

The Genetic Control of SEEDSTICK and LEUNIG-HOMOLOG in Seed and Fruit Development: New Insights into Cell Wall Control
 , , ,
, , ,  , , and
, , and
Abstract
1. Introduction
2. Results
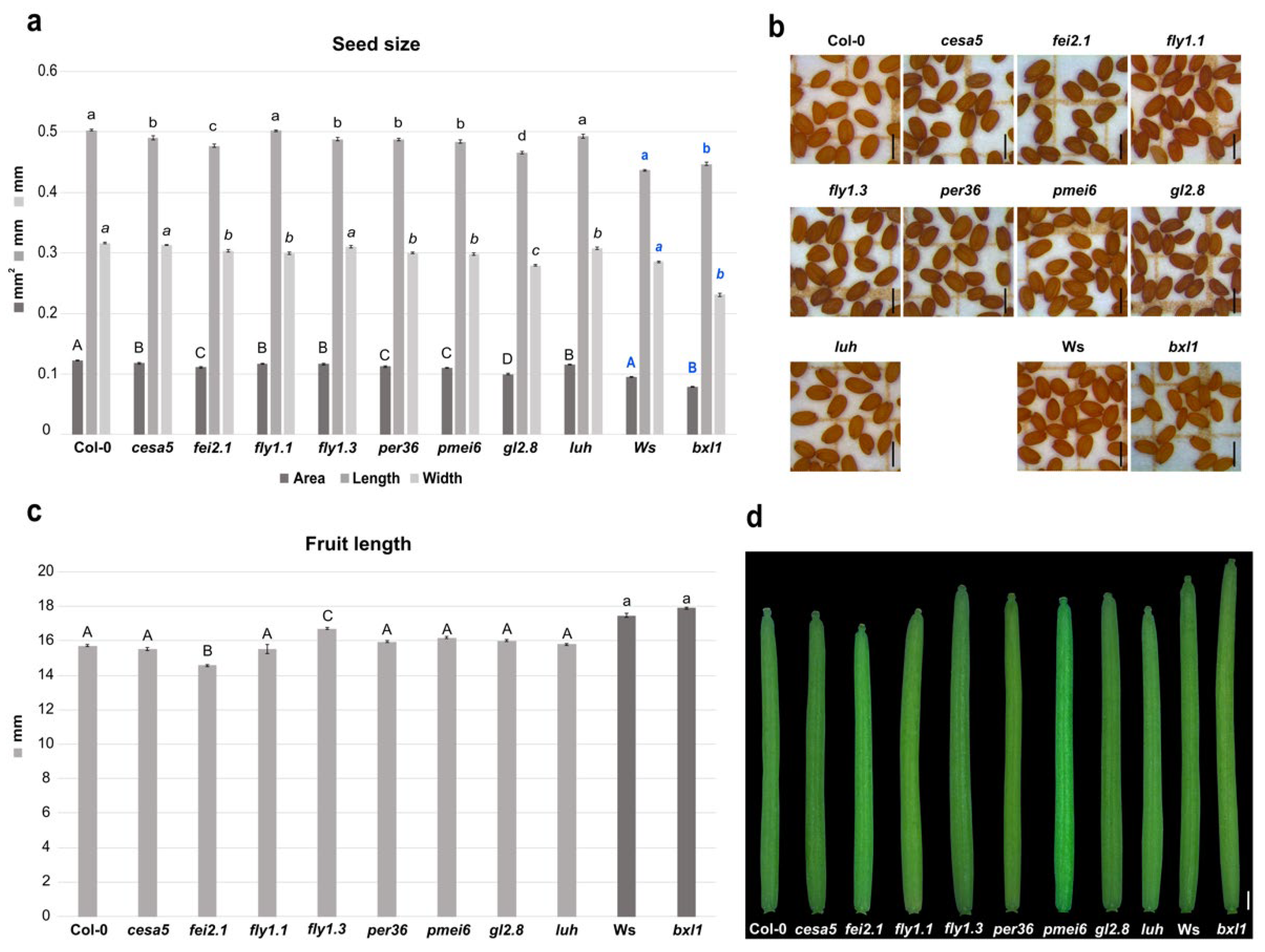
2.1. Cell Wall Structural Alterations Influence Seed and Silique Size
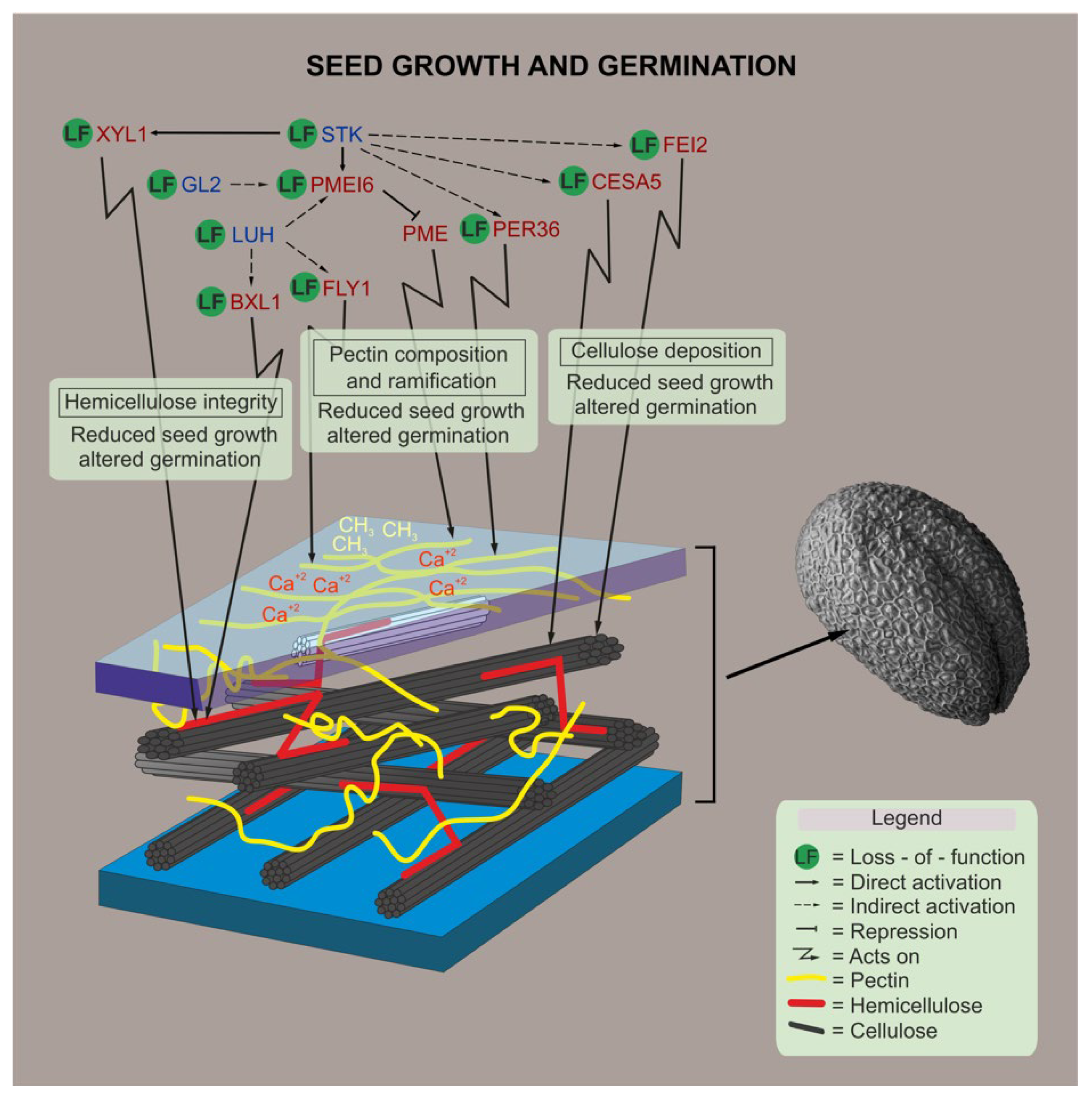
2.2. Transcriptional Cascades Controlling Cell Wall Modifications Are Important for Seed and Fruit Development
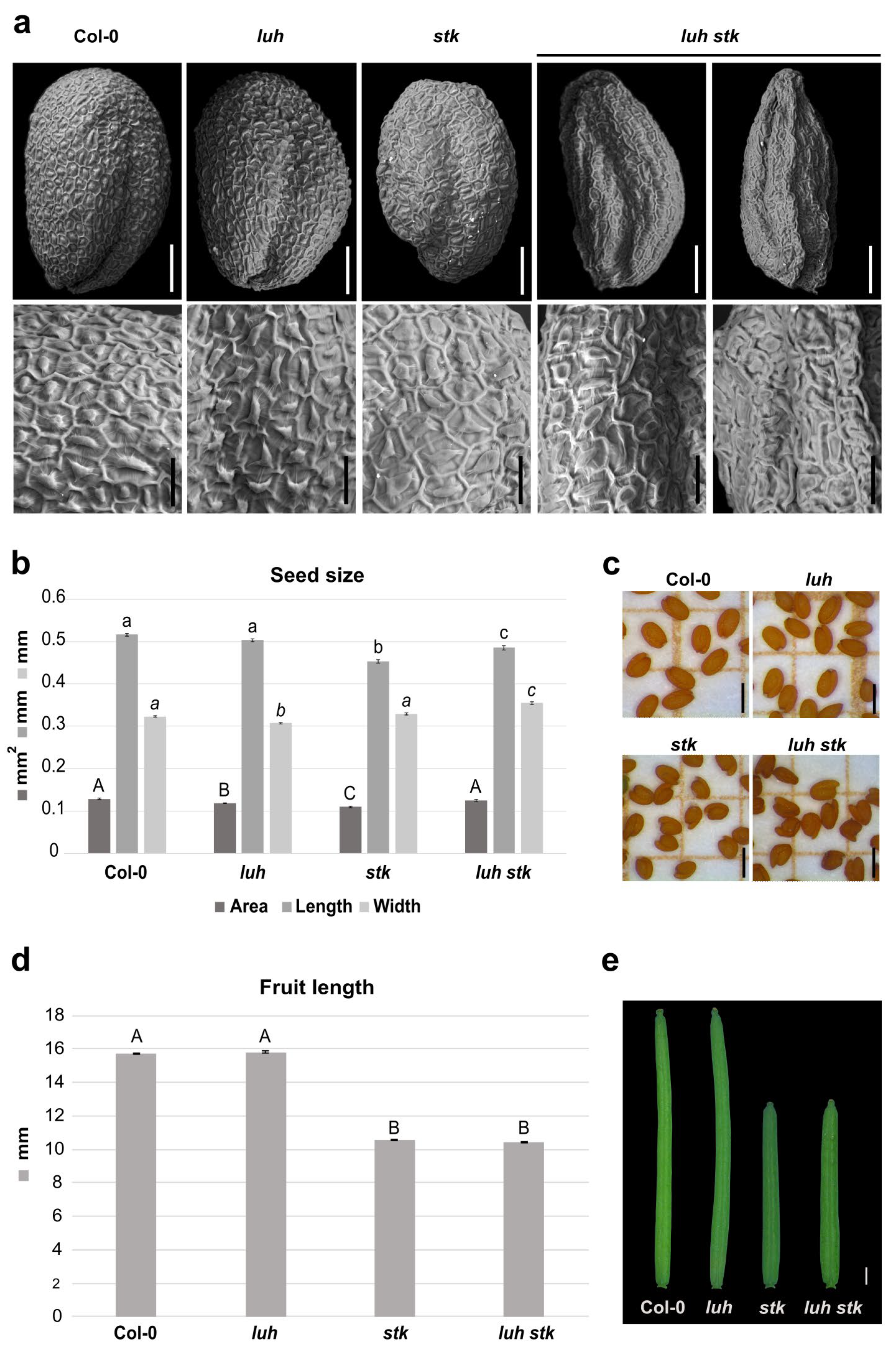
2.3. Dual Disruption of STK and LUH Impacts Seed Development
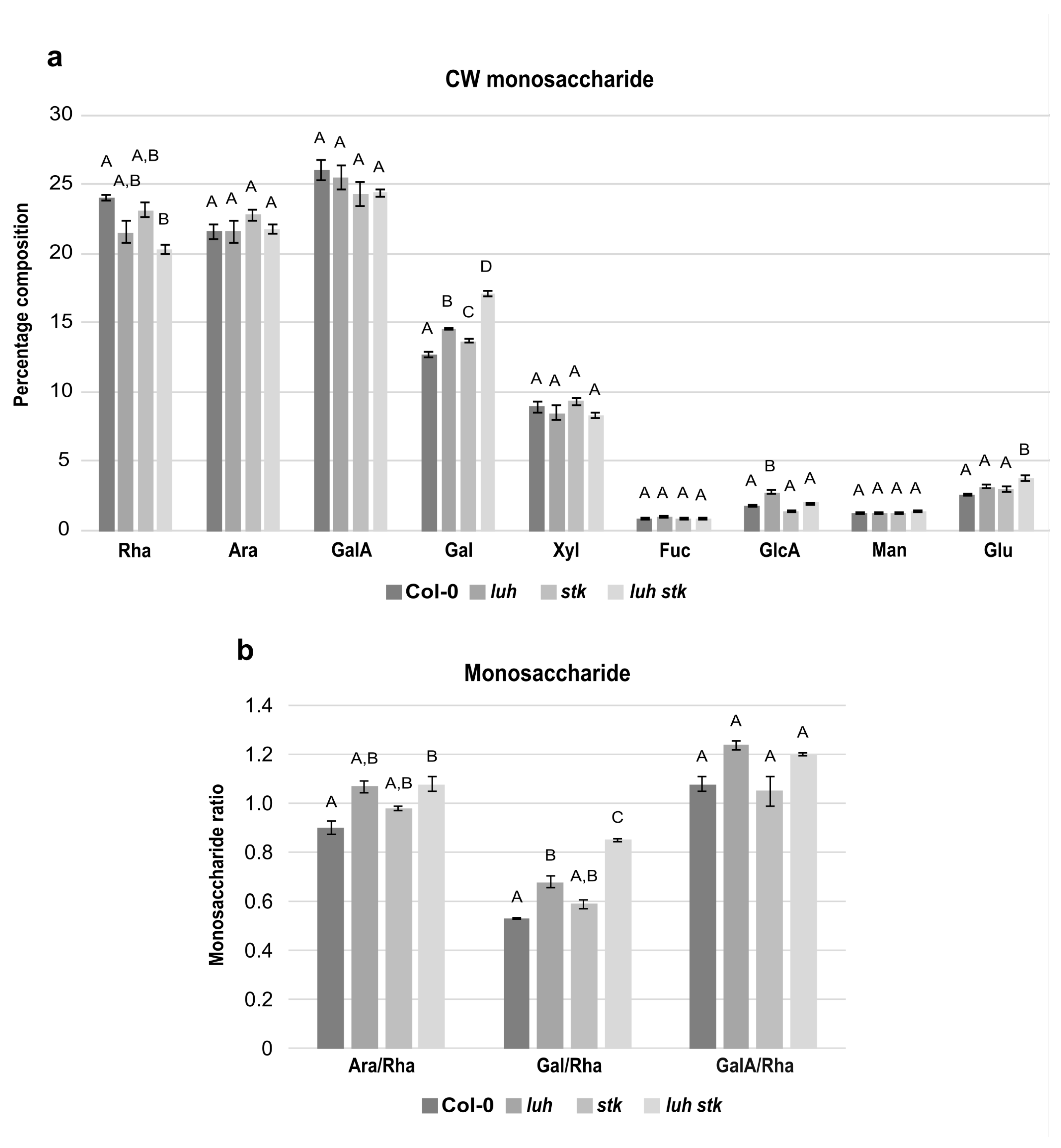
2.4. The stk and luh Mutant Seeds Have Altered Cell Wall Composition
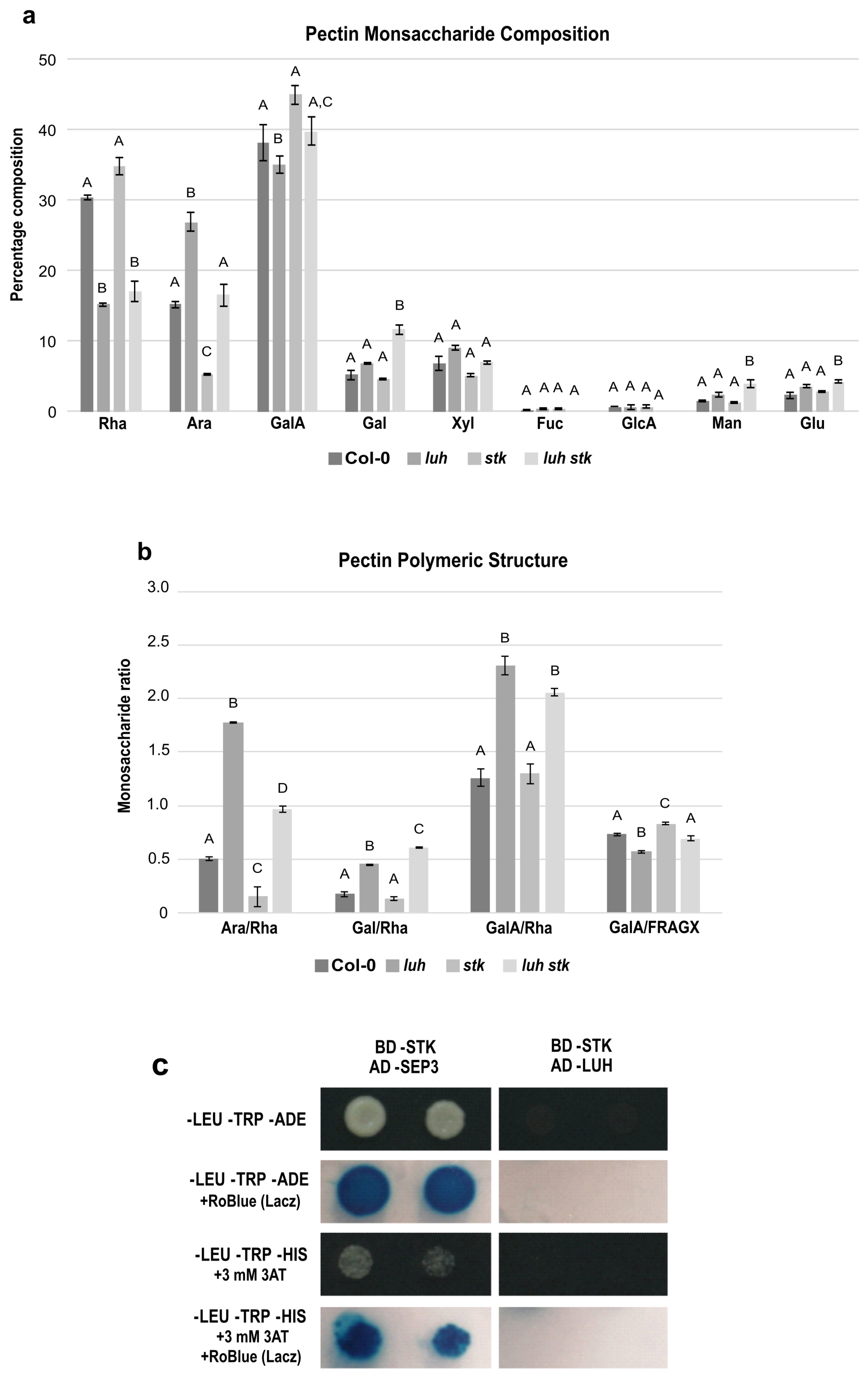
2.5. LUH and STK, Contrasting Control of Pectin Composition and Ramification
2.6. Structural Alterations of the Seed Coat Testa Cell Wall Influence Seed Germination
2.7. Proanthocyanidins (PAs) Accumulation Is Not Affected in CW Related Mutants
3. Discussion
3.1. Cell Wall Processes Modulate Seed and Fruit Growth
3.2. SEEDSTICK and LEUNIG-HOMOLOG Contribute to Seed Development
3.3. Cell Wall Processes Modulate Seed Germination
4. Materials and Methods
4.1. Plant Material
4.2. Seed and Silique Morphological Characterization
4.3. Germination Test
4.4. Yeast Two-Hybrid
4.5. Cell Wall Extraction and Pectin Enrichment Analyses from Mature Arabidopsis Seeds
4.6. Monosaccharide Composition Analysis
4.7. Proanthocyanidin and Mucilage Extrusion Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waterworth, W.M.; Bray, C.M.; West, C.E. The Importance of Safeguarding Genome Integrity in Germination and Seed Longevity. J. Exp. Bot. 2015, 66, 3549–3558. [Google Scholar] [CrossRef] [PubMed]
- Locascio, A.; Roig-Villanova, I.; Bernardi, J.; Varotto, S. Current Perspectives on the Hormonal Control of Seed Development in Arabidopsis and Maize: A Focus on Auxin. Front. Plant Sci. 2014, 5, 412. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Arroyo, G.; Paolo, D.; Ezquer, I.; Colombo, L. Networks Controlling Seed Size in Arabidopsis. Plant Reprod. 2015, 28, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, A.M.; Koltunow, A.; Payne, T.; Luo, M.; Tucker, M.R.; Dennis, E.S.; Peacock, W.J. Control of Early Seed Development. Annu. Rev. Cell Dev. Biol. 2001, 17, 677–699. [Google Scholar] [CrossRef] [PubMed]
- Sornay, E.; Dewitte, W.; Murray, J.A.H. Seed Size Plasticity in Response to Embryonic Lethality Conferred by Ectopic CYCD Activation Is Dependent on Plant Architecture. Plant Signal. Behav. 2016, 11, e1192741. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.B.; Huang, H.Y.; Hu, Y.W.; Zhu, S.W.; Wang, Z.Y.; Lin, W.H. Brassinosteroid Regulates Seed Size and Shape in Arabidopsis. Plant Physiol. 2013, 162, 1965–1977. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, M.F.; Kirkbride, R.C.; Stone, S.L.; Pelletier, J.M.; Bui, A.Q.; Yeung, E.C.; Hashimoto, M.; Fei, J.; Harada, C.M.; Munoz, M.D.; et al. Comprehensive Developmental Profiles of Gene Activity in Regions and Subregions of the Arabidopsis Seed. Proc. Natl. Acad. Sci. USA 2013, 110, E435–E444. [Google Scholar] [CrossRef]
- Dekkers, B.J.W.; Pearce, S.; van Bolderen-Veldkamp, R.P.; Marshall, A.; Widera, P.; Gilbert, J.; Drost, H.-G.; Bassel, G.W.; Müller, K.; King, J.R.; et al. Transcriptional Dynamics of Two Seed Compartments with Opposing Roles in Arabidopsis Seed Germination. Plant Physiol. 2013, 163, 205–215. [Google Scholar] [CrossRef]
- Aguirre, M.; Kiegle, E.; Leo, G.; Ezquer, I. Carbohydrate Reserves and Seed Development: An Overview. Plant Reprod. 2018, 31, 263–290. [Google Scholar] [CrossRef]
- Walker, M.; Tehseen, M.; Doblin, M.S.; Pettolino, F.A.; Wilson, S.M.; Bacic, A.; Golz, J.F. The Transcriptional Regulator LEUNIG_HOMOLOG Regulates Mucilage Release from the Arabidopsis Testa. Plant Physiol. 2011, 156, 46–60. [Google Scholar] [CrossRef]
