
Safety and Transfer Study: Transfer of Bromoform Present in Asparagopsis taxiformis to Milk and Urine of Lactating Dairy Cows



Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design, Diets, Feeding, and Housing
2.2. Sample Collection and Measurements
2.3. Method for Bromoform Analysis
2.4. Data Analysis
3. Results
3.1. Experimental Development
3.2. CHBr3 Measurements
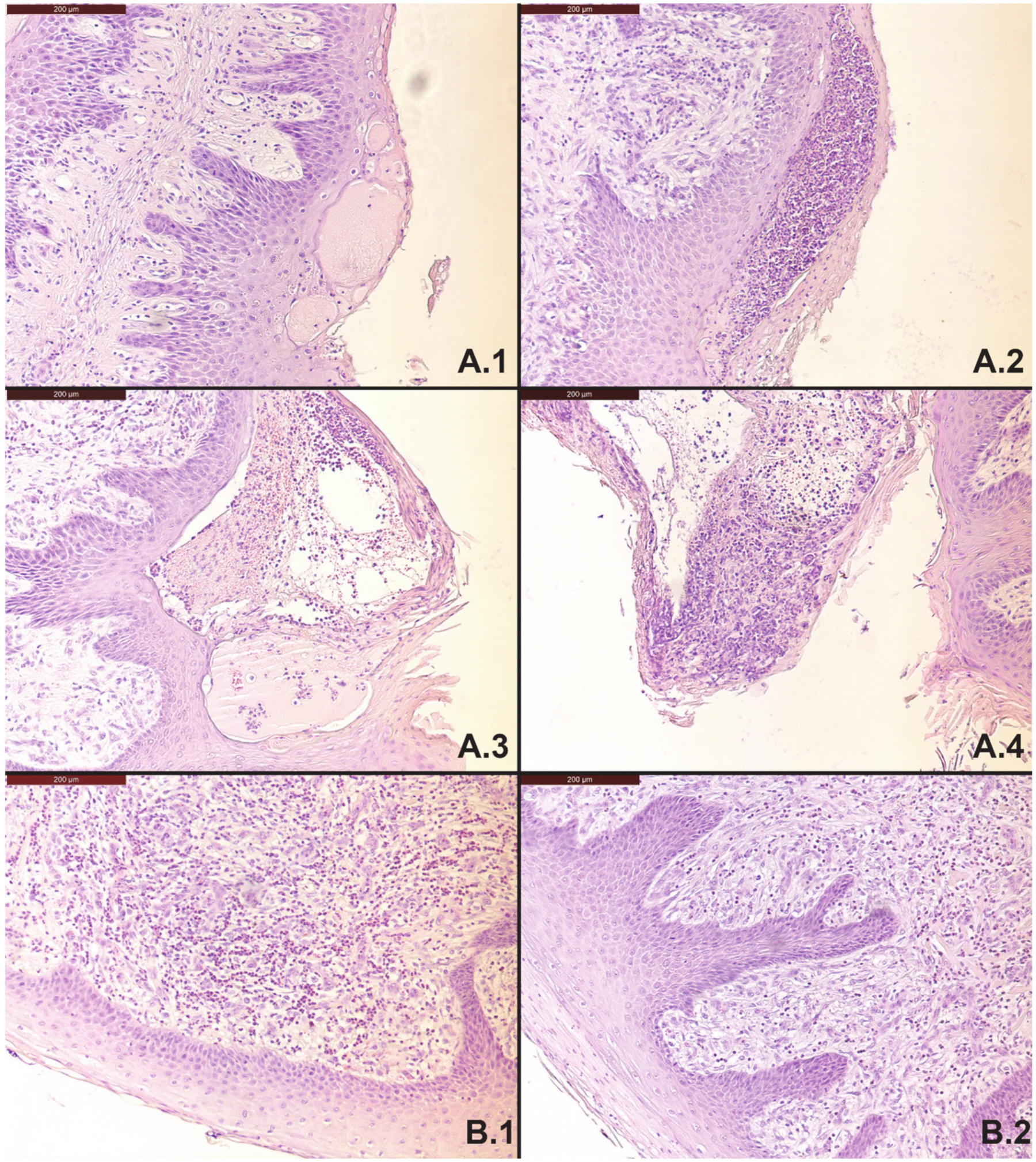
3.3. Pathology and Histology
3.4. Feed Intake
3.5. Milk Yield and Composition
4. Discussion
4.1. Bromoform Excretion and Residues
4.2. Effect on the Rumen Wall
4.3. Feed Intake and Refusal
4.4. Milk Yield and Composition
4.5. Concerns for Human Consumption
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hristov, A.N.; Oh, J.; Firkins, J.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Makkar, H.P.S.; Adesogan, A.T.; Yang, W.; Lee, C.; et al. Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef]
- Lanigan, G.W. Metabolism of pyrrolizidine alkaloids in the ovine rumen. IV. Effects of chloral hydrate and halogenated methanes on rumen methanogenesis and alkaloid metabolism in fistulated sheep. Aust. J. Agric. Res. 1972, 23, 1085–1091. [Google Scholar] [CrossRef]
- Machado, L.; Magnusson, M.; Paul, N.A.; De Nys, R.; Tomkins, N. Effects of marine and freshwater macroalgae on in vitro total gas and methane production. PLoS ONE 2014, 9, e85289. [Google Scholar] [CrossRef]
- Machado, L.; Tomkins, N.; Magnusson, M.; Midgley, D.J.; de Nys, R.; Rosewarne, C.P. In Vitro Response of Rumen Microbiota to the Antimethanogenic Red Macroalga Asparagopsis taxiformis. Microb. Ecol. 2018, 75, 811–818. [Google Scholar] [CrossRef]
- Kinley, R.D.; Vucko, M.J.; de Nys, R.; Machado, L.; Tomkins, N.W. The red macroalgae Asparagopsis taxiformis is a potent natural antimethanogenic that reduces methane production during in vitro fermentation with rumen fluid. Anim. Prod. Sci. 2016, 56, 282–289. [Google Scholar] [CrossRef]
- Machado, L.; Magnusson, M.; Paul, N.A.; Kinley, R.; de Nys, R.; Tomkins, N. Dose-response effects of Asparagopsis taxiformis and Oedogonium sp. on in vitro fermentation and methane production. J. Appl. Phycol. 2016, 28, 1443–1452. [Google Scholar] [CrossRef]
- Machado, L.; Magnusson, M.; Paul, N.A.; Kinley, R.; de Nys, R.; Tomkins, N. Identification of bioactives from the red seaweed Asparagopsis taxiformis that promote antimethanogenic activity in vitro. J. Appl. Phycol. 2016, 28, 3117–3126. [Google Scholar] [CrossRef]
