
Near Infrared Spectroscopic Evaluation of Starch Properties of Diverse Sorghum Populations

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Grain Samples
2.2. NIR Scanning
2.3. Starch and Amylose Content Determination
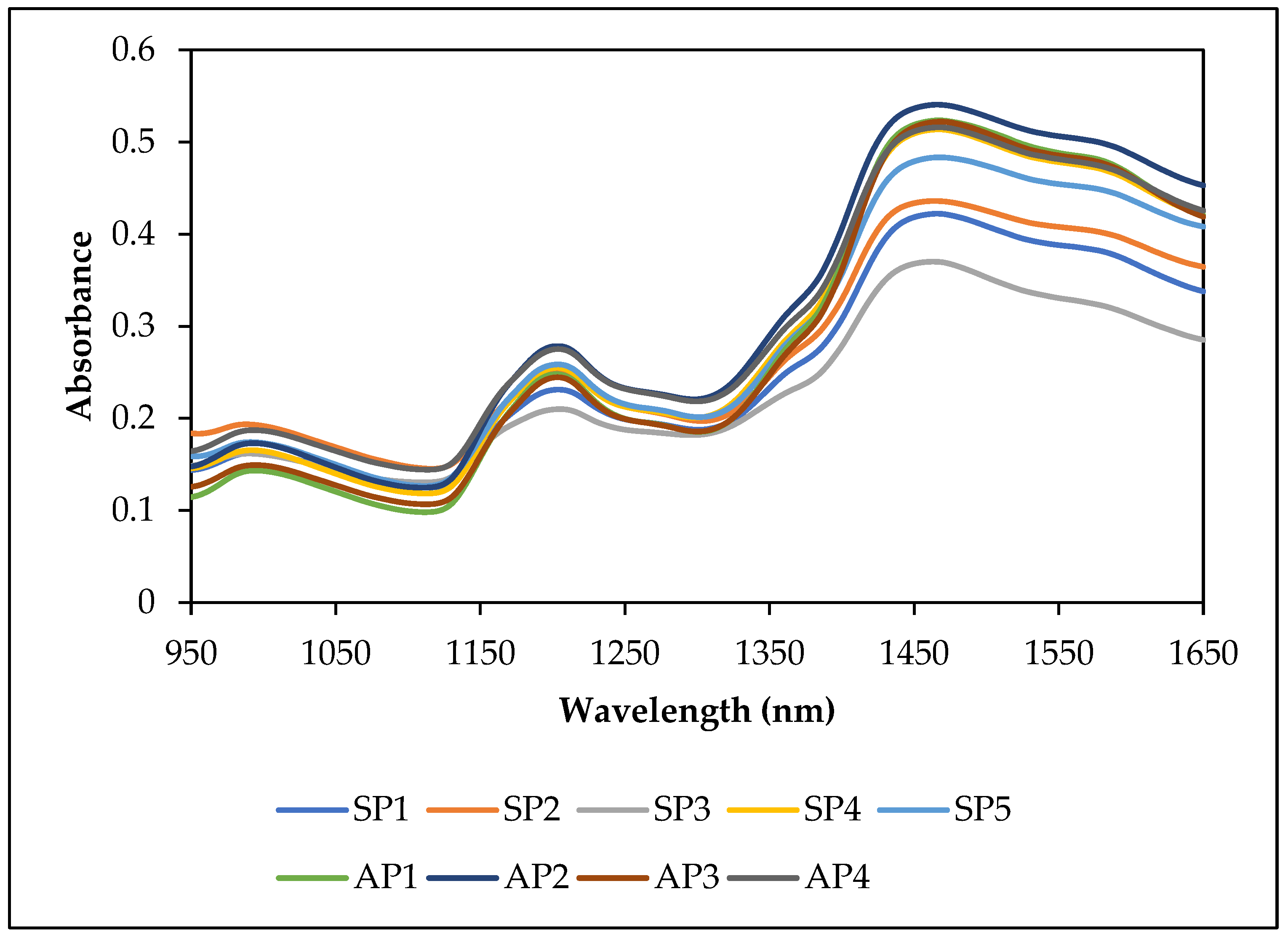
2.4. Spectral Data Acquisition and Data Analysis
2.5. Prediction of Moisture, Starch, Amylose and Protein Contents of New BREEDING Populations
3. Results and Discussion
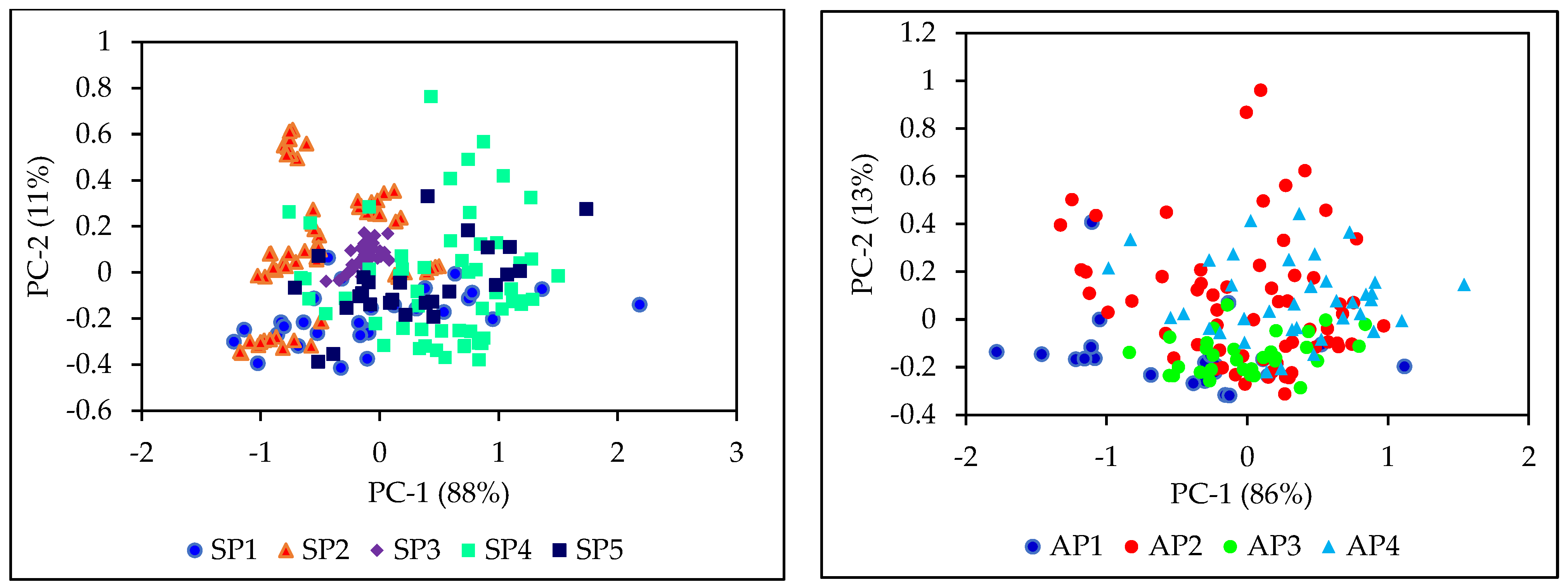
3.1. Diversity of Sample Populations
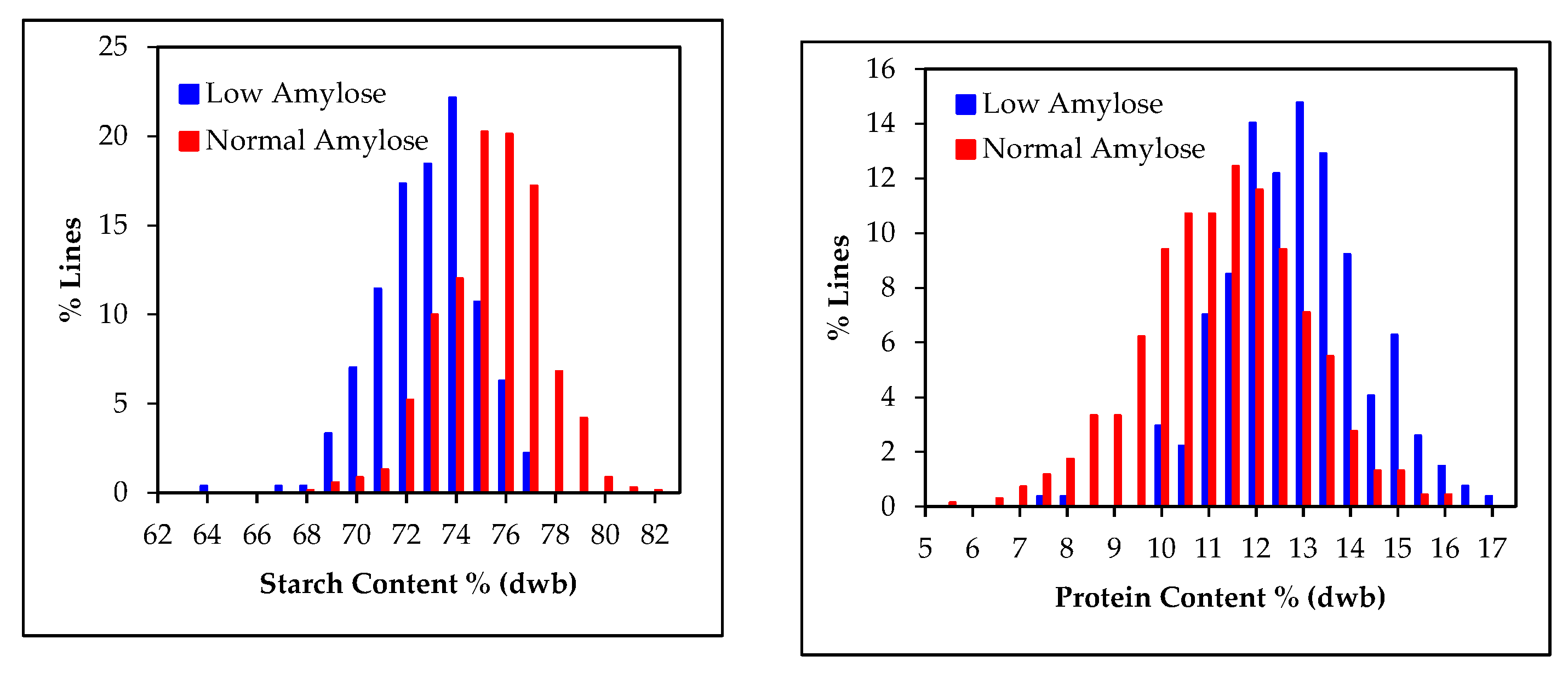
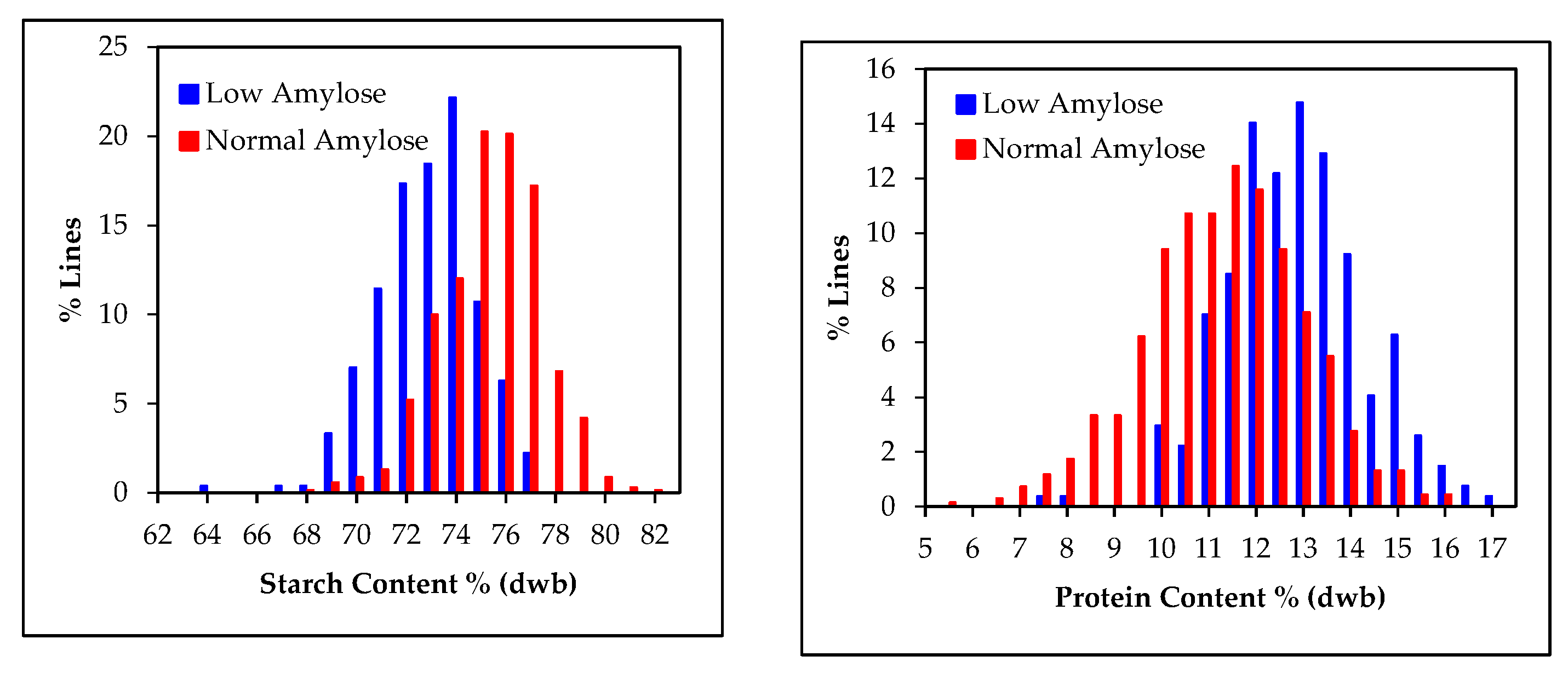
3.2. Starch and Amylose Contents in Grain Samples
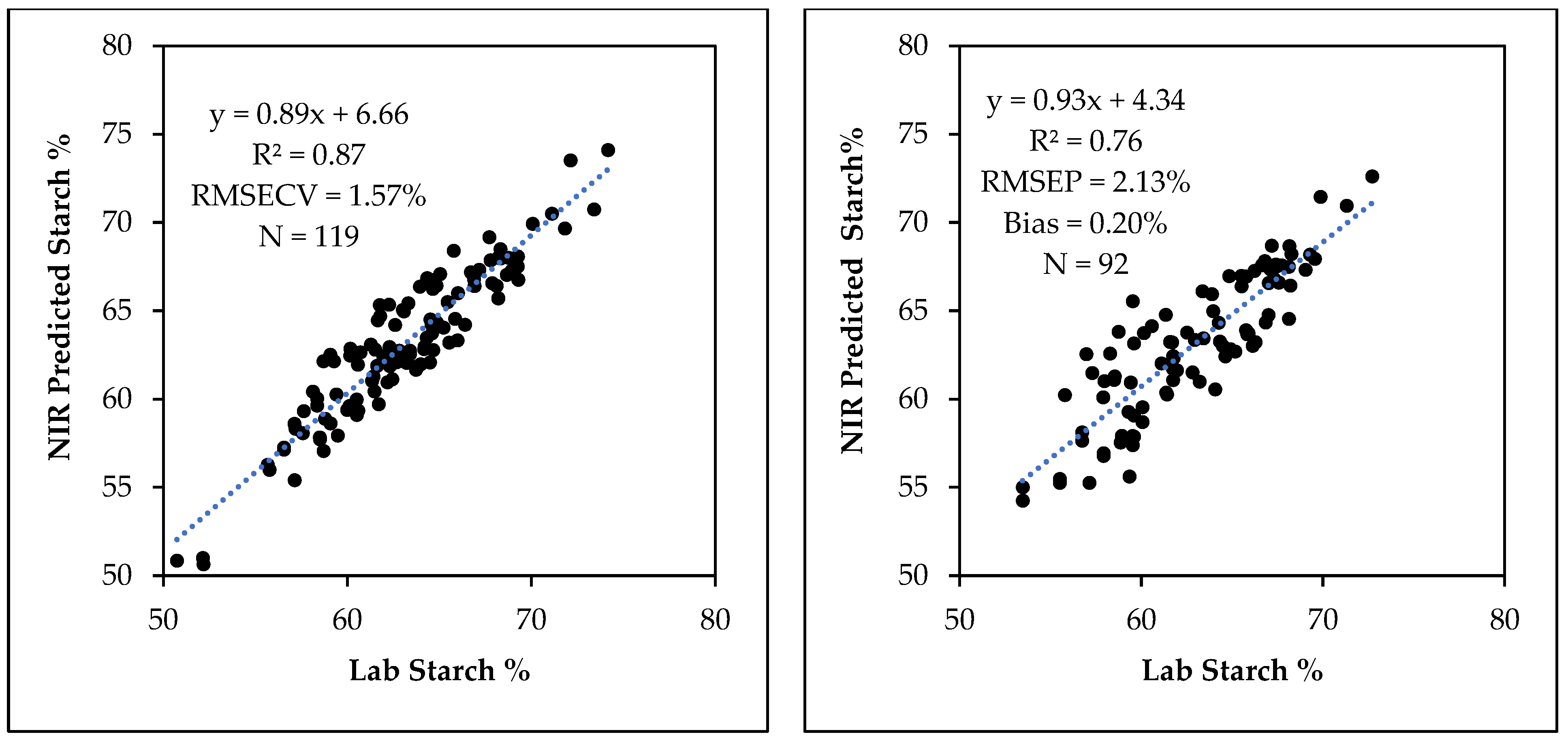
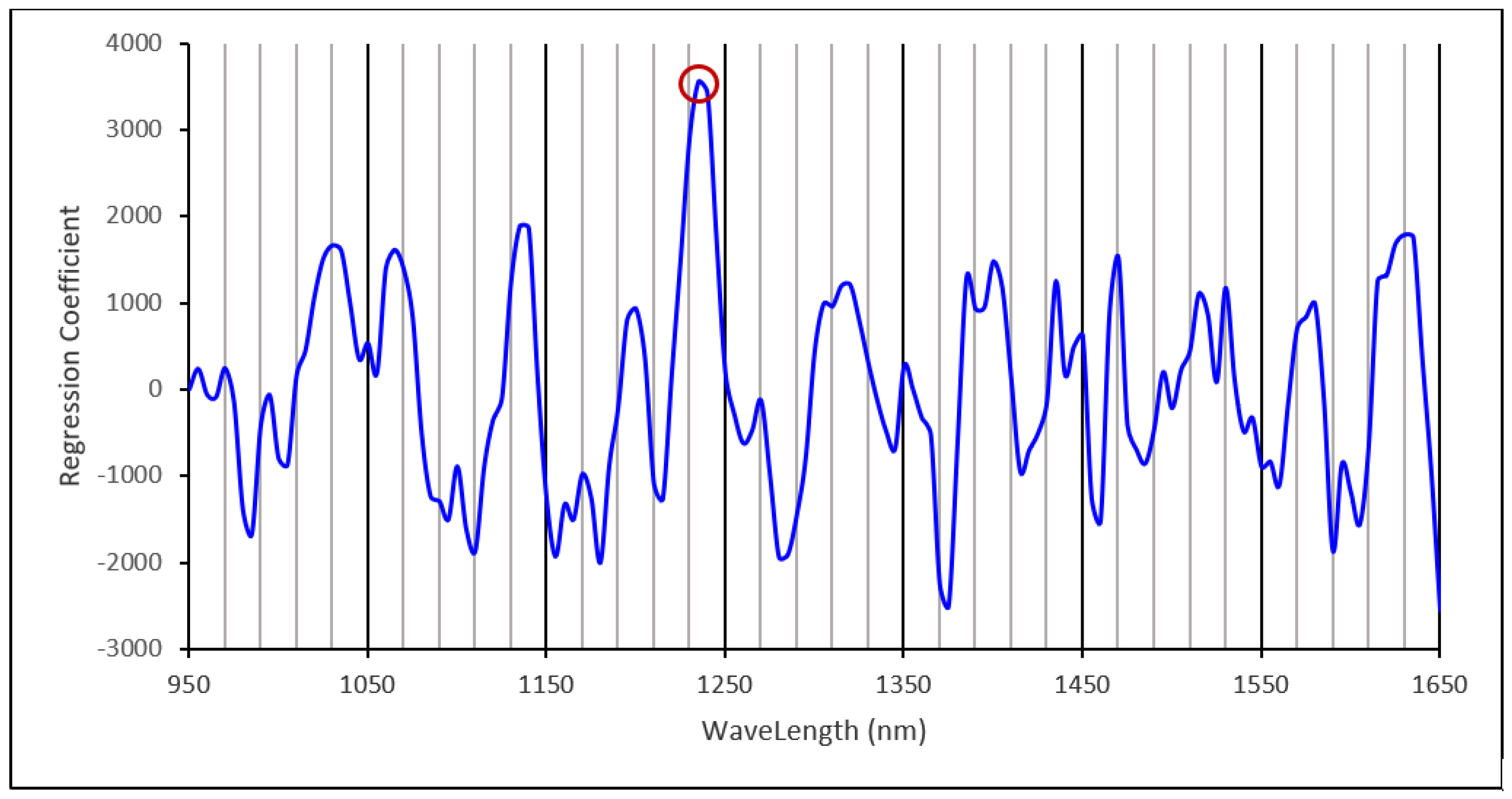
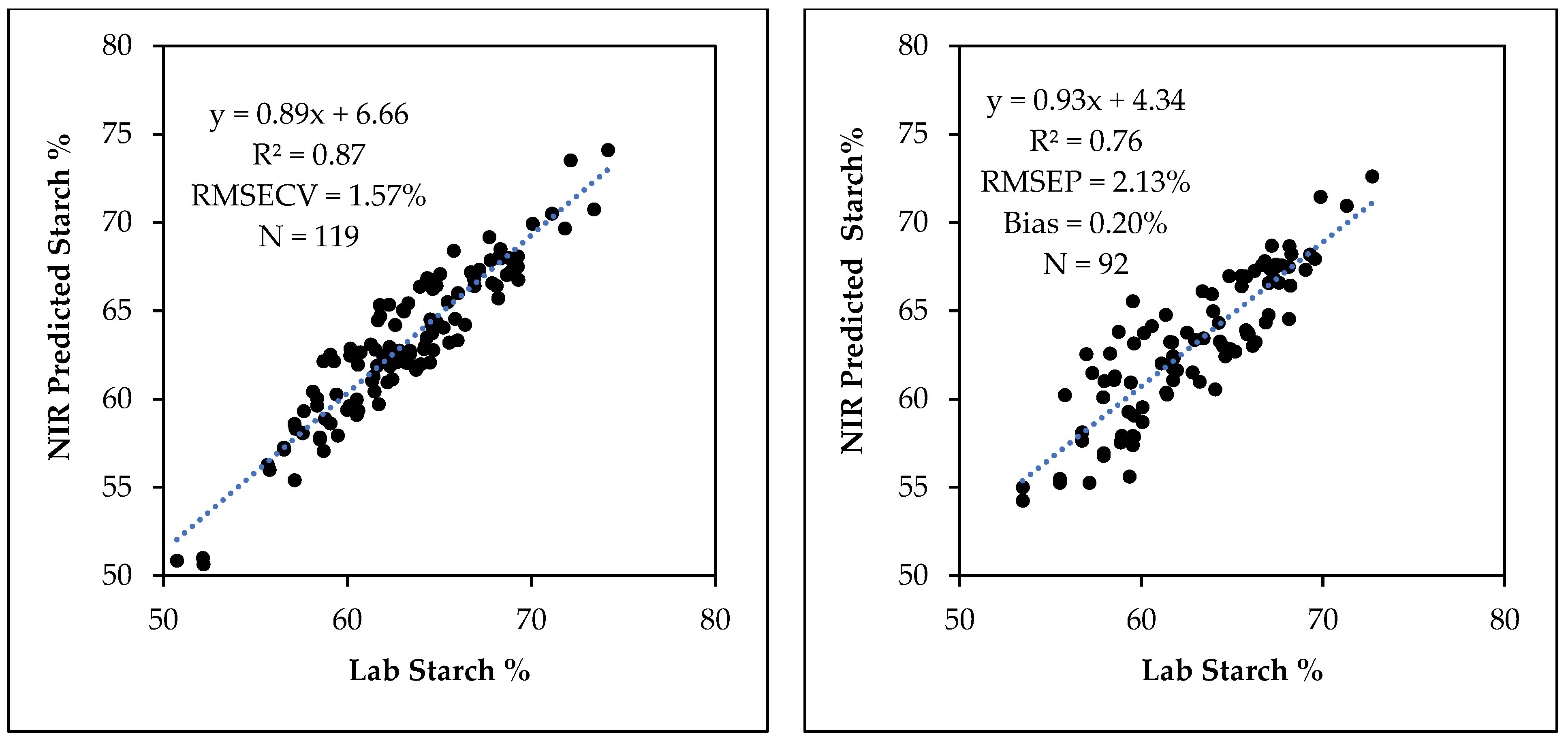
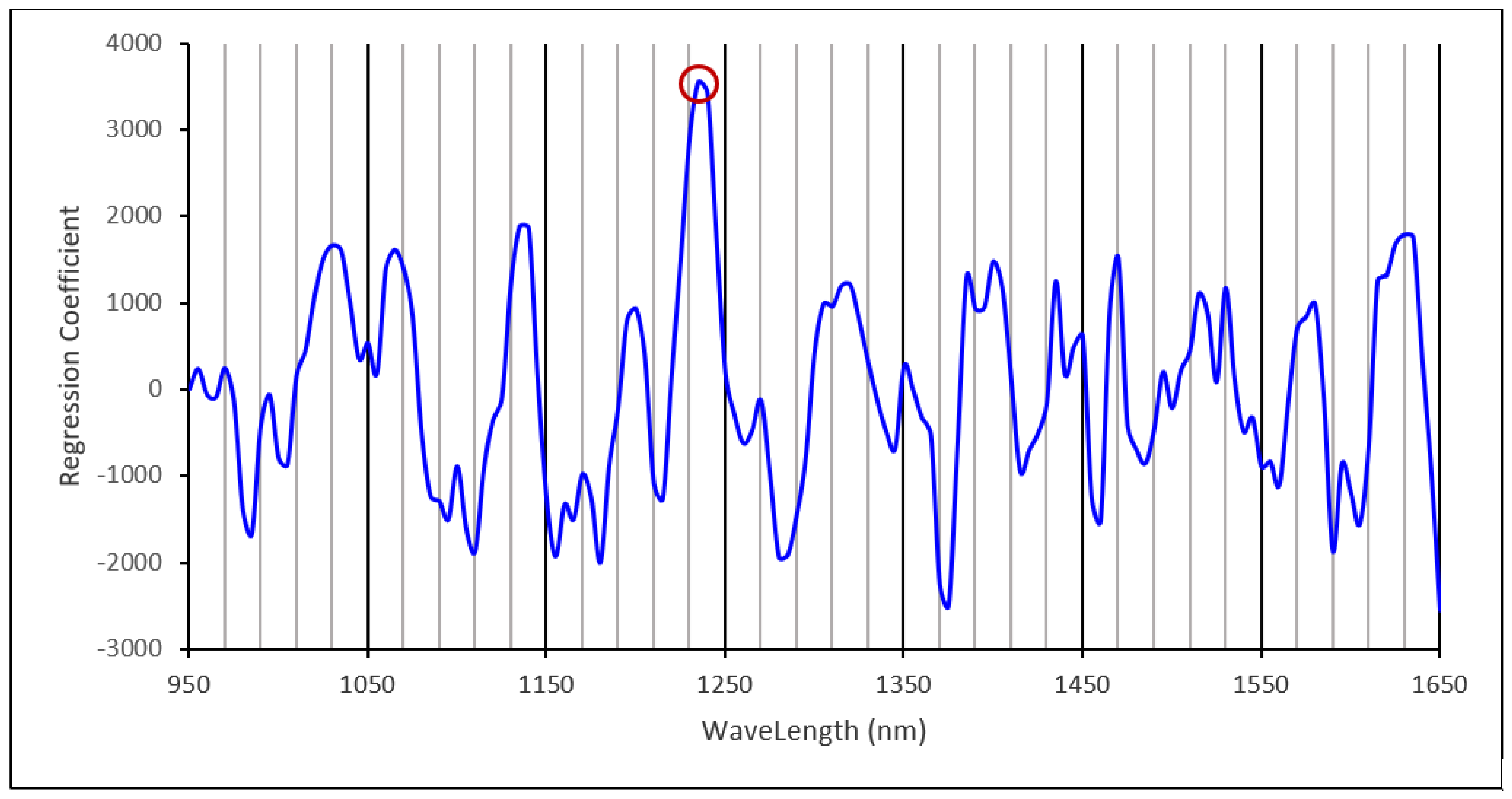
3.3. Starch Calibration Development and Model Validation
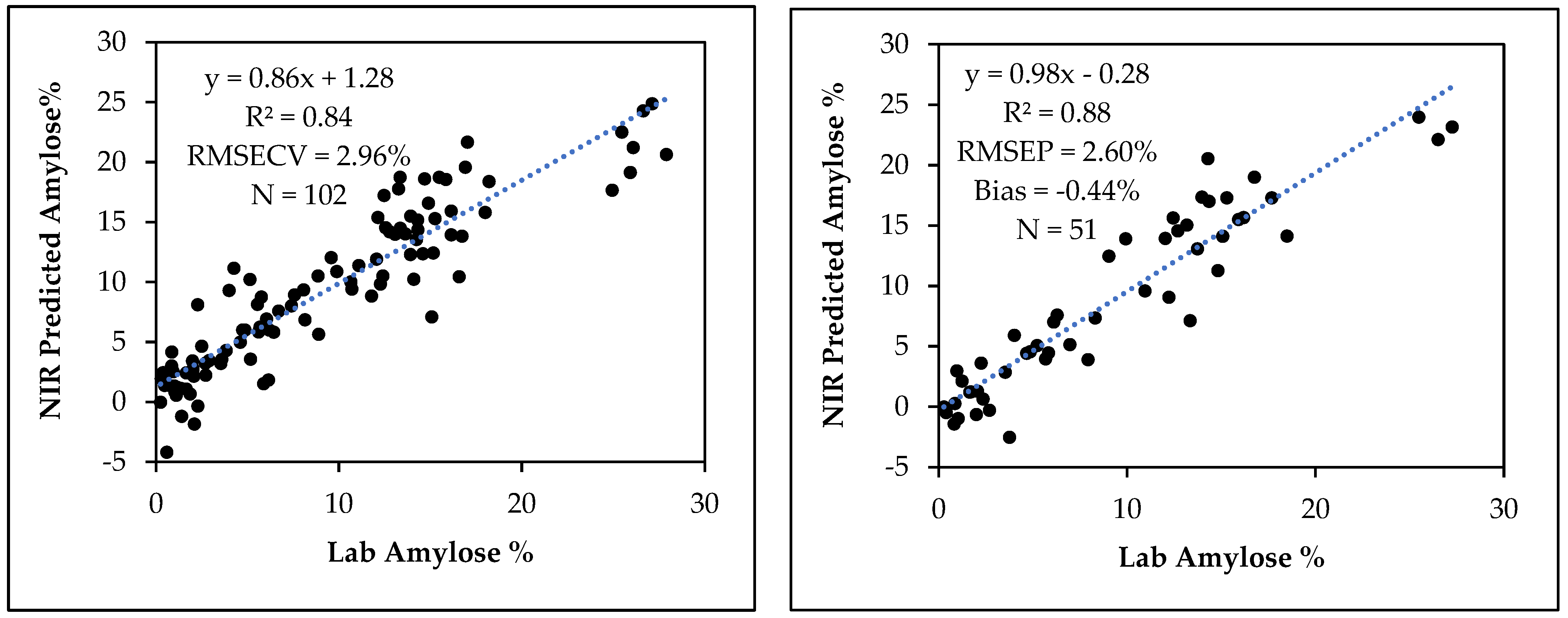
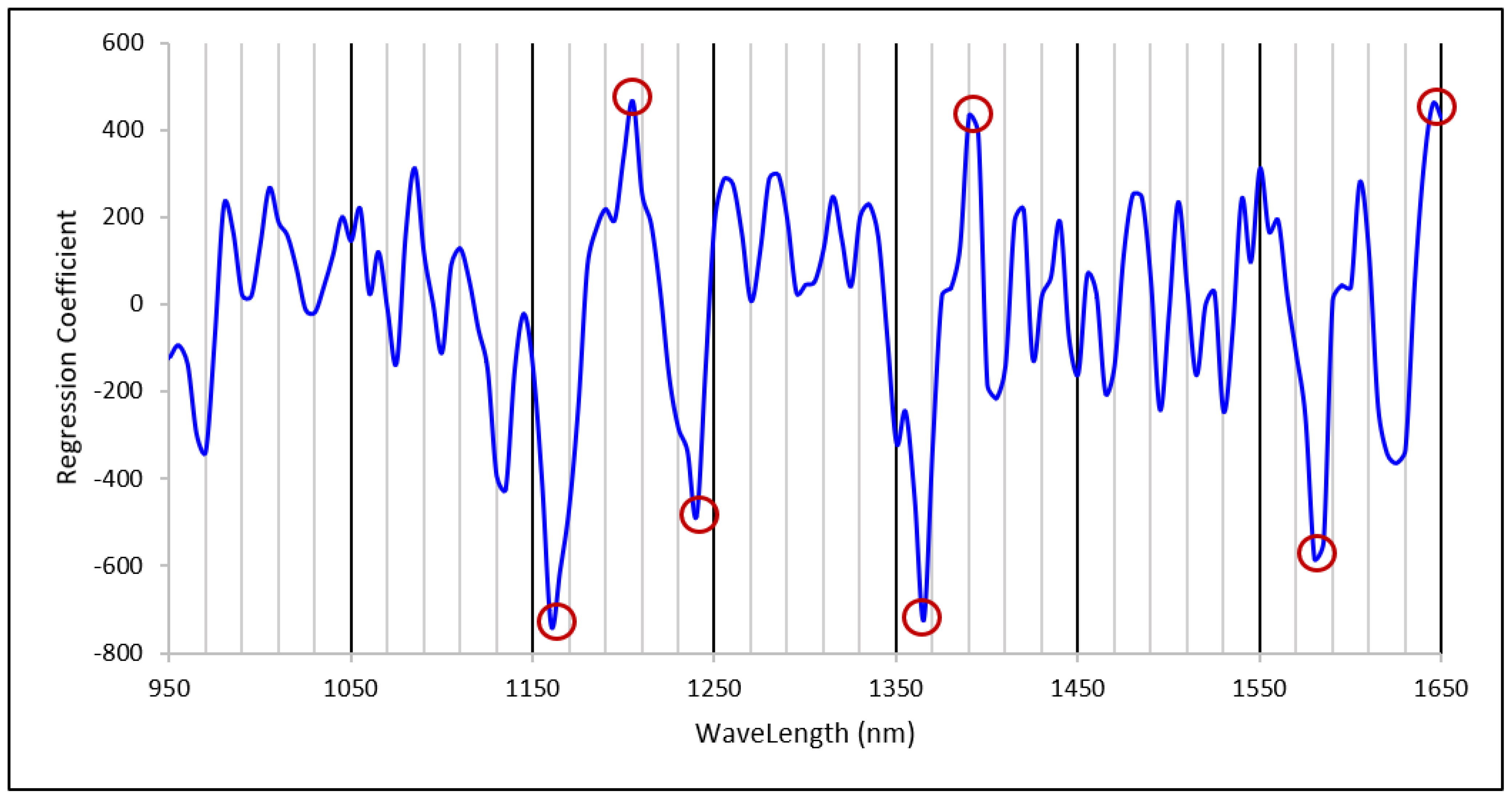
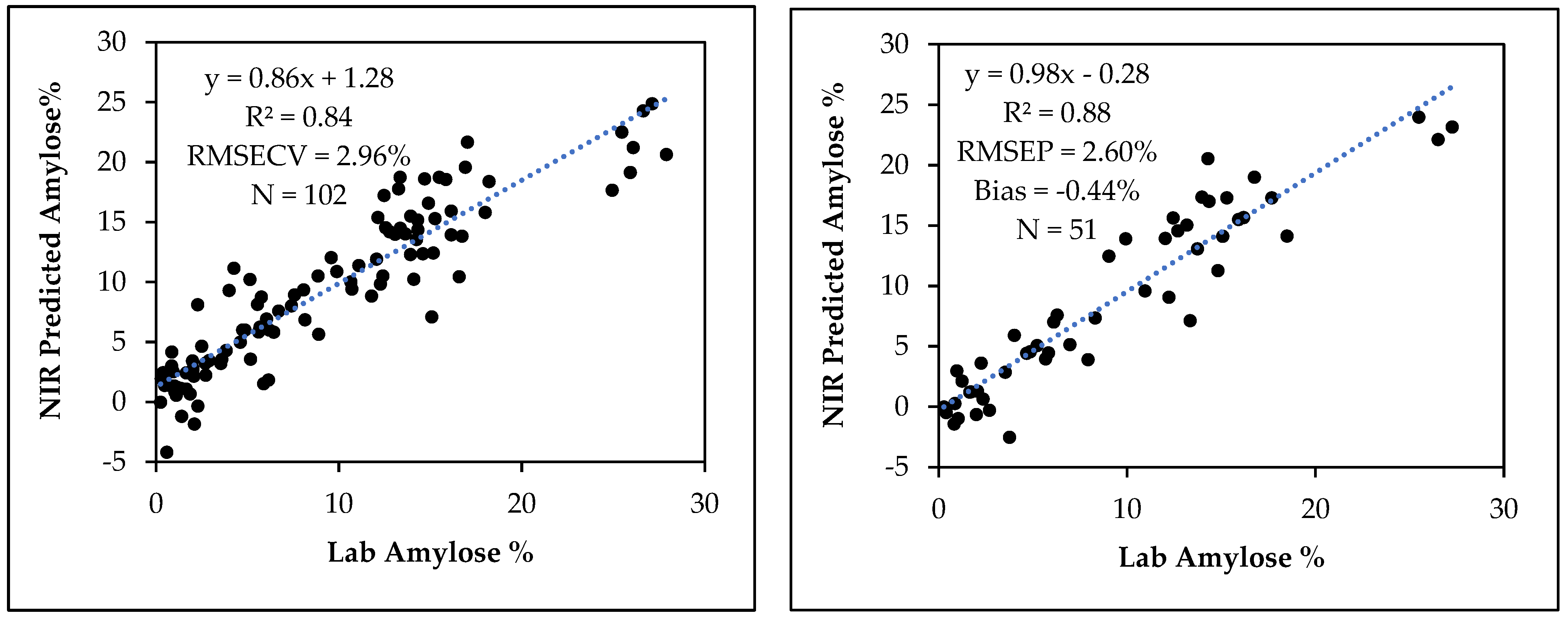
3.4. Amylose Calibration Development and Model Validation
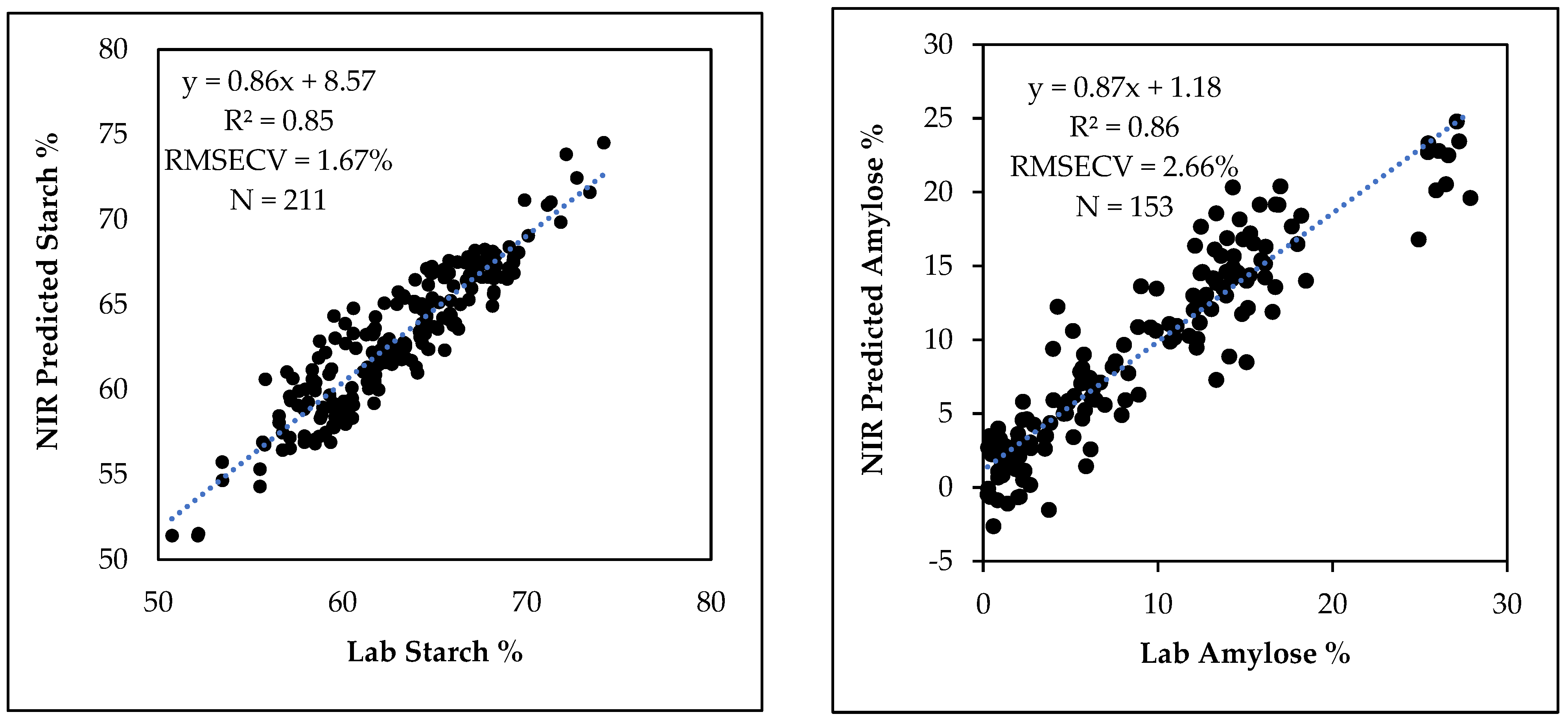
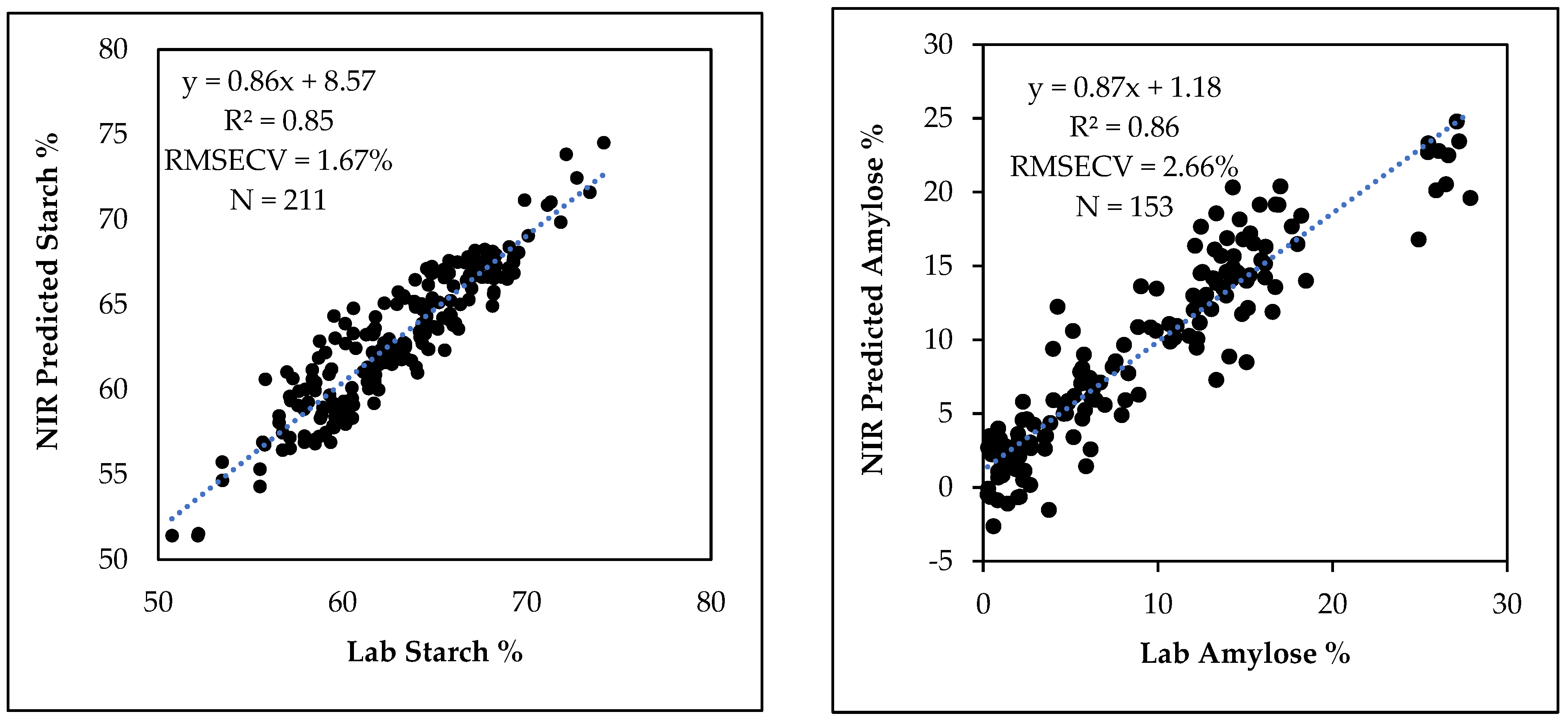
3.5. NIR Prediction of Starch and Amylose Contents in Breeding Populations
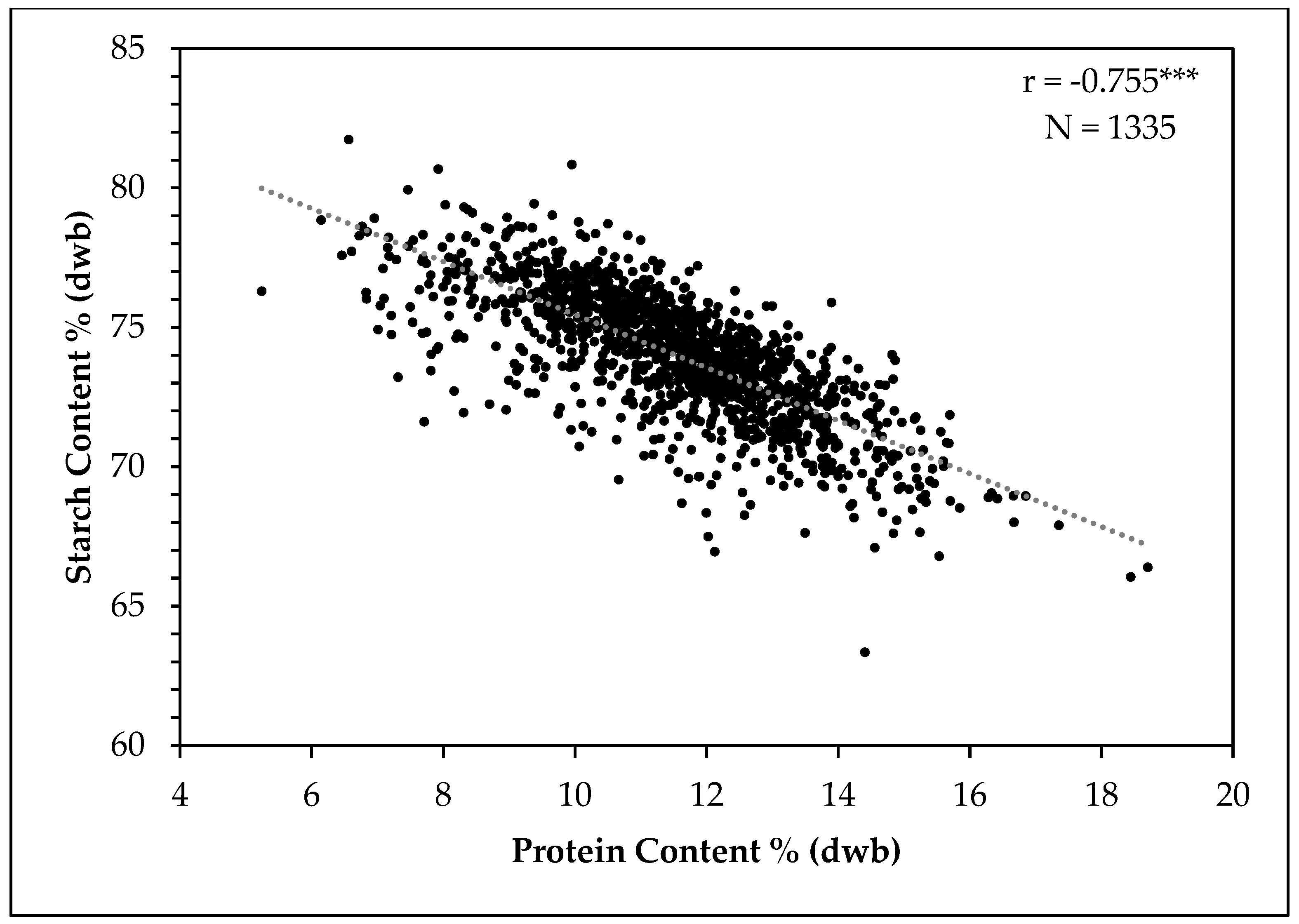
3.6. Relationship between, Starch, Amylose and Protein Contents in Grain Sorghum Populations
3.7. NIR Spectroscopy for High Throughput Phenotyping of Segregating Sorghum Populations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anglani, C. Sorghum for human food–A review. Plant Foods Hum. Nutr. 1998, 52, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Aruna, C.; Visarada, K.B. Other Industrial Uses of Sorghum. In Breeding Sorghum for Diverse End Uses; Aruna, C., Visarada, K.B.R.S., Venkatesh Bhat, B., Tonapi, V.A., Eds.; Woodhead Publishing: Cambridge, MA, USA, 2019; pp. 271–292. ISBN 9780081018804. [Google Scholar]
