
The Impact of Essential Oils Derived from Citrus Species to Control Botrytis cinerea and Their Potential Physiological Actions

 , ,
, ,  ,
,  , ,
, ,  ,
,
Abstract
1. Introduction
2. Results
2.1. Characterization of the Essential Oils
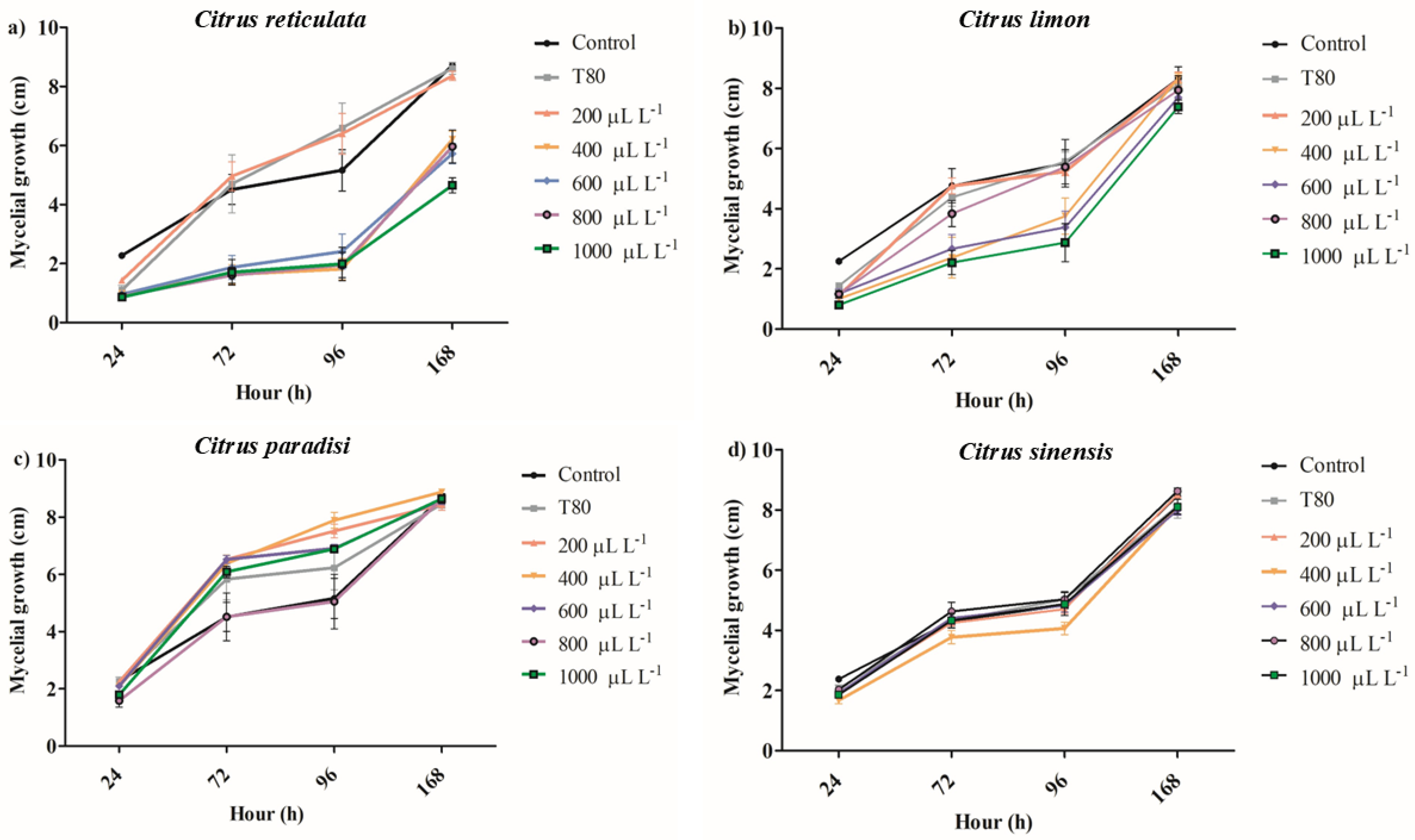
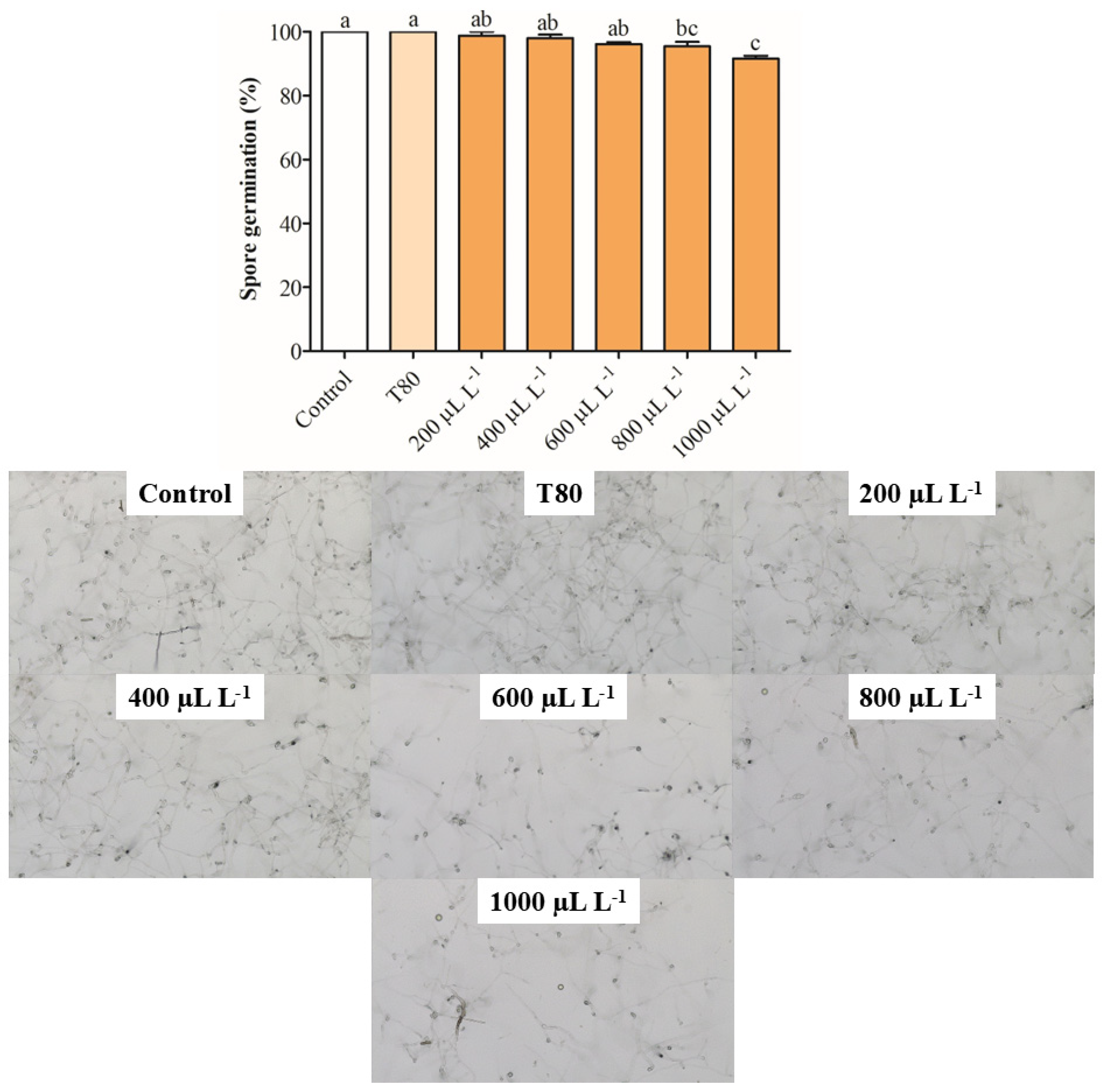
2.2. Inhibition of Mycelial Growth and Spore Germination
2.3. Dry Weight, Electrical Conductivity, and pH
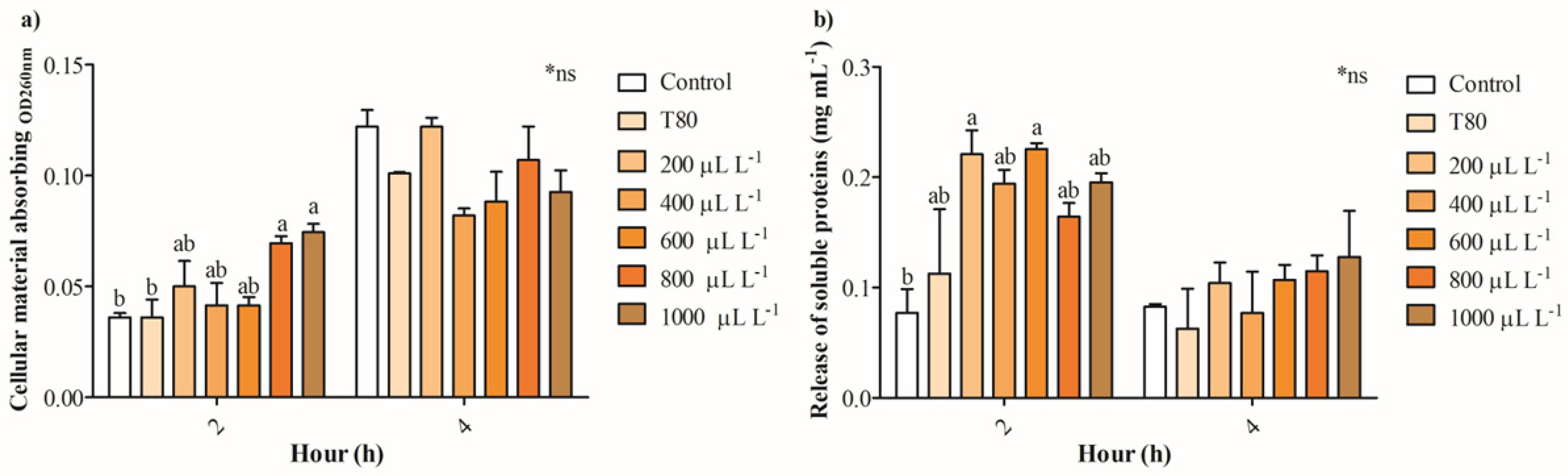
2.4. The Release of Intracellular Constituents and Potential Physiological Mechanisms
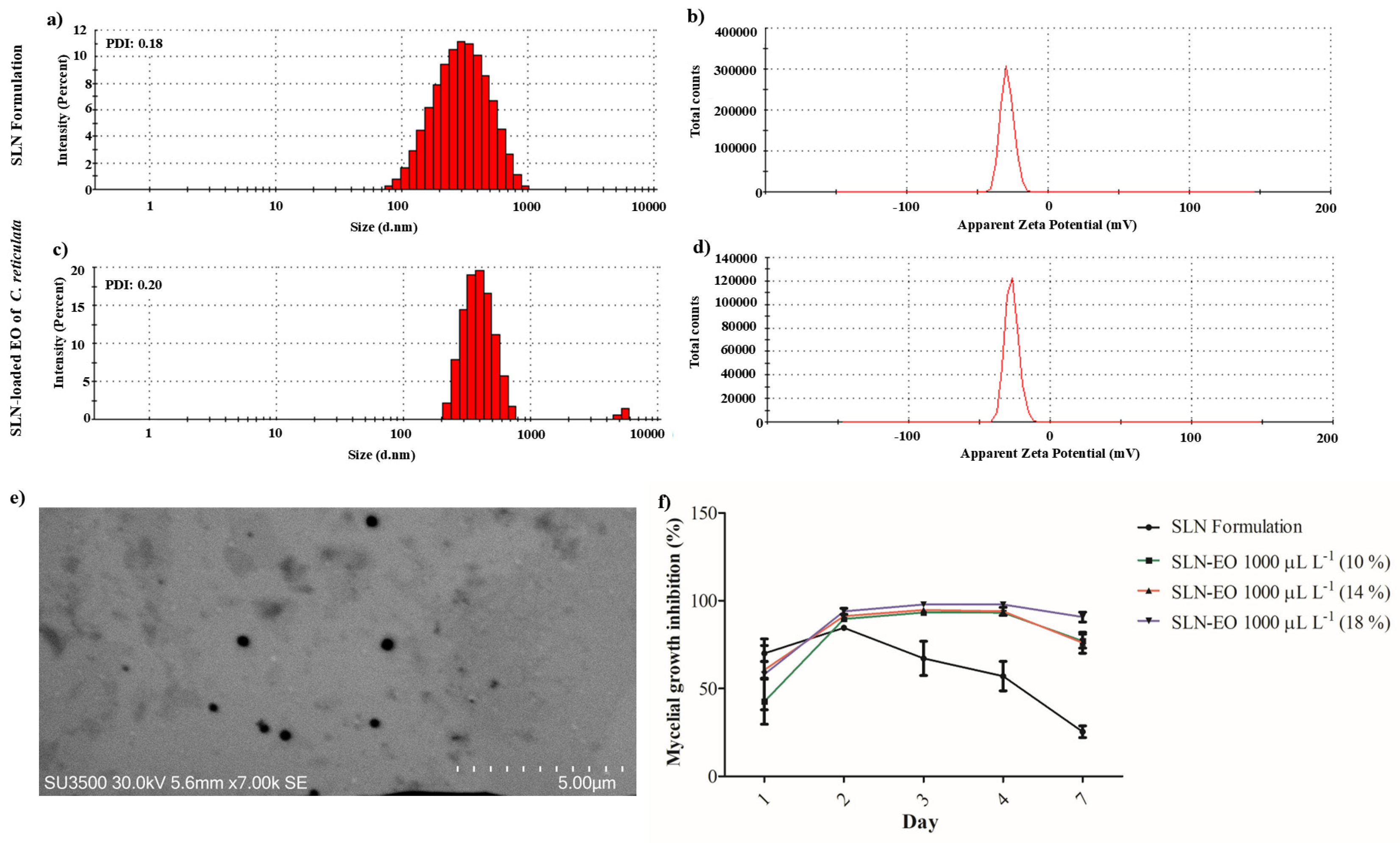
2.5. Characterization and Evaluation of SLN Loaded with the EO of C. reticulata
3. Discussion
4. Materials and Methods
4.1. Characterization of the Essential Oils by GC/MS
4.2. B. cinerea Strain
4.3. Inhibition of B. cinerea Mycelium Growth
4.4. Inhibition of Spore Germination of B. cinerea
4.5. Evaluation of Dry Weight, pH, and Electrical Conductivity to Investigate Potential Alterations in the Cell Membrane
4.6. Release of Constituents Absorbing at OD260nm and Soluble Proteins
4.7. Fluorescent Staining to Explore Mitochondrial Activity, Cell Viability, and Esterase Activity of Cultures of B. cinerea
4.8. Succinate Dehydrogenase Activity
4.9. Formulation and Characterization of SLN Loaded with the EO of C. reticulata
4.10. Antifungal Activity of SLN Loaded with the EO of C. reticulata
4.11. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| EO | Essential oil |
| SLN | Solid lipid nanoparticle |
| GC/MS | Gas–liquid chromatography coupled to mass spectrometry |
| OD | Optical density |
| PDI | Polydispersity index |
| STEM | Scanning transmission electron microscopy |
| GRAS | Generally recognized as safe |
| TCA | Tricarboxylic acid |
| ATP | Adenosine triphosphate |
| MTT | 3- (4,5-Dimethylthiazol-2-yl) 2,5-diphenyltetrazolium |
| FDA | Food and Drug Administration |
| PDA | Potato dextrose agar |
| PDB | Potato dextrose broth |
| PBS | Phosphate-buffered saline |
References
- Dean, R.; van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Dwivedi, M.; Singh, P.; Pandey, A.K. Botrytis fruit rot management: What have we achieved so far? Food Microbiol. 2024, 122, 104564. [Google Scholar] [CrossRef] [PubMed]
- Bi, K.; Liang, Y.; Mengiste, T.; Sharon, A. Killing Softly: A Roadmap of Botrytis cinerea Pathogenicity. Trends Plant Sci. 2023, 28, 211–222. [Google Scholar] [CrossRef]
- Li, T.; Zhou, J.; Li, J. Combined effects of temperature and humidity on the interaction between tomato and Botrytis cinerea revealed by integration of histological characteristics and transcriptome sequencing. Hortic. Res. 2023, 10, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Veloso, J.; Van Kan, J.A.L. Many Shades of Grey in Botrytis–Host Plant Interactions. Trends Plant Sci. 2018, 23, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Cheung, N.; Tian, L.; Liu, X.; Li, X. The Destructive Fungal Pathogen Botrytis cinerea—Insights from Genes Studied with Mutant Analysis. Pathogens 2020, 9, 923. [Google Scholar] [CrossRef]
- Abbey, J.A.; Percival, D.; Abbey, L.; Asiedu, S.K.; Prithiviraj, B.; Schilder, A. Biofungicides as alternative to synthetic fungicide control of grey mould (Botrytis cinerea)—Prospects and challenges. Biocontrol Sci. Technol. 2019, 29, 207–228. [Google Scholar] [CrossRef]
