Exploration of the Possible Relationships Between Gut and Hypothalamic Inflammation and Allopregnanolone: Preclinical Findings in a Post-Finasteride Rat Model

 , , ,
, , ,  , , ,
, , ,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Treatment and Experimental Design
2.3. Thiobarbituric Acid Reactive Substance
2.4. RNA Extraction and Gene Expression Analysis
2.5. Histological Analysis of Colon Tissues
2.6. SCFA Quantification and Analysis
2.7. Western Blotting Analysis
2.8. Metabolomic Profiling and Analysis
2.9. Statistics
3. Results
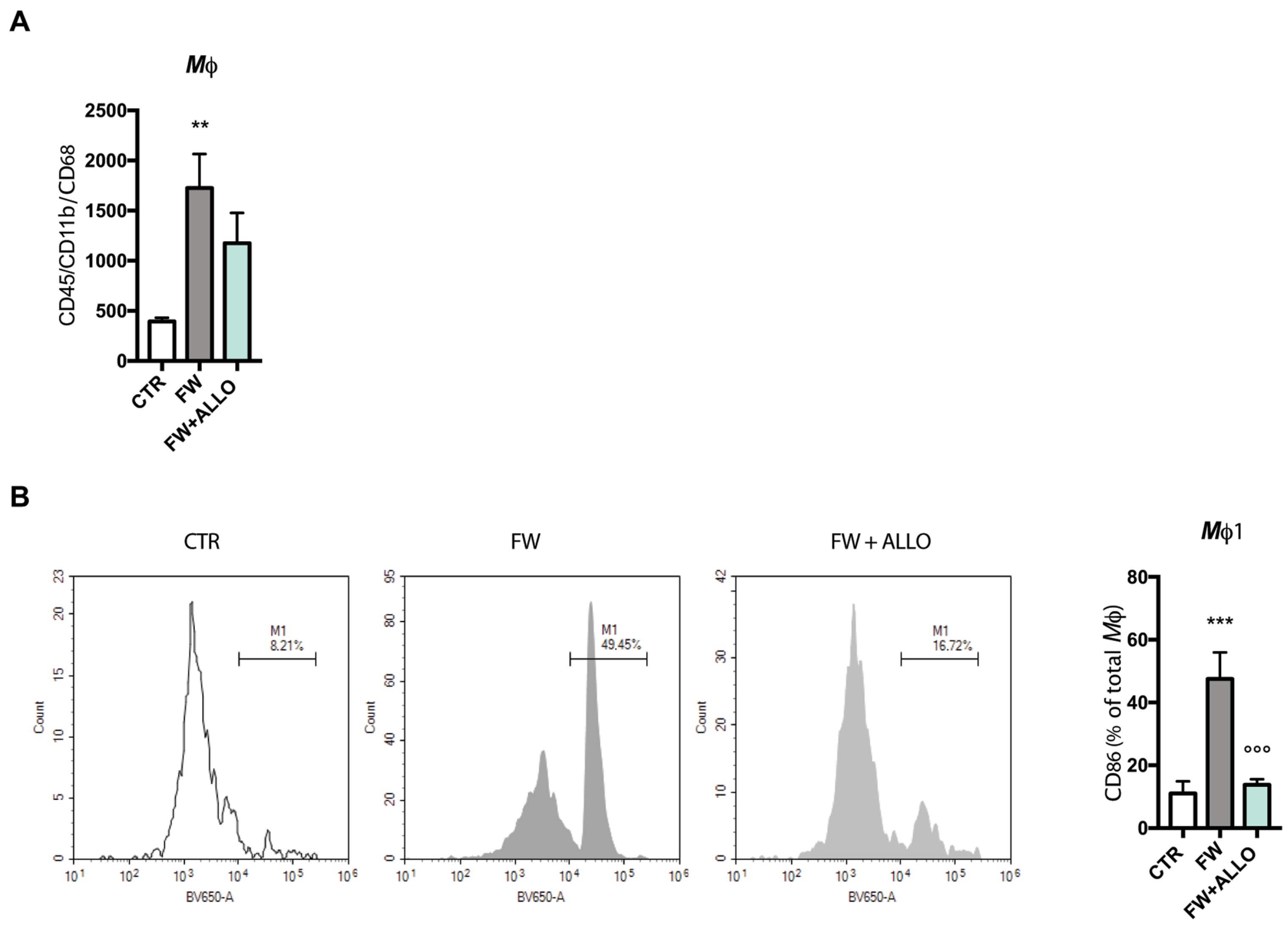
3.1. Allopregnanolone Treatment Reverses Finasteride Withdrawal-Induced Macrophage (CD86+) Polarization
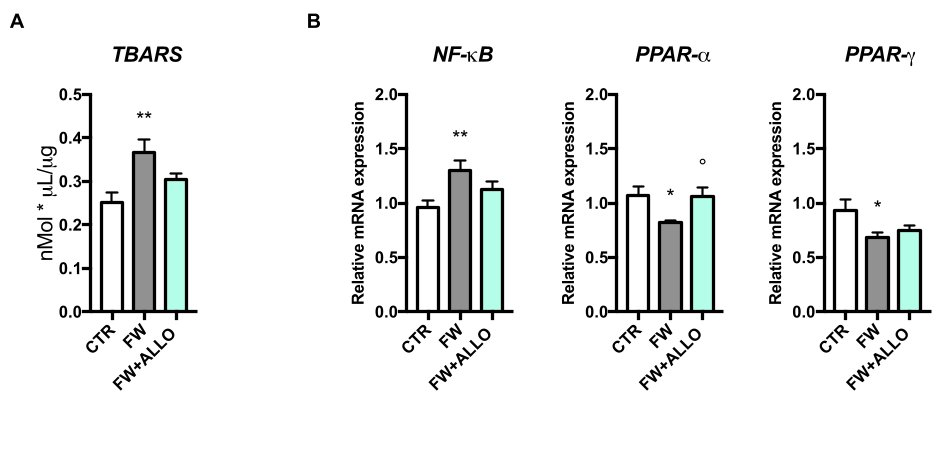
3.2. Finasteride Withdrawal Increases Oxidative Stress and Decreases PPAR Gene Expression in the Colon—Activation of Protective Mechanisms by Allopregnanolone
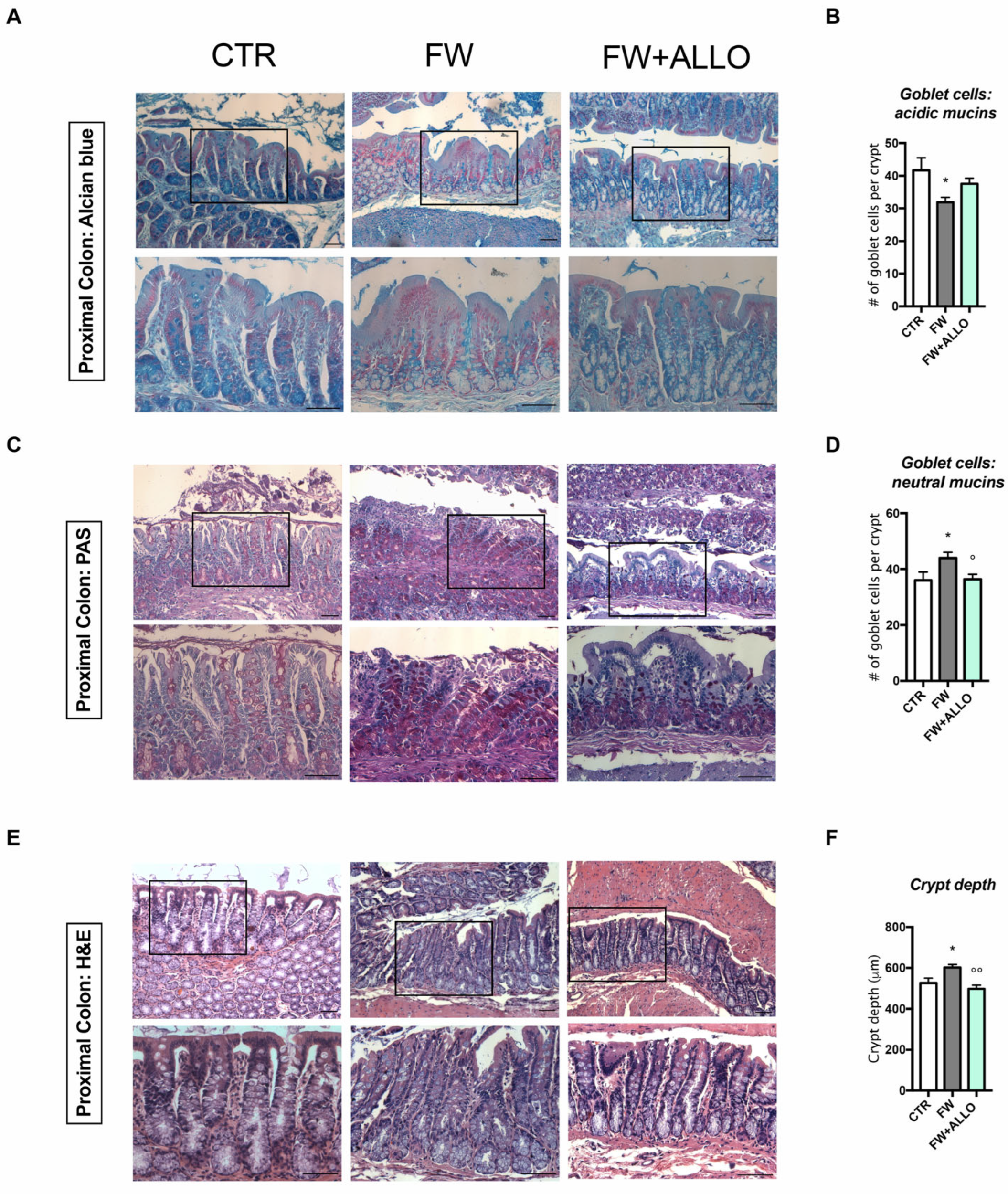
3.3. Finasteride Withdrawal Alters the Intestinal Epithelium Morphology Affecting Mucin Composition and Crypt Depth—Protective Effects of Allopregnanolone
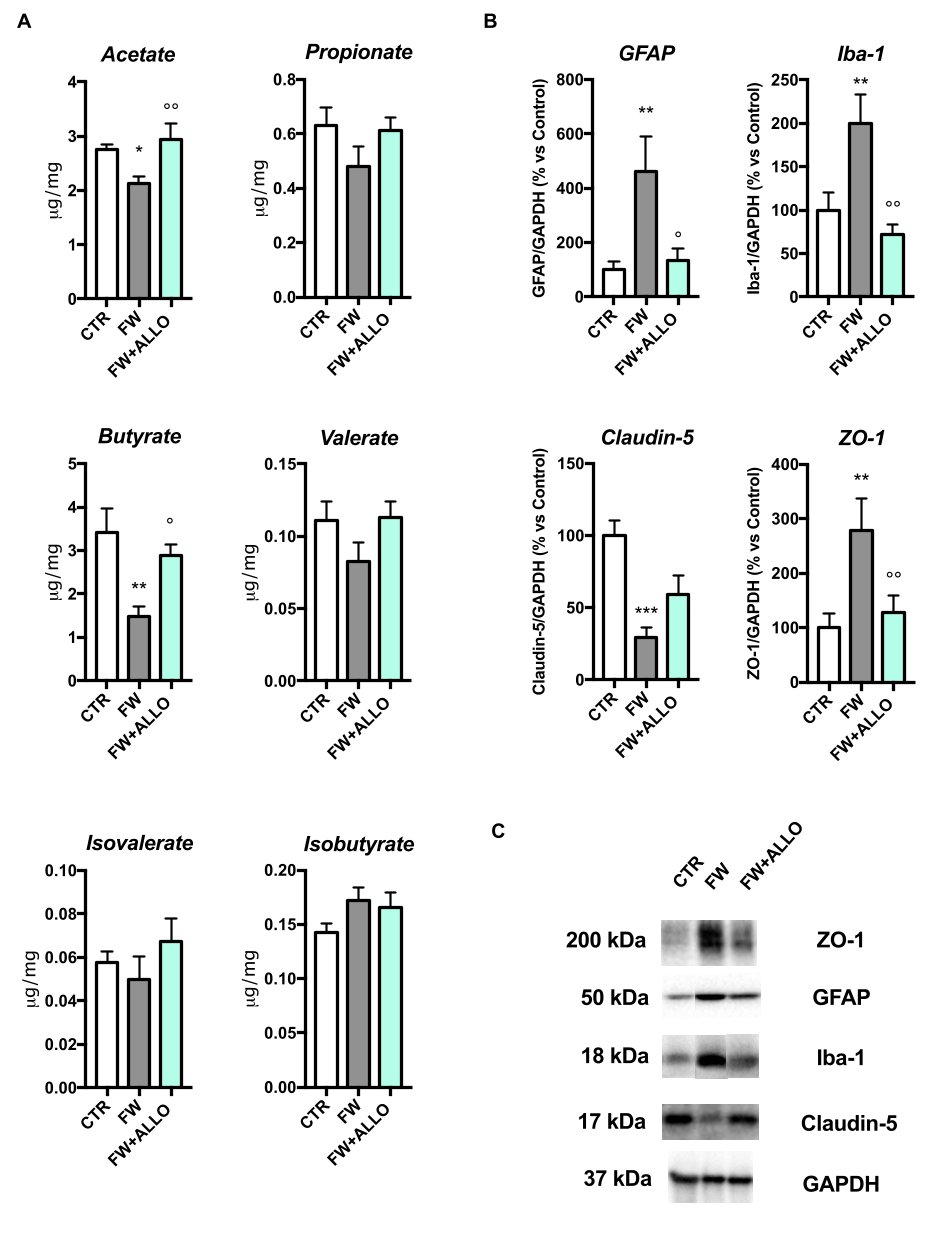
3.4. Finasteride Withdrawal Decreases Acetate and Butyrate Fecal Levels and Induces Hypothalamic Neuroinflammation—Protective Effects of Allopregnanolone Treatment
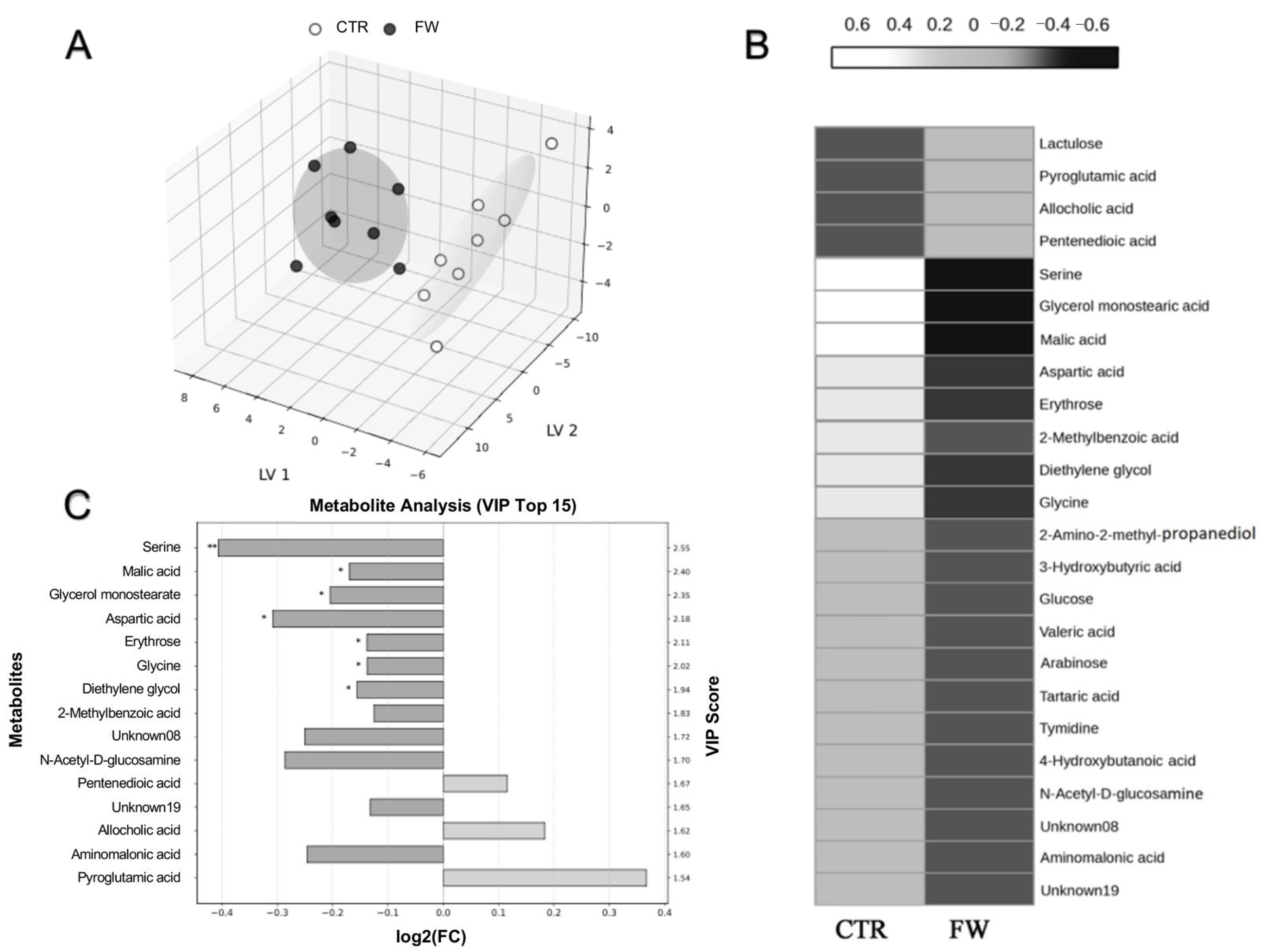
3.5. Metabolic Shifts in the Cecum Induced by Finasteride Withdrawal
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALLO | Allopregnanolone |
| BBB | Blood–brain barrier |
| CNS | Central nervous system |
| FITC-dextran | Fluorescein isothiocyanate–dextran |
| FW | Finasteride withdrawal |
| GABA | Gamma-aminobutyric acid |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GC-MS | Gas chromatography mass spectrometry |
| GFAP | Glial fibrillary acidic protein |
| HPA | Hypothalamic–pituitary–adrenal |
| Iba-1 | Ionized calcium-binding adaptor molecule-1 |
| IBD | Inflammatory bowel disease |
| Mφ | Macrophages |
| MDA | Malondialdeide |
| MDD | Major depressive disorder |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| PDD | Premenstrual dysphoric disorder |
| PFS | Post-finasteride syndrome |
| PPARs | Peroxisome proliferator-activated receptors |
| PPD | Postpartum depression |
| PTSD | Post-traumatic stress disorder |
| SCFAs | Short-chain fatty acids |
| TBARS | Thiobarbituric acid reactive substance |
| TJs | Tight junctions |
| ZO-1 | Zonula occludens-1 |
References