- Dahlberg, J.; Janoš, B.; Sikora, V.; Latković, D. Assessing sorghum [Sorghum bicolor (L.) Moench] germplasm for new traits: Food, fuels & unique uses. Maydica 2012, 56, 85–92. [Google Scholar]
- Taylor, J.R.; Schober, T.J.; Bean, S.R. Novel food and non-food uses for sorghum and millets. J. Cereal Sci. 2006, 44, 252–271. [Google Scholar] [CrossRef]
- Wang, D.; Bean, S.; McLaren, J.; Seib, P.; Madl, R.; Tuinstra, M.; Shi, Y.; Lenz, M.; Wu, X.; Zhao, R. Grain sorghum is a viable feedstock for ethanol production. J. Ind. Microbiol. Biotechnol. 2008, 35, 313–320. [Google Scholar] [CrossRef] [PubMed]
- de Morais Cardoso, L.; Pinheiro, S.S.; Martino, H.S.D.; Pinheiro-Sant’Ana, H.M. Sorghum (Sorghum bicolor L.): Nutrients, bioactive compounds, and potential impact on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 372–390. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Fang, Z. Sorghum grain: From genotype, nutrition, and phenolic profile to its health benefits and food applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2025–2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGinnis, M.J.; Painter, J.E. Sorghum: History, use, and health benefits. Nutr. Today 2020, 55, 38–44. [Google Scholar] [CrossRef]
- Stefoska-Needham, A.; Tapsell, L. Considerations for progressing a mainstream position for sorghum, a potentially sustainable cereal crop, for food product innovation pipelines. Trends Food Sci. Technol. 2020, 97, 249–253. [Google Scholar] [CrossRef]
- Khoddami, A.; Messina, V.; Vadabalija Venkata, K.; Farahnaky, A.; Blanchard, C.L.; Roberts, T.H. Sorghum in foods: Functionality and potential in innovative products. Crit. Rev. Food Sci. Nutr. 2021, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.R.; Duodu, K.G. Sorghum and millets: Grain-quality characteristics and management of quality requirements. In Cereal Grains, 2nd ed.; Wrigley, C., Batey, I., Miskelly, D., Eds.; Woodhead Publishing: Cambridge, MA, USA, 2017; pp. 317–351. ISBN 978-0-08-100719-8. [Google Scholar]