- Olea, A.F.; Bravo, A.; Martínez, R.; Thomas, M.; Sedan, C.; Espinoza, L.; Zambrano, E.; Carvajal, D.; Silva-Moreno, E.; Carrasco, H. Antifungal Activity of Eugenol Derivatives against Botrytis Cinerea. Molecules 2019, 24, 1239. [Google Scholar] [CrossRef]
- Petrasch, S.; Knapp, S.J.; van Kan, J.A.L.; Blanco-Ulate, B. Grey mould of strawberry, a devastating disease caused by the ubiquitous necrotrophic fungal pathogen Botrytis cinerea. Mol. Plant Pathol. 2019, 20, 877–892. [Google Scholar] [CrossRef]
- Allagui, M.B.; Moumni, M.; Romanazzi, G. Antifungal Activity of Thirty Essential Oils to Control Pathogenic Fungi of Postharvest Decay. Antibiotics 2024, 13, 28. [Google Scholar] [CrossRef]
- de Sousa, D.P.; Damasceno, R.O.S.; Amorati, R.; Elshabrawy, H.A.; de Castro, R.D.; Bezerra, D.P.; Nunes, V.R.V.; Gomes, R.C.; Lima, T.C. Essential Oils: Chemistry and Pharmacological Activities. Biomolecules 2023, 13, 1144. [Google Scholar] [CrossRef] [PubMed]
- Bolouri, P.; Salami, R.; Kouhi, S.; Kordi, M.; Asgari Lajayer, B.; Hadian, J.; Astatkie, T. Applications of Essential Oils and Plant Extracts in Different Industries. Molecules 2022, 27, 8999. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Sebghatollahi, Z.; Kamal, M.; Dhyani, A.; Shrivastava, A.; Singh, K.K.; Sinha, M.; Mahato, N.; Mishra, A.K.; Baek, K.-H. Citrus Essential Oils in Aromatherapy: Therapeutic Effects and Mechanisms. Antioxidants 2022, 11, 2374. [Google Scholar] [CrossRef] [PubMed]
- Tabti, L.; El Amine Dib, M.; Djabou, N.; Gaouar Benyelles, N.; Paolini, J.; Costa, J.; Muselli, A. Control of fungal pathogens of Citrus sinensis L. by essential oil and hydrosol of Thymus capitatus L. J. Appl. Bot. Food Qual. 2014, 87, 279–285. [Google Scholar]
- Saini, R.K.; Ranjit, A.; Sharma, K.; Prasad, P.; Shang, X.; Gowda, K.G.M.; Keum, Y.-S. Bioactive Compounds of Citrus Fruits: A Review of Composition and Health Benefits of Carotenoids, Flavonoids, Limonoids, and Terpenes. Antioxidants 2022, 11, 239. [Google Scholar] [CrossRef]
- Simas, D.L.; de Amorim, S.H.; Goulart, F.R.; Alviano, C.S.; Alviano, D.S.; da Silva, A.J.R. Citrus species essential oils and their components can inhibit or stimulate fungal growth in fruit. Ind. Crops Prod. 2017, 98, 108–115. [Google Scholar] [CrossRef]
- Tančinová, D.; Mašková, Z.; Mendelová, A.; Foltinová, D.; Barboráková, Z.; Medo, J. Antifungal Activities of Essential Oils in Vapor Phase against Botrytis cinerea and Their Potential to Control Postharvest Strawberry Gray Mold. Foods 2022, 11, 2945. [Google Scholar] [CrossRef]
- Almawash, S. Solid Lipid Nanoparticles, an Effective Carrier for Classical Antifungal Drugs. Saudi Pharm. J. 2023, 31, 1167–1180. [Google Scholar] [CrossRef]
- Sharma, S.; Mulrey, L.; Byrne, M.; Jaiswal, A.K.; Jaiswal, S. Encapsulation of Essential Oils in Nanocarriers for Active Food Packaging. Foods 2022, 11, 2337. [Google Scholar] [CrossRef]
- Peng, Y.; Li, S.J.; Yan, J.; Tang, Y.; Cheng, J.P.; Gao, A.J.; Yao, X.; Ruan, J.J.; Xu, B.L. Research Progress on Phytopathogenic Fungi and Their Role as Biocontrol Agents. Front. Microbiol. 2021, 12, 670135. [Google Scholar] [CrossRef]
