; Qi, P.X.; Ren, D. Effects of dietary antioxidant supplementation of feed, milk processing and storage on the lutein content and sensory quality of bovine milk. Int. J. Dairy Technol. 2018, 71, 849–856. [Google Scholar] [CrossRef]
- Paul, A.A.; Kumar, S.; Kumar, V.; Sharma, R. Milk Analog: Plant based alternatives to conventional milk, production, potential and health concerns. Crit. Rev. Food Sci. Nutr. 2020, 60, 3005–3023. [Google Scholar] [CrossRef]
- Chalupa-Krebzdak, S.; Long, C.J.; Bohrer, B.M. Nutrient density and nutritional value of milk and plant-based milk alternatives. Int. Dairy J. 2018, 87, 84–92. [Google Scholar] [CrossRef]
- McClements, D.J.; Newman, E.; McClements, I.F. Plant-based milks: A review of the science underpinning their design, fabrication, and performance. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2047–2067. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Hughes, J.; Grafenauer, S. Got Mylk? The Emerging Role of Australian Plant-Based Milk Alternatives as A Cow’s Milk Substitute. Nutrients 2020, 12, 1254. [Google Scholar] [CrossRef]
- Xavier, A.A.O.; Mercadante, A.Z.; Garrido-Fernández, J.; Pérez-Gálvez, A. Fat content affects bioaccessibility and efficiency of enzymatic hydrolysis of lutein esters added to milk and yogurt. Food Res. Int. 2014, 65, 171–176. [Google Scholar] [CrossRef]
- García-Cayuela, T.; Nuño-Escobar, B.; Welti-Chanes, J.; Cano, M.P. In vitro bioaccessibility of individual carotenoids from persimmon (Diospyros kaki, cv. Rojo Brillante) used as an ingredient in a model dairy food. J. Sci. Food Agric. 2018, 98, 3246–3254. [Google Scholar] [CrossRef]
- Cilla, A.; Alegría, A.; de Ancos, B.A.; Sánchez-Moreno, C.N.; Cano, M.P.; Plaza, L.; Clemente, G.; Lagarda, M.J.; Barberá, R. Bioaccessibility of tocopherols, carotenoids, and ascorbic acid from milk-and soy-based fruit beverages: Influence of food matrix and processing. J. Agric. Food Chem. 2012, 60, 7282–7290. [Google Scholar] [CrossRef]
- Rodríguez-Roque, M.J.; de Ancos, B.; Sánchez-Vega, R.; Sánchez-Moreno, C.; Cano, M.P.; Elez-Martínez, P.; Martín-Belloso, O. Food matrix and processing influence on carotenoid bioaccessibility and lipophilic antioxidant activity of fruit juice-based beverages. Food Funct. 2016, 7, 380–389. [Google Scholar] [CrossRef]
- Aryana, K.J.; Barnes, H.T.; Emmick, T.K.; McGrew, P.; Moser, B. Lutein is stable in strawberry yogurt and does not affect its characteristics. J. Food Sci. 2006, 71, S467–S472. [Google Scholar] [CrossRef]
- Kubo, M.T.K.; Maus, D.; Xavier, A.A.O.; Mercadante, A.Z.; Viotto, W.H. Transference of lutein during cheese making, color stability, and sensory acceptance of Prato cheese. Food Sci. Technol. 2013, 33, 82–88. [Google Scholar] [CrossRef]
- Domingos, L.; Xavier, A.; Mercadante, A.; Petenate, A.; Jorge, R.; Viotto, W. Oxidative stability of yogurt with added lutein dye. J. Dairy Sci. 2014, 97, 616–623. [Google Scholar] [CrossRef]
- Jones, S.; Aryana, K.; Losso, J.N. Storage stability of lutein during ripening of cheddar cheese. J. Dairy Sci. 2005, 88, 1661–1670. [Google Scholar] [CrossRef]
- Sobral, D.; Costa, R.G.B.; Machado, G.M.; de Paula, J.C.J.; Teodoro, V.A.M.; Nunes, N.M.; dos Santos Pires, A.C.; Pinto, M.S. Can lutein replace annatto in the manufacture of Prato cheese? LWT-Food Sci. Technol. 2016, 68, 349–355. [Google Scholar] [CrossRef]
