
Antimicrobial Indole-3-Carboxamido-Polyamine Conjugates Target Bacterial Membranes and Are Antibiotic Potentiators

 , ,
, , 
 and
and
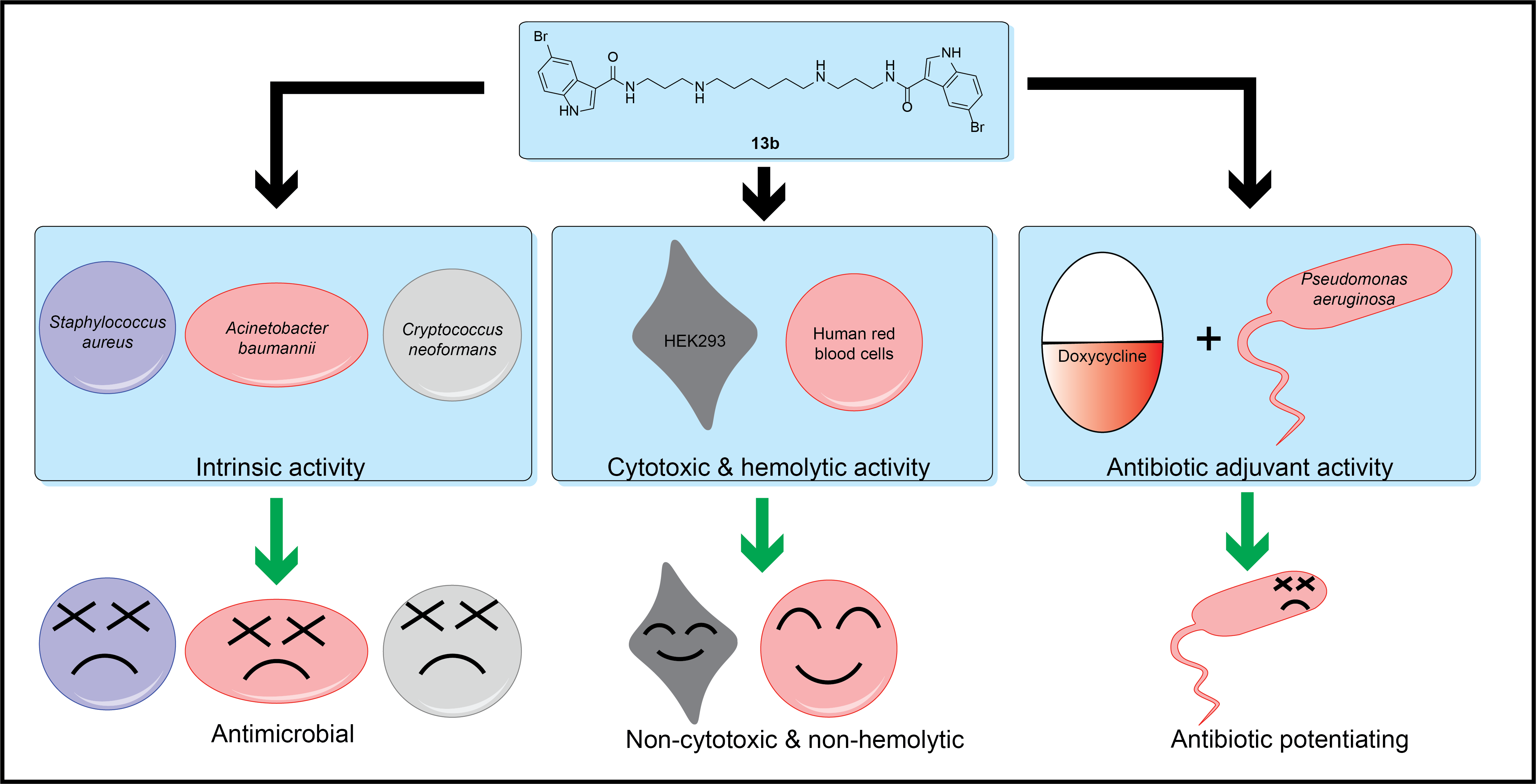
Abstract

1. Introduction
2. Materials and Methods
2.1. Chemistry Synthesis General Methods
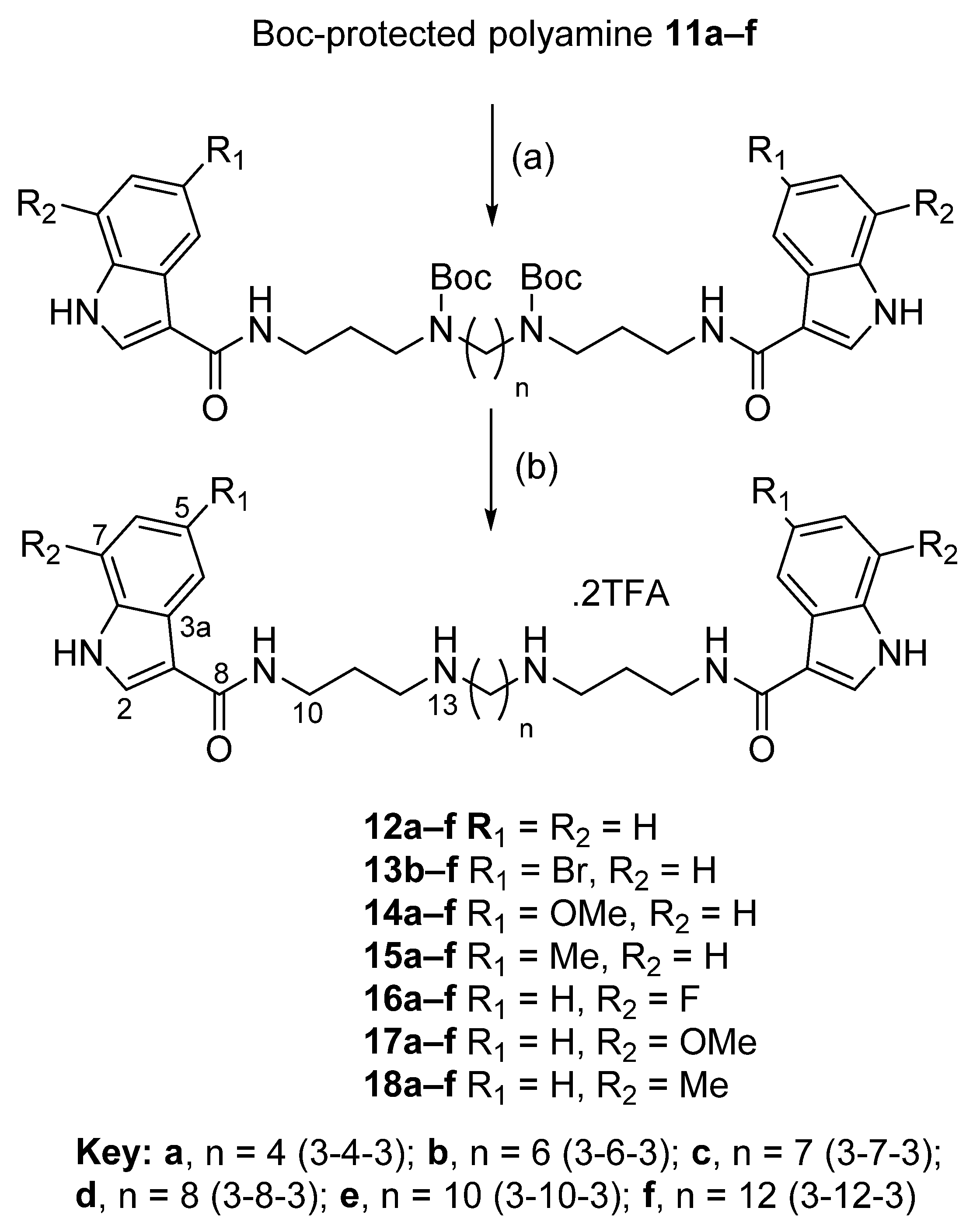
2.1.1. General Procedure A: Amide Bond Formation of Indole Analogues
2.1.2. General Procedure B—Boc Deprotection
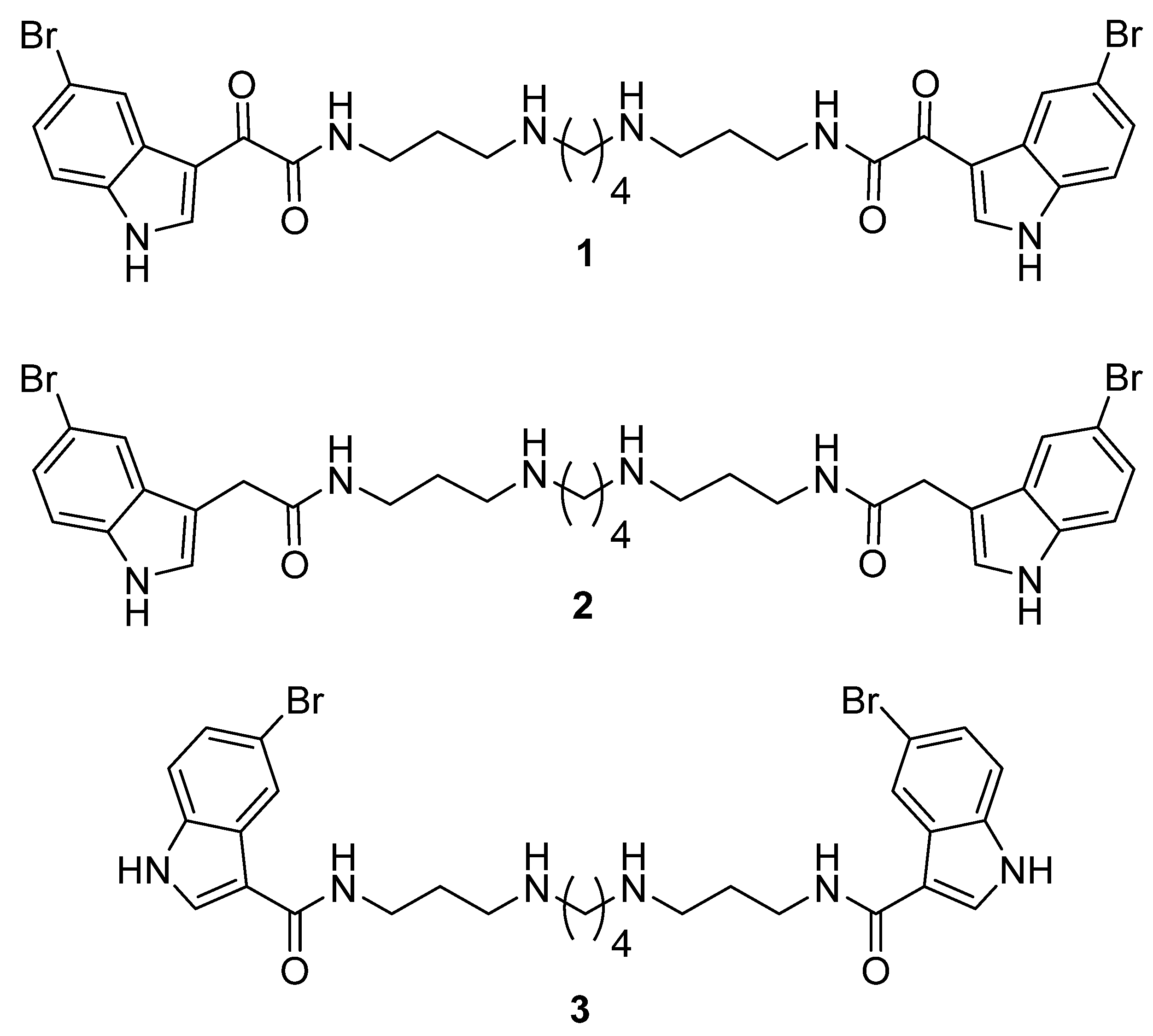
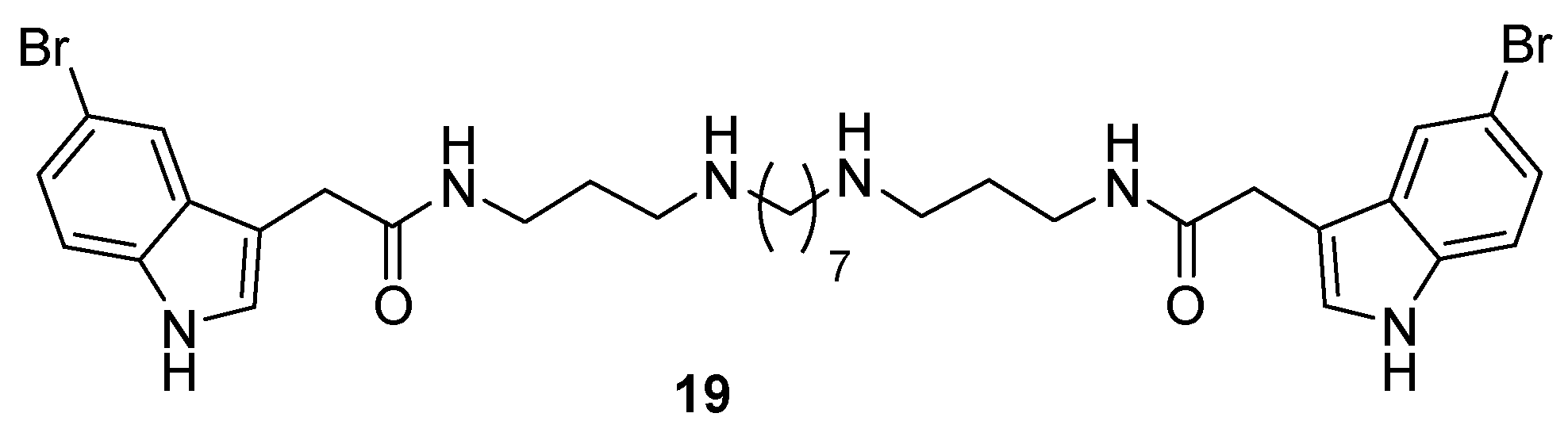
2.2. Synthesis of Compounds
2.3. Antimicrobial Assays
2.4. Real-Time Growth Curves
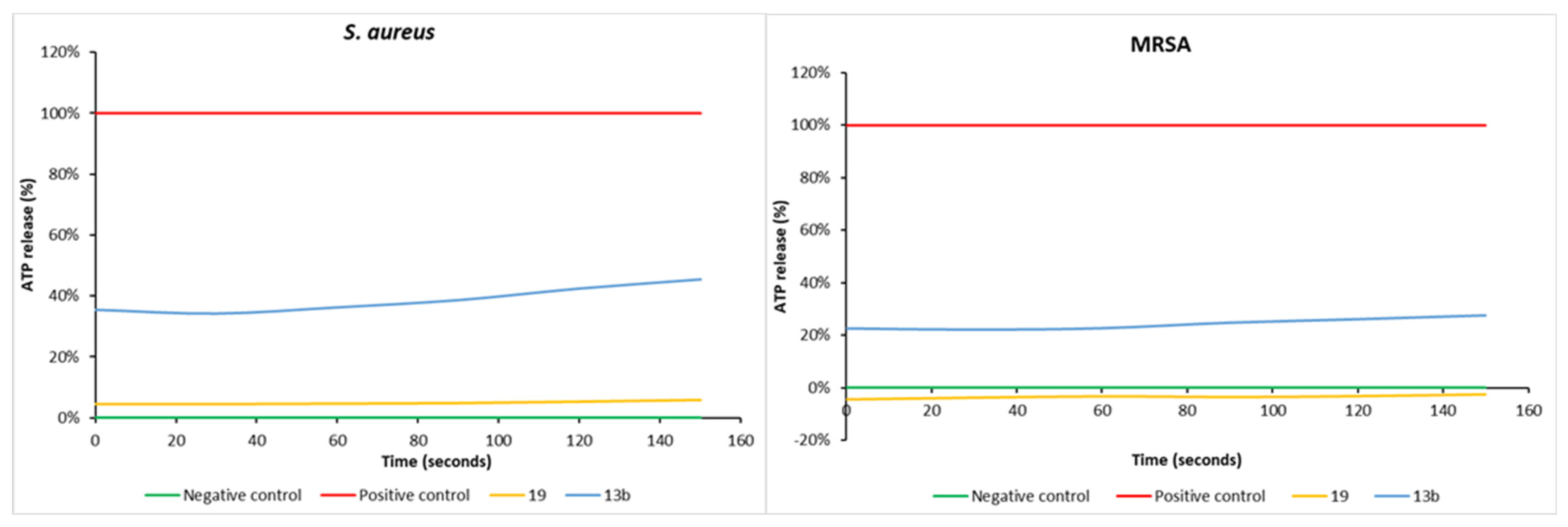
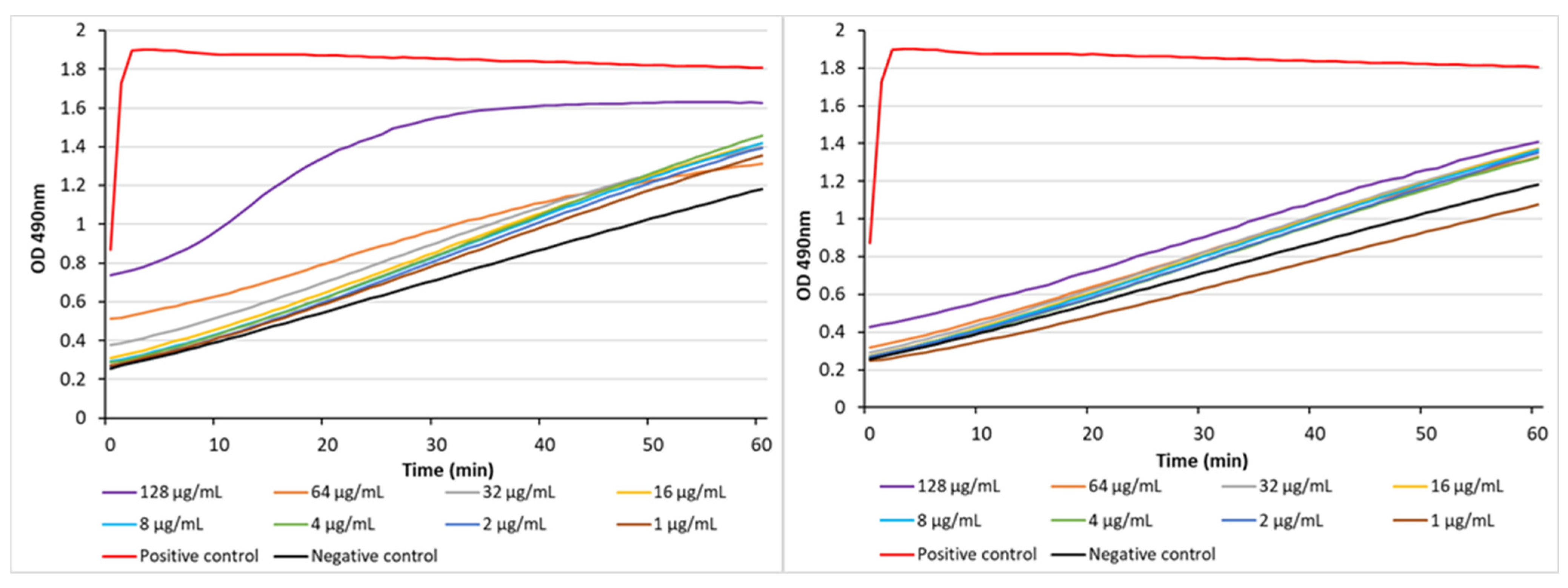