- Rodrigo, S.; García-Latorre, C.; Santamaria, O. Metabolites Produced by Fungi against Fungal Phytopathogens: Review, Implementation and Perspectives. Plants 2022, 11, 81. [Google Scholar] [CrossRef] [PubMed]
- Ekwomadu, T.I.; Mwanza, M. Fusarium Fungi Pathogens, Identification, Adverse Effects, Disease Management, and Global Food Security: A Review of the Latest Research. Agriculture 2023, 13, 1810. [Google Scholar] [CrossRef]
- Jackson-Davis, A.; White, S.; Kassama, L.S.; Coleman, S.; Shaw, A.; Mendonca, A.; Cooper, B.; Thomas-Popo, E.; Gordon, K.; London, L. A Review of Regulatory Standards and Advances in Essential Oils as Antimicrobials in Foods. J. Food Prot. 2023, 86, 100025. [Google Scholar] [CrossRef]
- Basak, S.; Guha, P. A review on antifungal activity and mode of action of essential oils and their delivery as nano-sized oil droplets in food system. J. Food Sci. Technol. 2018, 55, 4701–4710. [Google Scholar] [CrossRef]
- Taheri, P.; Soweizy, M.; Tarighi, S. Application of Essential Oils to Control Some Important Fungi and Bacteria Pathogenic on Cereals. J. Nat. Pestic. Res. 2023, 6, 100052. [Google Scholar] [CrossRef]
- Dosoky, N.S.; Setzer, W.N. Biological Activities and Safety of Citrus spp. Essential Oils. Int. J. Mol. Sci. 2018, 19, 1966. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.; Lei, Z.; Li, L.; Xie, R.; Xi, W.; Guan, Y.; Sumner, L.W.; Zhou, Z. Antifungal activity of citrus essential oils. J. Agric. Food Chem. 2014, 62, 3011–3033. [Google Scholar] [CrossRef]
- Moura, V.S.; Olandin, L.D.; Mariano, B.S.; Rodrigues, J.; Devite, F.T.; Arantes, A.C.C.; Queiroga, C.L.; Sartoratto, A.; de Azevedo, F.A.; Bastianel, M. Antifungal Activity of Citrus Essential Oil in Controlling Sour Rot in Tahiti Acid Lime Fruits. Plants 2024, 13, 3075. [Google Scholar] [CrossRef]
- Malik, S.K.; Kumar, S.; Singh, I.P.; Dhariwal, O.P.; Chaudhury, R. Socio-economic importance, domestication trends and in situ conservation of wild Citrus species of Northeast India. Genet. Resour. Crop Evol. 2013, 60, 1655–1671. [Google Scholar] [CrossRef]
- Palangasinghe, P.C.; Liyanage, W.K.; Wickramasinghe, M.P.; Palangasinghe, H.R.; Shih, H.-C.; Shiao, M.-S.; Chiang, Y.-C. Reviews on Asian citrus species: Exploring traditional uses, biochemistry, conservation, and disease resistance. Ecol. Genet. Genom. 2024, 32, 100269. [Google Scholar] [CrossRef]
- Vasquez-Gomez, K.L.; Mori-Mestanza, D.; Caetano, A.C.; Idrogo-Vasquez, G.; Culqui-Arce, C.; Auquiñivin-Silva, E.A.; Castro-Alayo, E.M.; Cruz-Lacerna, R.; Perez-Ramos, H.A.; Balcázar-Zumaeta, C.R.; et al. Exploring Chemical Properties of Essential Oils from Citrus Peels Using Green Solvent. Heliyon 2024, 10, e40088. [Google Scholar] [CrossRef] [PubMed]
- González-Mas, M.C.; Rambla, J.L.; López-Gresa, M.P.; Blázquez, M.A.; Granell, A. Volatile Compounds in Citrus Essential Oils: A Comprehensive Review. Front. Plant Sci. 2019, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- D’Almeida, R.E.; Sued, N.; Arena, M.E. Citrus paradisi and Citrus reticulata essential oils interfere with Pseudomonas aeruginosa quorum sensing in vivo on Caenorhabditis elegans. Phytomed. Plus 2022, 2, 100160. [Google Scholar] [CrossRef]
- Sanli, I.; Ozkan, G.; Şahin-Yeşilçubuk, N. Green extractions of bioactive compounds from citrus peels and their applications in the food industry. Food Res. Int. 2025, 212, 116352. [Google Scholar] [CrossRef]