- Traish, A.M.; Haider, K.S.; Doros, G.; Haider, A. Finasteride, not tamsulosin, increases severity of erectile dysfunction and decreases testosterone levels in men with benign prostatic hyperplasia. Horm. Mol. Biol. Clin. Investig. 2015, 23, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Diviccaro, S.; Melcangi, R.C.; Giatti, S. Post-finasteride syndrome: An emerging clinical problem. Neurobiol. Stress. 2020, 12, 100209. [Google Scholar] [CrossRef] [PubMed]
- Cilio, S.; Tsampoukas, G.; Morgado, A.; Ramos, P.; Minhas, S. Post-finasteride syndrome-a true clinical entity? Int. J. Impot. Res. 2025, 37, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Traish, A.M.; Melcangi, R.C.; Bortolato, M.; Garcia-Segura, L.M.; Zitzmann, M. Adverse effects of 5alpha-reductase inhibitors: What do we know, don’t know, and need to know? Rev. Endocr. Metab. Disord. 2015, 16, 177–198. [Google Scholar] [CrossRef]
- Basaria, S.; Jasuja, R.; Huang, G.; Wharton, W.; Pan, H.; Pencina, K.; Li, Z.; Travison, T.G.; Bhawan, J.; Gonthier, R.; et al. Characteristics of Men Who Report Persistent Sexual Symptoms After Finasteride Use for Hair Loss. J. Clin. Endocrinol. Metab. 2016, 101, 4669–4680. [Google Scholar] [CrossRef]
- Khera, M.; Than, J.K.; Anaissie, J.; Antar, A.; Song, W.; Losso, B.; Pastuszak, A.; Kohn, T.; Mirabal, J.R. Penile vascular abnormalities in young men with persistent side effects after finasteride use for the treatment of androgenic alopecia. Transl. Androl. Urol. 2020, 9, 1201–1209. [Google Scholar] [CrossRef]
- Healy, D.; Bahrick, A.; Bak, M.; Barbato, A.; Calabro, R.S.; Chubak, B.M.; Cosci, F.; Csoka, A.B.; D’Avanzo, B.; Diviccaro, S.; et al. Diagnostic criteria for enduring sexual dysfunction after treatment with antidepressants, finasteride and isotretinoin. Int. J. Risk Saf. Med. 2022, 33, 65–76. [Google Scholar] [CrossRef]
- Melcangi, R.C.; Santi, D.; Spezzano, R.; Grimoldi, M.; Tabacchi, T.; Fusco, M.L.; Diviccaro, S.; Giatti, S.; Carra, G.; Caruso, D.; et al. Neuroactive steroid levels and psychiatric and andrological features in post-finasteride patients. J. Steroid. Biochem. Mol. Biol. 2017, 171, 229–235. [Google Scholar] [CrossRef]
- Giatti, S.; Foglio, B.; Romano, S.; Pesaresi, M.; Panzica, G.; Garcia-Segura, L.M.; Caruso, D.; Melcangi, R.C. Effects of Subchronic Finasteride Treatment and Withdrawal on Neuroactive Steroid Levels and Their Receptors in the Male Rat Brain. Neuroendocrinology 2016, 103, 746–757. [Google Scholar] [CrossRef]
- Diviccaro, S.; Herian, M.; Cioffi, L.; Audano, M.; Mitro, N.; Caruso, D.; Giatti, S.; Melcangi, R.C. Exploring rat corpus cavernosum alterations induced by finasteride treatment and withdrawal. Andrology 2024, 12, 674–681. [Google Scholar] [CrossRef]