- Tester, R.F.; Karkalas, J.; Qi, X. Starch—composition, fine structure and architecture. J. Cereal Sci. 2004, 39, 151–165. [Google Scholar] [CrossRef]
- Ai, Y.; Medic, J.; Jiang, H.; Wang, D.; Jane, J.L. Starch characterization and ethanol production of sorghum. J. Agric. Food Chem. 2011, 59, 7385–7392. [Google Scholar] [CrossRef] [PubMed]
- Beta, T.; Corke, H. Noodle quality as related to sorghum starch properties. Cereal Chem. 2001, 78, 417–420. [Google Scholar] [CrossRef]
- Miller, O.H.; Burns, E.E. Starch characteristics of selected grain sorghums as related to human foods. J. Food Sci. 1970, 35, 666–668. [Google Scholar] [CrossRef]
- Lichtenwalner, R.E.; Ellis, E.B.; Rooney, L.W. Effect of incremental dosages of the waxy gene of sorghum on digestibility. J. Anim. Sci. 1978, 46, 1113–1119. [Google Scholar] [CrossRef] [Green Version]
- McCleary, B.V.; Charmier, L.M.; McKie, V.A. Measurement of starch: Critical evaluation of current methodology. Starch-Stärke 2019, 71, 1800146. [Google Scholar] [CrossRef] [Green Version]
- Osborne, B.G. Applications of near infrared spectroscopy in quality screening of early-generation material in cereal breeding programmes. J. Near Infrared Spec. 2006, 14, 93–101. [Google Scholar] [CrossRef]
- Cozzolino, D.; Degner, S.; Eglinton, J. A review on the role of vibrational spectroscopy as an analytical method to measure starch biochemical and biophysical properties in cereals and starchy foods. Foods 2014, 3, 605–621. [Google Scholar] [CrossRef] [Green Version]
- Boyles, R.E.; Pfeiffer, B.K.; Cooper, E.A.; Rauh, B.L.; Zielinski, K.J.; Myers, M.T.; Brenton, Z.; Rooney, W.L.; Kresovich, S. Genetic dissection of sorghum grain quality traits using diverse and segregating populations. Theor. Appl. Genet. 2017, 130, 697–716. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Danao, M.G.C.; Chen, S.F.; Li, S.; Singh, V.; Brown, P.J. Prediction of starch content and ethanol yields of sorghum grain using near infrared spectroscopy. J. Near Infrared Spec. 2015, 23, 85–92. [Google Scholar] [CrossRef]