- Eun, J.-B.; Maruf, A.; Das, P.R.; Nam, S.-H. A review of encapsulation of carotenoids using spray drying and freeze drying. Crit. Rev. Food Sci. Nutr. 2020, 60, 3547–3572. [Google Scholar] [CrossRef]
- Rehman, A.; Tong, Q.; Jafari, S.M.; Assadpour, E.; Shehzad, Q.; Aadil, R.M.; Iqbal, M.W.; Rashed, M.M.; Mushtaq, B.S.; Ashraf, W. Carotenoid-loaded nanocarriers: A comprehensive review. Adv. Colloid Interface Sci. 2020, 275, 102048. [Google Scholar] [CrossRef]
- de Campo, C.; Assis, R.Q.; da Silva, M.M.; Costa, T.M.H.; Paese, K.; Guterres, S.S.; de Oliveira Rios, A.; Flôres, S.H. Incorporation of zeaxanthin nanoparticles in yogurt: Influence on physicochemical properties, carotenoid stability and sensory analysis. Food Chem. 2019, 301, 125230. [Google Scholar] [CrossRef]
- McClements, D.J. Nanoscale nutrient delivery systems for food applications: Improving bioactive dispersibility, stability, and bioavailability. J. Food Sci. 2015, 80, N1602–N1611. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, A.K.D.O.C.; de Carvalho Gomes, C.; de Araújo Amaral, M.L.Q.; de Medeiros, L.D.G.; Medeiros, I.; Porto, D.L.; Aragão, C.F.S.; Maciel, B.L.L.; de Araújo Morais, A.H.; Passos, T.S. Nanoencapsulation improved water solubility and color stability of carotenoids extracted from Cantaloupe melon (Cucumis melo L.). Food Chem. 2019, 270, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Donhowe, E.G.; Flores, F.P.; Kerr, W.L.; Wicker, L.; Kong, F. Characterization and in vitro bioavailability of β-carotene: Effects of microencapsulation method and food matrix. LWT-Food Sci. Technol. 2014, 57, 42–48. [Google Scholar] [CrossRef]
- McClements, D.J. Recent developments in encapsulation and release of functional food ingredients: Delivery by design. Curr. Opin. Food Sci. 2018, 23, 80–84. [Google Scholar] [CrossRef]
- Osorio, L.L.D.R.; Flórez-López, E.; Grande-Tovar, C.D. The Potential of Selected Agri-Food Loss and Waste to Contribute to a Circular Economy: Applications in the Food, Cosmetic and Pharmaceutical Industries. Molecules 2021, 26, 515. [Google Scholar] [CrossRef] [PubMed]
- Joglekar, S.N.; Pathak, P.D.; Mandavgane, S.A.; Kulkarni, B.D. Process of fruit peel waste biorefinery: A case study of citrus waste biorefinery, its environmental impacts and recommendations. Environ. Sci. Pollut. Res. 2019, 26, 34713–34722. [Google Scholar] [CrossRef]
- Sharma, M.; Usmani, Z.; Gupta, V.K.; Bhat, R. Valorization of fruits and vegetable wastes and by-products to produce natural pigments. Crit. Rev. Biotechnol. 2021, 41, 535–563. [Google Scholar] [CrossRef]
- Rizk, E.M.; El-Kady, A.T.; El-Bialy, A.R. Charactrization of carotenoids (lyco-red) extracted from tomato peels and its uses as natural colorants and antioxidants of ice cream. Ann. Agric. Sci. 2014, 59, 53–61. [Google Scholar] [CrossRef]
- Pandya, D.; Akbari, S.; Bhatt, H.; Joshi, D. Standardization of solvent extraction process for Lycopene extraction from tomato pomace. J. Appl. Biotechnol. Bioeng. 2017, 2, 00019. [Google Scholar] [CrossRef]