- Diviccaro, S.; Giatti, S.; Cioffi, L.; Falvo, E.; Herian, M.; Caruso, D.; Melcangi, R.C. Gut Inflammation Induced by Finasteride Withdrawal: Therapeutic Effect of Allopregnanolone in Adult Male Rats. Biomolecules 2022, 12, 1567. [Google Scholar] [CrossRef] [PubMed]
- Diviccaro, S.; Giatti, S.; Borgo, F.; Barcella, M.; Borghi, E.; Trejo, J.L.; Garcia-Segura, L.M.; Melcangi, R.C. Treatment of male rats with finasteride, an inhibitor of 5alpha-reductase enzyme, induces long-lasting effects on depressive-like behavior, hippocampal neurogenesis, neuroinflammation and gut microbiota composition. Psychoneuroendocrinology 2019, 99, 206–215. [Google Scholar] [CrossRef]
- Borgo, F.; Macandog, A.D.; Diviccaro, S.; Falvo, E.; Giatti, S.; Cavaletti, G.; Melcangi, R.C. Alterations of gut microbiota composition in post-finasteride patients: A pilot study. J. Endocrinol. Investig. 2021, 44, 1263–1273. [Google Scholar] [CrossRef]
- Backstrom, T.; Doverskog, M.; Blackburn, T.P.; Scharschmidt, B.F.; Felipo, V. Allopregnanolone and its antagonist modulate neuroinflammation and neurological impairment. Neurosci. Biobehav. Rev. 2024, 161, 105668. [Google Scholar] [CrossRef]
- Diviccaro, S.; Cioffi, L.; Falvo, E.; Giatti, S.; Melcangi, R.C. Allopregnanolone: An overview on its synthesis and effects. J. Neuroendocrinol. 2022, 34, e12996. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.C.; Buffington, S.A.; Salas, R. Microbiota-Gut-Brain Axis in Psychiatry: Focus on Depressive Disorders. Curr. Epidemiol. Rep. 2024, 11, 222–232. [Google Scholar] [CrossRef]
- Cussotto, S.; Sandhu, K.V.; Dinan, T.G.; Cryan, J.F. The Neuroendocrinology of the Microbiota-Gut-Brain Axis: A Behavioural Perspective. Front. Neuroendocrinol. 2018, 51, 80–101. [Google Scholar] [CrossRef] [PubMed]
- Serra, D.; Almeida, L.M.; Dinis, T.C.P. The Impact of Chronic Intestinal Inflammation on Brain Disorders: The Microbiota-Gut-Brain Axis. Mol. Neurobiol. 2019, 56, 6941–6951. [Google Scholar] [CrossRef]
- Gasaly, N.; de Vos, P.; Hermoso, M.A. Impact of Bacterial Metabolites on Gut Barrier Function and Host Immunity: A Focus on Bacterial Metabolism and Its Relevance for Intestinal Inflammation. Front Immunol 2021, 12, 658354. [Google Scholar] [CrossRef]
- Harusato, A.; Geem, D.; Denning, T.L. Macrophage Isolation from the Mouse Small and Large Intestine. Methods Mol. Biol. 2016, 1422, 171–180. [Google Scholar]
- Fraga, C.G.; Leibovitz, B.E.; Tappel, A.L. Lipid peroxidation measured as thiobarbituric acid-reactive substances in tissue slices: Characterization and comparison with homogenates and microsomes. Free Radic. Biol. Med. 1988, 4, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Bialkowska, A.B.; Ghaleb, A.M.; Nandan, M.O.; Yang, V.W. Improved Swiss-rolling Technique for Intestinal Tissue Preparation for Immunohistochemical and Immunofluorescent Analyses. J. Vis. Exp. 2016, 113, 54161. [Google Scholar]
