
Characterization of Kefir Produced in Household Conditions: Physicochemical and Nutritional Profile, and Storage Stability

 , ,
, ,  ,
,  ,
,  ,
,  ,
, 
 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Kefir Grains Storage and Kefir Production
2.2. Activity of Kefir Grains
2.2.1. Biomass Growth
2.2.2. Acidification Kinetic
2.3. Viable Microorganisms and Inhibitory Activity Test
2.3.1. Bacterial and Yeasts Counts
2.3.2. Inhibitory Activity Test
2.4. Physicochemical Characteristics of Kefir Beverage
2.4.1. Particle Size, Polydispersity Index, and Zeta Potential
2.4.2. Fourier Transform Infrared Spectroscopy (FTIR)
2.4.3. Viscosity and pH
2.5. Nutritional Analysis of Kefir Beverage
2.6. Statistical Analysis
3. Results
3.1. Activity of Kefir Grains
3.1.1. Biomass Growth
3.1.2. Acidification Kinetic
3.2. Viable Microorganisms and Inhibitory Activity Test
3.3. Physicochemical Characteristics of Kefir Beverage
3.3.1. Particle Size, PdI, and Zeta Potential
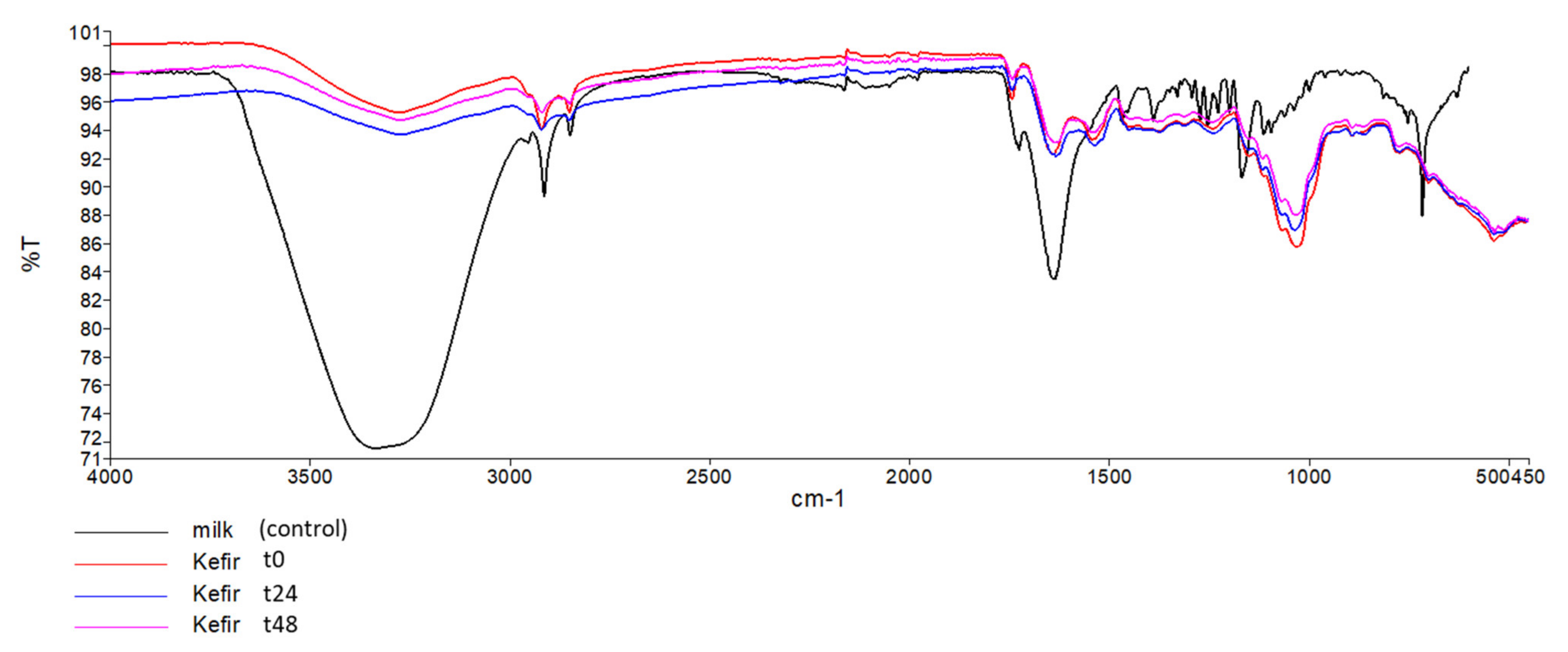
3.3.2. FTIR
3.3.3. Viscosity and pH
3.4. Nutritional Analysis of Kefir Beverage
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wouters, J.T.M.; Ayad, E.H.E.; Hugenholtz, J.; Smit, G. Microbes from raw milk for fermented dairy products. Int. Dairy J. 2002, 12, 91–109. [Google Scholar] [CrossRef]
- Bourrie, B.C.T.; Willing, B.P.; Cotter, P.D. The Microbiota and Health Promoting Characteristics of the Fermented Beverage Kefir. Front. Microbiol. 2016, 7, 647. [Google Scholar] [CrossRef]
- Vinderola, C.G.; Duarte, J.; Thangavel, D.; Perdigón, G.; Farnworth, E.; Matar, C. Immunomodulating capacity of kefir. J. Dairy Res. 2005, 72, 195–202. [Google Scholar] [CrossRef]
- Farnworth, E.R. Kefir—A complex probiotic. Food Sci. Technol. Bull. Funct. Foods 2005, 2, 1–17. [Google Scholar] [CrossRef]
- Ahmed, Z.; Wang, Y.; Ahmad, A.; Khan, S.T.; Nisa, M.; Ahmad, H.; Afreen, A. Kefir and Health: A Contemporary Perspective. Crit. Rev. Food Sci. Nutr. 2013, 53, 422–434. [Google Scholar] [CrossRef]
- Hertzler, S.R.; Clancy, S.M. Kefir improves lactose digestion and tolerance in adults with lactose maldigestion. J. Am. Diet. Assoc. 2003, 103, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, K.L.; Carvalho, J.C.T.; Schneedorf, J.M. Anti-inflammatory properties of kefir and its polysaccharide extract. Inflammopharmacology 2005, 13, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Iraporda, C.; Romanin, D.E.; Rumbo, M.; Garrote, G.L.; Abraham, A.G. The role of lactate on the immunomodulatory properties of the nonbacterial fraction of kefir. Food Res. Int. 2014, 62, 247–253. [Google Scholar] [CrossRef]
