Fatty Acid Methyl Ester (FAME) Profiling Identifies Carbapenemase-Producing Klebsiella pneumoniae Belonging to Clonal Complex 258

 , ,
, ,
Abstract
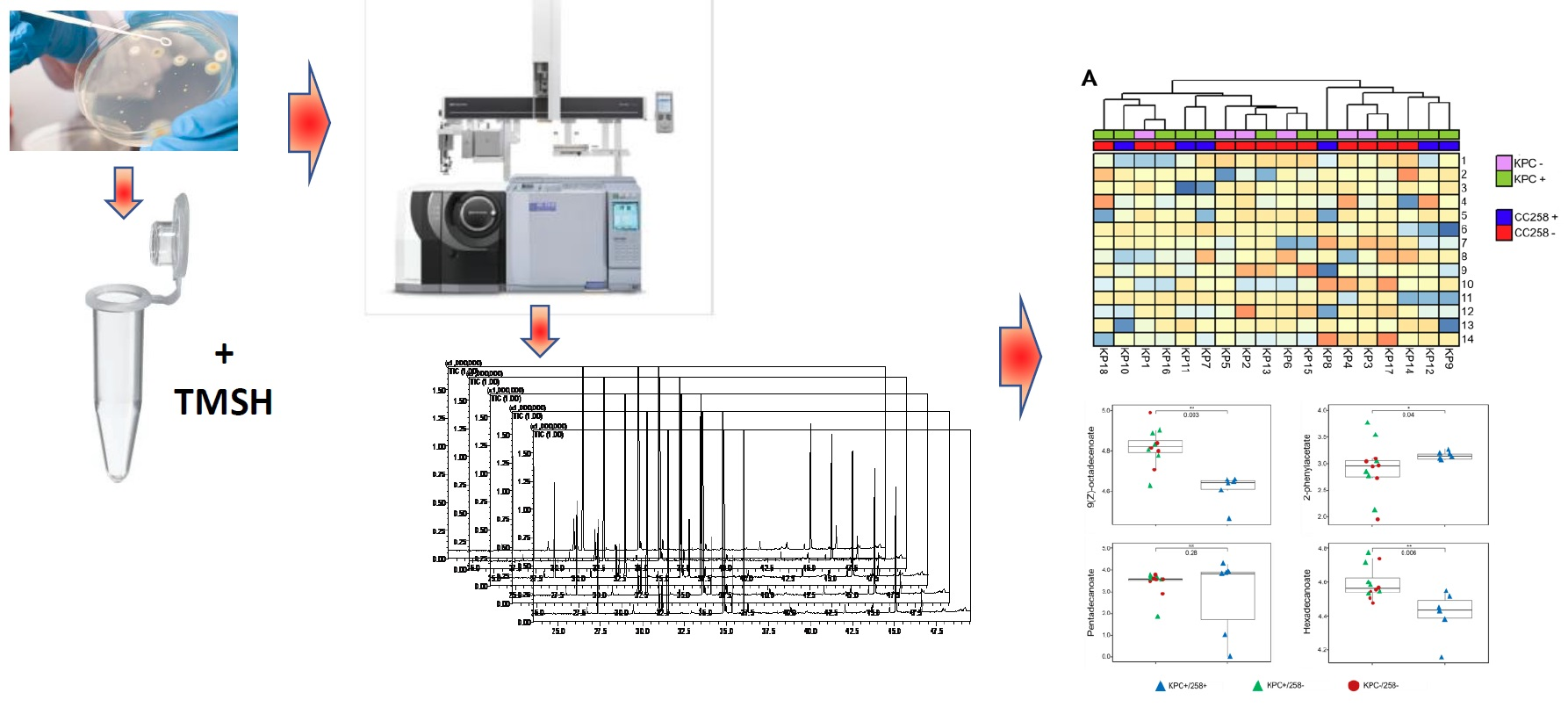
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Bacterial Strains, Culture Conditions, and Sample Preparation
2.3. Analytical Instrumentation
2.4. Data Elaboration and Statistical Analysis
3. Results and Discussion
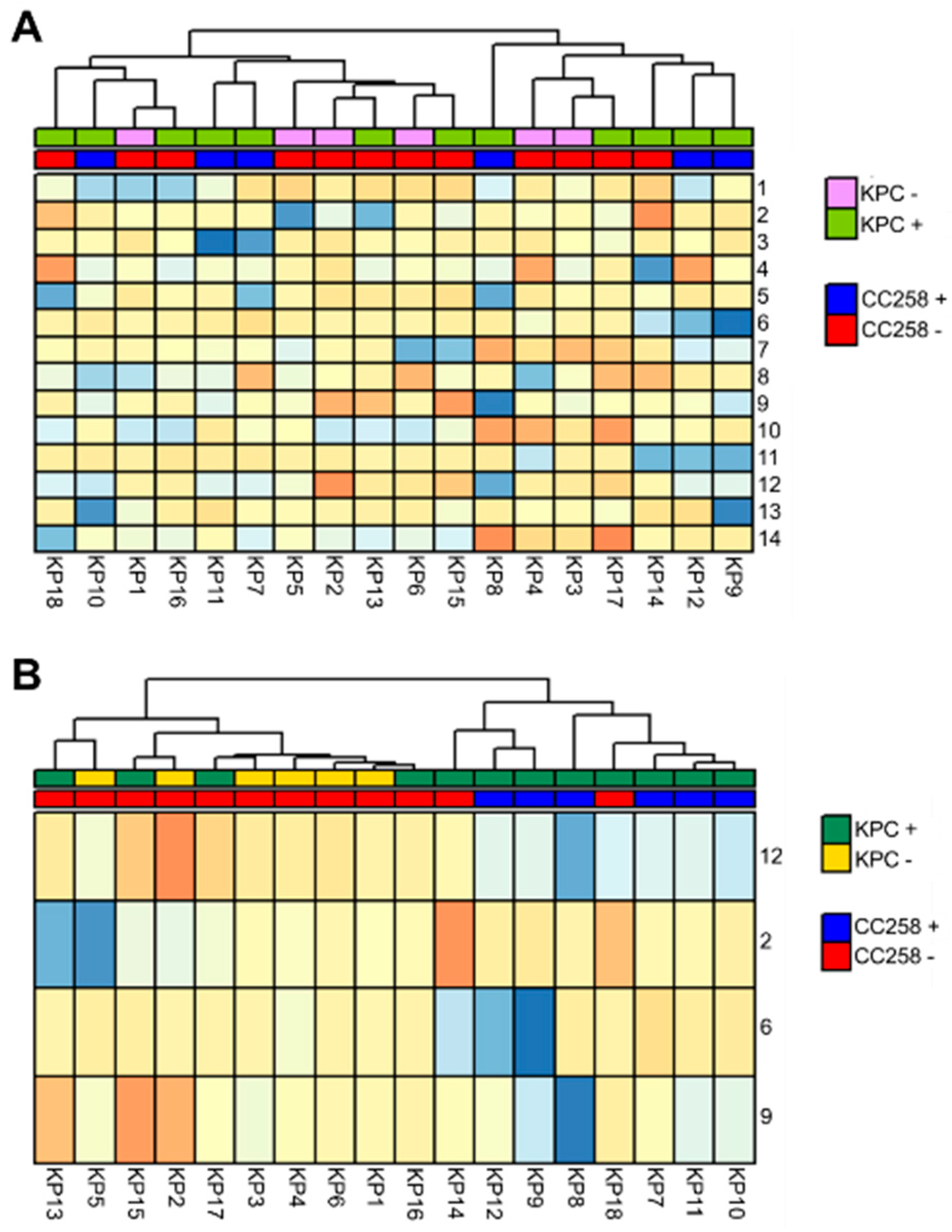
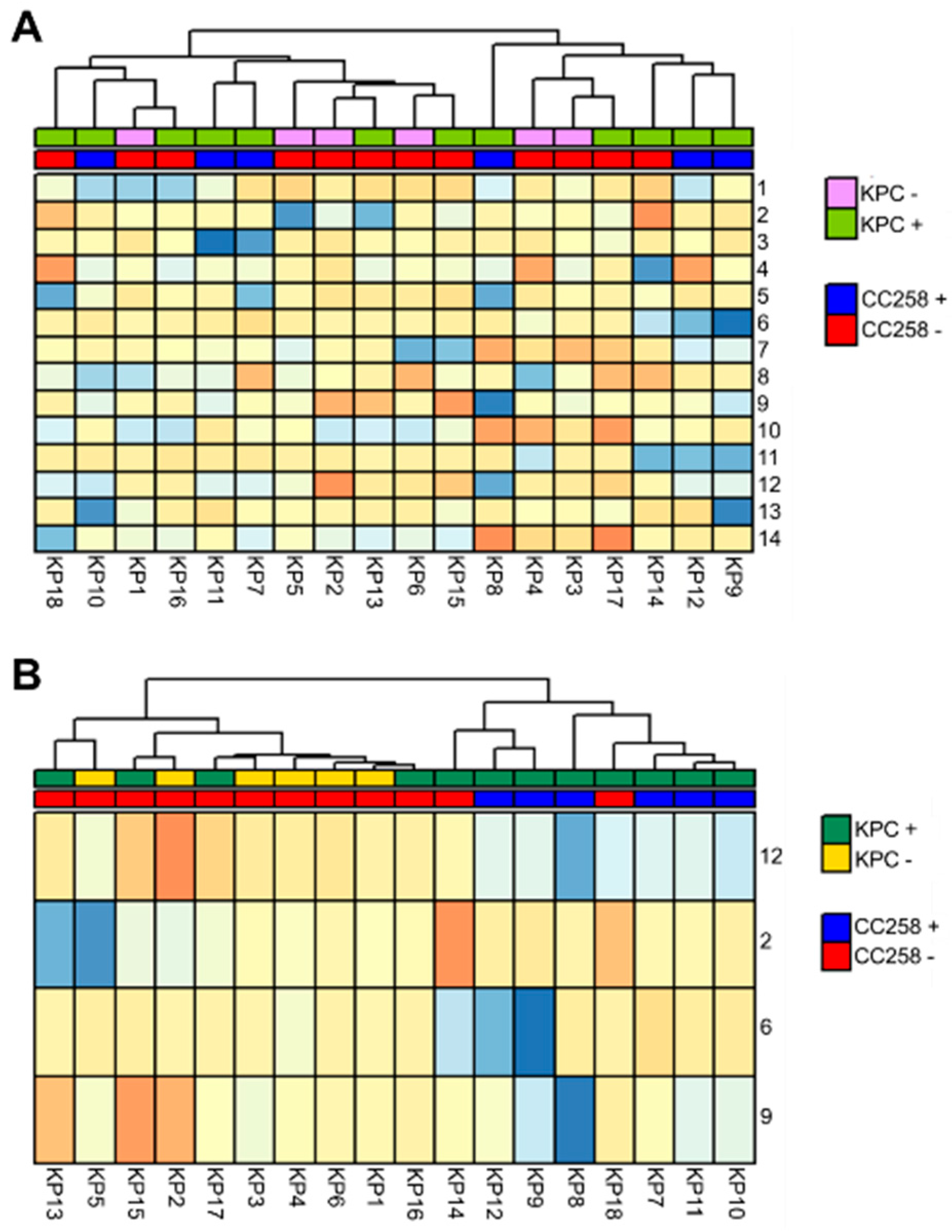
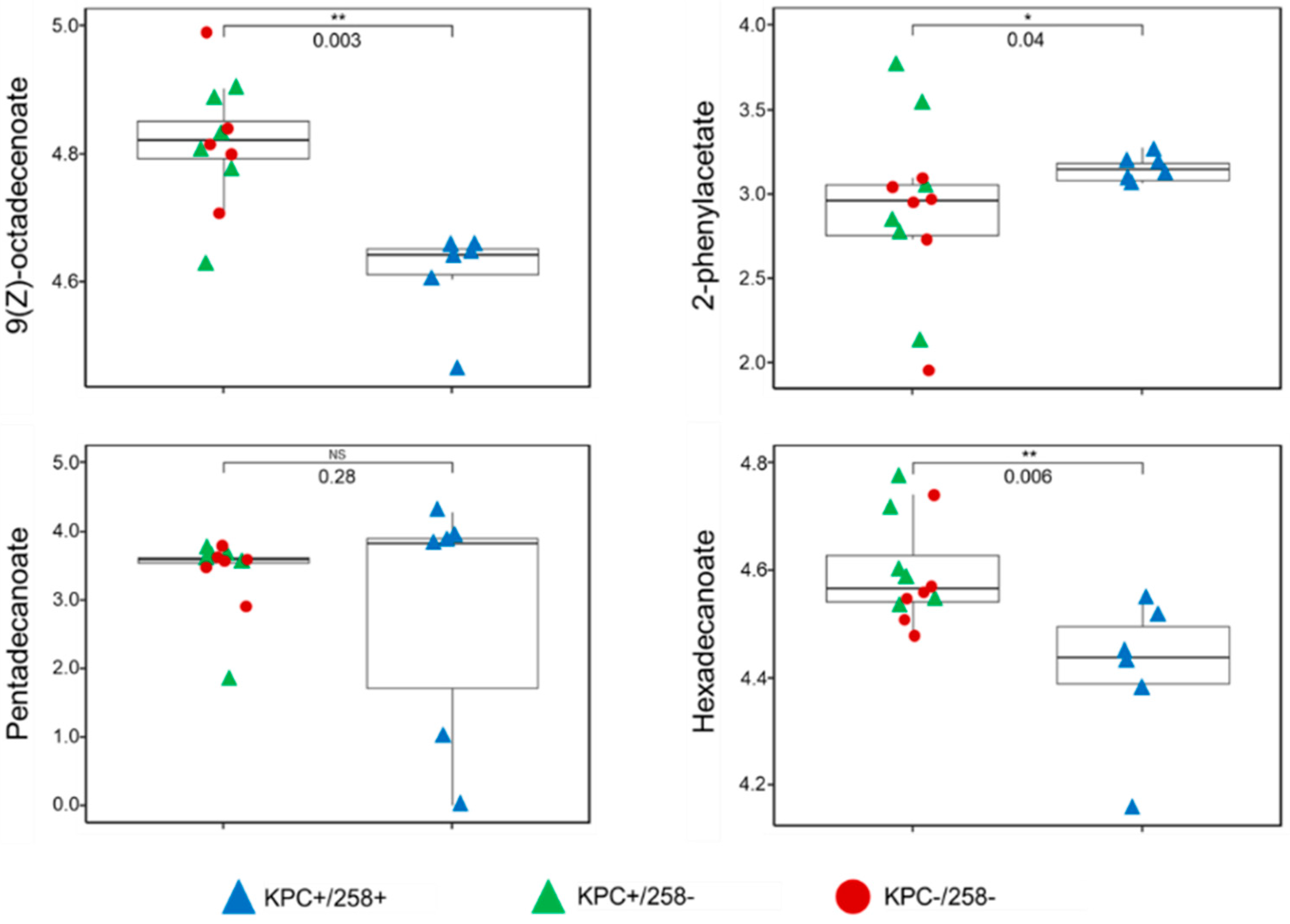
3.1. The FAME Profile of Klebsiella pneumoniae Clinical Isolates
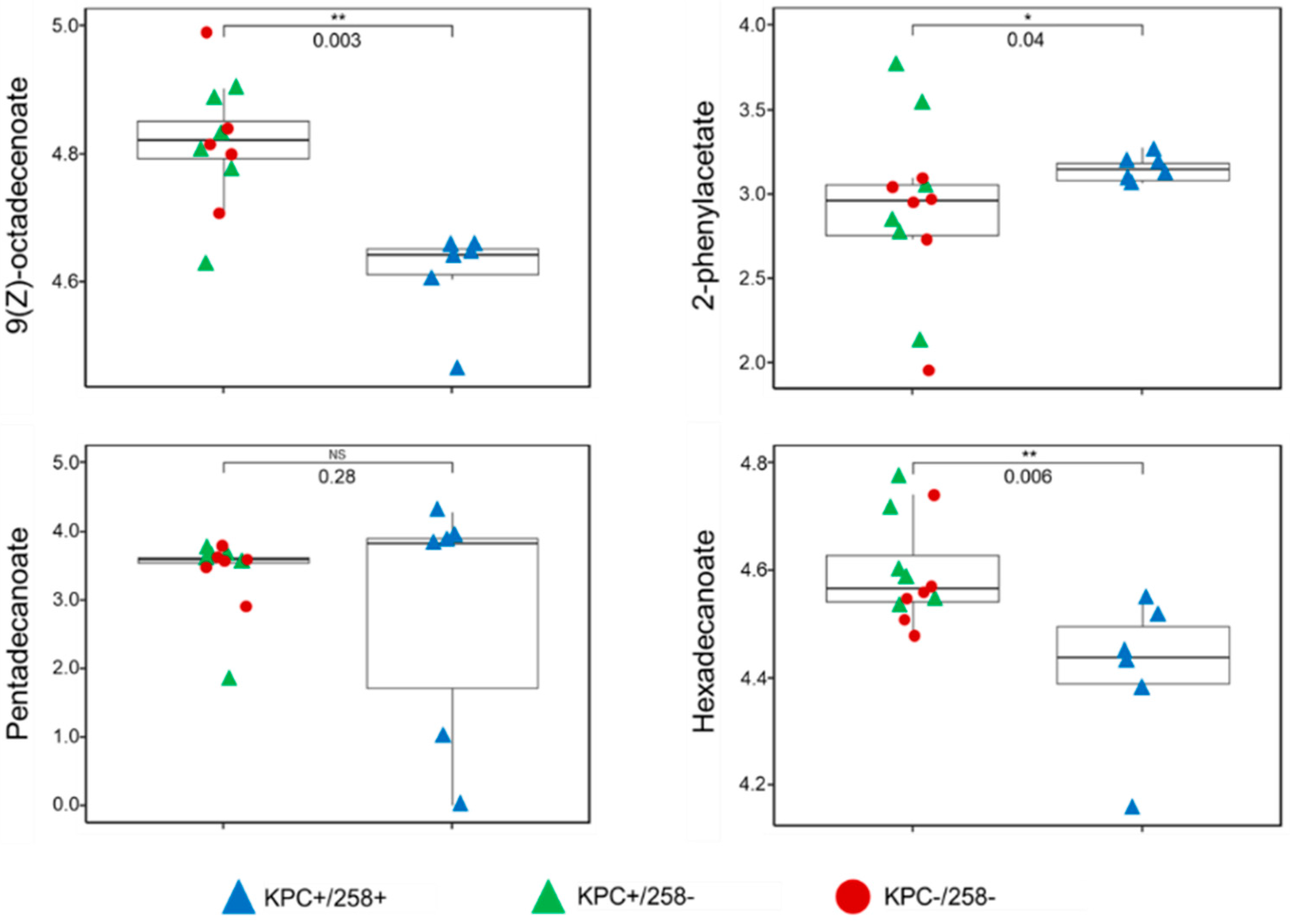
3.2. Discrimination between CC258 and Non-CC258 via FAME Profiles
3.3. Study Strengths and Limitations
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Todd, R.; Kiehlbauch, J.; Walters, M.; Kallen, A. Pan-Resistant New Delhi Metallo-Beta-Lactamase-Producing Klebsiella pneumoniae-Washoe County, Nevada, 2016. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 33. [Google Scholar] [CrossRef]
- Elemam, A.; Rahimian, J.; Mandell, W. Infection with Panresistant Klebsiella pneumoniae: A Report of 2 Cases and a Brief Review of the Literature. Clin. Infect. Dis. 2009, 49, 271–274. [Google Scholar] [CrossRef]
