Kinetic Modelling of the Coproduction Process of Fumaric and Malic Acids by Rhizopus arrhizus NRRL 1526

 and
and 
Abstract
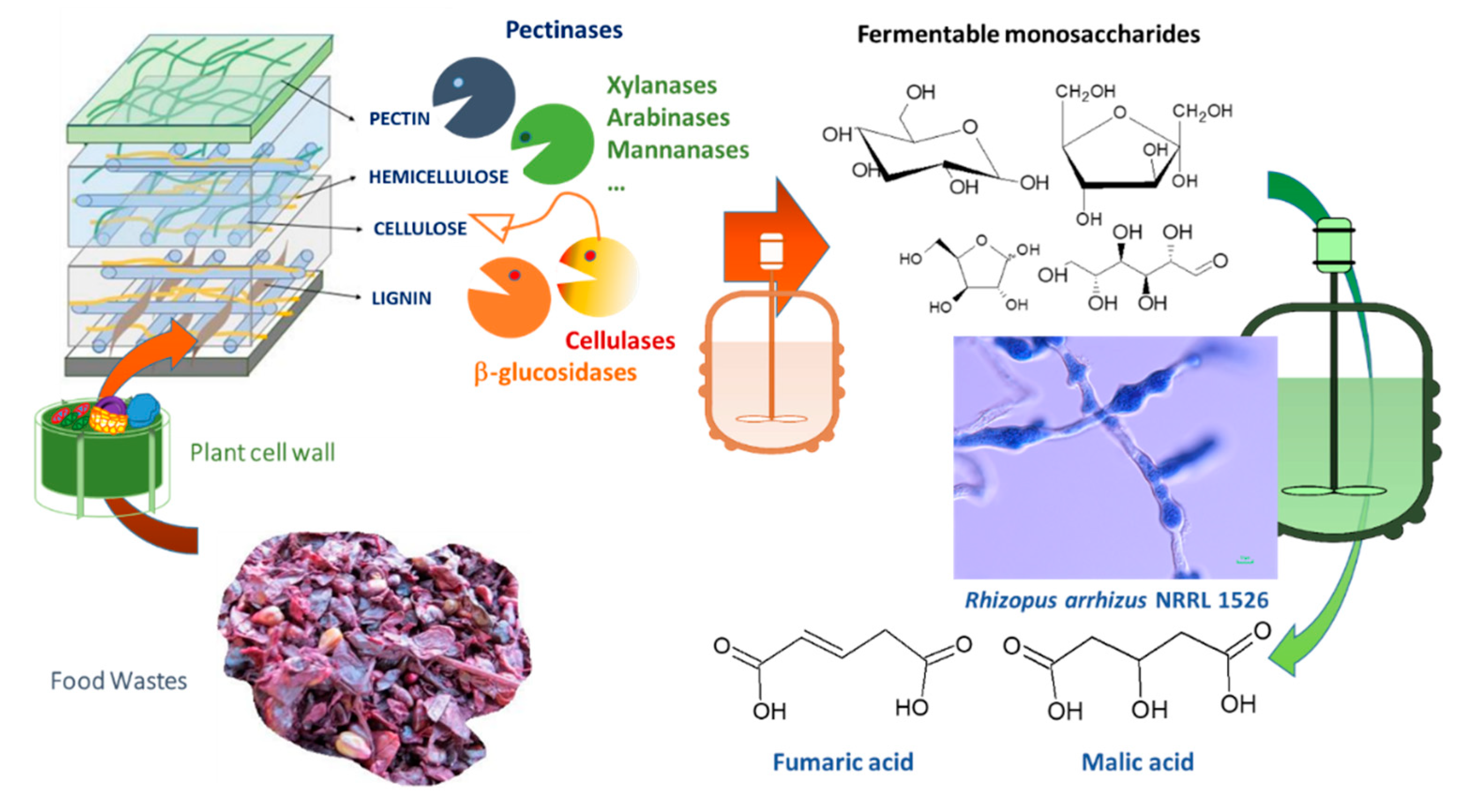
1. Introduction
2. Materials and Methods
2.1. Microorganism and Spore Production
2.2. Culture Mediums
2.3. Culture Conditions
2.4. Analitical Methods
2.5. Mathematic Methods
3. Results and Discussion
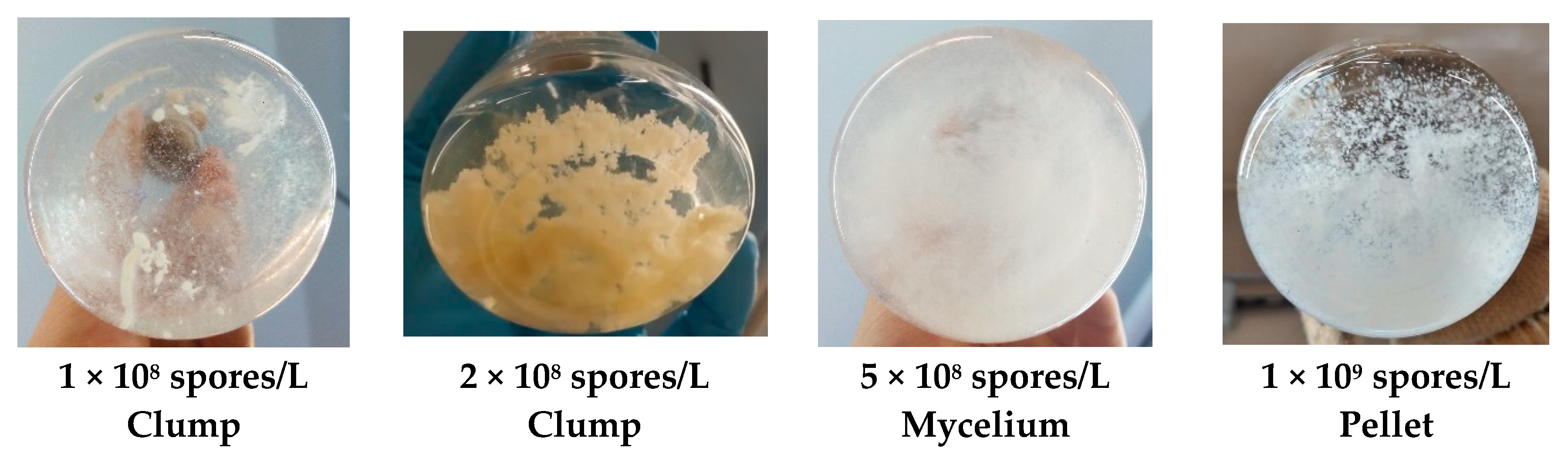
3.1. Fungi Morphology Selection at the Inoculum Stage
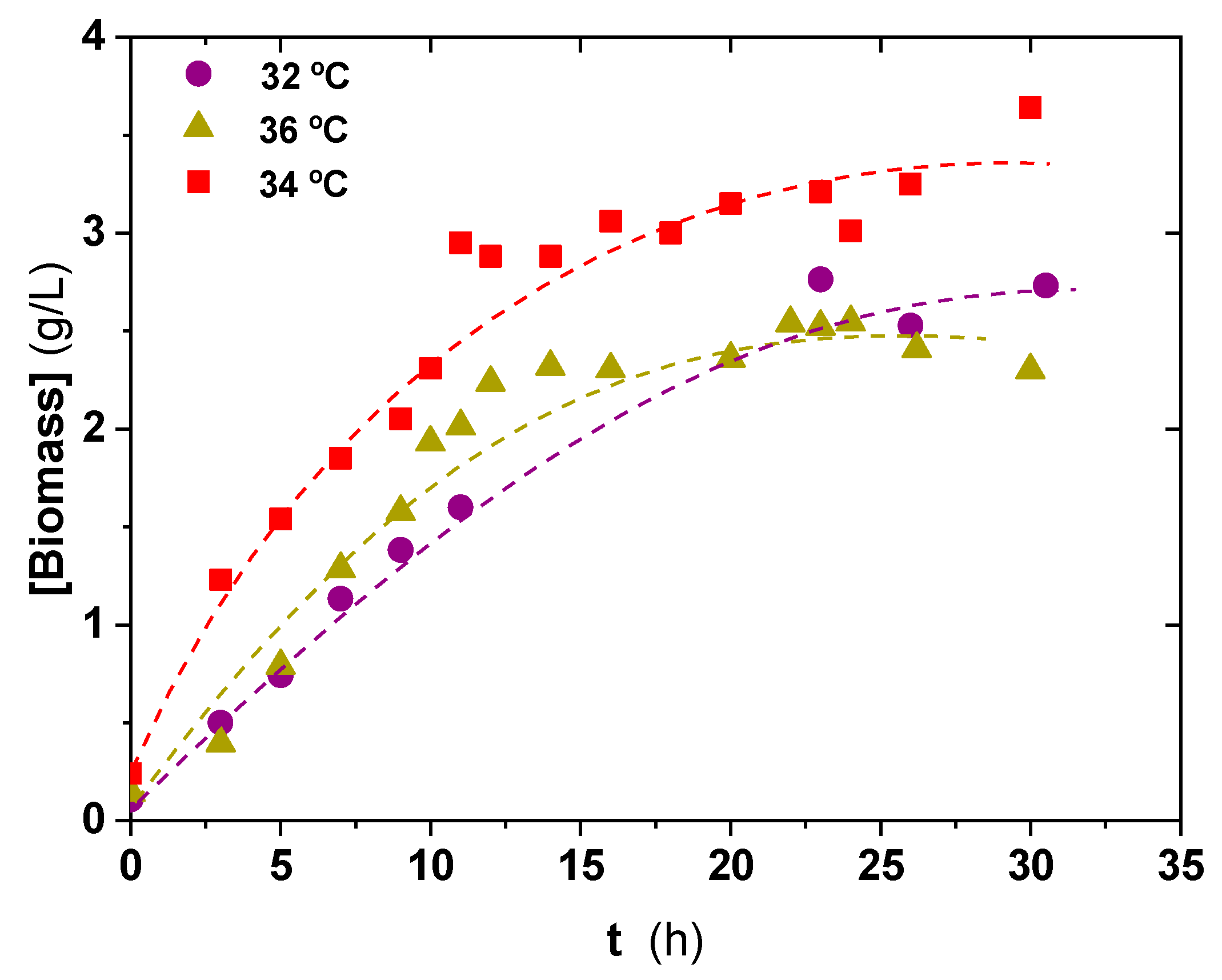
3.2. Temperature and pH Effect on Inoculum Biomass Growth
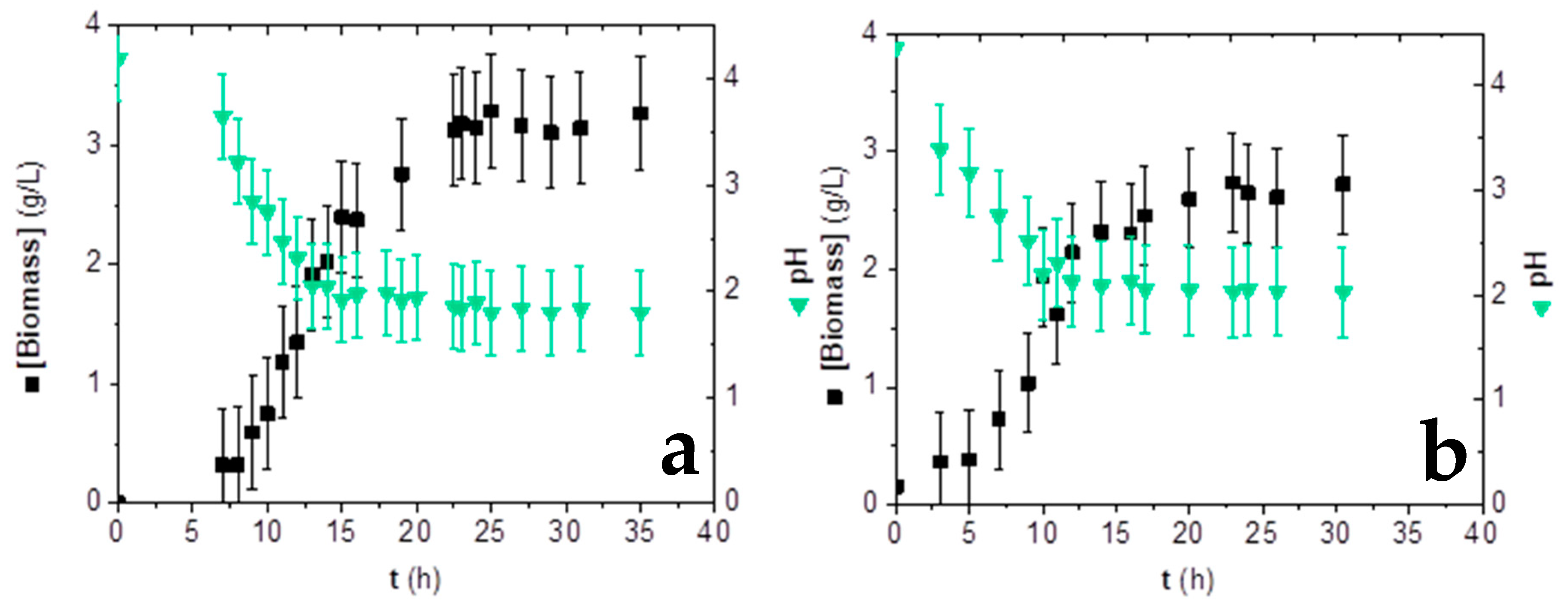
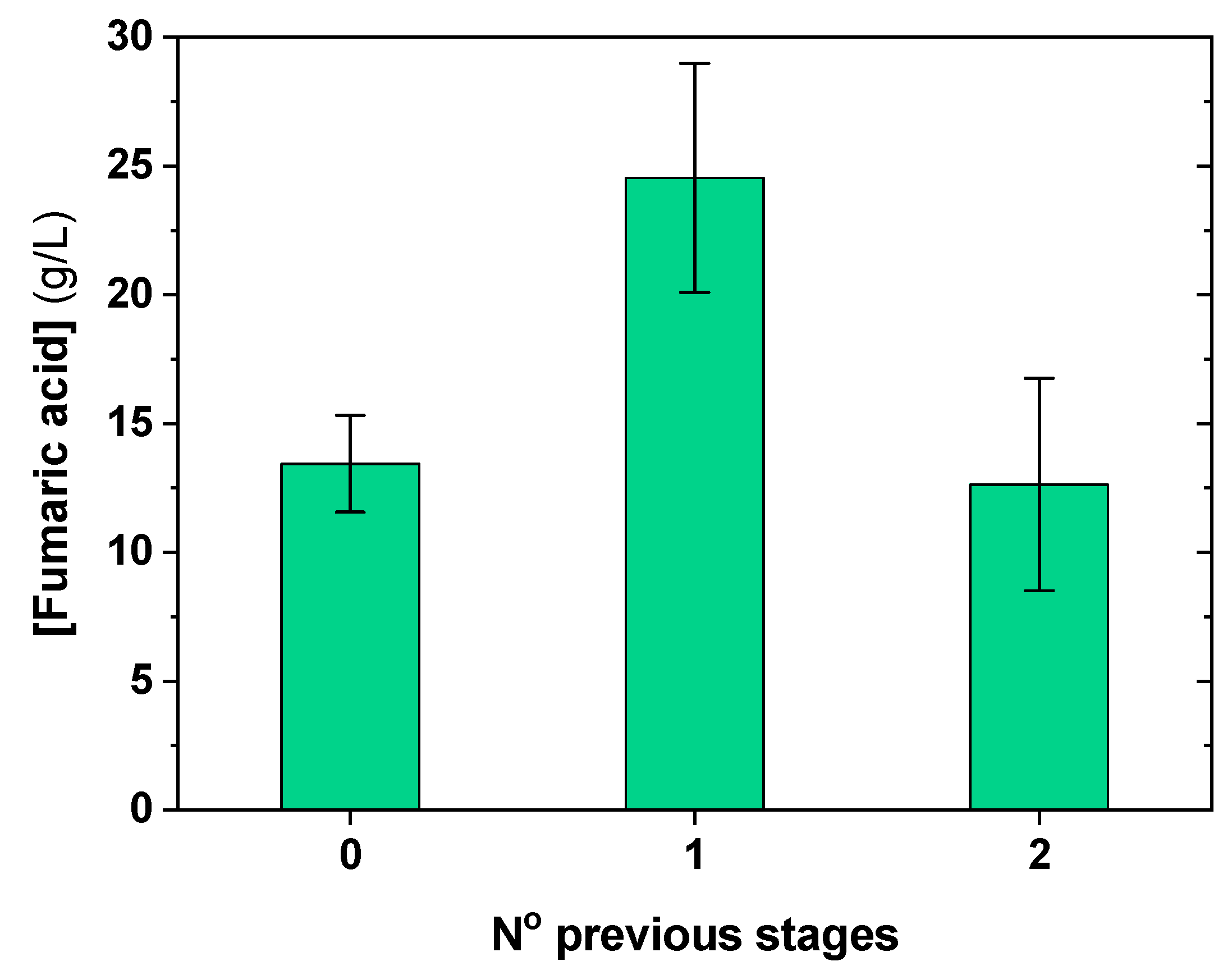
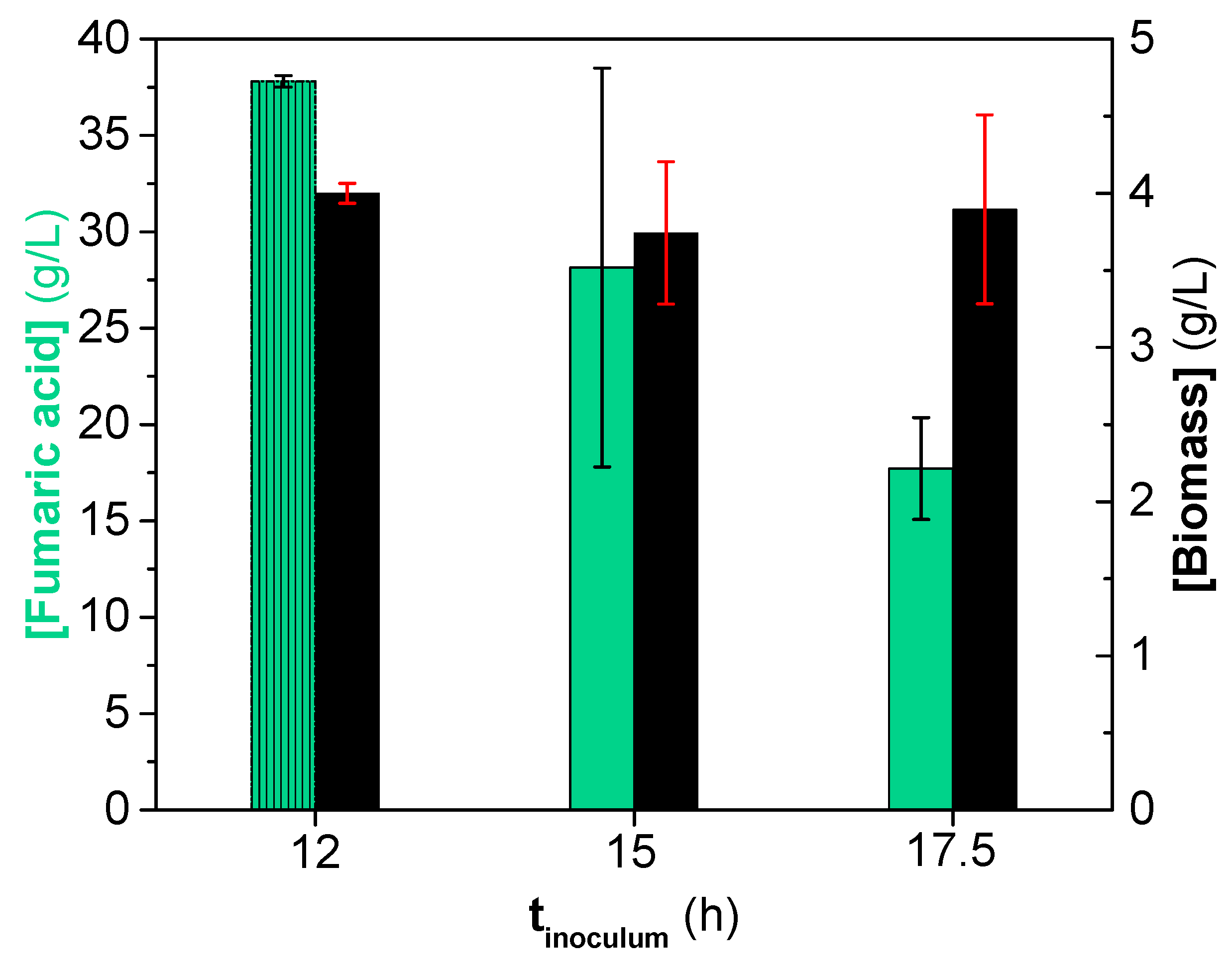
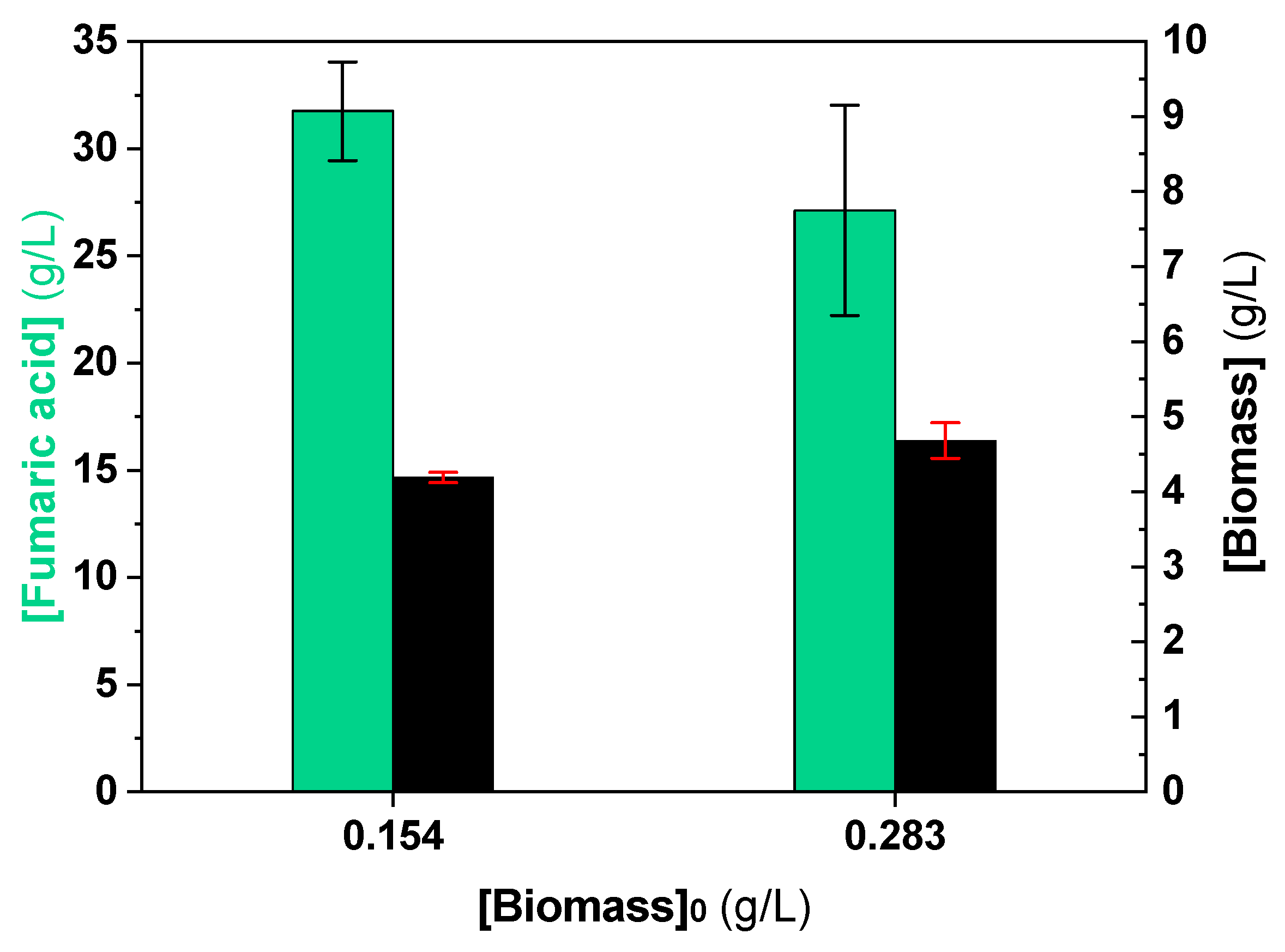
3.3. Influence of Inoculum Stages on Fumaric Acid Production
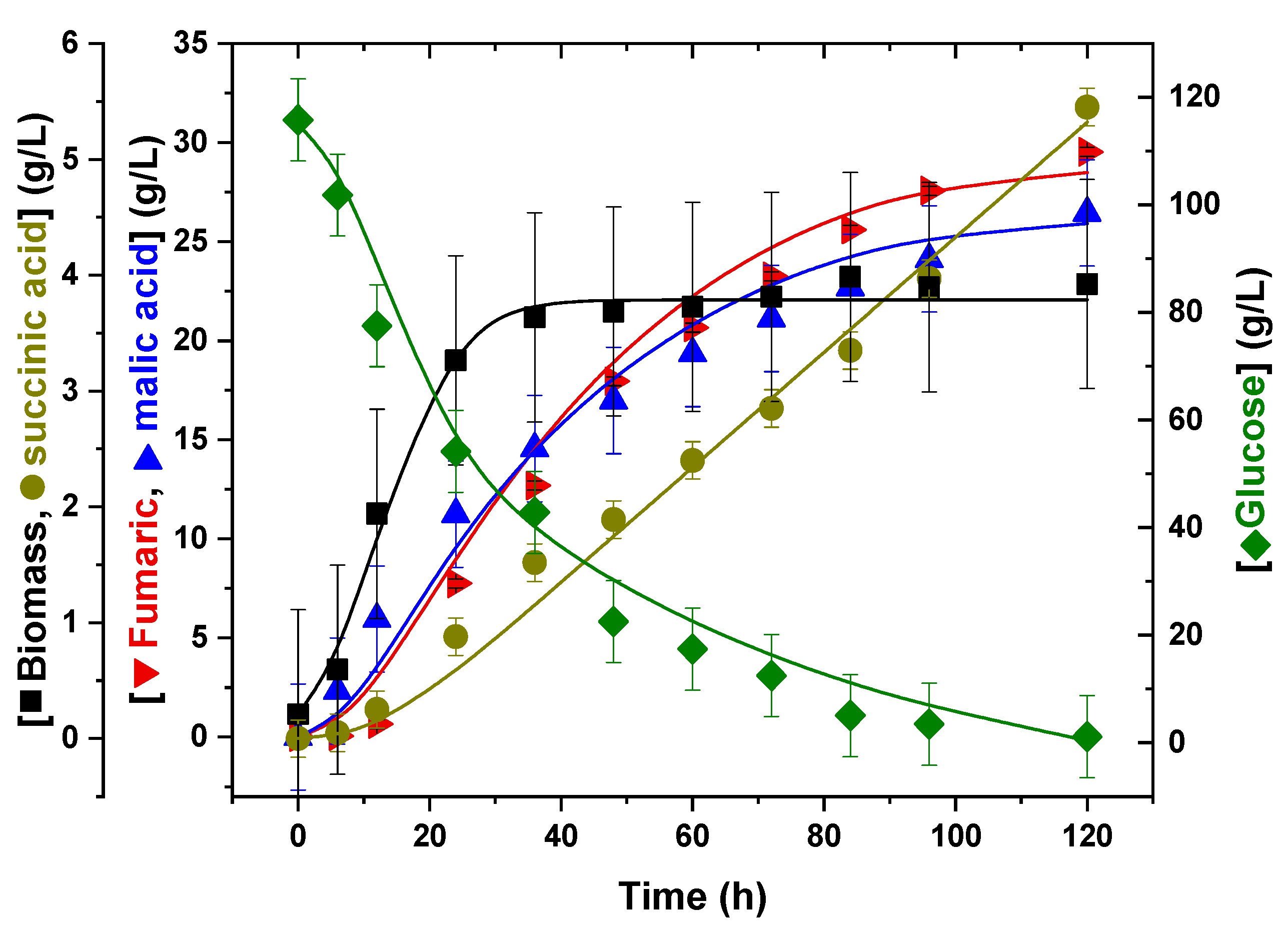
3.4. Kinetic Modelling