2.5. Measurement of the ATP Efflux
2.6. Determination of the MICs of Antibiotics in the Presence of Synergizing Compounds
2.7. Nitrocefin Hydrolysis Assay
2.8. Cytotoxicity Assays
2.9. Hemolytic Assay
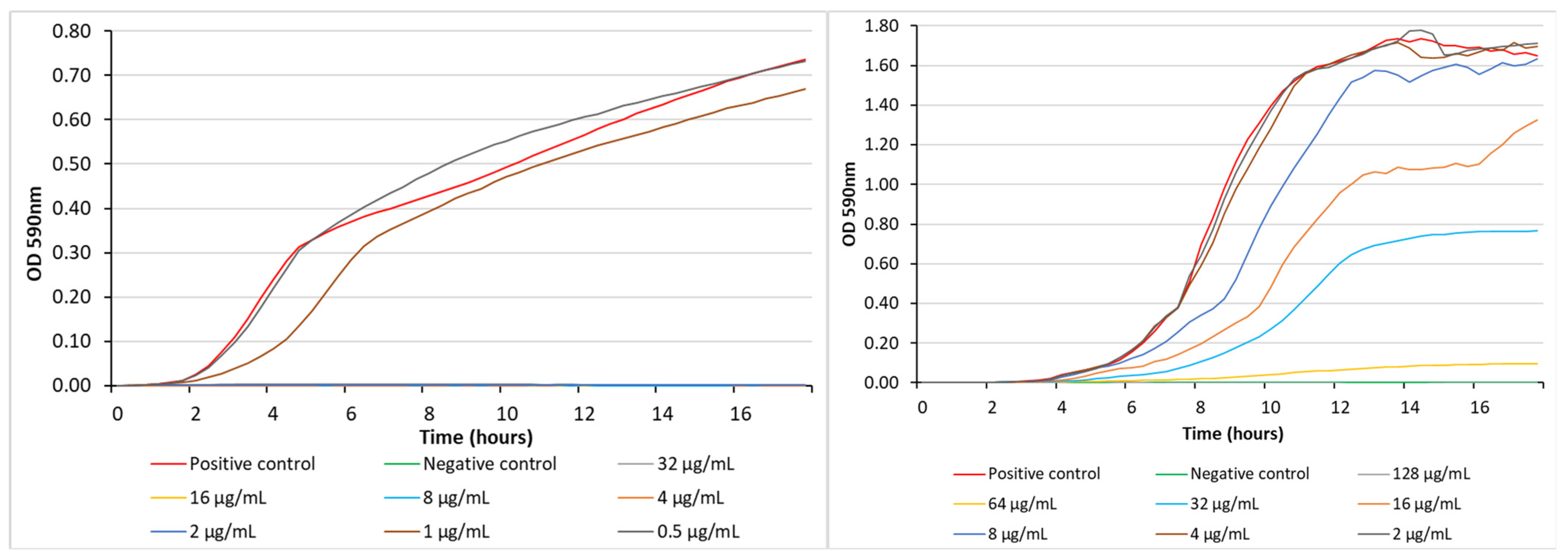
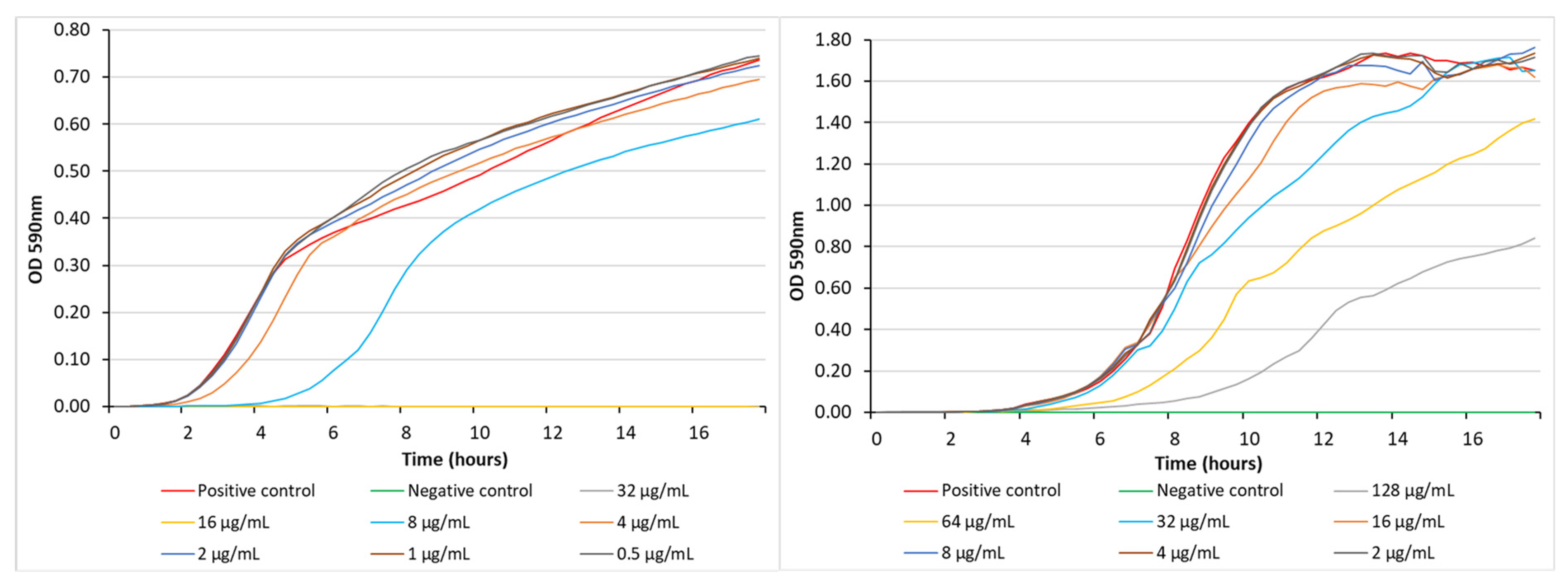
3. Results and Discussion
Chemistry
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silver, L.L. Challenges of Antibacterial Discovery. Clin. Microbiol. Rev. 2011, 24, 71–109. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. The Science of Antibiotic Discovery. Cell 2020, 181, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.B. Confronting the Challenges of Discovery of Novel