- Sonnevend, Á.; Ghazawi, A.; Hashmey, R.; Haidermota, A.; Girgis, S.; Alfaresi, M.; Omar, M.; Paterson, D.L.; Zowawi, H.M.; Pál, T. Multihospital Occurrence of Pan-Resistant Klebsiella pneumoniae Sequence Type 147 with an ISEcp1-Directed blaOXA-181 Insertion in the mgrB Gene in the United Arab Emirates. Antimicrob. Agents Chemother. 2017, 61, e00418-17. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.D.; Goodman, K.E.; Harris, A.D.; Tekle, T.; Roberts, A.; Taiwo, A.; Simner, P.J.; Tamma, P.D.; Corresponding, M.H.S. Comparing the Outcomes of Patients with Carbapenemase-Producing and Non-Carbapenemase-Producing Carbapenem-Resistant Enterobacteriaceae Bacteremia. Clin. Infect. Dis. 2017, 64, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Van Duin, D.; Doi, Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence 2017, 8, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Kitchel, B.; Rasheed, J.K.; Patel, J.B.; Srinivasan, A.; Navon-Venezia, S.; Carmeli, Y.; Brolund, A.; Giske, C.G. Molecular epidemiology of KPC-producing Klebsiella pneumoniae isolates in the United States: Clonal expansion of multilocus sequence type 258. Antimicrob. Agents Chemother. 2009, 53, 3365–3370. [Google Scholar] [CrossRef] [PubMed]
- Adler, A.; Paikin, S.; Sterlin, Y.; Glick, J.; Edgar, R.; Aronov, R.; Schwaber, M.J.; Carmelia, Y. A swordless knight: Epidemiology and molecular characteristics of the blaKPC-negative sequence type 258 Klebsiella pneumoniae clone. J. Clin. Microbiol. 2012, 50, 3180–3185. [Google Scholar] [CrossRef]