- Bihan, D.G.; Rydzak, T.; Wyss, M.; Pittman, K.; McCoy, K.D.; Lewis, I.A. Method for absolute quantification of short chain fatty acids via reverse phase chromatography mass spectrometry. PLoS ONE 2022, 17, e0267093. [Google Scholar] [CrossRef] [PubMed]
- Backstrom, T.; Bixo, M.; Johansson, M.; Nyberg, S.; Ossewaarde, L.; Ragagnin, G.; Savic, I.; Stromberg, J.; Timby, E.; van Broekhoven, F.; et al. Allopregnanolone and mood disorders. Prog. Neurobiol. 2014, 113, 88–94. [Google Scholar] [CrossRef]
- King, S.R. Emerging roles for neurosteroids in sexual behavior and function. J. Androl. 2008, 29, 524–533. [Google Scholar] [CrossRef]
- Schule, C.; Romeo, E.; Uzunov, D.P.; Eser, D.; di Michele, F.; Baghai, T.C.; Pasini, A.; Schwarz, M.; Kempter, H.; Rupprecht, R. Influence of mirtazapine on plasma concentrations of neuroactive steroids in major depression and on 3alpha-hydroxysteroid dehydrogenase activity. Mol. Psychiatry 2006, 11, 261–272. [Google Scholar] [CrossRef]
- Rasmusson, A.M.; Pinna, G.; Paliwal, P.; Weisman, D.; Gottschalk, C.; Charney, D.; Krystal, J.; Guidotti, A. Decreased cerebrospinal fluid allopregnanolone levels in women with posttraumatic stress disorder. Biol. Psychiatry 2006, 60, 704–713. [Google Scholar] [CrossRef]
- Timby, E.; Backstrom, T.; Nyberg, S.; Stenlund, H.; Wihlback, A.N.; Bixo, M. Women with premenstrual dysphoric disorder have altered sensitivity to allopregnanolone over the menstrual cycle compared to controls-a pilot study. Psychopharmacology 2016, 233, 2109–2117. [Google Scholar] [CrossRef]
- Osborne, L.M.; Gispen, F.; Sanyal, A.; Yenokyan, G.; Meilman, S.; Payne, J.L. Lower allopregnanolone during pregnancy predicts postpartum depression: An exploratory study. Psychoneuroendocrinology 2017, 79, 116–121. [Google Scholar] [CrossRef]
- Nappi, R.E.; Abbiati, I.; Luisi, S.; Ferdeghini, F.; Polatti, F.; Genazzani, A.R. Serum allopregnanolone levels relate to FSFI score during the menstrual cycle. J. Sex. Marital. Ther. 2003, 29 (Suppl 1.), 95–102. [Google Scholar] [CrossRef]
- Maguire, J.L.; Mennerick, S. Neurosteroids: Mechanistic considerations and clinical prospects. Neuropsychopharmacology 2024, 49, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. FDA approves first oral drug for postpartum depression, but rejects it for major depressive disorder. Nat. Rev. Drug Discov. 2023, 22, 774. [Google Scholar] [CrossRef] [PubMed]