- Rodrigues, K.L.; Caputo, L.R.G.; Carvalho, J.C.T.; Evangelista, J.; Schneedorf, J.M. Antimicrobial and healing activity of kefir and kefiran extract. Int. J. Antimicrob. Agents 2005, 25, 404–408. [Google Scholar] [CrossRef]
- Liu, J.-R.; Chen, M.-J.; Lin, C.-W. Antimutagenic and Antioxidant Properties of Milk−Kefir and Soymilk−Kefir. J. Agric. Food Chem. 2005, 53, 2467–2474. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Gu, F.; Ruan, H.; Chen, Q.; He, J.; He, G. Induction of apoptosis of gastric cancer cells SGC7901 in vitro by a cell-free fraction of Tibetan kefir. Int. Dairy J. 2013, 30, 14–18. [Google Scholar] [CrossRef]
- Huseini, H.F.; Rahimzadeh, G.; Fazeli, M.R.; Mehrazma, M.; Salehi, M. Evaluation of wound healing activities of kefir products. Burns 2012, 38, 719–723. [Google Scholar] [CrossRef]
- Carasi, P.; Racedo, S.M.; Jacquot, C.; Romanin, D.E.; Serradell, M.A.; Urdaci, M.C. Impact of Kefir Derived Lactobacillus kefiri on the Mucosal Immune Response and Gut Microbiota. J. Immunol. Res. 2015, 2015, 361604. [Google Scholar] [CrossRef] [PubMed]
- Garrote, G.L.; Abraham, A.G.; De Antoni, G.L. Inhibitory Power of Kefir: The Role of Organic Acids. J. Food Prot. 2000, 63, 364–369. [Google Scholar] [CrossRef]
- Iraporda, C.; Júnior, M.A.; Neumann, E.; Nunes, Á.C.; Nicoli, J.R.; Abraham, A.G.; Garrote, G.L. Biological activity of the non-microbial fraction of kefir: Antagonism against intestinal pathogens. J. Dairy Res. 2017, 84, 339–345. [Google Scholar] [CrossRef]
- Simova, E.; Beshkova, D.; Angelov, A.; Hristozova, T.; Frengova, G.; Spasov, Z. Lactic acid bacteria and yeasts in kefir grains and kefir made from them. J. Ind. Microbiol. Biotechnol. 2002, 28, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Vardjan, T.; Lorbeg, P.M.; Rogelj, I.; Majhenič, A.Č. Characterization and stability of lactobacilli and yeast microbiota in kefir grains. J. Dairy Sci. 2013, 96, 2729–2736. [Google Scholar] [CrossRef]
- Witthuhn, R.C.; Cilliers, A.; Britz, T.J. Evaluation of different preservation techniques on the storage potential of Kefir grains. J. Dairy Res. 2005, 72, 125–128. [Google Scholar] [CrossRef]
- Garrote, G.; Abraham, A.; De Antoni, G. Preservation of Kefir Grains, a Comparative Study. LWT 1997, 30, 77–84. [Google Scholar] [CrossRef]