Antibacterial Agents. Bioorg. Med. Chem. Lett. 2014, 24, 3683–3689. [Google Scholar] [CrossRef] [PubMed]
- Tommasi, R.; Brown, D.G.; Walkup, G.K.; Manchester, J.I.; Miller, A.A. ESKAPEing the Labyrinth of Antibacterial Discovery. Nat. Rev. Drug Discov. 2015, 14, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Payne, D.J.; Gwynn, M.N.; Holmes, D.J.; Pompliano, D.L. Drugs for Bad Bugs: Confronting the Challenges of Antibacterial Discovery. Nat. Rev. Drug Discov. 2007, 6, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Alekshun, M.N.; Levy, S.B. Molecular Mechanisms of Antibacterial Multidrug Resistance. Cell 2007, 128, 1037–1050. [Google Scholar] [CrossRef]
- Melander, R.J.; Melander, C. The Challenge of Overcoming Antibiotic Resistance: An Adjuvant Approach? ACS Infect. Dis. 2017, 3, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Farha, M.A.; Brown, E.D. Discovery of Antibiotic Adjuvants. Nat. Biotechnol. 2013, 31, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Antibiotic Adjuvants: Rescuing Antibiotics from Resistance. Trends Microbiol. 2016, 24, 862–871. [Google Scholar] [CrossRef]
- Melander, R.J.; Mattingly, A.E.; Nemeth, A.M.; Melander, C. Overcoming Intrinsic Resistance in Gram-Negative Bacteria Using Small Molecule Adjuvants. Bioorg. Med. Chem. Lett. 2023, 80, 129113. [Google Scholar] [CrossRef]