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Nomenclature
| CG | Glucose concentration (gglucose/L) |
| Cx | Biomass concentration (gbiomass/L) |
| CXm | Biomass concentration at stationary phase (logistic equation parameter) (gbiomass/L) |
| F95 | Fisher distribution parameter with 95% confident |
| HPLC | High Performance Liquid Chromatography |
| t | Time (h) |
| mS | Reaction rate coefficient of maintenance reaction (L∙h−1∙gbiomass−1) |
| kF | Reaction rate coefficient of fumaric acid production (gfumaric acid∙L∙h−1∙gglucose−1∙gbiomass−1) |
| kM | Reaction rate coefficient of malic acid production (gmalic acid∙L∙h−1∙gglucose−1∙gbiomass−1) |
| kS | Reaction rate coefficient of succinic acid production (gsuccinic acid∙L∙h−1∙gbiomass−1) |
| r1 | Reaction rate of reaction 1 (biomass production) (gbiomass/(L∙h)) |
| r2 | Reaction rate of reaction 2 (maintenance reaction) (-gglucose/(L∙h)) |
| r3 | Reaction rate of reaction 3 (fumaric acid production) (gfumaric acid/(L∙h)) |
| r4 | Reaction rate of reaction 4 (malic acid production) (gmalic acid/(L∙h)) |
| r5 | Reaction rate of reaction 5 (succinic acid production) (gsuccinic acid/(L∙h)) |
| RMSE | Root Mean Squared Error |
| RF | Reaction rate of fumaric acid production (gfumaric acid/(L∙h)) |
| RG | Reaction rate of glucose consumption (gglucose/(L∙h)) |
| RM | Reaction rate of malic acid production (gmalic acid/(L∙h)) |
| RS | Reaction rate of succinic acid production (gsuccinic acid/(L∙h)) |
| RX | Reaction rate of biomass production (gbiomass/(L∙h)) |
| SSR | Sum of Squared Residuals |
| YGF | Glucose consumption to fumaric acid production parameter (gglucose/gfumaric acid) |
| YGM | Glucose consumption to malic acid production parameter (gglucose/gmalic acid) |