- López-García, C.M.; Raya-González, J.; López-Bucio, J.S.; Guevara-García, Á.A.; López-Bucio, J. ALTERED MERISTEM PROGRAM 1 Plays a Role in Seed Coat Development, Root Growth, and Post-Embryonic Epidermal Cell Elongation in Arabidopsis. J. Plant Growth Regul. 2016, 35, 1141–1158. [Google Scholar] [CrossRef]
- Müller, K.; Tintelnot, S.; Leubner-Metzger, G. Endosperm-Limited Brassicaceae Seed Germination: Abscisic Acid Inhibits Embryo-Induced Endosperm Weakening of Lepidium sativum (Cress) and Endosperm Rupture of Cress and Arabidopsis thaliana. Plant Cell Physiol. 2006, 47, 864–877. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination-Still a Mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the Mark: Early Seed Germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed Germination and Vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef]
- Steinbrecher, T.; Leubner-Metzger, G. The Biomechanics of Seed Germination. J. Exp. Bot. 2017, 68, 765–783. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Growth of the Plant Cell Wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef]
- Manfield, I.W.; Orfila, C.; McCartney, L.; Harholt, J.; Bernal, A.J.; Scheller, H.V.; Gilmartin, P.M.; Mikkelsen, J.D.; Paul Knox, J.; Willats, W.G.T. Novel Cell Wall Architecture of Isoxaben-Habituated Arabidopsis Suspension-Cultured Cells: Global Transcript Profiling and Cellular Analysis. Plant J. 2004, 40, 260–275. [Google Scholar] [CrossRef]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- Driouich, A.; Follet-Gueye, M.-L.; Bernard, S.; Kousar, S.; Chevalier, L.; Vicré-Gibouin, M.; Lerouxel, O. Golgi-Mediated Synthesis and Secretion of Matrix Polysaccharides of the Primary Cell Wall of Higher Plants. Front. Plant Sci. 2012, 3, 79. [Google Scholar] [CrossRef]
- Polko, J.K.; Kieber, J.J. The Regulation of Cellulose Biosynthesis in Plants. Plant Cell 2019, 31, 282–296. [Google Scholar] [CrossRef] [PubMed]
- Wilkop, T.; Pattathil, S.; Ren, G.; Davis, D.J.; Bao, W.; Duan, D.; Peralta, A.G.; Domozych, D.S.; Hahn, M.G.; Drakakaki, G. A Hybrid Approach Enabling Large-Scale Glycomic Analysis of Post-Golgi Vesicles Reveals a Transport Route for Polysaccharides. Plant Cell 2019, 31, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, J.S.; North, H.M. Sticking to Cellulose: Exploiting Arabidopsis Seed Coat Mucilage to Understand Cellulose Biosynthesis and Cell Wall Polysaccharide Interactions. New Phytol. 2017, 214, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Willats, W.G.T.; McCartney, L.; Mackie, W.; Knox, J.P. Pectin: Cell Biology and Prospects for Functional Analysis. Plant Cell Walls 2001, 47, 9–27. [Google Scholar]
- Broxterman, S.E.; Schols, H.A. Interactions between Pectin and Cellulose in Primary Plant Cell Walls. Carbohydr. Polym. 2018, 192, 263–272. [Google Scholar] [CrossRef]
- Müller, K.; Levesque-Tremblay, G.; Bartels, S.; Weitbrecht, K.; Wormit, A.; Usadel, B.; Haughn, G.; Kermode, A.R. Demethylesterification of Cell Wall Pectins in Arabidopsis Plays a Role in Seed Germination. Plant Physiol. 2013, 161, 305–316. [Google Scholar] [CrossRef]
- Shigeyama, T.; Watanabe, A.; Tokuchi, K.; Toh, S.; Sakurai, N.; Shibuya, N.; Kawakami, N. α-Xylosidase Plays Essential Roles in Xyloglucan Remodelling, Maintenance of Cell Wall Integrity, and Seed Germination in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 5615–5629. [Google Scholar] [CrossRef]
- Di Marzo, M.; Viana, V.E.; Banfi, C.; Cassina, V.; Corti, R.; Herrera-Ubaldo, H.; Babolin, N.; Guazzotti, A.; Kiegle, E.; Gregis, V.; et al. Cell Wall Modifications by α-XYLOSIDASE1 Are Required for Control of Seed and Fruit Size in Arabidopsis. J. Exp. Bot. 2022, 73, 1499–1515. [Google Scholar] [CrossRef]
- Ezquer, I.; Mizzotti, C.; Nguema-Ona, E.; Gotté, M.; Beauzamy, L.; Viana, V.E.; Dubrulle, N.; Costa de Oliveira, A.; Caporali, E.; Koroney, A.-S.; et al. The Developmental Regulator SEEDSTICK Controls Structural and Mechanical Properties of the Arabidopsis Seed Coat. Plant Cell 2016, 28, 2478–2492. [Google Scholar] [CrossRef]