- Griebel, S.; Adedayo, A.; Tuinstra, M.R. Genetic diversity for starch quality and alkali spreading value in sorghum. Plant Genome 2021, 14, e20067. [Google Scholar] [CrossRef] [PubMed]
- De Alencar Figueiredo, L.F.; Davrieux, F.; Fliedel, G.; Rami, J.F.; Chantereau, J.; Deu, M.; Courtois, B.; Mestres, C. Development of NIRS equations for food grain quality traits through exploitation of a core collection of cultivated sorghum. J. Agric. Food Chem. 2006, 54, 8501–8509. [Google Scholar] [CrossRef] [PubMed]
- Casa, A.M.; Pressoir, G.; Brown, P.J.; Casa, A.M.; Pressoir, G.; Brown, P.J. Community resources and strategies for association mapping in sorghum. Crop Sci. 2008, 48, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Megazyme. Total Starch Assay Procedure (Amyloglucosidase/α-Amylase Method). K-TSTA-100A, Procedure (b). 2020. Available online: https://www.megazyme.com/documents/Assay_Protocol/K-TSTA-100A_DATA.pdf (accessed on 9 January 2021).
- Morrison, W.R.; Laignelet, B. An improved colorimetric procedure for determining apparent and total amylose in cereal and other starches. J. Cereal Sci. 1983, 1, 9–20. [Google Scholar] [CrossRef]
- Chrastil, J. Improved colorimetric determination of amylose in starches or flours. Carbohydr. Res. 1987, 159, 154–158. [Google Scholar] [CrossRef]
- McDonald, R.S.; Wilks, P.A., Jr. JCAMP-DX: A standard form for exchange of infrared spectra in computer readable form. Appl. Spectrosc. 1988, 42, 151–162. [Google Scholar] [CrossRef]
- Martens, H.; Stark, E. Extended multiplicative signal correction and spectral interference subtraction: New preprocessing methods for near infrared spectroscopy. J. Pharmaceut. Biomed. Anal. 1991, 9, 625–635. [Google Scholar] [CrossRef]