| YGS | Glucose consumption to succinic acid production parameter (gglucose/gsuccinic acid) |
| YGX | Glucose consumption to biomass production parameter (gglucose/gbiomass) |
| YMX | Malic acid production associated to biomass growth parameter (gmalic acid/gbiomass) |
| VE | Variation Explained (%) |
| WWII | World War II |
| µ | Specific growth rate (h−1) |
References
- Parajuli, R.; Dalgaard, T.; Jørgensen, U.; Adamsen, A.P.S.; Knudsen, M.T.; Birkved, M.; Gylling, M.; Schjørring, J.K. Biorefining in the prevailing energy and materials crisis: A review of sustainable pathways for biorefinery value chains and sustainability assessment methodologies. Renew. Sustain. Energy Rev. 2015, 43, 244–263. [Google Scholar] [CrossRef]
- Moncada, B.J.; Aristizábal, M.V.; Cardona, A.C.A. Design strategies for sustainable biorefineries. Biochem. Eng. J. 2016, 116, 122–134. [Google Scholar] [CrossRef]
- Nanda, S.; Rana, R.; Sarangi, P.K.; Dalai, A.K.; Kozinski, J.A. A broad introduction to first-, second-, and third-generation biofuels. Recent Adv. Biofuels Bioenergy Util. 2018, 1–25. [Google Scholar] [CrossRef]
- Straathof, A.J.J.; Wahl, S.A.; Benjamin, K.R.; Takors, R.; Wierckx, N.; Noorman, H.J. Grand Research Challenges for Sustainable Industrial Biotechnology. Trends Biotechnol. 2019, 37, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Esteban, J.; Ladero, M. Food waste as a source of value-added chemicals and materials: A biorefinery perspective. Int. J. Food Sci. Technol. 2018, 53, 1095–1108. [Google Scholar] [CrossRef]
- Sebastian, J.; Hegde, K.; Kumar, P.; Rouissi, T.; Brar, S.K. Bioproduction of fumaric acid: An insight into microbial strain improvement strategies. Crit. Rev. Biotechnol. 2019, 39, 817–834. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, P.; Garcia-Garcia, G.; Angelis-Dimakis, A.; Campbell, G.M.; Rahimifard, S. Synergies in the co-location of food manufacturing and biorefining. Food Bioprod. Process. 2019, 117, 340–359. [Google Scholar] [CrossRef]
- Martin-Dominguez, V.; Estevez, J.; De Borja Ojembarrena, F.; Santos, V.E.; Ladero, M. Fumaric acid production: A biorefinery perspective. Fermentation 2018, 4, 33. [Google Scholar] [CrossRef]
- Roa Engel, C.A.; Straathof, A.J.J.; Zijlmans, T.W.; Van Gulik, W.M.; Van Der Wielen, L.A.M. Fumaric acid production by fermentation. Appl. Microbiol. Biotechnol. 2008, 78, 379–389. [Google Scholar] [CrossRef]
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass Volume I—Results of Screening for Potential Candidates from Sugars and Synthesis Gas Energy Efficiency and Renewable Energy; Northwest Natl. Lab.: Golden, CO, USA, 1992.