- Wood, J.M.; Kennedy, F.S.; Wolfe, R.S. The Reaction of Multihalogenated Hydrocarbons with Free and Bound Reduced Vitamin B 12. Biochemistry 1968, 7, 1707–1713. [Google Scholar] [CrossRef] [PubMed]
- Kinley, R.D.; Martinez-Fernandez, G.; Matthews, M.K.; de Nys, R.; Magnusson, M.; Tomkins, N.W. Mitigating the carbon footprint and improving productivity of ruminant livestock agriculture using a red seaweed. J. Clean. Prod. 2020, 259, 120836. [Google Scholar] [CrossRef]
- Li, X.; Hayley, N.; Kinley, R.D.; Laurence, M.; Wilmot, M.; Bender, H.; de Nys, R.; Tomkins, N.W. Asparagopsis taxiformis decreases enteric methane production from sheep. Anim. Prod. Sci. 2018, 58, 681–688. [Google Scholar] [CrossRef]
- Roque, B.M.; Salwen, J.K.; Kinley, R.; Kebreab, E. Inclusion of Asparagopsis armata in lactating dairy cows’ diet reduces enteric methane emission by over 50 percent. J. Clean. Prod. 2019, 234, 132–138. [Google Scholar] [CrossRef]
- Anders, M.W.; Stevens, J.L.; Sprague, R.W.; Shaath, Z.; Ahmed, A.E. Metabolism of haloforms to carbon monoxide. II. In Vivo Studies. Amercian Soc. Pharmacol. Exp. Ther. 1978, 6, 556–560. [Google Scholar] [CrossRef]
- Condie, L.W.; Smallwood, C.L.; Laurie, R.D. Comparative Renal and Hepatotoxicity of Halomethanes: Bromodichloromethane, Bromoform, Chloroform, Dibromochloromethane and Methylene Chloride. Drug Chem. Toxicol. 1983, 6, 563–578. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Additives and Products or Substances Used in Animal Feed (FEEDAP). Guidance for establishing the safety of additives for the consumer. EFSA J. 2012, 10, 2537. [Google Scholar] [CrossRef][Green Version]
- Biancarosa, I.; Espe, M.; Bruckner, C.G.; Heesch, S.; Liland, N.; Waagbø, R.; Torstensen, B.; Lock, E.J. Amino acid composition, protein content, and nitrogen-to-protein conversion factors of 21 seaweed species from Norwegian waters. J. Appl. Phycol. 2017, 29, 1001–1009. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Abrahamse, P.A.; Dijkstra, J.; Vlaeminck, B.; Tamminga, S. Frequent allocation of rotationally grazed dairy cows changes grazing behavior and improves productivity. J. Dairy Sci. 2008, 91, 2033–2045. [Google Scholar] [CrossRef] [PubMed]
- Mink, F.L.; Brown, T.J.; Rickabaugh, J. Partition coefficients for the trihalomethanes among blood, urine, water, milk and air. Sci. Total Environ. 2002, 284, 237–247. [Google Scholar] [CrossRef]