- Bowers, J.R.; Kitchel, B.; Driebe, E.M.; MacCannell, D.R.; Roe, C.; Lemmer, D.; De Man, T.; Rasheed, J.K.; Engelthaler, D.M.; Keim, P.; et al. Genomic analysis of the emergence and rapid global dissemination of the clonal group 258 Klebsiella pneumoniae pandemic. PLoS ONE 2015, 10, e0133727. [Google Scholar] [CrossRef]
- Chmelnitsky, I.; Shklyar, M.; Hermesh, O.; Navon-Venezia, S.; Edgar, R.; Carmeli, Y. Unique genes identified in the epidemic extremely drug-resistant KPC-producing Klebsiella pneumoniae sequence type 258. J. Antimicrob. Chemother. 2013, 68, 74–83. [Google Scholar] [CrossRef]
- Bruchmann, S.; Muthukumarasamy, U.; Pohl, S.; Preusse, M.; Bielecka, A.; Nicolai, T.; Hamann, I.; Hillert, R.; Kola, A.; Gastmeier, P.; et al. Deep transcriptome profiling of clinical Klebsiella pneumoniae isolates reveals strain and sequence type-specific adaptation. Environ. Microbiol. 2015, 17, 4690–4710. [Google Scholar] [CrossRef]
- Rees, C.A.; Nasir, M.; Smolinska, A.; Lewis, A.E.; Kane, K.R.; Kossmann, S.E.; Sezer, O.; Zucchi, P.C.; Doi, Y.; Hirsch, E.B.; et al. Detection of high-risk carbapenem-resistant Klebsiella pneumoniae and Enterobacter cloacae isolates using volatile molecular profiles. Sci. Rep. 2018, 8, 13297. [Google Scholar] [CrossRef]