- Garrote, G.L.; Abraham, A.G.; De Antoni, G.L. Chemical and microbiological characterisation of kefir grains. J. Dairy Res. 2001, 68, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Leite, A.M.O.; Leite, D.C.A.; Del Aguila, E.M.; Alvares, T.S.; Peixoto, R.S.; Miguel, M.A.L.; Silva, J.T.; Paschoalin, V.M.F. Microbiological and chemical characteristics of Brazilian kefir during fermentation and storage processes. J. Dairy Sci. 2013, 96, 4149–4159. [Google Scholar] [CrossRef]
- Pogačić, T.; Šinko, S.; Zamberlin, Š.; Samaržija, D. Microbiota of Kefir Grains. Mljekarstvo 2013, 63, 3–14. [Google Scholar]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Bengoa, A.A.; Iraporda, C.; Garrote, G.L.; Abraham, A.G. Kefir micro-organisms: Their role in grain assembly and health properties of fermented milk. J. Appl. Microbiol. 2019, 126, 686–700. [Google Scholar] [CrossRef] [PubMed]
- Garrote, G.L.; Abraham, A.G.; De Antoni, G.L. Characteristics of kefir prepared with different grain [ratio] milk ratios. J. Dairy Res. 1998, 65, 149–154. [Google Scholar] [CrossRef]
- Rattray, F.P.; O’Connell, M.J. Fermented Milks—Kefir. In Encyclopedia of Dairy Sciences; Fuquay, J.W., Fox, P.F., McSweeney, P.L.H., Eds.; Academic Press Inc.: London, UK, 2011; pp. 518–524. [Google Scholar]
- Guzel-Seydim, Z.; Kok-Tas, T.; Ertekin-Filiz, B.; Seydim, A.C. Effect of different growth conditions on biomass increase in kefir grains. J. Dairy Sci. 2011, 94, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Schoevers, A.; Britz, T.J. Influence of different culturing conditions on kefir grain increase. Int. J. Dairy Technol. 2003, 56, 183–187. [Google Scholar] [CrossRef]
- Rea, M.C.; Lennartsson, T.; Dillon, P.; Drinan, F.D.; Reville, W.J.; Heapes, M.; Cogan, T.M. Irish kefir-like grains: Their structure, microbial composition and fermentation kinetics. J. Appl. Bacteriol. 1996, 81, 83–94. [Google Scholar] [CrossRef]
- Nielsen, B.; Gürakan, G.C.; Ünlü, G. Kefir: A Multifaceted Fermented Dairy Product. Probiotics Antimicrob. Proteins 2014, 6, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Wszolek, M.; Tamime, A.; Muir, D.; Barclay, M. Properties of Kefir made in Scotland and Poland using Bovine, Caprine and Ovine Milk with Different Starter Cultures. LWT 2001, 34, 251–261. [Google Scholar] [CrossRef]
- Vieira, C.P.; Álvares, T.S.; Gomes, L.S.; Torres, A.G.; Paschoalin, V.M.F.; Conte-Junior, C.A. Kefir Grains Change Fatty Acid Profile of Milk during Fermentation and Storage. PLoS ONE 2015, 10, e0139910. [Google Scholar] [CrossRef]