- De-Montijo-Prieto, S.; Razola-Díaz, M.d.C.; Gómez-Caravaca, A.M.; Guerra-Hernandez, E.J.; Jiménez-Valera, M.; Garcia-Villanova, B.; Ruiz-Bravo, A.; Verardo, V. Essential Oils from Fruit and Vegetables, Aromatic Herbs, and Spices: Composition, Antioxidant, and Antimicrobial Activities. Biology 2021, 10, 1091. [Google Scholar] [CrossRef]
- Badawy, M.E.I.; Abdelgaleil, S.A.M. Composition and antimicrobial activity of essential oils isolated from Egyptian plants against plant pathogenic bacteria and fungi. Ind. Crops Prod. 2014, 52, 776–782. [Google Scholar] [CrossRef]
- Brito, C.; Hansen, H.; Espinoza, L.; Faúndez, M.; Olea, A.F.; Pino, S.; Díaz, K. Assessing the Control of Postharvest Gray Mold Disease on Tomato Fruit Using Mixtures of Essential Oils and Their Respective Hydrolates. Plants 2021, 10, 1719. [Google Scholar] [CrossRef] [PubMed]
- Mbili, N.C.; Opara, U.L.; Lennox, C.L.; Vries, F.A. Citrus and lemongrass essential oils inhibit Botrytis cinerea on ‘Golden Delicious’, ‘Pink Lady’ and ‘Granny Smith’ apples. J. Plant Dis. Prot. 2017, 124, 499–511. [Google Scholar] [CrossRef]
- Lin, H.; Li, Z.; Sun, Y.; Zhang, Y.; Wang, S.; Zhang, Q.; Cai, T.; Xiang, W.; Zeng, C.; Tang, J. D-Limonene: Promising and Sustainable Natural Bioactive Compound. Appl. Sci. 2024, 14, 4605. [Google Scholar] [CrossRef]
- Zochedh, A.; Priya, M.; Shunmuganarayanan, A.; Thandavarayan, K.; Sultan, A.B. Investigation on structural, spectroscopic, DFT, biological activity and molecular docking simulation of essential oil Gamma-Terpinene. J. Mol. Struct. 2022, 1268, 133651. [Google Scholar] [CrossRef]
- Połeć, K.; Wyżga, B.; Olechowska, K.; Hąc-Wydro, K. On the synergy/antagonism of selected terpenes in the effect on lipid membranes studied in model systems. J. Mol. Liq. 2022, 349, 118473. [Google Scholar] [CrossRef]
- Lee, D.-H.; Lee, M.-W.; Cho, S.B.; Hwang, K.; Park, I.-K. Antifungal mode of action of bay, allspice, and ajowan essential oils and their constituents against Colletotrichum gloeosporioides via overproduction of reactive oxygen species and downregulation of ergosterol biosynthetic genes. Ind. Crops Prod. 2023, 197, 116684. [Google Scholar] [CrossRef]
- Li, J.; Tian, X.; Gao, Y.; Shang, S.; Feng, J.; Zhang, X. A value-added use of volatile turpentine: Antifungal activity and QSAR study of β-pinene derivatives against three agricultural fungi. RSC Adv. 2015, 5, 66947–66955. [Google Scholar] [CrossRef]
- Allizond, V.; Cavallo, L.; Roana, J.; Mandras, N.; Cuffini, A.M.; Tullio, V.; Banche, G. In Vitro Antifungal Activity of Selected Essential Oils against Drug-Resistant Clinical Aspergillus spp. Strains. Molecules 2023, 28, 7259. [Google Scholar] [CrossRef]
- Hou, H.; Zhang, X.; Zhao, T.; Zhou, L. Effects of Origanum vulgare essential oil and its two main components, carvacrol and thymol, on the plant pathogen Botrytis cinerea. PeerJ 2020, 8, e9626. [Google Scholar] [CrossRef]
- Xiong, Y.; Li, S.; Warner, R.D.; Fang, Z. Effect of oregano essential oil and resveratrol nanoemulsion loaded pectin edible coating on the preservation of pork loin in modified atmosphere packaging. Food Control 2020, 114, 107226. [Google Scholar] [CrossRef]