- McCurry, M.D.; D’Agostino, G.D.; Walsh, J.T.; Bisanz, J.E.; Zalosnik, I.; Dong, X.; Morris, D.J.; Korzenik, J.R.; Edlow, A.G.; Balskus, E.P.; et al. Gut bacteria convert glucocorticoids into progestins in the presence of hydrogen gas. Cell 2024, 187, 2952–2968. [Google Scholar] [CrossRef] [PubMed]
- Seifi, M.; Brown, J.F.; Mills, J.; Bhandari, P.; Belelli, D.; Lambert, J.J.; Rudolph, U.; Swinny, J.D. Molecular and functional diversity of GABA-A receptors in the enteric nervous system of the mouse colon. J. Neurosci. 2014, 34, 10361–10378. [Google Scholar] [CrossRef]
- Auteri, M.; Zizzo, M.G.; Serio, R. GABA and GABA receptors in the gastrointestinal tract: From motility to inflammation. Pharmacol. Res. 2015, 93, 11–21. [Google Scholar] [CrossRef]
- Seifi, M.; Swinny, J.D. Developmental and age-dependent plasticity of GABA(A) receptors in the mouse colon: Implications in colonic motility and inflammation. Auton. Neurosci. 2019, 221, 102579. [Google Scholar] [CrossRef]
- Reyes-Garcia, M.G.; Hernandez-Hernandez, F.; Hernandez-Tellez, B.; Garcia-Tamayo, F. GABA (A) receptor subunits RNA expression in mice peritoneal macrophages modulate their IL-6/IL-12 production. J. Neuroimmunol. 2007, 188, 64–68. [Google Scholar] [CrossRef]
- Zhang, K.; Guo, J.; Yan, W.; Xu, L. Macrophage polarization in inflammatory bowel disease. Cell Commun. Signal 2023, 21, 367. [Google Scholar] [CrossRef]
- Yang, S.; Zhao, M.; Jia, S. Macrophage: Key player in the pathogenesis of autoimmune diseases. Front Immunol 2023, 14, 1080310. [Google Scholar] [CrossRef]
- Isidro, R.A. Appleyard CB: Colonic macrophage polarization in homeostasis, inflammation, and cancer. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G59–G73. [Google Scholar] [CrossRef]
- Dubuquoy, L.; Rousseaux, C.; Thuru, X.; Peyrin-Biroulet, L.; Romano, O.; Chavatte, P.; Chamaillard, M.; Desreumaux, P. PPARgamma as a new therapeutic target in inflammatory bowel diseases. Gut 2006, 55, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Annese, V.; Rogai, F.; Settesoldi, A.; Bagnoli, S. PPARgamma in Inflammatory Bowel Disease. PPAR Res. 2012, 2012, 620839. [Google Scholar] [CrossRef]
- Pinna, G. Role of PPAR-Allopregnanolone Signaling in Behavioral and Inflammatory Gut-Brain Axis Communications. Biol. Psychiatry 2023, 94, 609–618. [Google Scholar] [CrossRef]
- Locci, A.; Pinna, G. Stimulation of Peroxisome Proliferator-Activated Receptor-alpha by N-Palmitoylethanolamine Engages Allopregnanolone Biosynthesis to Modulate Emotional Behavior. Biol. Psychiatry 2019, 85, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Zulinska, S.; Strosznajder, A.K.; Strosznajder, J.B. Current View on PPAR-alpha and Its Relation to Neurosteroids in Alzheimer’s Disease and Other Neuropsychiatric Disorders: Promising Targets in a Therapeutic Strategy. Int. J Mol. Sci. 2024, 25. [Google Scholar] [CrossRef]