- Cadelis, M.M.; Liu, T.; Sue, K.; Rouvier, F.; Bourguet-Kondracki, M.-L.; Brunel, J.M.; Copp, B.R. Structure–Activity Relationship Studies of Indolglyoxyl-Polyamine Conjugates as Antimicrobials and Antibiotic Potentiators. Pharmaceuticals 2023, 16, 823. [Google Scholar] [CrossRef] [PubMed]
- Sue, K.; Cadelis, M.M.; Gill, E.S.; Rouvier, F.; Bourguet-Kondracki, M.-L.; Brunel, J.M.; Copp, B.R. Indole-3-Acetamido-Polyamines as Antimicrobial Agents and Antibiotic Adjuvants. Biomolecules 2023, 13, 1226. [Google Scholar] [CrossRef] [PubMed]
- Cadelis, M.M.; Li, S.A.; Bourguet-Kondracki, M.; Blanchet, M.; Douafer, H.; Brunel, J.M.; Copp, B.R. Spermine Derivatives of Indole-3-carboxylic Acid, Indole-3-acetic Acid and Indole-3-acrylic Acid as Gram-Negative Antibiotic Adjuvants. ChemMedChem 2021, 16, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Blaskovich, M.A.T.; Zuegg, J.; Elliott, A.G.; Cooper, M.A. Helping Chemists Discover New Antibiotics. ACS Infect. Dis. 2015, 1, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Katsu, T.; Nakagawa, H.; Yasuda, K. Interaction between Polyamines and Bacterial Outer Membranes as Investigated with Ion-Selective Electrodes. Antimicrob. Agents Chemother. 2002, 46, 1073–1079. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yasuda, K.; Ohmizo, C.; Katsu, T. Mode of Action of Novel