- da Rocha Neto, A.C.; Navarro, B.B.; Canton, L.; Maraschin, M.; Di Piero, R.M. Antifungal activity of palmarosa (Cymbopogon martinii), tea tree (Melaleuca alternifolia) and star anise (Illicium verum) essential oils against Penicillium expansum and their mechanisms of action. LWT-Food Sci. Technol. 2019, 105, 385–392. [Google Scholar] [CrossRef]
- Pavoni, L.; Maggi, F.; Mancianti, F.; Nardoni, S.; Ebani, V.V.; Cespi, M.; Bonacucina, G.; Palmieri, G.F. Microemulsions: An effective encapsulation tool to enhance the antimicrobial activity of selected EOs. J. Drug Deliv. Sci. Technol. 2019, 53, 101101. [Google Scholar] [CrossRef]
- Mani-López, E.; Cortés-Zavaleta, O.; López-Malo, A. A review of the methods used to determine the target site or the mechanism of action of essential oils and their components against fungi. SN Appl. Sci. 2021, 3, 44. [Google Scholar] [CrossRef]
- Kou, Z.; Zhang, J.; Lan, Q.; Liu, L.; Su, X.; Islam, R.; Tian, Y. Antifungal activity and mechanism of palmarosa essential oil against pathogen Botrytis cinerea in the postharvest onions. J. Appl. Microbiol. 2023, 134, lxad290. [Google Scholar] [CrossRef]
- Wang, L.; Hu, W.; Deng, J.; Liu, X.; Zhou, J.; Li, X. Antibacterial activity of Litsea cubeba essential oil and its mechanism against Botrytis cinerea. RSC Adv. 2019, 9, 28987–28995. [Google Scholar] [CrossRef] [PubMed]
- Fincheira, P.; Jofré, I.; Espinoza, J.; Levío-Raimán, M.; Tortella, G.; Oliveira, H.; Diez, M.; Quiroz, A.; Rublar, O. The efficient activity of plant essential oils for inhibiting Botrytis cinerea and Penicillium expansum: Mechanistic insights into antifungal activity. Microbiol. Res. 2023, 277, 127486. [Google Scholar] [CrossRef] [PubMed]
- Rani, P.; Nidhin, P.T.; Ali, S.M.; Bera, A.; Katoch, M. The antifungal mechanism of Monarda citriodora essential oil, hexanal and their combined vapours on Aspergillus foetidus. Biocatal. Agric. Biotechnol. 2023, 54, 102894. [Google Scholar] [CrossRef]
- Medina-Romero, Y.M.; Rodriguez-Canales, M.; Rodriguez-Monroy, M.A.; Hernandez-Hernandez, A.B.; Delgado-Buenrostro, N.L.; Chirino, Y.I.; Cruz-Sanchez, T.; Garcia-Tovar, C.G.; Canales-Martinez, M.M. Effect of the Essential Oils of Bursera morelensis and Lippia graveolens and Five Pure Compounds on the Mycelium, Spore Production, and Germination of Species of Fusarium. J. Fungi 2022, 8, 617. [Google Scholar] [CrossRef]
- Yan, J.; Wu, H.; Chen, K.; Feng, J.; Zhang, Y. Antifungal Activities and Mode of Action of Cymbopogon citratus, Thymus vulgraris, and Origanum heracleoticum Essential Oil Vapors against Botrytis cinerea and Their Potential Application to Control Postharvest Strawberry Gray Mold. Foods 2021, 10, 2451. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Shao, X.; Wei, Y.; Li, Y.; Xu, F.; Wang, H. Solidago canadensis L. Essential Oil Vapor Effectively Inhibits Botrytis cinerea Growth and Preserves Postharvest Quality of Strawberry as a Food Model System. Front. Microbiol. 2016, 7, 1179. [Google Scholar] [CrossRef]
- Uggeri, J.; Gatti, R.; Belletti, S.; Scandroglio, R.; Corradini, R.; Rotoli, B.M.; Orlandini, G. Calcein-AM is a detector of intracellular oxidative activity. Histochem. Cell Biol. 2000, 122, 499–505. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, J.; Kong, W.; Zhao, G.; Yang, M. Mechanisms of antifungal and anti-aflatoxigenic properties of essential oil derived from turmeric (Curcuma longa L.) on Aspergillus flavus. Food Chem. 2007, 220, 1–8. [Google Scholar] [CrossRef]