- Low, Y.M.; Yap, I.K.S.; Abdul Jabar, K.; Md Yusof, M.Y.; Chong, C.W.; Teh, C.S.J. Genotypic and metabolic approaches towards the segregation of Klebsiella pneumoniae strains producing different antibiotic resistant enzymes. Metabolomics 2017, 13, 65. [Google Scholar] [CrossRef]
- Müller, K.D.; Nalik, H.P.; Schmid, E.N.; Husmann, H.; Schomburg, G. Fast identification of mycobacterium species by GC analysis with trimethylsulfonium hydroxide (TMSH) for transesterification. J. High Resolut. Chromatogr. 1993, 16, 161–165. [Google Scholar] [CrossRef]
- Breiman, L.E.O. Random Forest. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P.S.B. Multilocus Sequence Typing of Klebsiella pneumoniae Nosocomial Isolates. J. Clin. Microbiol. 2005, 43, 4178–4182. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C.; Kalsi, R.K.; Williams, P.P.; Carey, R.B.; Stocker, S.; Lonsway, D.; Rasheed, J.K.; Biddle, J.W.; McGowan, J.E.; Hanna, B. Carbapenem resistance in Klebsiella pneumoniae not detected by automated susceptibility testing. Emerg. Infect. Dis. 2006, 12, 1209–1213. [Google Scholar] [CrossRef]