- Putnik, P.; Bursać Kovačević, D.; Režek Jambrak, A.; Barba, F.J.; Cravotto, G.; Binello, A.; Lorenzo, J.M.; Shpigelman, A. Innovative “green” and novel strategies for the extraction of bioactive added value compounds from citrus wastes—A review. Molecules 2017, 22, 680. [Google Scholar] [CrossRef]
- Pattnaik, M.; Pandey, P.; Martin, G.J.; Mishra, H.N.; Ashokkumar, M. Innovative technologies for extraction and microencapsulation of bioactives from plant-based food waste and their applications in functional food development. Foods 2021, 10, 279. [Google Scholar] [CrossRef]
- Abid, Y.; Azabou, S.; Jridi, M.; Khemakhem, I.; Bouaziz, M.; Attia, H. Storage stability of traditional Tunisian butter enriched with antioxidant extract from tomato processing by-products. Food Chem. 2017, 233, 476–482. [Google Scholar] [CrossRef]
- Šeregelj, V.; Pezo, L.; Šovljanski, O.; Lević, S.; Nedović, V.; Markov, S.; Tomić, A.; Čanadanović-Brunet, J.; Vulić, J.; Šaponjac, V.T. New concept of fortified yogurt formulation with encapsulated carrot waste extract. LWT 2021, 138, 110732. [Google Scholar] [CrossRef]
- Gies, M.; Descalzo, A.M.; Servent, A.; Dhuique-Mayer, C. Incorporation and stability of carotenoids in a functional fermented maize yogurt-like product containing phytosterols. LWT 2019, 111, 105–110. [Google Scholar] [CrossRef]
- Jackson, J.G.; Lien, E.L.; White, S.J.; Bruns, N.J.; Kuhlman, C.F. Major carotenoids in mature human milk: Longitudinal and diurnal patterns. J. Nutr. Biochem. 1998, 9, 2–7. [Google Scholar] [CrossRef]
- Giuliano, A.R.; Neilson, E.M.; Yap, H.-H.; Baier, M.; Canfield, L.M. Quantitation of and inter/intra-individual variability in major carotenoids of mature human milk. J. Nutr. Biochem. 1994, 5, 551–556. [Google Scholar] [CrossRef]
- de Azeredo, V.B.; Trugo, N.M. Retinol, carotenoids, and tocopherols in the milk of lactating adolescents and relationships with plasma concentrations. Nutrition 2008, 24, 133–139. [Google Scholar] [CrossRef]
- Sommerburg, O.; Meissner, K.; Nelle, M.; Lenhartz, H.; Leichsenring, M. Carotenoid supply in breast-fed and formula-fed neonates. Eur. J. Pediatrics 2000, 159, 86–90. [Google Scholar] [CrossRef]
- Canfield, L.M.; Clandinin, M.T.; Davies, D.P.; Fernandez, M.C.; Jackson, J.; Hawkes, J.; Goldman, W.J.; Pramuk, K.; Reyes, H.; Sablan, B. Multinational study of major breast milk carotenoids of healthy mothers. Eur. J. Nutr. 2003, 42, 133–141. [Google Scholar] [CrossRef]
- Hammond, B.R., Jr. Possible role for dietary lutein and zeaxanthin in visual development. Nutr. Rev. 2008, 66, 695–702. [Google Scholar] [CrossRef]
- Henriksen, B.S.; Chan, G.; Hoffman, R.O.; Sharifzadeh, M.; Ermakov, I.V.; Gellermann, W.; Bernstein, P.S. Interrelationships between maternal carotenoid status and newborn infant macular pigment optical density and carotenoid status. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5568–5578. [Google Scholar] [CrossRef]
- Vishwanathan, R.; Kuchan, M.J.; Sen, S.; Johnson, E.J. Lutein and preterm infants with decreased concentrations of brain carotenoids. J. Pediatric Gastroenterol. Nutr. 2014, 59, 659–665. [Google Scholar] [CrossRef]