- Tian, F.; Woo, S.Y.; Lee, S.Y.; Park, S.B.; Zheng, Y.; Chun, H.S. Antifungal Activity of Essential Oil and Plant-Derived Natural Compounds against Aspergillus flavus. Antibiotics 2022, 11, 1727. [Google Scholar] [CrossRef]
- Li, Y.; Shao, X.; Xu, J.; Wei, Y.; Xu, F.; Wang, H. Tea tree oil exhibits antifungal activity against Botrytis cinerea by affecting mitochondria. Food Chem. 2017, 234, 62–67. [Google Scholar] [CrossRef]
- Ghasemi, M.; Turnbull, T.; Sebastian, S.; Kempson, I. The MTT Assay: Utility, Limitations, Pitfalls, and Interpretation in Bulk and Single-Cell Analysis. Int. J. Mol. Sci. 2021, 22, 12827. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, X.; Zhang, H.; Wang, Z.; Xu, H. The research progress in and perspective of potential fungicides: Succinate dehydrogenase inhibitors. Bioorg. Med. Chem. 2021, 50, 116476. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Shao, X.; Li, Y.; Wei, Y.; Xu, F.; Wang, H. Metabolomic Analysis and Mode of Action of Metabolites of Tea Tree Oil Involved in the Suppression of Botrytis cinerea. Front. Microbiol. 2017, 8, 1017. [Google Scholar] [CrossRef]
- Li, Z.; Shao, X.; Wei, Y.; Dai, K.; Xu, J.; Xu, F.; Wang, H. Transcriptome analysis of Botrytis cinerea in response to tea tree oil and its two characteristic components. Appl. Microbiol. Biotechnol. 2020, 104, 2163–2178. [Google Scholar] [CrossRef]
- Sedeek, M.S.; Al-Mahallawi, A.M.; Hussien, R.A.A.; Ali, A.M.A.; Naguib, I.A.; Mansour, M.K. Hexosomal Dispersion: A Nano-Based Approach to Boost the Antifungal Potential of Citrus Essential Oils against Plant Fungal Pathogens. Molecules 2021, 26, 6284. [Google Scholar] [CrossRef]
- Cavanagh, H.M.A. Antifungal Activity of the Volatile Phase of Essential Oils: A Brief Review. Nat. Prod. Commun. 2007, 2, 1297–1302. [Google Scholar] [CrossRef]
- Fuentes, J.M.; Jofré, I.; Tortella, G.; Benavides-Mendoza, A.; Diez, M.C.; Rubilar, O.; Fincheira, P. The mechanistic insights of essential oil of Mentha piperita to control Botrytis cinerea and the prospection of lipid nanoparticles to its application. Microbiol. Res. 2024, 286, 127792. [Google Scholar] [CrossRef]
- Akanda, M.; Mithu, M.D.S.H.; Douroumis, D. Solid lipid nanoparticles: An effective lipid-based technology for cancer treatment. J. Drug Deliv. Sci. Technol. 2023, 86, 104709. [Google Scholar] [CrossRef]
- Takechi-Haraya, Y.; Ohgita, T.; Demizu, Y.; Saito, H.; Izutsu, K.-i.; Sakai-Kato, K. Current Status and Challenges of Analytical Methods for Evaluation of Size and Surface Modification of Nanoparticle-Based Drug Formulations. AAPS PharmSciTech 2022, 23, 150. [Google Scholar] [CrossRef]
- Bouqellah, N.A.; Abdulmajeed, A.M.; Rashed, A.F.K.; Mattar, E.; Al-Sarraj, F.; Abdulfattah, A.M.; Hassan, M.M.; Baazeem, A.; Al-Harthi, H.F.; Musa, A.; et al. Optimizing encapsulation of garlic and cinnamon essential oils in silver nanoparticles for enhanced antifungal activity against Botrytis cinerea pathogenic disease. Physiol. Mol. Plant Pathol. 2025, 136, 102522. [Google Scholar] [CrossRef]