- Figueira, D.; Cavalheiro, J.; Sommer Ferreira, B. Purification of polymer-grade fumaric acid from fermented spent sulfite liquor. Fermentation 2017, 3, 13. [Google Scholar] [CrossRef]
- Papadaki, A.; Papapostolou, H.; Alexandri, M.; Kopsahelis, N.; Papanikolaou, S.; de Castro, A.M.; Freire, D.M.G.; Koutinas, A.A. Fumaric acid production using renewable resources from biodiesel and cane sugar production processes. Environ. Sci. Pollut. Res. 2018, 25, 35960–35970. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, R.A.; Moyer, A.J.; Smith, M.L.; Kelley, S.E. Production of fumaric acid by Rhizopus arrhizus. Appl. Microbiol. 1959, 7, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, R.A.; Lagoda, A.A.; Misenheimer, T.J.; Smith, M.L.; Anderson, R.F.; Jackson, R.W. Production of Fumaric Acid in 20-Liter Fermentors. Appl. Microbiol. 1962, 10, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Ng, T. Fermentation Process for Carboxylic Acids. US Patent 4,877,731, 31 October 1989. [Google Scholar]
- Xu, Q.; Li, S.; Huang, H.; Wen, J. Key technologies for the industrial production of fumaric acid by fermentation. Biotechnol. Adv. 2012, 30, 1685–1696. [Google Scholar] [CrossRef]
- Eidt, L.; Kuenz, A.; Prüße, U. Biotechnologische produktion von fumarsäure: Prozessoptimierung und kontrolle der morphologie. Chemie Ing. Technik 2018, 90, 1272. [Google Scholar] [CrossRef]
- Naude, A.; Nicol, W. Malic acid production through the whole-cell hydration of fumaric acid with immobilised Rhizopus oryzae. Biochem. Eng. J. 2018, 137, 152–161. [Google Scholar] [CrossRef]
- Sun, C.; Xu, L.; Chen, X.; Qiu, T.; Zhou, T. Sustainable recovery of valuable metals from spent lithium-ion batteries using DL-malic acid: Leaching and kinetics aspect. Waste Manag. Res. 2018, 36, 113–120. [Google Scholar] [CrossRef]
- Pinto Carneiro, S.; Moine, L.; Tessier, B.; Nicolas, V.; dos Santos, O.; dos Santos, D.H.; Fattal, E. Pyrazinoic acid-Poly(malic acid) biodegradable nanoconjugate for efficient intracellular delivery. Precis. Nanomed. 2019, 2, 303–317. [Google Scholar] [CrossRef]
- Castro, V.I.; Mano, F.; Reis, R.L.; Paiva, A.; Duarte, A.R.C. Synthesis and physical and thermodynamic properties of lactic acid and malic acid-based natural deep eutectic solvents. J. Chem. Eng. Data 2019, 63, 2548–2556. [Google Scholar] [CrossRef]
- Naude, A.; Nicol, W. Fumaric acid fermentation with immobilised Rhizopus oryzae: Quantifying time-dependent variations in catabolic flux. Process Biochem. 2017, 56, 8–20. [Google Scholar] [CrossRef]
- Zhang, K.; Yang, S.T.; Chalmers, J.J.; Wood, D. Fumaric Acid Fermentation by Rhizopus oryzae with Integrated Separation Technologies. Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2012; pp. 1–210. [Google Scholar]
- Acedos, M.G.; Ramon, A.; de la Morena, S.; Santos, V.E.; Garcia-Ochoa, F. Isobutanol production by a recombinant biocatalyst Shimwellia blattae (p424IbPSO): Study of the operational conditions. Biochem. Eng. J. 2018, 133, 21–27. [Google Scholar] [CrossRef]
- De la Morena, S.; Acedos, M.G.; Santos, V.E.; Garcia-Ochoa, F. Dihydroxyacetone production from glycerol using Gluconobacter oxydans: Study of medium composition and operational conditions in shaken flasks. Biotechnol. Prog. 2019, 35, e2803. [Google Scholar] [CrossRef] [PubMed]
- De la Torre, I.; Ladero, M.; Santos, V.E. Production of d-lactic acid by Lactobacillus delbrueckii ssp. delbrueckii from orange peel waste: Techno-economical assessment of nitrogen sources. Appl. Microbiol. Biotechnol. 2018, 102, 10511–10521. [Google Scholar] [PubMed]
- Zhang, K.; Yu, C.; Yang, S.T. Effects of soybean meal hydrolysate as the nitrogen source on seed culture morphology and fumaric acid production by Rhizopus oryzae. Process Biochem. 2015, 50, 173–179. [Google Scholar] [CrossRef]