- Gul, O.; Mortas, M.; Atalar, I.; Dervisoglu, M.; Kahyaoglu, T. Manufacture and characterization of kefir made from cow and buffalo milk, using kefir grain and starter culture. J. Dairy Sci. 2015, 98, 1517–1525. [Google Scholar] [CrossRef]
- Magra, T.I.; Antoniou, K.D.; Psomas, E.I. Effect of milk fat, kefir grain inoculum and storage time on the flow properties and microbiological characteristics of kefir. J. Texture Stud. 2012, 43, 299–308. [Google Scholar] [CrossRef]
- Hecer, C.; Ulusoy, B.; Kaynarca, D. Effect of Different Fermentation Conditions on Composition of Kefir Microbiota. Int. Food Res. J. 2019, 26, 401–409. [Google Scholar]
- Irigoyen, A.; Arana, I.; Castiella, M.; Torre, P.; Ibáñez, F. Microbiological, physicochemical, and sensory characteristics of kefir during storage. Food Chem. 2005, 90, 613–620. [Google Scholar] [CrossRef]
- Zajšek, K.; Goršek, A. Modelling of batch kefir fermentation kinetics for ethanol production by mixed natural microflora. Food Bioprod. Process. 2010, 88, 55–60. [Google Scholar] [CrossRef]
- Barukčić, I.; Gracin, L.; Jambrak, A.R.; Božanić, R. Comparison of chemical, rheological and sensory properties of kefir produced by kefir grains and commercial kefir starter. Mljekarstvo 2017, 67, 169–176. [Google Scholar] [CrossRef]
- De Vrese, M.; Keller, B.; Barth, C.A. Enhancement of intestinal hydrolysis of lactose by microbial β-galactosidase (EC 3.2.1.23) of kefir. Br. J. Nutr. 1992, 67, 67–75. [Google Scholar] [CrossRef]
- Guzel-Seydim, Z.; Seydim, A.C.; Greene, A.K. Organic Acids and Volatile Flavor Components Evolved During Refrigerated Storage of Kefir. J. Dairy Sci. 2000, 83, 275–277. [Google Scholar] [CrossRef]
- Otles, S.; Cagindi, O. Kefir: A Probiotic Dairy-Composition, Nutritional and Therapeutic Aspects. Pak. J. Nutr. 2003, 2, 54–59. [Google Scholar] [CrossRef]
- Walsh, A.M.; Crispie, F.; Kilcawley, K.; O’Sullivan, O.; O’Sullivan, M.G.; Claesson, M.J.; Cotter, P.D. Microbial Succession and Flavor Production in the Fermented Dairy Beverage Kefir. mSystems 2016, 1, e00052-16. [Google Scholar] [CrossRef]
- Codex Alimentarius International Food Standards. Codex Standard for Fermented Milks (Codex Stan CXS 243-2003) Codex Alimentarius; FAO: Rome, Italy; ISBN 5856420187. Available online: http://www.fao.org/fao-who-codexalimentarius/codex-texts/list-standards/en/ (accessed on 12 January 2021).