- Peiris, K.H.; Bean, S.R.; Chiluwal, A.; Perumal, R.; Jagadish, S.K. Moisture effects on robustness of sorghum grain protein near-infrared spectroscopy calibration. Cereal Chem. 2019, 96, 678–688. [Google Scholar] [CrossRef]
- Dowell, F.E.; Maghirang, E.B.; Graybosch, R.A.; Berzonsky, W.A.; Delwiche, S.R. Selecting and sorting waxy wheat kernels using near-infrared spectroscopy. Cereal Chem. 2009, 86, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Yong, H.; Shuijuan, F. Short-wave near-infrared spectroscopy analysis of major compounds in milk powder and wavelength assignment. Anal. Chim. Acta 2008, 610, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Osborne, B.G.; Fearn, T.; Hindle, P.H. Practical NIR Spectroscopy with Applications in Food and Beverage Analysis; Longman Scientific and Technical: Singapore, 1993; 227p. [Google Scholar]
- Williams, P.C. Implementation of near-infrared technology. In Near-Infrared Technology in the Agricultural and Food Industries; Williams, P., Norris, K.H., Eds.; American Association of Cereal Chemist: St. Paul, MN, USA, 2001; pp. 145–169. [Google Scholar]
- Fertig, C.C.; Podczeck, F.; Jee, R.D.; Smith, M.R. Feasibility study for the rapid determination of the amylose content in starch by near-infrared spectroscopy. Eur. J. Pharm. Sci. 2004, 21, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Dardenne, P. Some considerations about NIR spectroscopy: Closing speech at NIR-2009. NIR News 2010, 21, 8–14. [Google Scholar] [CrossRef]
- De Maesschalck, R.; Jouan-Rimbaud, D.; Massart, D.L. The mahalanobis distance. Chemometr. Intell. Lab. Syst. 2000, 50, 1–18. [Google Scholar] [CrossRef]
- Subramanian, V.; Jambunathan, R. Properties of sorghum grain and their relationship to roti quality. Sorghum in the eighties. In Proceedings of the International Symposium on Sorghum Grain Quality, ICRISAT Center, Patanchuru, India, 28–31 October 1981; pp. 281–288. [Google Scholar]