- Pinyopich, A.; Ditta, G.S.; Savidge, B.; Liljegren, S.J.; Baumann, E.; Wisman, E.; Yanofsky, M.F. Assessing the Redundancy of MADS-Box Genes during Carpel and Ovule Development. Nature 2003, 424, 85–88. [Google Scholar] [CrossRef]
- Saez-Aguayo, S.; Ralet, M.C.; Berger, A.; Botran, L.; Ropartz, D.; Marion-Poll, A.; North, H.M. PECTIN METHYLESTERASE INHIBITOR6 Promotes Arabidopsis Mucilage Release by Limiting Methylesterification of Homogalacturonan in Seed Coat Epidermal Cells. Plant Cell 2013, 25, 308–323. [Google Scholar] [CrossRef] [PubMed]
- Golz, J.F.; Allen, P.J.; Li, S.F.; Parish, R.W.; Jayawardana, N.U.; Bacic, A.; Doblin, M.S. Layers of Regulation–Insights into the Role of Transcription Factors Controlling Mucilage Production in the Arabidopsis Seed Coat. Plant Sci. 2018, 272, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Ren, A.; Tang, X.; Qi, G.; Xu, Z.; Chai, G.; Hu, R.; Zhou, G.; Kong, Y. MYB52 Negatively Regulates Pectin Demethylesterification in Seed Coat Mucilage. Plant Physiol. 2018, 176, 2737–2749. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.S.; Kolevski, B.; Smyth, D.R. TRANSPARENT TESTA GLABRA2, a Trichome and Seed Coat Development Gene of Arabidopsis, Encodes a WRKY Transcription Factor. Plant Cell 2002, 14, 1359–1375. [Google Scholar] [CrossRef]
- Huang, J.; DeBowles, D.; Esfandiari, E.; Dean, G.; Carpita, N.C.; Haughn, G.W. The Arabidopsis Transcription Factor LUH/MUM1 Is Required for Extrusion of Seed Coat Mucilage. Plant Physiol. 2011, 156, 491–502. [Google Scholar] [CrossRef]
- Wang, M.; Xu, Z.; Ahmed, R.I.; Wang, Y.; Hu, R.; Zhou, G.; Kong, Y. Tubby-like Protein 2 Regulates Homogalacturonan Biosynthesis in Arabidopsis Seed Coat Mucilage. Plant Mol. Biol. 2019, 99, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, M.; Di Marzo, M.; Guazzotti, A.; de Folter, S.; Kater, M.M.; Colombo, L. Gynoecium Size and Ovule Number Are Interconnected Traits That Impact Seed Yield. J. Exp. Bot. 2020, 71, 2479–2489. [Google Scholar] [CrossRef]
- North, H.M.; Berger, A.; Saez-Aguayo, S.; Ralet, M.-C.C. Understanding Polysaccharide Production and Properties Using Seed Coat Mutants: Future Perspectives for the Exploitation of Natural Variants. Ann. Bot. 2014, 114, 1251–1263. [Google Scholar] [CrossRef]
- Voiniciuc, C.; Dean, G.H.; Griffiths, J.S.; Kirchsteiger, K.; Hwang, Y.T.; Gillett, A.; Dow, G.; Western, T.L.; Estelle, M.; Haughn, G.W. FLYING SAUCER 1 Is a Transmembrane RING E3 Ubiquitin Ligase That Regulates the Degree of Pectin Methylesterification in Arabidopsis Seed Mucilage. Plant Cell 2013, 25, 944–959. [Google Scholar] [CrossRef]
- Arsovski, A.A.; Popma, T.M.; Haughn, G.W.; Carpita, N.C.; McCann, M.C.; Western, T.L. AtBXL1 Encodes a Bifunctional Beta-D-Xylosidase/Alpha-L-Arabinofuranosidase Required for Pectic Arabinan Modification in Arabidopsis Mucilage Secretory Cells. Plant Physiol. 2009, 150, 1219–1234. [Google Scholar] [CrossRef]
- Harpaz-Saad, S.; Western, T.L.; Kieber, J.J. The FEI2-SOS5 Pathway and CELLULOSE SYNTHASE 5 Are Required for Cellulose Biosynthesis in the Arabidopsis Seed Coat and Affect Pectin Mucilage Structure. Plant Signal. Behav. 2012, 7, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, J.S.; Crepeau, M.J.; Ralet, M.C.; Seifert, G.J.; North, H.M. Dissecting Seed Mucilage Adherence Mediated by FEI2 and SOS5. Front. Plant Sci. 2016, 7, 1073. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, J.S.; Tsai, A.Y.L.; Xue, H.; Voiniciuc, C.; Šola, K.; Seifert, G.J.; Mansfield, S.D.; Haughn, G.W. SALT-OVERLY SENSITIVE5 Mediates Arabidopsis Seed Coat Mucilage Adherence and Organization through Pectins. Plant Physiol. 2014, 165, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Francoz, E.; Ranocha, P.; Le Ru, A.; Martinez, Y.; Fourquaux, I.; Jauneau, A.; Dunand, C.; Burlat, V. Pectin Demethylesterification Generates Platforms That Anchor Peroxidases to Remodel Plant Cell Wall Domains. Dev. Cell 2019, 48, 261–276. [Google Scholar] [CrossRef]