- Grønnevik, H.; Falstad, M.; Narvhus, J.A. Microbiological and chemical properties of Norwegian kefir during storage. Int. Dairy J. 2011, 21, 601–606. [Google Scholar] [CrossRef]
- Kim, Y.J.; Liu, R.H. Increase of Conjugated Linoleic Acid Content in Milk by Fermentation with Lactic Acid Bacteria. J. Food Sci. 2002, 67, 1731–1737. [Google Scholar] [CrossRef]
- Hamet, M.F.; Londero, A.; Medrano, M.; Vercammen, E.; Van Hoorde, K.; Garrote, G.L.; Huys, G.; Vandamme, P.; Abraham, A.G. Application of culture-dependent and culture-independent methods for the identification of Lactobacillus kefiranofaciens in microbial consortia present in kefir grains. Food Microbiol. 2013, 36, 327–334. [Google Scholar] [CrossRef]
- Diosma, G.; Romanin, D.E.; Rey-Burusco, M.F.; Londero, A.; Garrote, G.L. Yeasts from kefir grains: Isolation, identification, and probiotic characterization. World J. Microbiol. Biotechnol. 2014, 30, 43–53. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2020. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 20th ed.; Latimer, G., Ed.; AOAC International: Gaithersburg, MD, USA, 2016; ISBN 0935584870. [Google Scholar]
- Barros, L.; Pereira, E.; Calhelha, R.C.; Dueñas, M.; Carvalho, A.M.; Santos-Buelga, C.; Ferreira, I.C. Bioactivity and chemical characterization in hydrophilic and lipophilic compounds of Chenopodium ambrosioides L. J. Funct. Foods 2013, 5, 1732–1740. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. Food Energy—Methods of Analysis and Conversion Factors. Food and Nutrition Paper 77. 2003. Available online: http://www.fao.org/3/Y5022E/Y5022E00.htm (accessed on 3 March 2021).
- Instituto Portugês do Mar e da Atmosfera—IPMA. Clima de Portugal Continental. Available online: https://www.ipma.pt/pt/educativa/tempo.clima/ (accessed on 10 February 2021).
- Londero, A.; Hamet, M.F.; De Antoni, G.L.; Garrote, G.L.; Abraham, A.G. Kefir grains as a starter for whey fermentation at different temperatures: Chemical and microbiological characterisation. J. Dairy Res. 2012, 79, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Guzel-Seydim, Z.B.; Kok-Tas, T.; Greene, A.K.; Seydim, A.C. Review: Functional Properties of Kefir. Crit. Rev. Food Sci. Nutr. 2011, 51, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.G.; De Antoni, G.L. Characterization of kefir grains grown in cows’ milk and in soya milk. J. Dairy Res. 1999, 66, 327–333. [Google Scholar] [CrossRef] [PubMed]
- De Sainz, I.; Redondo-Solano, M.; Solano, G.; Ramírez, L. Short communication: Effect of different kefir grains on the attributes of kefir produced with milk from Costa Rica. J. Dairy Sci. 2020, 103, 215–219. [Google Scholar] [CrossRef]
- Pop, C.R.; Apostu, S.; Salanţă, L.; Rotar, A.M.; Sindic, M.; Mabon, N.; Socaciu, C. Influence of Different Growth Conditions on the Kefir Grains Production, used in the Kefiran Synthesis. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca Food Sci. Technol. 2014, 71, 147–153. [Google Scholar] [CrossRef][Green Version]