- Buffo, R.A.; Weller, C.L.; Parkhurst, A.M. Relationships among grain sorghum quality factors. Cereal Chem. 1998, 75, 100–104. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, D.H.; Hoffmann, L.; Rooney, W.L.; Herald, T.J.; Bean, S.; Boyles, R.; Brenton, Z.W.; Kresovich, S. Genetic architecture of kernel composition in global sorghum germplasm. BMC Genom. 2017, 18, 15. [Google Scholar] [CrossRef] [Green Version]
- Jampala, B.; Rooney, W.L.; Peterson, G.C.; Bean, S.; Hays, D.B. Estimating the relative effects of the endosperm traits of waxy and high protein digestibility on yield in grain sorghum. Field Crops Res. 2012, 139, 57–62. [Google Scholar] [CrossRef]
- Yerka, M.K.; Toy, J.J.; Funnell-Harris, D.L.; Sattler, S.E.; Pedersen, J.F. Registration of A/BN641 and RN642 waxy grain sorghum genetic stocks. J. Plant Regist. 2015, 9, 258–261. [Google Scholar] [CrossRef]
- Yerka, M.K.; Toy, J.J.; Funnell-Harris, D.L.; Sattler, S.E.; Pedersen, J.F. Registration of N619 to N640 grain sorghum lines with waxy or wild-type endosperm. J. Plant Regist. 2015, 9, 244–248. [Google Scholar] [CrossRef] [Green Version]
- Yerka, M.K.; Toy, J.J.; Funnell-Harris, D.L.; Sattler, S.E.; Pedersen, J.F. Evaluation of interallelic waxy, heterowaxy, and wild-type grain sorghum hybrids. Crop Sci. 2016, 56, 1–9. [Google Scholar] [CrossRef]
- Lane, H.M.; Murray, S.C. High Throughput can produce better decisions than high accuracy when phenotyping plant populations. Crop Sci. 2021, 61, 3301–3313. [Google Scholar] [CrossRef]
- Beta, T.; Corke, H. Genetic and environmental variation in sorghum starch properties. J. Cereal Sci. 2001, 34, 261–268. [Google Scholar] [CrossRef]
- Tester, R.F.; Karkalas, J. The effects of environmental conditions on the structural features and physico-chemical properties of starches. Starch-Stärke 2001, 53, 513–519. [Google Scholar] [CrossRef]