- Batterman, S.; Zhang, L.; Wang, S.; Franzblau, A. Absorption, distribution, and excretion of 14 C-trihalomethanes in mice and rats. Bull. Environ. Contam. Toxicol. 1986, 37, 752–758. [Google Scholar] [CrossRef]
- Ahmed, A.E.; Kubic, V.L.; Anders, M.W. Metabolism of haloforms to carbon monoxide. I. In vitro studies. Drug Metab. Dispos. 1977, 5, 198–204. [Google Scholar] [PubMed]
- Stevens, J.L.; Anders, M.W. Metabolism of haloforms to carbon monoxide-III. Studies on the mechanism of the reaction. Biochem. Pharmacol. 1979, 28, 3189–3194. [Google Scholar] [CrossRef]
- Stevens, J.L.; Anders, M.W. Metabolism of haloforms to carbon monoxide. IV. Studies on the reaction mechanism in vivo. Chem. Biol. Interact. 1981, 37, 365–374. [Google Scholar] [CrossRef]
- Parra, P.; Martinez, E.; SuÑol, C.; Artigas, F.; Tusell, J.M.; Gelpl, E.; Albaies, J. Analysis, Accumulation and Central Effects of Trihalomethanes. I. Bromoform. Toxicol. Environ. Chem. 1986, 11, 79–91. [Google Scholar] [CrossRef]
- Stefenoni, H.; Räisänen, S.; Melgar, A.; Lage, C.; Young, M.; Hristov, A. Dose-response effect of the macroalga Asparagopsis taxiformis on enteric methane emission in lactating dairy cows. Ruminant Nutrition: Ruminal Fermentation. In Proceedings of the American Dairy Science Association Annual Meeting, Cincinnati, OH, USA, 23–26 June 2019. [Google Scholar]
- Jensen, C.; Østergaard, S.; Schei, I.; Bertilsson, J.; Weisbjerg, M.R. A meta-analysis of milk production responses to increased net energy intake in Scandinavian dairy cows. Livest. Sci. 2015, 175, 59–69. [Google Scholar] [CrossRef]
- Van Knegsel, A.T.M.; van den Brand, H.; Dijkstra, J.; Tamminga, S.; Kemp, B. Effect of dietary energy source on energy balance, production, metabolic disorders and reproduction in lactating dairy cattle. Reprod. Nutr. Dev. 2005, 45, 665–688. [Google Scholar] [CrossRef] [PubMed]
- European Parliament. Commission Directive 2015/1787 amendment to Council Directive 98/83/EC on the quality of water intended for human consumption. Off. J. Eur. Communities 2015, 58, 6–17. [Google Scholar]
- United States Environmental Protection Agency. National Primary Drinking Water Regulations. Available online: https://web.archive.org/web/20210225100053/https://www.epa.gov/ground-water-and-drinking-water/national-primary-drinking-water-regulations (accessed on 25 February 2021).