- Irigoyen, A.; Ortigosa, M.; Torre, P.; Ibáñez, F. Influence of Different Technological Parameters in the Evolution of PH during Fermentation of Kefir. Milchwissenschaft 2003, 58, 631–633. [Google Scholar]
- Hahn, C.; Sramek, M.; Nöbel, S.; Hinrichs, J. Post-processing of concentrated fermented milk: Influence of temperature and holding time on the formation of particle clusters. Dairy Sci. Technol. 2012, 92, 91–107. [Google Scholar] [CrossRef]
- Beirami-Serizkani, F.; Hojjati, M.; Jooyandeh, H. The effect of microbial transglutaminase enzyme and Persian gum on the characteristics of traditional kefir drink. Int. Dairy J. 2021, 112, 104843. [Google Scholar] [CrossRef]
- Danaei, M.; Dehghankhold, M.; Ataei, S.; Davarani, F.H.; Javanmard, R.; Dokhani, A.; Khorasani, S.; Mozafari, M.R. Impact of Particle Size and Polydispersity Index on the Clinical Applications of Lipidic Nanocarrier Systems. Pharmaceutics 2018, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Teimouri, S.; Abbasi, S.; Scanlon, M.G. Stabilisation mechanism of various inulins and hydrocolloids: Milk-sour cherry juice mixture. Int. J. Dairy Technol. 2018, 71, 208–215. [Google Scholar] [CrossRef]
- Tan, T.J.; Wang, D.; Moraru, C.I. A physicochemical investigation of membrane fouling in cold microfiltration of skim milk. J. Dairy Sci. 2014, 97, 4759–4771. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, N.; Xu, Y.; Goodacre, R. Fourier transform infrared spectroscopy and multivariate analysis for the detection and quantification of different milk species. J. Dairy Sci. 2010, 93, 5651–5660. [Google Scholar] [CrossRef]
- Rimada, P.S.; Abraham, A.G. Kefiran improves rheological properties of glucono-δ-lactone induced skim milk gels. Int. Dairy J. 2006, 16, 33–39. [Google Scholar] [CrossRef]
- Şanli, T.; Sezgin, E.; Şenel, E.; Benli, M. The effect of transglutaminase on some physicochemical and sensory properties of the Turkish drinking yoghurt Ayran. Int. J. Dairy Technol. 2013, 66, 410–416. [Google Scholar] [CrossRef]
- Sarkar, S. Potential of kefir as a dietetic beverage—A review. Br. Food J. 2007, 109, 280–290. [Google Scholar] [CrossRef]
- INSA Tabela Composição Alimentos. 2019. Available online: http://portfir.insa.pt/foodcomp/search (accessed on 25 March 2021).
- Assadi, M.M.; Pourahmad, R.; Moazami, N. Use of isolated kefir starter cultures in kefir production. World J. Microbiol. Biotechnol. 2000, 16, 541–543. [Google Scholar] [CrossRef]
- Dallas, D.C.; Citerne, F.; Tian, T.; Silva, V.L.; Kalanetra, K.M.; Frese, S.A.; Robinson, R.C.; Mills, D.A.; Barile, D. Peptidomic analysis reveals proteolytic activity of kefir microorganisms on bovine milk proteins. Food Chem. 2016, 197, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Alm, L. Effect of Fermentation on Proteins of Swedish Fermented Milk Products. J. Dairy Sci. 1982, 65, 1696–1704. [Google Scholar] [CrossRef]