- Gonçalves, M.T.V.; Morota, G.; Costa, P.M.D.A.; Vidigal, P.M.P.; Barbosa, M.H.P.; Peternelli, L.A. Near-infrared spectroscopy outperforms genomics for predicting sugarcane feedstock quality traits. PLoS ONE 2021, 16, e0236853. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Population | N | Year | Location(s) | Type of Sample Population |
|---|---|---|---|---|
| Amylose | ||||
| AP1 | 22 | 2018 | Texas | Breeding Population |
| AP2 | 63 | 2019 | Kansas/Texas | Breeding Population |
| AP3 | 31 | 2020 | Texas | Breeding Population |
| AP4 | 37 | 2020 | Texas | Breeding Population |
| Starch | ||||
| SP1 | 29 | 2018 | Kansas | Diversity panel |
| SP2 | 61 | 2021 | Kansas | Hybrid |
| SP3 | 39 | 2019 | Kansas | Diversity panel |
| SP4 | 56 | 2019 | Kansas/Texas | Breeding Population |
| SP5 | 26 | 2020 | California | Breeding Population |
| Predictions | ||||
| Breeding 1 | 946 | 2020 | Texas | Breeding Populations |
| Breeding 2 | 391 | 2020 | California/Argentina/Mexico | Breeding Populations |
| Sample Set | Population | N * | Min | Max | Avg | SD |
|---|---|---|---|---|---|---|
| Starch samples | ||||||
| All | 211 | 50.73 | 74.17 | 62.99 | 4.31 | |
| SP1 | 29 | 55.79 | 65.54 | 61.44 | 2.56 | |
| SP2 | 61 | 50.73 | 64.56 | 58.82 | 2.96 | |
| SP3 | 39 | 63.93 | 69.55 | 67.23 | 1.40 | |
| SP4 | 56 | 58.69 | 68.10 | 63.14 | 2.28 | |
| SP5 | 26 | 59.26 | 74.17 | 67.85 | 3.95 | |
| Calibration set | 119 | 50.73 | 74.17 | 63.12 | 4.34 | |
| SP1 | 16 | 58.12 | 65.54 | 62.46 | 2.16 | |
| SP2 | 33 | 50.73 | 64.56 | 58.82 | 3.27 | |
| SP3 | 19 | 63.93 | 69.29 | 67.31 | 1.60 | |
| SP4 | 34 | 58.69 | 68.10 | 62.96 | 2.09 | |
| SP5 | 17 | 59.26 | 74.17 | 67.75 | 4.38 | |
| Validation set | 92 | 53.46 | 72.70 | 62.83 | 4.29 | |
| SP1 | 13 | 55.79 | 63.42 | 60.18 | 2.52 | |
| SP2 | 28 | 53.46 | 63.95 | 58.82 | 2.59 | |
| SP3 | 20 | 64.84 | 69.55 | 67.17 | 1.21 | |
| SP4 | 22 | 58.74 | 67.00 | 63.42 | 2.58 | |
| SP5 | 9 | 62.81 | 72.70 | 68.03 | 3.21 | |
| Amylose samples | ||||||
| All | 153 | 0.25 | 27.90 | 9.17 | 7.14 | |
| AP1 | 22 | 4.00 | 27.90 | 14.68 | 10.91 | |