- Gustafsson, J.K.; Johansson, M.E.V. The role of goblet cells and mucus in intestinal homeostasis. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 785–803. [Google Scholar] [CrossRef] [PubMed]
- Nowarski, R.; Jackson, R.; Gagliani, N.; de Zoete, M.R.; Palm, N.W.; Bailis, W.; Low, J.S.; Harman, C.C.; Graham, M.; Elinav, E.; et al. Epithelial IL-18 Equilibrium Controls Barrier Function in Colitis. Cell 2015, 163, 1444–1456. [Google Scholar] [CrossRef]
- Danese, S.; Fiocchi, C. Ulcerative colitis. N. Engl. J. Med. 2011, 365, 1713–1725. [Google Scholar] [CrossRef]
- Grabrucker, S.; Marizzoni, M.; Silajdzic, E.; Lopizzo, N.; Mombelli, E.; Nicolas, S.; Dohm-Hansen, S.; Scassellati, C.; Moretti, D.V.; Rosa, M.; et al. Microbiota from Alzheimer’s patients induce deficits in cognition and hippocampal neurogenesis. Brain 2023, 146, 4916–4934. [Google Scholar] [CrossRef]
- Song, C.; Chai, Z.; Chen, S.; Zhang, H.; Zhang, X.; Zhou, Y. Intestinal mucus components and secretion mechanisms: What we do and do not know. Exp. Mol. Med. 2023, 55, 681–691. [Google Scholar] [CrossRef]
- Erben, U.; Loddenkemper, C.; Doerfel, K.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kuhl, A.A. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557–4576. [Google Scholar] [PubMed]
- Qiu, J.; Liu, R.; Ma, Y.; Li, Y.; Chen, Z.; He, H.; Chen, J.; Tong, L.; Huang, C.; You, Q. Lipopolysaccharide-Induced Depression-Like Behaviors Is Ameliorated by Sodium Butyrate via Inhibiting Neuroinflammation and Oxido-Nitrosative Stress. Pharmacology 2020, 105, 550–560. [Google Scholar] [CrossRef]
- Resende, W.R.; Valvassori, S.S.; Reus, G.Z.; Varela, R.B.; Arent, C.O.; Ribeiro, K.F.; Bavaresco, D.V.; Andersen, M.L.; Zugno, A.I.; Quevedo, J. Effects of sodium butyrate in animal models of mania and depression: Implications as a new mood stabilizer. Behav. Pharmacol. 2013, 24, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Firoozi, D.; Masoumi, S.J.; Mohammad-Kazem Hosseini Asl, S.; Fararouei, M.; Jamshidi, S. Effects of Short Chain Fatty Acid-Butyrate Supplementation on the Disease Severity, Inflammation, and Psychological Factors in Patients with Active Ulcerative Colitis: A Double-Blind Randomized Controlled Trial. J. Nutr. Metab. 2025, 2025, 3165876. [Google Scholar] [CrossRef] [PubMed]
- Firoozi, D.; Masoumi, S.J.; Mohammad-Kazem Hosseini Asl, S.; Labbe, A.; Razeghian-Jahromi, I.; Fararouei, M.; Lankarani, K.B.; Dara, M. Effects of short-chain fatty acid-butyrate supplementation on expression of circadian-clock genes, sleep quality, and inflammation in patients with active ulcerative colitis: A double-blind randomized controlled trial. Lipids Health Dis. 2024, 23, 216. [Google Scholar] [CrossRef]