- Sampedro, J.; Sieiro, C.; Revilla, G.; González-Villa, T.; Zarra, I. Cloning and Expression Pattern of a Gene Encoding an α-Xylosidase Active against Xyloglucan Oligosaccharides from Arabidopsis. Plant Physiol. 2001, 126, 910–920. [Google Scholar] [CrossRef]
- Louvet, R.; Cavel, E.; Gutierrez, L.; Guénin, S.; Roger, D.; Gillet, F.; Guerineau, F.; Pelloux, J. Comprehensive Expression Profiling of the Pectin Methylesterase Gene Family during Silique Development in Arabidopsis Thaliana. Planta 2006, 224, 782–791. [Google Scholar] [CrossRef]
- Ruan, Y.-L.; Patrick, J.W.; Bouzayen, M.; Osorio, S.; Fernie, A.R. Molecular Regulation of Seed and Fruit Set. Trends Plant Sci. 2012, 17, 656–665. [Google Scholar] [CrossRef]
- Balanzà, V.; Roig-Villanova, I.; Di Marzo, M.; Masiero, S.; Colombo, L. Seed Abscission and Fruit Dehiscence Required for Seed Dispersal Rely on Similar Genetic Networks. Development 2016, 143, 3372–3381. [Google Scholar] [CrossRef]
- Herrera-Ubaldo, H.; de Folter, S. Exploring Cell Wall Composition and Modifications During the Development of the Gynoecium Medial Domain in Arabidopsis. Front. Plant Sci. 2018, 9, 454. [Google Scholar] [CrossRef]
- Vivian-Smith, A.; Koltunow, A.M. Genetic Analysis of Growth-Regulator-Induced Parthenocarpy in Arabidopsis. Plant Physiol. 1999, 121, 437–451. [Google Scholar] [CrossRef]
- Roeder, A.H.; Yanofsky, M.F. Fruit development in Arabidopsis. Arabidopsis Book 2006, 4, e0075. [Google Scholar] [CrossRef] [PubMed]
- Ferrándiz, C. Regulation of Fruit Dehiscence in Arabidopsis. J. Exp. Bot. 2002, 53, 2031–2038. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Ubaldo, H.; de Folter, S. Gynoecium and Fruit Development in Arabidopsis. Development 2022, 149, dev200120. [Google Scholar] [CrossRef] [PubMed]
- Doughty, J.; Aljabri, M.; Scott, R.J. Flavonoids and the Regulation of Seed Size in Arabidopsis. Biochem. Soc. Trans. 2014, 42, 364–369. [Google Scholar] [CrossRef]
- Li, N.; Li, Y. Maternal Control of Seed Size in Plants. J. Exp. Bot. 2015, 66, 1087–1097. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Catalysts of Plant Cell Wall Loosening. F1000Research 2016, 5. [Google Scholar] [CrossRef]
- Harpaz-Saad, S.; McFarlane, H.E.; Xu, S.; Divi, U.K.; Forward, B.; Western, T.L.; Kieber, J.J. Cellulose Synthesis via the FEI2 RLK/SOS5 Pathway and Cellulose Synthase 5 Is Required for the Structure of Seed Coat Mucilage in Arabidopsis. Plant J. 2011, 68, 941–953. [Google Scholar] [CrossRef]
- Pauly, M.; Keegstra, K. Biosynthesis of the Plant Cell Wall Matrix Polysaccharide Xyloglucan. Annu. Rev. Plant Biol. 2016, 67, 235–259. [Google Scholar] [CrossRef]
- Di Marzo, M.; Herrera-Ubaldo, H.; Caporali, E.; Novák, O.; Strnad, M.; Balanzà, V.; Ezquer, I.; Mendes, M.A.; de Folter, S.; Colombo, L. SEEDSTICK Controls Arabidopsis Fruit Size by Regulating Cytokinin Levels and FRUITFULL. Cell Rep. 2020, 30, 2846–2857. [Google Scholar] [CrossRef]
- Paolo, D.; Rotasperti, L.; Schnittger, A.; Masiero, S.; Colombo, L.; Mizzotti, C. The Arabidopsis Mads-Domain Transcription Factor Seedstick Controls Seed Size via Direct Activation of E2fa. Plants 2021, 10, 192. [Google Scholar] [CrossRef]
- Bui, M.; Lim, N.; Sijacic, P.; Liu, Z. LEUNIG_HOMOLOG and LEUNIG Regulate Seed Mucilage Extrusion in Arabidopsis. J. Integr. Plant Biol. 2011, 53, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Houben, K.; Jolie, R.P.; Fraeye, I.; Van Loey, A.M.; Hendrickx, M.E. Comparative Study of the Cell Wall Composition of Broccoli, Carrot, and Tomato: Structural Characterization of the Extractable Pectins and Hemicelluloses. Carbohydr. Res. 2011, 346, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Daszkowska-Golec, A. Arabidopsis Seed Germination under Abiotic Stress as a Concert of Action of Phytohormones. OMICS 2011, 15, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Longo, C.; Holness, S.; De Angelis, V.; Lepri, A.; Occhigrossi, S.; Ruta, V.; Vittorioso, P. From the Outside to the Inside: New Insights on the Main Factors That Guide Seed Dormancy and Germination. Genes 2020, 12, 52. [Google Scholar] [CrossRef] [PubMed]