- Burreson, B.J.; Moore, R.E.; Roller, P.P. Volatile Halogen Compounds in the Alga Asparagopsis taxiformis (Rhodophyta). J. Agric. Food Chem. 1976, 24, 856–861. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ingredients | Low | Medium | High |
|---|---|---|---|
| A. taxiformis (g DM) | 67 | 133 | 333 |
| Dextrose (g DM) | 67 | 133 | 333 |
| Wheat (g as fed basis) | 500 | 500 | 500 |
| Dehydrated beet pulp (g as fed basis) | 500 | 500 | 500 |
| Water (liters) | 2 | 2 | 2 |
| Nutrients | Grass Silage 2 | Maize Silage | Soybean Meal | Compound Feed 3 | Wheat Straw | A. taxiformis |
|---|---|---|---|---|---|---|
| DM (g/kg) | 466 | 348 | 874 | 878 | 886 | 951 |
| Ash | 121 | 47 | 70 | 45 | 67 | 510 |
| Crude Protein | 203 | 69 | 513 | 115 | 36 | 146 4 |
| Crude Fat | 43 | 30 | 13 | 10 | 11 | 7.5 |
| Crude Fiber | 219 | 181 | 46 | 100 | 404 | 65 |
| NDF | 430 | 377 | 242 | 257 | 757 | ND |
| ADF | 237 | 209 | 142 | 123 | 454 | ND |
| ADL | 14 | 14 | 4 | 8 | 51 | ND |
| Reducing Sugar | 95 | 12 | 117 | 128 | 30 | 11 |
| Starch | ND | 323 | ND | ND | ND | ND |
| CHBr3 (mg/kg DM) | ND | ND | ND | ND | ND | 1.26 |
| Treatments | ||||
|---|---|---|---|---|
| Day | Low | Medium | High | |
| CHBr3 Milk | −2 | <5 | < 5 | <5 |
| (µg/L) | −1 | <5 | <5 | <5 |
| 1 | 9.1 1 | 11 | <5 | |
| 9 | <5 | <5 | 35 2 | |
| 10 | <5 | <5 | <5 | |
| 17 | <5 3 | <53 | <5 3 | |
| CHBr3 Urine | −2 | <1 | <1 | <1 |
| (µg/L) | −1 | <1 | <1 | <1 |
| 1 | 10 | 50 | 114 | |
| 10 | 89 4 | 148 | 127 | |
| 17 | <2 3,5 | <2 3,5 | <2 3,5 | |
| CHBr3 Feces | −2 | <20 | <20 | <20 |
| (µg/kg) | −1 | <20 | <20 | <20 |
| 1 | <20 | <20 | <20 | |
| 10 | <20 | <20 | <20 | |
| 17 | <20 | <20 | <20 | |
| Treatments | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low | Medium | High | ||||||||||
| Cow | L1 | L2 | L3 | L4 | L5 | L6 | L7 | L8 | M1 | M2 | H1 | H2 |
| Parity | 2 | 1 | 1 | 1 | 1 | 5 | 1 | 1 | 4 | 4 | 2 | 1 |
| DIM | 281 | 577 | 538 | 326 | 344 | 521 | 159 | 126 | 280 | 348 | 281 | 282 |
| Day | ||||||||||||
| −7 | 1 | 12.3 | 12.2 | 13.5 | 13.2 | 8.4 | 10.2 | 10.1 | 14.1 | 13.7 | 12.9 | 11.3 |
| −6 | 1 | 15.8 | 16.4 | 19.2 | 15.8 | 18.1 | 16.2 | 13.1 | 20.5 | 18.6 | 17.7 | 15.4 |