- Ferreira-Halder, C.V.; Faria, A.V.S.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best. Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]
- Stilling, R.M.; van de Wouw, M.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. The neuropharmacology of butyrate: The bread and butter of the microbiota-gut-brain axis? Neurochem. Int. 2016, 99, 110–132. [Google Scholar] [CrossRef]
- Filiano, A.J.; Gadani, S.P.; Kipnis, J. Interactions of innate and adaptive immunity in brain development and function. Brain. Res. 2015, 1617, 18–27. [Google Scholar] [CrossRef]
- Huuskonen, J.; Suuronen, T.; Nuutinen, T.; Kyrylenko, S.; Salminen, A. Regulation of microglial inflammatory response by sodium butyrate and short-chain fatty acids. Br. J. Pharmacol. 2004, 141, 874–880. [Google Scholar] [CrossRef]
- Soliman, M.L.; Ohm, J.E.; Rosenberger, T.A. Acetate reduces PGE2 release and modulates phospholipase and cyclooxygenase levels in neuroglia stimulated with lipopolysaccharide. Lipids 2013, 48, 651–662. [Google Scholar] [CrossRef]
- Soliman, M.L.; Puig, K.L.; Combs, C.K.; Rosenberger, T.A. Acetate reduces microglia inflammatory signaling in vitro. J. Neurochem. 2012, 123, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Hirschberg, S.; Gisevius, B.; Duscha, A.; Haghikia, A. Implications of Diet and The Gut Microbiome in Neuroinflammatory and Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 3109. [Google Scholar] [CrossRef]
- Giatti, S.; Cioffi, L.; Diviccaro, S.; Piazza, R.; Melcangi, R.C. Analysis of the finasteride treatment and its withdrawal in the rat hypothalamus and hippocampus at whole-transcriptome level. J. Endocrinol. Investig. 2024, 47, 2565–2574. [Google Scholar] [CrossRef]
- Schneeberger, E.E.; Lynch, R.D. The tight junction: A multifunctional complex. Am. J. Physiol. Cell Physiol. 2004, 286, C1213–C1228. [Google Scholar] [CrossRef] [PubMed]
- Greene, C.; Hanley, N.; Campbell, M. Claudin-5: Gatekeeper of neurological function. Fluids Barriers CNS 2019, 16, 3. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Wang, Z.; Liu, Y.; Wang, P.; Xue, Y. Specific role of tight junction proteins claudin-5, occludin, and ZO-1 of the blood-brain barrier in a focal cerebral ischemic insult. J. Mol. Neurosci. 2011, 44, 130–139. [Google Scholar] [CrossRef]
- Mullier, A.; Bouret, S.G.; Prevot, V.; Dehouck, B. Differential distribution of tight junction proteins suggests a role for tanycytes in blood-hypothalamus barrier regulation in the adult mouse brain. J. Comp. Neurol. 2010, 518, 943–962. [Google Scholar] [CrossRef]