| AP2 | 63 | 0.29 | 18.21 | 8.27 | 5.71 | |
| AP3 | 31 | 0.25 | 12.27 | 3.70 | 3.56 | |
| AP4 | 37 | 1.40 | 18.48 | 12.01 | 4.98 | |
| Calibration set | 102 | 0.25 | 27.90 | 9.20 | 7.19 | |
| AP1 | 16 | 4.00 | 27.90 | 14.36 | 10.89 | |
| AP2 | 41 | 0.29 | 18.21 | 8.74 | 5.92 | |
| AP3 | 24 | 0.25 | 12.27 | 4.07 | 3.77 | |
| AP4 | 21 | 1.40 | 18.00 | 12.03 | 4.89 | |
| Validation set | 51 | 0.28 | 27.25 | 9.12 | 7.09 | |
| AP1 | 6 | 4.02 | 27.25 | 15.52 | 11.95 | |
| AP2 | 22 | 0.86 | 16.75 | 7.41 | 5.33 | |
| AP3 | 7 | 0.28 | 6.29 | 2.44 | 2.52 | |
| AP4 | 16 | 1.99 | 18.48 | 11.98 | 5.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peiris, K.H.S.; Wu, X.; Bean, S.R.; Perez-Fajardo, M.; Hayes, C.; Yerka, M.K.; Jagadish, S.V.K.; Ostmeyer, T.; Aramouni, F.M.; Tesso, T.; et al. Near Infrared Spectroscopic Evaluation of Starch Properties of Diverse Sorghum Populations. Processes 2021, 9, 1942. https://doi.org/10.3390/pr9111942
Peiris KHS, Wu X, Bean SR, Perez-Fajardo M, Hayes C, Yerka MK, Jagadish SVK, Ostmeyer T, Aramouni FM, Tesso T, et al. Near Infrared Spectroscopic Evaluation of Starch Properties of Diverse Sorghum Populations. Processes. 2021; 9(11):1942. https://doi.org/10.3390/pr9111942
Chicago/Turabian StylePeiris, Kamaranga H. S., Xiaorong Wu, Scott R. Bean, Mayra Perez-Fajardo, Chad Hayes, Melinda K. Yerka, S. V. Krishna Jagadish, Troy Ostmeyer, Fadi M. Aramouni, Tesfaye Tesso, and et al. 2021. "Near Infrared Spectroscopic Evaluation of Starch Properties of Diverse Sorghum Populations" Processes 9, no. 11: 1942. https://doi.org/10.3390/pr9111942
APA StylePeiris, K. H. S., Wu, X., Bean, S. R., Perez-Fajardo, M., Hayes, C., Yerka, M. K., Jagadish, S. V. K., Ostmeyer, T., Aramouni, F. M., Tesso, T., Perumal, R., Rooney, W. L., Kent, M. A., & Bean, B. (2021). Near Infrared Spectroscopic Evaluation of Starch Properties of Diverse Sorghum Populations. Processes, 9(11), 1942. https://doi.org/10.3390/pr9111942