- Papadaki, A.; Androutsopoulos, N.; Patsalou, M.; Koutinas, M.; Kopsahelis, N.; de Castro, A.M.; Papanikolaou, S.; Koutinas, A.A. Biotechnological production of fumaric acid: The effect of morphology of Rhizopus arrhizus NRRL 2582. Fermentation 2017, 3, 33. [Google Scholar] [CrossRef]
- Zhou, Z.; Du, G.; Hua, Z.; Zhou, J.; Chen, J. Optimization of fumaric acid production by Rhizopus delemar based on the morphology formation. Biores. Technol. 2011, 102, 9345–9349. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, L.; Yang, S.T. Fumaric acid recovery and purification from fermentation broth by activated carbon adsorption followed with desorption by acetone. Ind. Eng. Chem. Res. 2014, 53, 12802–12808. [Google Scholar] [CrossRef]
- Cao, N.; Du, J.; Gong, C.S.; Tsao, G.T. Simultaneous production and recovery of fumaric acid from immobilized Rhizopus oryzae with a rotary biofilm contactor and an adsorption column. Appl. Environ. Microbiol. 1996, 62, 2926–2931. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium A | Medium B | Medium C | ||
|---|---|---|---|---|
| Component | Concentration (g/L) | Component | Concentration (g/L) | Concentration (g/L) |
| Glucose | 4 | Glucose | 40 | 130 |
| Urea | 0.6 | (NH4)2SO4 | 4 | 1.8 |
| MgSO4·7H2O | 0.3 | MgSO4·7H2O | 0.4 | 0.4 |
| ZnSO4·7H2O | 0.088 | ZnSO4·7H2O | 0.044 | 0.44 |
| K2PO4 | 0.4 | K2PO4 | 1.6 | 0.3 |
| FeSO4·7H2O | 0.25 | FeCl3·6H2O | 0.0075 | 0.0075 |
| CuSO4·5H2O | 0.00782 | Tartaric acid | 0.0084 | 0.0075 |
| MnSO4·H2O | 0.038 | CaCO3 | ― | 50 |
| Lactose × H2O | 6.32 | Corn steep liquor | 0.5 mL/L | 0.5 mL/L |
| KCl | 0.4 | |||
| NaCl | 40 | |||
| Agar-agar | 30 | |||
| Peptone | 1.6 | |||
| Corn steep liquor | 1 mL/L | |||
| Glycerol | 10 mL/L | |||
| Parameter | Units | Value | ||
|---|---|---|---|---|
| CXm | gbiomass/L | 3.79 | ± | 0.35 |
| h−1 | 0.23 | ± | 0.15 | |
| mS | L∙h−1∙gbiomass−1 | 3.47 × 10−2 | ± | 8.43 × 10−3 |
| kF | gfumaric acid∙L∙h−1∙gglucose−1∙gbiomass−1 | 2.80 × 10−3 | ± | 1.25 × 10−3 |
| kM | gmalic acid∙L∙h−1∙gglucose−1∙gbiomass−1 | 2.29 × 10−3 | ± | 1.81 × 10−3 |
| kS | gsuccinic acid∙L∙h−1∙gbiomass−1 | 1.31 × 10−2 | ± | 3.95 × 10−3 |
| YGF | gglucose/gfumaric acid | 6.38 × 10−1 | ± | 1.16 × 10−2 |
| YGM | gglucose/gmalic acid | 1.07 | ± | 0.52 |
| YGS | gglucose/gsuccinic acid | 1.65 | ± | 0.34 |
| YGX | gglucose/gbiomass | 14.4 | ± | 3.34 |
| YMX | gmalic acid/gbiomass | 0.73 | ± | 0.33 |
| Parameter | Value |
|---|---|
| Degrees of freedom | 45 |
| Root Mean Square Error (RMSE) | 21.13 |
| F95 | 37.76 |
| Variation Explained (VE) (%) | 98.12 |
| Sum of Squared Residuals (SSR) | 359.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Dominguez, V.; Bouzas-Santiso, L.; Martinez-Peinado, N.; Santos, V.E.; Ladero, M. Kinetic Modelling of the Coproduction Process of Fumaric and Malic Acids by Rhizopus arrhizus NRRL 1526. Processes 2020, 8, 188. https://doi.org/10.3390/pr8020188
Martin-Dominguez V, Bouzas-Santiso L, Martinez-Peinado N, Santos VE, Ladero M. Kinetic Modelling of the Coproduction Process of Fumaric and Malic Acids by Rhizopus arrhizus NRRL 1526. Processes. 2020; 8(2):188. https://doi.org/10.3390/pr8020188
Chicago/Turabian StyleMartin-Dominguez, Victor, Laura Bouzas-Santiso, Nieves Martinez-Peinado, Victoria E. Santos, and Miguel Ladero. 2020. "Kinetic Modelling of the Coproduction Process of Fumaric and Malic Acids by Rhizopus arrhizus NRRL 1526" Processes 8, no. 2: 188. https://doi.org/10.3390/pr8020188
APA StyleMartin-Dominguez, V., Bouzas-Santiso, L., Martinez-Peinado, N., Santos, V. E., & Ladero, M. (2020). Kinetic Modelling of the Coproduction Process of Fumaric and Malic Acids by Rhizopus arrhizus NRRL 1526. Processes, 8(2), 188. https://doi.org/10.3390/pr8020188