| −5 | 1 | 13.2 | 16.2 | 18.2 | 14.5 | 17.8 | 15.3 | 10.1 | 19.2 | 18.1 | 15.3 | 14.6 |
| −4 | 1 | 16.1 | 15.7 | 16.2 | 15.5 | 19.6 | 17.9 | 11.4 | 15.6 | 16.6 | 15.8 | 16.0 |
| −3 | 1 | 14.9 | 16.9 | 17.4 | 15.3 | 18.5 | 15.0 | 14.4 | 17.8 | 17.0 | 15.3 | 14.4 |
| −2 | 18.0 | 14.2 | 15.5 | 18.0 | 13.4 | 19.8 | 16.6 | 14.3 | 17.4 | 18.4 | 15.3 | 16.6 |
| −1 | 17.2 | 15.3 | 14.7 | 17.7 | 8.9 | 18.2 | 14.2 | 14.3 | 18.4 | 15.8 | 15.5 | 13.8 |
| 1 | 16.8 | 16.3 | 16.3 | 16.8 | 9.3 | 19.8 | 15.7 | 14.7 | 15.9 | 17.6 | 10.3 | 5.4 |
| 2 | 17.0 | 13.5 | 14.5 | 16.3 | 13.8 | 20.1 | 15.0 | 13.2 | 13.6 | 15.9 | 15.2 | 6.6 |
| 3 | 14.6 | 12.9 | 14.0 | 12.8 | 12.1 | 16.1 | 12.8 | 12.0 | 11.2 | 16.1 | 14.3 | 11.3 |
| 4 | 18.5 | 15.4 | 16.3 | 13.2 | 14.5 | 20.7 | 13.8 | 11.5 | 15.9 | 19.0 | 16.5 | 14.0 |
| 5 | 15.5 | 16.3 | 16.6 | 15.6 | 9.9 | 19.3 | 11.4 | 12.8 | 15.8 | 17.6 | 17.5 | 15.4 |
| 6 | 20.3 | 15.7 | 14.0 | 16.7 | 17.7 | 22.0 | 9.5 | 13.2 | 16.1 | 20.0 | 15.5 | 18.5 |
| 7 2 | 9.9 | 9.9 | 9.9 | 10.0 | 9.9 | 10.0 | 5.0 | 5.0 | 4.8 | 4.9 | 3.6 | 3.1 |
| 8 2 | 14.7 | 9.8 | 9.9 | 4.0 | 9.8 | 14.8 | 4.9 | 14.3 | 2.1 | 3.0 | 3.1 | 1.2 |
| 9 2 | 14.3 | 7.0 | 11.0 | 10.2 | 13.8 | 19.5 | 7.6 | 7.7 | 2.8 | 8.6 | 7.7 | 5.3 |
| 10 2 | 9.6 | 9.8 | 9.8 | 9.8 | 9.6 | 19.6 | 9.8 | 9.6 | 8.1 | 9.8 | 9.7 | 9.8 |
| 11 | 13.6 | 13.3 | 14.9 | 12.0 | 14.6 | 14.8 | 14.5 | 14.7 | 8.8 | 14.9 | 8.9 | 11.4 |
| 12 | 14.2 | 14.6 | 14.4 | 14.8 | 14.7 | 14.8 | 14.7 | 14.8 | 5.6 | 14.8 | 13.4 | 9.9 |
| 13 | 4.7 | 14.6 | 14.0 | 14.7 | 14.6 | 9.7 | 4.7 | 4.8 | 3 | 14.7 | 14.7 | 4 |
| 14 | 17.4 | 18.8 | 18.7 | 19.3 | 19.0 | 19.3 | 17.8 | 18.5 | 3 | 19.7 | 18.1 | 11.2 |
| 15 | 17.1 | 16.2 | 17.4 | 18.8 | 17.6 | 19.6 | 18.5 | 18.5 | 3 | 16.9 | 16.6 | 13.0 |
| 16 | 23.9 | 17.2 | 17.7 | 24.1 | 18.0 | 27.8 | 23.0 | 21.9 | 3 | 19.7 | 16.7 | 22.4 |
| 17 | 19.8 | 16.3 | 19.3 | 21.9 | 20.4 | 20.5 | 18.7 | 22.1 | 3 | 20.2 | 17.0 | 17.5 |
| 18 | 16.5 | 16.3 | 16.3 | 20.7 | 16.1 | 21.0 | 15.8 | 16.2 | 3 | 20.6 | 15.7 | 17.6 |
| 19 | 19.3 | 17.5 | 19.0 | 19.6 | 19.4 | 24.7 | 16.6 | 19.2 | 3 | 19.1 | 18.7 | 16.7 |
| Treatments | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low | Medium | High | ||||||||||