- Silva, E.F.d.; Santos, F.A.L.d.; Pires, H.M.; Bastos, L.M.; Ribeiro, L.N.d.M. Lipid Nanoparticles Carrying Essential Oils for Multiple Applications as Antimicrobials. Pharmaceutics 2025, 17, 178. [Google Scholar] [CrossRef] [PubMed]
- Zhaveh, S.; Mohsenifar, A.; Beiki, M.; Khalili, S.T.; Abdollahi, A.; Rahmani-Cherati, T.; Tabatabaei, M. Encapsulation of Cuminum cyminum essential oils in chitosan-caffeic acid nanogel with enhanced antimicrobial activity against Aspergillus flavus. Ind. Crops Prod. 2015, 69, 251–256. [Google Scholar] [CrossRef]
- Sivalingam, S.; Sharmila, J.S.; Golla, G.; Arunachalam, L.; Singh, T.; Karthikeyan, G.; Shabthi, A.; Malaichamy, K. Encapsulation of essential oil to prepare environment friendly nanobio-fungicide against Fusarium oxysporum f.sp. lycopersici: An experimental and molecular dynamics approach. Colloids Surf. A Physicochem. Eng. Asp. 2024, 681, 132681. [Google Scholar] [CrossRef]
- Vakili-Ghartavol, M.; Arouiee, H.; Golmohammadzadeh, S.; Naseri, M. Synthesis, Characterization, and in Vitro Antifungal Activity of Solid Lipid Nanoparticles Containing Mentha × piperita L. Essential Oil. J. Agric. Sci. Technol. 2024, 26, 667–680. [Google Scholar]
- Espinoza, J.; Soto, I.; Arriagada, J.; Lizama, M.; Aninao, N.; Aniñir, W.; Ungerfeld, E.M.; Chacón-Fuentes, M.; Quiroz, A. Volatile Cues from Fresh Cattle Dung Can Drive Horn Fly Egg-Laying and Fecal Attraction to Horn Flies, Haematobia irritans (Diptera: Muscidae). Insects 2025, 16, 129. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Kovats, E.; Keulemans, A.I.M. The Kováts Retention Index System. Anal. Chem. 1964, 36, 31A–41A. [Google Scholar]
- Areco, A.V.; Achimón, F.; Almirón, C.; Nally, C.M.; Zunino, P.M.; Yaryura, P. Antifungal activity of essential oils rich in ketones against Botrytis cinerea: New strategy for biocontrol. Biocatal. Agric. Biotechnol. 2024, 59, 103233. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, S.; Du, S.; Chen, S.; Sun, H. Antifungal activity of thymol and carvacrol against postharvest pathogens Botrytis cinerea. J. Food Sci. Technol. 2019, 56, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Plaza, V.; Pasten, A.; López-Ramírez, L.A.; Mora-Montes, H.M.; Rubio-Astudillo, J.; Silva-Moreno, E.; Castillo, L. Botrytis cinerea PMT4 Is Involved in O-Glycosylation, Cell Wall Organization, Membrane Integrity, and Virulence. J. Fungi 2025, 11, 71. [Google Scholar] [CrossRef]
- Lai, Q.; Sun, X.; Li, L.; Li, D.; Wang, M.; Shi, H. Toxicity effects of procymidone, iprodione and their metabolite of 3,5-dichloroaniline to zebrafish. Chemosphere 2021, 272, 129577. [Google Scholar] [CrossRef]
- Ji, D.; Chen, T.; Ma, D.; Liu, J.; Xu, Y.; Tian, S. Inhibitory effects of methyl thujate on mycelial growth of Botrytis cinerea and possible mechanisms. Postharvest Biol. Technol. 2018, 142, 46–54. [Google Scholar] [CrossRef]
- Freimoser, F.M.; Jakob, C.A.; Aebi, M.; Tuor, U. The MTT [3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide] Assay Is a Fast and Reliable Method for Colorimetric Determination of Fungal Cell Densities. Appl. Environ. Microbiol. 1999, 65, 3727–3729. [Google Scholar] [CrossRef] [PubMed]