- Saha, D.; Choyal, P.; Mishra, U.N.; Dey, P.; Bose, B.; MD, P.; Gupta, N.K.; Mehta, B.K.; Kumar, P.; Pandey, S.; et al. Drought Stress Responses and Inducing Tolerance by Seed Priming Approach in Plants. Plant Stress 2022, 4, 100066. [Google Scholar] [CrossRef]
- Western, T.L. The Sticky Tale of Seed Coat Mucilages: Production, Genetics, and Role in Seed Germination and Dispersal. Seed Sci. Res. 2012, 22, 1–25. [Google Scholar] [CrossRef]
- Macquet, A.; Ralet, M.-C.; Loudet, O.; Kronenberger, J.; Mouille, G.; Marion-Poll, A.; North, H.M. A Naturally Occurring Mutation in an Arabidopsis Accession Affects a β-d-Galactosidase That Increases the Hydrophilic Potential of Rhamnogalacturonan I in Seed Mucilage. Plant Cell 2007, 19, 3990–4006. [Google Scholar] [CrossRef]
- Macquet, A.; Ralet, M.-C.; Kronenberger, J.; Marion-Poll, A.; North, H.M. In Situ, Chemical and Macromolecular Study of the Composition of Arabidopsis Thaliana Seed Coat Mucilage. Plant Cell Physiol. 2007, 48, 984–999. [Google Scholar] [CrossRef]
- Poulain, D.; Botran, L.; North, H.M.; Ralet, M.-C. Composition and Physicochemical Properties of Outer Mucilage from Seeds of Arabidopsis Natural Accessions. AoB Plants 2019, 11, plz031. [Google Scholar] [CrossRef]
- Voiniciuc, C.; Günl, M.; Schmidt, M.H.W.; Usadel, B. Highly Branched Xylanmade by IRREGULAR XYLEM14 and MUCILAGE-RELATED21 Links Mucilage to Arabidopsis Seeds. Plant Physiol. 2015, 169, 2481–2495. [Google Scholar] [CrossRef]
- Kunieda, T.; Shimada, T.; Kondo, M.; Nishimura, M.; Nishitani, K.; Hara-Nishimura, I. Spatiotemporal Secretion of PEROXIDASE36 Is Required for Seed Coat Mucilage Extrusion in Arabidopsis. Plant Cell 2013, 25, 1355–1367. [Google Scholar] [CrossRef] [PubMed]
- Kunieda, T.; Hara-Nishimura, I.; Demura, T.; Haughn, G.W. Arabidopsis FLYING SAUCER 2 Functions Redundantly with FLY1 to Establish Normal Seed Coat Mucilage. Plant Cell Physiol. 2020, 61, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Western, T.L.; Burn, J.; Tan, W.L.; Skinner, D.J.; Martin-McCaffrey, L.; Moffatt, B.A.; Haughn, G.W. Isolation and Characterization of Mutants Defective in Seed Coat Mucilage Secretory Cell Development in Arabidopsis. Plant Physiol. 2001, 127, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Haughn, G.; Chaudhury, A. Genetic Analysis of Seed Coat Development in Arabidopsis. Trends Plant Sci. 2005, 10, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Debeaujon, I.; Léon-Kloosterziel, K.M.; Koornneef, M. Influence of the Testa on Seed Dormancy, Germination, and Longevity in Arabidopsis. Plant Physiol. 2000, 122, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Mizzotti, C.; Ezquer, I.; Paolo, D.; Rueda-Romero, P.; Guerra, R.F.; Battaglia, R.; Rogachev, I.; Aharoni, A.; Kater, M.M.; Caporali, E.; et al. SEEDSTICK Is a Master Regulator of Development and Metabolism in the Arabidopsis Seed Coat. PLoS Genet. 2014, 10, e1004856. [Google Scholar] [CrossRef]
- Paolo, D.; Orozco-arroyo, G.; Rotasperti, L.; Masiero, S.; Colombo, L.; de Folter, S.; Ambrose, B.A.; Caporali, E.; Ezquer, I.; Mizzotti, C. Genetic Interaction of SEEDSTICK, GORDITA and AUXIN RESPONSE FACTOR 2 during Seed Development. Genes 2021, 12, 1189. [Google Scholar] [CrossRef]
- Creff, A.; Brocard, L.; Ingram, G. A Mechanically Sensitive Cell Layer Regulates the Physical Properties of the Arabidopsis Seed Coat. Nat. Commun. 2015, 6, 6382. [Google Scholar] [CrossRef]
- Levesque-Tremblay, G.; Pelloux, J.; Braybrook, S.A.; Müller, K. Tuning of Pectin Methylesterification: Consequences for Cell Wall Biomechanics and Development. Planta 2015, 242, 791–811. [Google Scholar] [CrossRef]
- Bashline, L.; Lei, L.; Li, S.; Gu, Y. Cell Wall, Cytoskeleton, and Cell Expansion in Higher Plants. Mol. Plant 2014, 7, 586–600. [Google Scholar] [CrossRef]