| Cow | L1 | L2 | L3 | L4 | L5 | L6 | L7 | L8 | M1 | M2 | H1 | H2 |
| Day | ||||||||||||
| −6 | 1 | 11.7 | 10.4 | 18.3 | 10.0 | 11.8 | 7.8 | 7.6 | 22.9 | 12.7 | 15.3 | 19.9 |
| −5 | 1 | 12.1 | 10.7 | 16.7 | 16.4 | 16.6 | 19.9 | 13.2 | 22.5 | 13.1 | 14.7 | 18.3 |
| −4 | 1 | 11.6 | 9.9 | 15.9 | 15.1 | 13.0 | 21.1 | 13.4 | 23.8 | 11.7 | 16.8 | 17.4 |
| −3 | 1 | 14.0 | 9.6 | 14.8 | 19.3 | 14.7 | 21.0 | 13.5 | 23.6 | 11.7 | 18.8 | 19.7 |
| −2 | 7.05 | 13.7 | 10.5 | 14.6 | 17.1 | 14.9 | 21.3 | 8.9 5 | 23.4 | 11.7 | 19.5 | 20.2 |
| −1 | 22.6 | 14.5 | 8.5 | 16.6 | 17.0 | 16.0 | 22.2 | 16.3 | 20.4 | 11.8 | 18.4 | 19.5 |
| 1 | 22.9 | 13.3 | 8.6 | 17.5 | 16.4 | 15.2 | 21.4 | 16.0 | 22.0 | 11.6 | 17.1 | 18.9 |
| 2 | 23.6 | 13.2 | 9.2 | 15.7 | 16.6 | 15.0 | 21.8 | 15.6 | 21.8 | 11.5 | 13.1 | 13.6 |
| 3 | 23.1 | 12.9 | 13.9 | 18.9 | 18.7 | 15.3 | 21.4 | 15.8 | 20.9 | 13.6 | 17.8 | 13.1 |
| 4 | 22.4 | 16.3 | 14.9 | 20.8 | 20.6 | 13.5 | 20.3 | 16.2 | 19.6 | 14.5 | 21.8 | 16.4 |
| 5 | 22.8 | 14.6 | 14.8 | 21.5 | 19.5 | 15.0 | 19.6 | 16.4 | 18.5 | 12.8 | 22.5 | 17.8 |
| 6 | 22.2 | 15.6 | 15.0 | 20.4 | 20.1 | 14.4 | 19.3 | 13.2 | 15.8 | 15.5 | 19.8 | 18.0 |
| 7 2 | 23.1 | 16.4 | 14.2 | 23.8 | 20.9 | 14.4 | 16.4 | 15.5 | 17.3 | 14.5 | 18.3 | 19.1 |
| 8 2 | 21.6 | 13.4 | 12.4 | 14.3 | 19.0 | 12.0 | 14.8 | 13.3 | 15.2 | 9.4 | 10.5 | 15.4 |
| 9 2 | 21.2 | 13.7 | 10.1 | 13.0 | 17.1 | 13.1 | 13.9 | 15.6 | 11.3 | 7.7 | 9.7 | 10.4 |
| 10 2 | 19.6 | 8.5 | 7.7 | 15.3 | 17.6 | 12.4 | 15.5 | 15.5 | 9.6 | 5.6 | 7.3 | 12.0 |
| 11 | 18.3 | 10.1 | 13.4 | 6.4 | 15.9 | 15.0 | 15.9 | 15.1 | 11.5 | 6.8 | 10.1 | 13.8 |
| 12 | 18.6 | 11.7 | 12.4 | 9.4 | 16.2 | 11.4 | 16.3 | 14.8 | 9.5 | 9.0 | 4.3 5 | 14.3 |
| 13 | 18.4 | 12.1 | 12.7 | 19.7 | 16.6 | 13.1 | 17.5 | 15.7 | 4 | 6.8 | 10.6 | 14.3 |
| 14 | 22.2 | 13.6 | 14.6 | 16.4 | 20.7 | 9.1 | 18.4 | 17.0 | 4 | 9.0 | 14.0 | 13.6 |
| 15 | 21.2 | 14.0 | 14.6 | 18.4 | 18.4 | 3.4 5 | 14.8 | 16.4 | 4 | 9.9 | 7.1 5 | 14.1 |
| 16 | 3 | 3 | 14.6 | 20.0 | 20.7 | 3.9 5 | 18.4 | 16.7 | 4 | 10.3 | 3 | 14.8 |
| 17 | 9.4 5 | 8.9 5 | 15.8 | 21.5 | 20.3 | 7.1 | 19.7 | 15.9 | 4 | 10.5 | 8.4 5 | 16.0 |