- Fincheira, P.; Quiroz, A.; Medina, C.; Tortella, G.; Hermosilla, E.; Diez, M.C.; Rubilar, O. Plant growth induction by volatile organic compound released from solid lipid nanoparticles and nanostructured lipid carriers. Colloids Surf. A Physicochem. Eng. Asp. 2020, 596, 124739. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT | RI | Compound | C. reticulata (%) | C. limon (%) | C. paradisi (%) | C. sinensis (%) | Identification |
|---|---|---|---|---|---|---|---|
| 8.65 | 929 | α-Pinene | 2.54 | 2.58 | 0.57 | 0.42 | RI, MS, Std-I |
| 9.05 | 943 | Camphene | - | 0.03 | - | - | RI, MS |
| 9.83 | 969 | β-Pinene | 2.23 | 21.89 | 0.12 | 0.14 | RI, MS, Std-I |
| 10.79 | 998 | β-Myrcene | 0.61 | 0.61 | 0.35 | 0.18 | RI, MS |
| 11.33 | 1017 | o-Cymene | 1.08 | 0.17 | - | - | RI, MS |
| 11.51 | 1024 | Limonene | 66.61 | 59.65 | 98.51 | 95.23 | RI, MS, Std-I |
| 12.37 | 1054 | γ-Terpinene | 26.29 | 12.85 | - | - | RI, MS, Std-I |
| 15.05 | 1144 | Limonene epoxide | - | - | 3.08 | RI, MS | |
| 19.32 | 1294 | Carvone | - | - | - | 0.83 | RI, MS |
| 21.60 | 1382 | Copaene | 0.10 | - | 0.09 | 0.03 | RI, MS |
| 22.63 | 1422 | Caryophyllene | 0.06 | 0.66 | 0.21 | RI, MS | |
| 22.92 | 1434 | β-copaene | 0.03 | 0.06 | 0.01 | RI, MS | |
| 23.09 | 1441 | α-Bergamotene | - | 0.68 | - | - | RI, MS |
| 23.49 | 1457 | Humulene | 0.02 | 0.03 | 0.02 | - | RI, MS |
| 24.49 | 1497 | Valencene | - | 0.05 | - | - | RI, MS |
| 26.49 | 1582 | Caryophyllene oxide | 0.07 | 0.09 | 0.02 | - | RI, MS |
| Identified compounds | 99.64 | 99.29 | 99.95 | 99.92 | |||
| Total Monoterpenes | 99.36 | 97.78 | 99.55 | 99.88 | |||
| Monoterpene hydrocarbons | 99.36 | 97.78 | 99.55 | 95.97 | |||
| Oxygenated monoterpenes | - | - | - | 3.91 | |||
| Total Sesquiterpenes | 0.28 | 1.51 | 0.40 | 0.04 | |||
| Sesquiterpene hydrocarbons | 0.21 | 1.42 | 0.38 | 0.04 | |||
| Oxygenated sesquiterpenes | 0.07 | 0.09 | 0.02 | - | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campos, S.; Espinoza, J.; Fuentes, J.M.; Jofré-Fernández, I.; Tortella, G.; Navarro, D.; Quiroz, A.; Diez, M.C.; Rubilar, O.; Fincheira, P. The Impact of Essential Oils Derived from Citrus Species to Control Botrytis cinerea and Their Potential Physiological Actions. Plants 2025, 14, 1859. https://doi.org/10.3390/plants14121859
Campos S, Espinoza J, Fuentes JM, Jofré-Fernández I, Tortella G, Navarro D, Quiroz A, Diez MC, Rubilar O, Fincheira P. The Impact of Essential Oils Derived from Citrus Species to Control Botrytis cinerea and Their Potential Physiological Actions. Plants. 2025; 14(12):1859. https://doi.org/10.3390/plants14121859
Chicago/Turabian StyleCampos, Sebastián, Javier Espinoza, Juan Mauricio Fuentes, Ignacio Jofré-Fernández, Gonzalo Tortella, Diego Navarro, Andrés Quiroz, María Cristina Diez, Olga Rubilar, and Paola Fincheira. 2025. "The Impact of Essential Oils Derived from Citrus Species to Control Botrytis cinerea and Their Potential Physiological Actions" Plants 14, no. 12: 1859. https://doi.org/10.3390/plants14121859
APA StyleCampos, S., Espinoza, J., Fuentes, J. M., Jofré-Fernández, I., Tortella, G., Navarro, D., Quiroz, A., Diez, M. C., Rubilar, O., & Fincheira, P. (2025). The Impact of Essential Oils Derived from Citrus Species to Control Botrytis cinerea and Their Potential Physiological Actions. Plants, 14(12), 1859. https://doi.org/10.3390/plants14121859