- Tranchida, P.Q.; Franchina, F.A.; Dugo, P.; Mondello, L. A flow-modulated comprehensive gas chromatography-mass spectrometry method for the analysis of fatty acid profiles in marine and biological samples. J. Chromatogr. A 2012, 1255, 171–176. [Google Scholar] [CrossRef]
- Purcaro, G.; Tranchida, P.Q.; Dugo, P.; Camera, L.E.; Bisignano, G.; Conte, L.; Mondello, L. Characterization of bacterial lipid profiles by using rapid sample preparation and fast comprehensive two-dimensional gas chromatography in combination with mass spectrometry. J. Sep. Sci. 2010, 33, 2334–2340. [Google Scholar] [CrossRef]
- Beccaria, M.; Franchina, F.A.; Nasir, M.; Mellors, T.; Hill, J.E.; Purcaro, G. Investigation of mycobacteria fatty acid profile using different ionization energies in GC-MS. Anal. Bioanal. Chem. 2018, 410, 7987–7996. [Google Scholar] [CrossRef]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in1H NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef]
- Kruskal, W.H.; Wallis, W.A.; Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Dunn, O.J. Multiple Comparisons Using Rank Sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Fulco, A.J. Fatty acid metabolism in bacteria. Prog. Lipid Res. 1983, 22, 133–160. [Google Scholar] [CrossRef]
- Dunnick, J.K.; O’Leary, W.M. Correlation of bacteria lipid composition with antibiotic resistance. J. Bacteriol. 1970, 101, 892–900. [Google Scholar] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Strain ID | CC258 (+/−) | blaKPC (+/−) | Sequence Type (ST) |