| 18 | 22.3 | 14.2 | 14.9 | 21.0 | 20.4 | 14.2 | 20.6 | 16.1 | 4 | 12.0 | 17.2 | 16.6 |
| 19 | 21.6 | 16.5 | 13.6 | 22.8 | 21.3 | 8.1 | 20.6 | 16.3 | 4 | 13.1 | 18.8 | 16.3 |
| Treatments | ||||
|---|---|---|---|---|
| Day | Low | Medium 1 | High | |
| Yield (gram/day) | ||||
| Fat | 1 | 806 | 788 | 771 |
| 8 | 746 | 752 | 730 | |
| 15 | 733 | 595 | 485 | |
| Protein | 1 | 652 | 695 | 650 |
| 8 | 608 | 521 | 493 | |
| 15 | 595 | 436 | 385 | |
| Lactose | 1 | 738 | 751 | 838 |
| 8 | 671 | 504 | 564 | |
| 15 | 677 | 379 | 474 | |
| Milk composition (%) | ||||
| Fat | 1 | 4.91 | 4.69 | 4.28 |
| 8 | 4.94 | 6.12 | 5.64 | |
| 15 | 4.84 | 6.01 | 4.57 | |
| Protein | 1 | 3.97 | 4.14 | 3.61 |
| 8 | 4.02 | 4.23 | 3.81 | |
| 15 | 3.93 | 4.40 | 3.63 | |
| Lactose | 1 | 4.49 | 4.47 | 4.65 |
| 8 | 4.45 | 4.10 | 4.35 | |
| 15 | 4.47 | 3.83 | 4.47 | |
| Urea (mg/dL) | 1 | 17.6 | 18.5 | 19.6 |
| 8 | 23.1 | 22.9 | 23.0 | |
| 15 | 14.2 | 13.3 | 14.6 | |
| SCC (cells/mL) | 1 | 56,709 | 64,841 | 36,325 |
| 8 | 66,923 | 276,219 | 50,757 | |
| 15 | 65,742 | 170,338 | 63,600 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muizelaar, W.; Groot, M.; van Duinkerken, G.; Peters, R.; Dijkstra, J. Safety and Transfer Study: Transfer of Bromoform Present in Asparagopsis taxiformis to Milk and Urine of Lactating Dairy Cows. Foods 2021, 10, 584. https://doi.org/10.3390/foods10030584
Muizelaar W, Groot M, van Duinkerken G, Peters R, Dijkstra J. Safety and Transfer Study: Transfer of Bromoform Present in Asparagopsis taxiformis to Milk and Urine of Lactating Dairy Cows. Foods. 2021; 10(3):584. https://doi.org/10.3390/foods10030584
Chicago/Turabian StyleMuizelaar, Wouter, Maria Groot, Gert van Duinkerken, Ruud Peters, and Jan Dijkstra. 2021. "Safety and Transfer Study: Transfer of Bromoform Present in Asparagopsis taxiformis to Milk and Urine of Lactating Dairy Cows" Foods 10, no. 3: 584. https://doi.org/10.3390/foods10030584
APA StyleMuizelaar, W., Groot, M., van Duinkerken, G., Peters, R., & Dijkstra, J. (2021). Safety and Transfer Study: Transfer of Bromoform Present in Asparagopsis taxiformis to Milk and Urine of Lactating Dairy Cows. Foods, 10(3), 584. https://doi.org/10.3390/foods10030584