- Jewell, V.; Mayes, C.; Tubman, T.; Northrop-Clewes, C.; Thurnham, D. A comparison of lutein and zeaxanthin concentrations in formula and human milk samples from Northern Ireland mothers. Eur. J. Clin. Nutr. 2004, 58, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Lipkie, T.E.; Banavara, D.; Shah, B.; Morrow, A.L.; McMahon, R.J.; Jouni, Z.E.; Ferruzzi, M.G. Caco-2 accumulation of lutein is greater from human milk than from infant formula despite similar bioaccessibility. Mol. Nutr. Food Res. 2014, 58, 2014–2022. [Google Scholar] [CrossRef] [PubMed]
- Mackey, A.D.; Albrecht, D.; Oliver, J.; Williams, T.; Long, A.C.; Price, P.T. Plasma carotenoid concentrations of infants are increased by feeding a milk-based infant formula supplemented with carotenoids. J. Sci. Food Agric. 2013, 93, 1945–1952. [Google Scholar] [CrossRef]
- Kon, I.Y.; Gmoshinskaya, M.V.; Safronova, A.I.; Alarcon, P.; Vandenplas, Y. Growth and tolerance assessment of a lutein-fortified infant formula. Pediatric Gastroenterol. Hepatol. Nutr. 2014, 17, 104. [Google Scholar] [CrossRef] [PubMed]
- Capeding, R.; Gepanayao, C.P.; Calimon, N.; Lebumfacil, J.; Davis, A.M.; Stouffer, N.; Harris, B.J. Lutein-fortified infant formula fed to healthy term infants: Evaluation of growth effects and safety. Nutr. J. 2010, 9, 22. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Carotenoid | Lipid Transporter | References |
|---|---|---|
| β-Carotene | SR-BI, NPC1L1 | [43,44,45] |
| α-Carotene | SR-BI, NPC1L1 | [43,44] |
| β-Cryptoxanthin | SR-BI, NPC1L1 | [43,44] |
| Lutein | SR-BI, NPC1L1, CD-36 | [44,46,47,48] |
| Zeaxanthin | SR-BI, NPC1L1 | [44,45] |
| Phytoene | SR-BI | [49] |
| Phytofluene | SR-BI | [49] |
| Lycopene | SR-BI, NPC1L1 *, CD-36 | [44,47,50] |
| Food Matrix | Carotenoid | Processing Technique | Carotenoid Retention (%) | Reference |
|---|---|---|---|---|
| Gouda cheese | β-Carotene | Cheese making (initial 12 days) | ↓ 62% | [62] |
| Ripening (26 weeks) | ↓ 9% | |||
| Edammer cheese | β-Carotene | Cheese making (initial 12 days) | ↓ 30% | [62] |
| Ripening (20 weeks) | ↓ 60% | |||
| Prato cheese | Lutein | Prato cheese making (after milk pasteurization and at milk coagulation) | ↓ <10% | [112,115] |
| Cheddar cheese | Lutein | Ripening (24 weeks) | x | [114] |
| Yogurt | β-Carotene, β-Cryptoxanthin, Lycopene | Pasteurization | ↓ 20–27% | [132] |
| Fermentation | ↓ <3% | |||
| Milk | Lutein | HTST | ↓ <3% | [102] |
| UHT | ↓ 8% | |||
| Milk | Lutein + Vitamin E | HTST | ↓ <3% | [102] |
| UHT | ↓ 3% | |||
| Milk | β-Carotene | Pasteurization | x | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conboy Stephenson, R.; Ross, R.P.; Stanton, C. Carotenoids in Milk and the Potential for Dairy Based Functional Foods. Foods 2021, 10, 1263. https://doi.org/10.3390/foods10061263
Conboy Stephenson R, Ross RP, Stanton C. Carotenoids in Milk and the Potential for Dairy Based Functional Foods. Foods. 2021; 10(6):1263. https://doi.org/10.3390/foods10061263
Chicago/Turabian StyleConboy Stephenson, Ruth, R. Paul Ross, and Catherine Stanton. 2021. "Carotenoids in Milk and the Potential for Dairy Based Functional Foods" Foods 10, no. 6: 1263. https://doi.org/10.3390/foods10061263
APA StyleConboy Stephenson, R., Ross, R. P., & Stanton, C. (2021). Carotenoids in Milk and the Potential for Dairy Based Functional Foods. Foods, 10(6), 1263. https://doi.org/10.3390/foods10061263