- Goujon, T.; Minic, Z.; El Amrani, A.; Lerouxel, O.; Aletti, E.; Lapierre, C.; Joseleau, J.-P.; Jouanin, L. AtBXL1, a Novel Higher Plant (Arabidopsis Thaliana) Putative Beta-Xylosidase Gene, Is Involved in Secondary Cell Wall Metabolism and Plant Development. Plant J. 2003, 33, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Re-Constructing Our Models of Cellulose and Primary Cell Wall Assembly. Curr. Opin. Plant Biol. 2014, 22, 122–131. [Google Scholar] [CrossRef]
- Peaucelle, A.; Braybrook, S.; Höfte, H. Cell Wall Mechanics and Growth Control in Plants: The Role of Pectins Revisited. Front. Plant Sci. 2012, 3, 121. [Google Scholar] [CrossRef]
- Bidhendi, A.J.; Geitmann, A. Relating the Mechanics of the Primary Plant Cell Wall to Morphogenesis. J. Exp. Bot. 2016, 67, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Liners, F.; Letesson, J.-J.; Didembourg, C.; Cutsem, P. Van Monoclonal Antibodies against Pectin: Recognition of a Conformation Induced by Calcium. Plant Physiol. 1989, 91, 1419–1424. [Google Scholar] [CrossRef]
- Palin, R.; Geitmann, A. The Role of Pectin in Plant Morphogenesis. Biosystems 2012, 109, 397–402. [Google Scholar] [CrossRef]
- Ralet, M.-C.; Crépeau, M.-J.; Vigouroux, J.; Tran, J.; Berger, A.; Sallé, C.; Granier, F.; Botran, L.; North, H.M. Xylans Provide the Structural Driving Force for Mucilage Adhesion to the Arabidopsis Seed Coat. Plant Physiol. 2016, 171, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hong, M. Solid-State NMR Investigations of Cellulose Structure and Interactions with Matrix Polysaccharides in Plant Primary Cell Walls. J. Exp. Bot. 2016, 67, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Gibeaut, D.M.; Bacic, A.; Findlay, K.; Roberts, K.; Hamilton, A.; Baulcombe, D.C.; Fincher, G.B. Virus-Induced Silencing of a Plant Cellulose Synthase Gene. Plant Cell 2000, 12, 691–706. [Google Scholar] [CrossRef]
- Mendu, V.; Griffiths, J.S.; Persson, S.; Stork, J.; Downie, A.B.; Voiniciuc, C.; Haughn, G.W.; DeBolt, S. Subfunctionalization of Cellulose Synthases in Seed Coat Epidermal Cells Mediates Secondary Radial Wall Synthesis and Mucilage Attachment. Plant Physiol. 2011, 157, 441–453. [Google Scholar] [CrossRef]
- Desprez, T.; Juraniec, M.; Crowell, E.F.; Jouy, H.; Pochylova, Z.; Parcy, F.; Höfte, H.; Gonneau, M.; Vernhettes, S. Organization of Cellulose Synthase Complexes Involved in Primary Cell Wall Synthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2007, 104, 15572–15577. [Google Scholar] [CrossRef] [PubMed]
- Persson, S.; Paredez, A.; Carroll, A.; Palsdottir, H.; Doblin, M.; Poindexter, P.; Khitrov, N.; Auer, M.; Somerville, C.R. Genetic Evidence for Three Unique Components in Primary Cell-Wall Cellulose Synthase Complexes in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 15566–15571. [Google Scholar] [CrossRef] [PubMed]
- Stork, J.; Harris, D.; Griffiths, J.; Williams, B.; Beisson, F.; Li-Beisson, Y.; Mendu, V.; Haughn, G.; Debolt, S. CELLULOSE SYNTHASE9 Serves a Nonredundant Role in Secondary Cell Wall Synthesis in Arabidopsis Epidermal Testa Cells. Plant Physiol. 2010, 153, 580–589. [Google Scholar] [CrossRef]
- Yang, B.; Hofmann, F.; Usadel, B.; Voiniciuc, C. Seed Hemicelluloses Tailor Mucilage Properties and Salt Tolerance. New Phytol. 2021, 229, 1946–1954. [Google Scholar] [CrossRef] [PubMed]
- Siedlecka, A.; Wiklund, S.; Péronne, M.-A.; Micheli, F.; Lesniewska, J.; Sethson, I.; Edlund, U.; Richard, L.; Sundberg, B.; Mellerowicz, E.J. Pectin Methyl Esterase Inhibits Intrusive and Symplastic Cell Growth in Developing Wood Cells of Populus. Plant Physiol. 2008, 146, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Windsor, J.B.; Symonds, V.V.; Mendenhall, J.; Lloyd, A.M. Arabidopsis Seed Coat Development: Morphological Differentiation of the Outer Integument. Plant J. 2000, 22, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Stadler, R.; Lauterbach, C.; Sauer, N. Cell-to-Cell Movement of Green Fluorescent Protein Reveals Post-Phloem Transport in the Outer Integument and Identifies Symplastic Domains in Arabidopsis Seeds and Embryos. Plant Physiol. 2005, 139, 701–712. [Google Scholar] [CrossRef]