- Neis, E.P.J.G.; DeJong, C.H.C.; Rensen, S.S. The Role of Microbial Amino Acid Metabolism in Host Metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef] [PubMed]
- Guzel-Seydim, Z.B.; Seydim, A.C.; Greene, A.K.; Taş, T. Determination of antimutagenic properties of acetone extracted fermented milks and changes in their total fatty acid profiles including conjugated linoleic acids. Int. J. Dairy Technol. 2006, 59, 209–215. [Google Scholar] [CrossRef]
- Molina, S.; Moran-Valero, M.; Martin, D.; Vázquez, L.; Vargas, T.; Torres, C.; De Molina, A.R.; Reglero, G. Antiproliferative effect of alkylglycerols as vehicles of butyric acid on colon cancer cells. Chem. Phys. Lipids 2013, 175–176, 50–56. [Google Scholar] [CrossRef]
- Guerzoni, M.E.; Lanciotti, R.; Cocconcelli, P.S. Alteration in cellular fatty acid composition as a response to salt, acid, oxidative and thermal stresses in Lactobacillus helveticus. Microbiology 2001, 147, 2255–2264. [Google Scholar] [CrossRef]
- Nantapo, C.T.W.; Muchenje, V.; Hugo, A. Atherogenicity index and health-related fatty acids in different stages of lactation from Friesian, Jersey and Friesian × Jersey cross cow milk under a pasture-based dairy system. Food Chem. 2014, 146, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Cais-Sokolińska, D.; Wójtowski, J.; Pikul, J.; Danków, R.; Majcher, M.; Teichert, J.; Bagnicka, E. Formation of volatile compounds in kefir made of goat and sheep milk with high polyunsaturated fatty acid content. J. Dairy Sci. 2015, 98, 6692–6705. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Kefir Beverage | |
|---|---|
| LAB (CFU/mL) | 7 × 107 |
| Yeasts (CFU/mL) | 2 × 106 |
| Coliforms (CFU/mL) | Absent |
| Control | Kefir | |||
|---|---|---|---|---|
| t0 | t24 | t48 | ||
| Diameter (nm) | 280 ± 5 b | 439 ± 42 a | 256 ± 6 b | 249 ± 1 b |
| PdI | 0.18 ± 0.01 b | 0.295 ± 0.006 a | 0.231 ± 0.008 c | 0.22 ± 0.02 c |
| Zeta potential (mV) | −35 ± 2 a | −38 ± 1 a | −31 ± 2 b | −30 ± 3 b |
| Control | Kefir | |||
|---|---|---|---|---|
| t0 | t24 | t48 | ||
| pH | 6.60 ± 0.00 a | 4.60 ± 0.05 b | 4.54 ± 0.02 b,c | 4.50 ± 0.04 c |
| Viscosity (mPa.s) | 2.11 ± 0.01 b | 32 ± 4 a | 26 ± 2 c | 24 ± 4 c |
| Control | Kefir | |||
|---|---|---|---|---|
| t0 | t24 | t48 | ||
| Energy (kcal/100 mL) | 48.2 ± 0.4 a | 43.8 ± 0.6 b | 44.5 ± 0.8 b | 44 ± 2 b |
| Carbohydrates (% w/v) | 5.14 ± 0.08 | 4.9 ± 0.2 | 5.0 ± 0.1 | 5.0 ± 0.2 |
| Lactose (% w/v) | 4.74 ± 0.05 a | 4.1 ± 0.2 b | 3.75 ± 0.08 b | 3.8 ± 0.2 b |
| Proteins (% w/v) | 2.8 ± 0.1 | 3.2 ± 0.2 | 3.1 ± 0.1 | 3.15 ± 0.05 |
| Total Fat (% w/v) | 1.81 ± 0.03 | 1.28 ± 0.04 | 1.32 ± 0.09 | 1.3 ± 0.3 |
| Lactic acid (% w/v) | 0.02 ± 0.00 b | 0.59 ± 0.07 a | 0.63 ± 0.01 a | 0.61 ± 0.05 a |
| Ash (% w/v) | 0.50 ± 0.01 a | 0.58 ± 0.02 b | 0.59 ± 0.01 a,b | 0.59 ± 0.02 b |
| Dry matter (% w/w) | 10.28 ± 0.04 a | 9.9 ± 0.1 b | 10.05 ± 0.09 b | 10.0 ± 0.2 b |
| Control | Kefir | |||
|---|---|---|---|---|