|---|---|---|---|
| KP1 | - | - | 14 |
| KP2 | - | - | 35 |
| KP3 | - | - | 36 |
| KP4 | - | - | 113 |
| KP5 | - | - | 307 |
| KP6 | - | - | 485 |
| KP7 | + | + | 258 |
| KP8 | + | + | 258 |
| KP9 | + | + | 258 |
| KP10 | + | + | 258 |
| KP11 | + | + | 258 |
| KP12 | + | + | 258 |
| KP13 | + | - | 42 |
| KP14 | + | - | 42 |
| KP15 | + | - | 42 |
| KP16 | + | - | 101 |
| KP17 | + | - | 307 |
| KP18 | + | - | 307 |
| # | Compound | CAS | MS% | LRIExp | LRILib |
|---|---|---|---|---|---|
| 1 | Methyl 3-methylsulfanylpropanoate | 13532-18-8 | 81 | 1021 | 1023 |
| 2 | Methyl 2-phenylacetate | 101-41-7 | 91 | 1173 | 1178 |
| 3 | Methyl dodecanoate * | 111-82-0 | 95 | 1522 | 1523 |
| 4 | Methyl tetradecanoate * | 124-10-7 | 94 | 1723 | 1724 |
| 5 | Methyl pentadecanoate a | - | - | 1773 | - |
| 6 | Methyl pentadecanoate * | 7132-64-1 | 90 | 1823 | 1825 |
| 7 | Methyl 3-hydroxytetradecanoate * | 55682-83-2 | 91 | 1869 | 1870 |
| 8 | Methyl 9(Z)-hexadecenoate * | 1120-25-8 | 93 | 1901 | 1903 |
| 9 | Methyl hexadecanoate * | 112-39-0 | 92 | 1925 | 1925 |
| 10 | Methyl 10-heptadecenoate * | 77745-60-9 | 90 | 2009 | 2009 |
| 11 | Methyl heptadecanoate * | 1731-92-6 | 95 | 2024 | 2026 |
| 12 | Methyl 9(Z)-octadecenoate * | 112-62-9 | 92 | 2103 | 2098 |