- Shi, L.; Katavic, V.; Yu, Y.; Kunst, L.; Haughn, G. Arabidopsis glabra2 Mutant Seeds Deficient in Mucilage Biosynthesis Produce More Oil. Plant J. 2011, 69, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Western, T.L.; Young, D.S.; Dean, G.H.; Tan, W.L.; Samuels, A.L.; Haughn, G.W. MUCILAGE-MODIFIED4 Encodes a Putative Pectin Biosynthetic Enzyme Developmentally Regulated by APETALA2, TRANSPARENT TESTA GLABRA1, and GLABRA2 in the Arabidopsis Seed Coat. Plant Physiol. 2004, 134, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Francoz, E.; Ranocha, P.; Nguyen-Kim, H.; Jamet, E.; Burlat, V.; Dunand, C. Roles of Cell Wall Peroxidases in Plant Development. Phytochemistry 2015, 112, 15–21. [Google Scholar] [CrossRef]
- Sitaraman, J.; Bui, M.; Liu, Z. LEUNIG_HOMOLOG and LEUNIG Perform Partially Redundant Functions during Arabidopsis Embryo and Floral Development. Plant Physiol. 2008, 147, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Park, J.; Kim, K.; Choi, G. The Transcriptional Coregulator LEUNIG_HOMOLOG Inhibits Light-Dependent Seed Germination in Arabidopsis. Plant Cell 2015, 27, 2301–2313. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wu, L.; Li, L.; Zhong, M.; Tang, Y.; Cao, G.; Lin, K.; Ye, Y. Aluminum-Induced Alterations to the Cell Wall and Antioxidant Enzymes Involved in the Regulation of the Aluminum Tolerance of Chinese Fir (Cunninghamia lanceolata). Front. Plant Sci. 2022, 13, 891117. [Google Scholar] [CrossRef] [PubMed]
- Zwieniecki, M.A.; Melcher, P.J.; Holbrook, N.M. Hydrogel Control of Xylem Hydraulic Resistance in Plants. Science 2001, 291, 1059–1062. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Tanabata, T.; Shibaya, T.; Hori, K.; Ebana, K.; Yano, M. SmartGrain: High-Throughput Phenotyping Software for Measuring Seed Shape through Image Analysis. Plant Physiol. 2012, 160, 1871–1880. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Millar, A.A.; Jacobsen, J.V.; Ross, J.J.; Helliwell, C.A.; Poole, A.T.; Scofield, G.; Reid, J.B.; Gubler, F. Seed Dormancy and ABA Metabolism in Arabidopsis and Barley: The Role of ABA 8′-Hydroxylase. Plant J. 2006, 45, 942–954. [Google Scholar] [CrossRef]
- De Folter, S.; Immink, R.G.H.; Kieffer, M.; Parenicová, L.; Henz, S.R.; Weigel, D.; Busscher, M.; Kooiker, M.; Colombo, L.; Kater, M.M.; et al. Comprehensive Interaction Map of the Arabidopsis MADS Box Transcription Factors. Plant Cell 2005, 17, 1424–1433. [Google Scholar] [CrossRef]
- De Folter, S.; Immink, R.G.H. Yeast Protein–Protein Interaction Assays and Screens. Methods Mol. Biol. 2011, 754, 145–165. [Google Scholar]
- Paz-Ares, J.; Consortium, R. REGIA, an EU Project on Functional Genomics of Transcription Factors from Arabidopsis thaliana. Comp. Funct. Genom. 2002, 3, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.P.; Nguema-Ona, E.; Chevalier, L.; Lindsey, G.G.; Brandt, W.F.; Lerouge, P.; Farrant, J.M.; Driouich, A. Response of the Leaf Cell Wall to Desiccation in the Resurrection Plant Myrothamnus flabellifolius. Plant Physiol. 2006, 141, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Ponce, N.M.A.; Ziegler, V.H.; Stortz, C.A.; Sozzi, G.O. Compositional Changes in Cell Wall Polysaccharides from Japanese Plum (Prunus salicina Lindl.) during Growth and On-Tree Ripening. J. Agric. Food Chem. 2010, 58, 2562–2570. [Google Scholar] [CrossRef]
- Renard, C.M.G.C.; Ginies, C. Comparison of the Cell Wall Composition for Flesh and Skin from Five Different Plums. Food Chem. 2009, 114, 1042–1049. [Google Scholar] [CrossRef]
- Popov, S.; Smirnov, V.; Kvashninova, E.; Khlopin, V.; Vityazev, F.; Golovchenko, V. Isolation, Chemical Characterization and Antioxidant Activity of Pectic Polysaccharides of Fireweed (Epilobium angustifolium L.). Molecules 2021, 26, 7290. [Google Scholar] [CrossRef] [PubMed]