| Fatty Acids (%) | t0 | t24 | t48 | |
| C6:0 | 3.86 ± 0.05 | 3.6 ± 0.2 | 3.3 ± 0.1 | 3.7 ± 0.1 |
| C8:0 | 2.12 ± 0.03 a | 2.07 ± 0.07 a | 1.95 ± 0.05 b | 2.18 ± 0.05 a |
| C10:0 | 4.41 ± 0.02 | 4.5 ± 0.3 | 4.1 ± 0.2 | 4.6 ± 0.1 |
| C11:0 | 0.10 ± 0.00 | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 |
| C12:0 | 5.5 ± 0.1 b | 5.3 ± 0.1 b | 5.2 ± 0.2 b | 5.6 ± 0.1 a |
| C13:0 | 0.12 ± 0.01 | 0.10 ± 0.00 | 0.10 ± 0.01 | 0.11 ± 0.02 |
| C14:0 | 14.62 ± 0.02 | 14.3 ± 0.2 | 14.2 ± 0.3 | 14.1 ± 0.9 |
| C14:1 | 1.15 ± 0.02 | 1.18 ± 0.06 | 1.20 ± 0.07 | 1.11 ± 0.04 |
| C15:0 | 1.18 ± 0.00 b | 1.17 ± 0.02 b | 1.14 ± 0.03 b | 1.23 ± 0.01 a |
| C15:1 | 0.25 ± 0.01 b | 0.27 ± 0.01 a,b | 0.26 ± 0.01 b | 0.28 ± 0.01 a |
| C16:0 | 39.34 ± 0.02 b | 38.4 ± 0.3 c | 38.9 ± 0.5 c | 39.9 ± 0.2 a |
| C16:1 | 1.57 ± 0.01 a | 1.61 ± 0.06 a | 1.57 ± 0.08 a | 1.38 ± 0.07 b |
| C17:0 | 0.61 ± 0.02 b | 0.61 ± 0.02 b | 0.61 ± 0.05 b | 0.70 ± 0.05 a |
| C18:0 | 5.71 ± 0.02 | 5.64 ± 0.06 | 5.7 ± 0.1 | 5.74 ± 0.09 |
| C18:1n-9 | 18.4 ± 0.1 b | 19.4 ± 0.3 a | 19.8 ± 0.3 a | 18.0 ± 0.7 a |
| C18:2n-6 | 0.79 ± 0.03 | 1.4 ± 0.3 | 1.52 ± 0.07 | 0.7 ± 0.7 |
| C18:3n-3 | 0.023 ± 0.001 b | 0.123 ± 0.003 a | 0.12 ± 0.00 a | 0.15 ± 0.01 a |
| C20:0 | 0.21 ± 0.01 | 0.32 ± 0.03 | 0.31 ± 0.06 | 0.32 ± 0.08 |
| SFA | 77.81 ± 0.07 b | 76.1 ± 0.5 a | 75.6 ± 0.3 a | 78.5 ± 1.2 a |
| MUFA | 21.37 ± 0.09 b | 22.4 ± 0.3 a | 22.8 ± 0.3 a | 20.7 ± 0.7 b |
| PUFA | 0.82 ± 0.03 | 1.4 ± 0.4 | 1.6 ± 0.1 | 0.8 ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves, E.; Ntungwe, E.N.; Gregório, J.; Rodrigues, L.M.; Pereira-Leite, C.; Caleja, C.; Pereira, E.; Barros, L.; Aguilar-Vilas, M.V.; Rosado, C.; et al. Characterization of Kefir Produced in Household Conditions: Physicochemical and Nutritional Profile, and Storage Stability. Foods 2021, 10, 1057. https://doi.org/10.3390/foods10051057
Alves E, Ntungwe EN, Gregório J, Rodrigues LM, Pereira-Leite C, Caleja C, Pereira E, Barros L, Aguilar-Vilas MV, Rosado C, et al. Characterization of Kefir Produced in Household Conditions: Physicochemical and Nutritional Profile, and Storage Stability. Foods. 2021; 10(5):1057. https://doi.org/10.3390/foods10051057
Chicago/Turabian StyleAlves, Emília, Epole N. Ntungwe, João Gregório, Luis M. Rodrigues, Catarina Pereira-Leite, Cristina Caleja, Eliana Pereira, Lillian Barros, M. Victorina Aguilar-Vilas, Catarina Rosado, and et al. 2021. "Characterization of Kefir Produced in Household Conditions: Physicochemical and Nutritional Profile, and Storage Stability" Foods 10, no. 5: 1057. https://doi.org/10.3390/foods10051057
APA StyleAlves, E., Ntungwe, E. N., Gregório, J., Rodrigues, L. M., Pereira-Leite, C., Caleja, C., Pereira, E., Barros, L., Aguilar-Vilas, M. V., Rosado, C., & Rijo, P. (2021). Characterization of Kefir Produced in Household Conditions: Physicochemical and Nutritional Profile, and Storage Stability. Foods, 10(5), 1057. https://doi.org/10.3390/foods10051057