- Almeida, F.B.; Pinna, G.; Barros, H.M.T. The Role of HPA Axis and Allopregnanolone on the Neurobiology of Major Depressive Disorders and PTSD. Int. J. Mol. Sci. 2021, 22, 5495. [Google Scholar] [CrossRef]
- Kurita, K.; Ohta, H.; Shirakawa, I.; Tanaka, M.; Kitaura, Y.; Iwasaki, Y.; Matsuzaka, T.; Shimano, H.; Aoe, S.; Arima, H.; et al. Macrophages rely on extracellular serine to suppress aberrant cytokine production. Sci. Rep. 2021, 11, 11137. [Google Scholar] [CrossRef]
- Wang, F.; Zhou, H.; Deng, L.; Wang, L.; Chen, J.; Zhou, X. Serine Deficiency Exacerbates Inflammation and Oxidative Stress via Microbiota-Gut-Brain Axis in D-Galactose-Induced Aging Mice. Mediators Inflamm. 2020, 2020, 5821428. [Google Scholar] [CrossRef]
- Amaral, A.U.; Ferreira, G.C.; Seminotti, B.; Leipnitz, G.; Wajner, M. Glutaric acid neurotoxicity: Mechanisms and actions. In Handbook of Neurotoxicity; Kostrzewa, R.M., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 1–35. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diviccaro, S.; Oleari, R.; Amoruso, F.; Fontana, F.; Cioffi, L.; Chrostek, G.; Abenante, V.; Troisi, J.; Cariboni, A.; Giatti, S.; et al. Exploration of the Possible Relationships Between Gut and Hypothalamic Inflammation and Allopregnanolone: Preclinical Findings in a Post-Finasteride Rat Model. Biomolecules 2025, 15, 1044. https://doi.org/10.3390/biom15071044
Diviccaro S, Oleari R, Amoruso F, Fontana F, Cioffi L, Chrostek G, Abenante V, Troisi J, Cariboni A, Giatti S, et al. Exploration of the Possible Relationships Between Gut and Hypothalamic Inflammation and Allopregnanolone: Preclinical Findings in a Post-Finasteride Rat Model. Biomolecules. 2025; 15(7):1044. https://doi.org/10.3390/biom15071044
Chicago/Turabian StyleDiviccaro, Silvia, Roberto Oleari, Federica Amoruso, Fabrizio Fontana, Lucia Cioffi, Gabriela Chrostek, Vera Abenante, Jacopo Troisi, Anna Cariboni, Silvia Giatti, and et al. 2025. "Exploration of the Possible Relationships Between Gut and Hypothalamic Inflammation and Allopregnanolone: Preclinical Findings in a Post-Finasteride Rat Model" Biomolecules 15, no. 7: 1044. https://doi.org/10.3390/biom15071044
APA StyleDiviccaro, S., Oleari, R., Amoruso, F., Fontana, F., Cioffi, L., Chrostek, G., Abenante, V., Troisi, J., Cariboni, A., Giatti, S., & Melcangi, R. C. (2025). Exploration of the Possible Relationships Between Gut and Hypothalamic Inflammation and Allopregnanolone: Preclinical Findings in a Post-Finasteride Rat Model. Biomolecules, 15(7), 1044. https://doi.org/10.3390/biom15071044