- Jofuku, K.D.; Omidyar, P.K.; Gee, Z.; Okamuro, J.K. Control of Seed Mass and Seed Yield by the Floral Homeotic Gene APETALA2. Proc. Natl. Acad. Sci. USA 2005, 102, 3117–3122. [Google Scholar] [CrossRef] [PubMed]
- Gubler, F.; Millar, A.A.; Jacobsen, J. V Dormancy Release, ABA and Pre-Harvest Sprouting. Curr. Opin. Plant Biol. 2005, 8, 183–187. [Google Scholar] [CrossRef]
- Navarro, D.A.; Cerezo, A.S.; Stortz, C.A. NMR Spectroscopy and Chemical Studies of an Arabinan-Rich System from the Endosperm of the Seed of Gleditsia triacanthos. Carbohydr. Res. 2002, 337, 255–263. [Google Scholar] [CrossRef]
- Mescia, T.B.; Louro, R.P.; Barbedo, C.J.; Carbonero, E.R.; Figueiredo-Ribeiro, R.D.C.L.; Braga, M.R. Changes in Cell Wall Composition and Ultrastructure Related to Desiccation during the Seed Maturation of Paubrasilia echinata (Brazilwood). Protoplasma 2022, 259, 1255–1269. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Germination Test | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Type | Genotype | Freshly Harvested Seeds | Vernalized Seeds | ||||||||||
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | ||||||||
| % | SD | % | SD | % | SD | % | SD | % | SD | % | SD | ||
| Wild-type | Col-0 | 12.7 | 2.5 | 75.3 | 2.5 | 99.3 | 1.1 | 45.0 | 1.7 | 99.0 | 1.7 | 100 | 0 |
| Ws | 19.3 | 3.2 | 50.3 | 2.5 | 70.7 | 1.5 | 39.7 | 1.5 | 59.7 | 4.2 | 88.7 | 2.5 | |
| Transcriptional regulators | stk | 23.7 *,+ | 0.6 | 54.7 *,+ | 1.5 | 90.7 * | 2.1 | 41.0 + | 3.0 | 76.7 *,+ | 1.5 | 100 | 0 |
| luh | 80.0 * | 2.0 | 96.3 *,+ | 3.8 | 100 + | 0 | 60.0 *,+ | 1.7 | 98.0 + | 1.0 | 100 | 0 | |
| luh stk | 84.0 * | 5.3 | 88.3 * | 0.6 | 92.0 | 4.6 | 83.7 * | 5.8 | 99.3 | 1.1 | 99.7 | 0.6 | |
| gl2.8 | 6.0 * | 2.0 | 52.0 * | 1.0 | 92.0 | 2.0 | 29.7 * | 1.5 | 53.0 * | 2.0 | 75.7 * | 1.5 | |
| Cellulose biosynthesis | cesa5 | 8.7 | 1.1 | 73.3 | 3.5 | 100 | 0 | 31.3 * | 1.1 | 89.3 * | 1.1 | 97.7 | 2.5 |
| fei2 | 21.7 * | 2.3 | 95.0 * | 1.0 | 98.0 | 2.0 | 72.0 * | 5.0 | 99.3 | 0.6 | 99.7 | 0.6 | |
| Pectin biosynthesis and maturation | pmei6 | 0 * | 0 | 75.3 | 1.1 | 81.3 * | 6.1 | 56.0 * | 1.0 | 90.0 * | 2.0 | 97.3 | 3.0 |
| fly1.1 | 31.7 * | 1.5 | 91.7 * | 1.5 | 99.0 | 1.7 | 90.3 * | 1.5 | 99.3 | 1.1 | 100 | 0 | |
| fly1.3 | 9.3 | 4.2 | 72.3 | 6.5 | 76.0 * | 7.5 | 92.0 * | 2.0 | 98.0 | 2.0 | 100 | 0 | |
| Mucilage extrusion | per36 | 0 * | 0 | 18.3 * | 2.1 | 57.0 * | 3.0 | 68.0 * | 1.7 | 86.7 * | 6.1 | 93.3 * | 2.1 |
| Hemicellulose | bxl1 | 0 * | 0 | 16.7 * | 2.1 | 28.0 * | 2.0 | 0 * | 0 | 100 * | 0 | 100* | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Marzo, M.; Babolin, N.; Viana, V.E.; de Oliveira, A.C.; Gugi, B.; Caporali, E.; Herrera-Ubaldo, H.; Martínez-Estrada, E.; Driouich, A.; de Folter, S.; et al. The Genetic Control of SEEDSTICK and LEUNIG-HOMOLOG in Seed and Fruit Development: New Insights into Cell Wall Control. Plants 2022, 11, 3146. https://doi.org/10.3390/plants11223146
Di Marzo M, Babolin N, Viana VE, de Oliveira AC, Gugi B, Caporali E, Herrera-Ubaldo H, Martínez-Estrada E, Driouich A, de Folter S, et al. The Genetic Control of SEEDSTICK and LEUNIG-HOMOLOG in Seed and Fruit Development: New Insights into Cell Wall Control. Plants. 2022; 11(22):3146. https://doi.org/10.3390/plants11223146
Chicago/Turabian StyleDi Marzo, Maurizio, Nicola Babolin, Vívian Ebeling Viana, Antonio Costa de Oliveira, Bruno Gugi, Elisabetta Caporali, Humberto Herrera-Ubaldo, Eduardo Martínez-Estrada, Azeddine Driouich, Stefan de Folter, and et al. 2022. "The Genetic Control of SEEDSTICK and LEUNIG-HOMOLOG in Seed and Fruit Development: New Insights into Cell Wall Control" Plants 11, no. 22: 3146. https://doi.org/10.3390/plants11223146
APA StyleDi Marzo, M., Babolin, N., Viana, V. E., de Oliveira, A. C., Gugi, B., Caporali, E., Herrera-Ubaldo, H., Martínez-Estrada, E., Driouich, A., de Folter, S., Colombo, L., & Ezquer, I. (2022). The Genetic Control of SEEDSTICK and LEUNIG-HOMOLOG in Seed and Fruit Development: New Insights into Cell Wall Control. Plants, 11(22), 3146. https://doi.org/10.3390/plants11223146