| 13 | Methyl octadecenoate * | 112-61-8 | 95 | 2124 | 2126 |
| 14 | Methyl cis-9,10-methyleneoctadecanoate * | 3971-54-8 | 92 | 2211 | 2208 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rees, C.A.; Beccaria, M.; Franchina, F.A.; Hill, J.E.; Purcaro, G. Fatty Acid Methyl Ester (FAME) Profiling Identifies Carbapenemase-Producing Klebsiella pneumoniae Belonging to Clonal Complex 258. Separations 2019, 6, 32. https://doi.org/10.3390/separations6020032
Rees CA, Beccaria M, Franchina FA, Hill JE, Purcaro G. Fatty Acid Methyl Ester (FAME) Profiling Identifies Carbapenemase-Producing Klebsiella pneumoniae Belonging to Clonal Complex 258. Separations. 2019; 6(2):32. https://doi.org/10.3390/separations6020032
Chicago/Turabian StyleRees, Christiaan A., Marco Beccaria, Flavio A. Franchina, Jane E. Hill, and Giorgia Purcaro. 2019. "Fatty Acid Methyl Ester (FAME) Profiling Identifies Carbapenemase-Producing Klebsiella pneumoniae Belonging to Clonal Complex 258" Separations 6, no. 2: 32. https://doi.org/10.3390/separations6020032
APA StyleRees, C. A., Beccaria, M., Franchina, F. A., Hill, J. E., & Purcaro, G. (2019). Fatty Acid Methyl Ester (FAME) Profiling Identifies Carbapenemase-Producing Klebsiella pneumoniae Belonging to Clonal Complex 258. Separations, 6(2), 32. https://doi.org/10.3390/separations6020032