Polyamines Increasing the Permeability of Bacterial Outer Membrane. Int. J. Antimicrob. Agents 2004, 24, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.A.; McCormack, C.E.M.; Wuest, W.M. Deriving Novel Quaternary Ammonium Compound Disinfectant Scaffolds from a Natural Product: Mechanistic Insights of the Quaternization of Ianthelliformisamine C. ChemMedChem 2023, 18, e202300253. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Pachaiyappan, B.; Gruber, J.D.; Schmidt, M.G.; Zhang, Y.-M.; Woster, P.M. Antibacterial Diamines Targeting Bacterial Membranes. J. Med. Chem. 2016, 59, 3140–3151. [Google Scholar] [CrossRef] [PubMed]
- Mammari, N.; Salles, E.; Beaussart, A.; El-Kirat-Chatel, S.; Varbanov, M. Squalamine and Its Aminosterol Derivatives: Overview of Biological Effects and Mechanisms of Action of Compounds with Multiple Therapeutic Applications. Microorganisms 2022, 10, 1205. [Google Scholar] [CrossRef] [PubMed]
- Kazakova, O.; Giniyatullina, G.; Babkov, D.; Wimmer, Z. From Marine Metabolites to the Drugs of the Future: Squalamine, Trodusquemine, Their Steroid and Triterpene Analogues. Int. J. Mol. Sci. 2022, 23, 1075. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
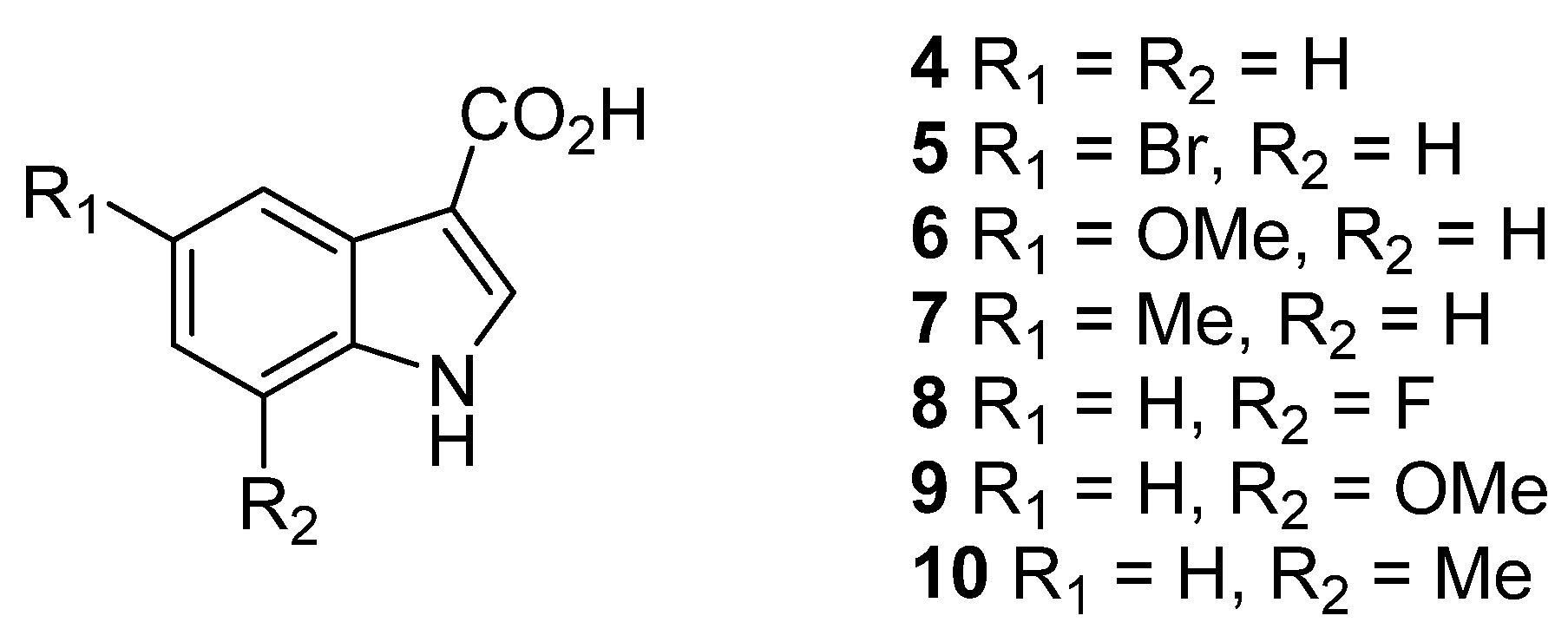{kind=link}
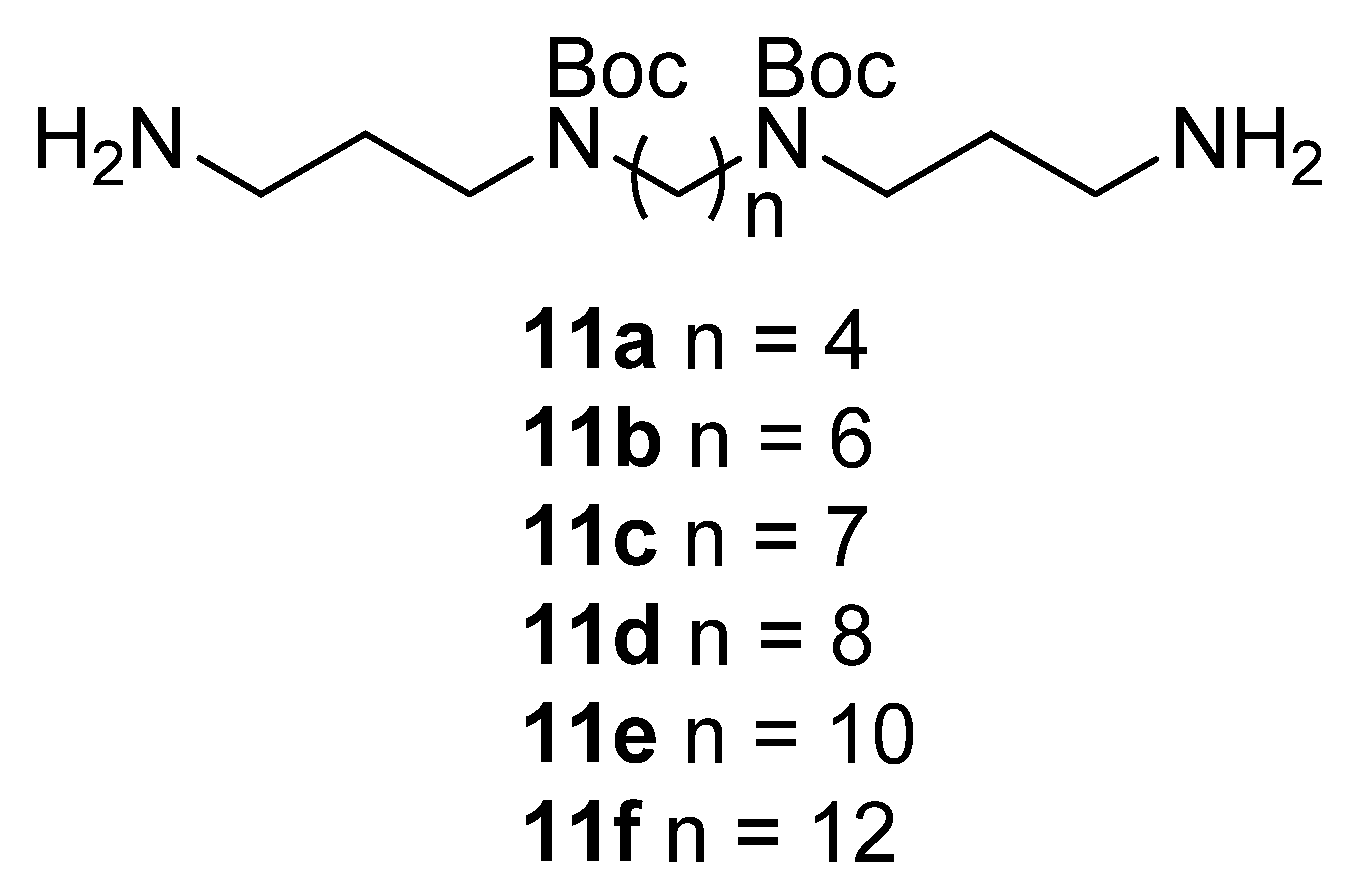{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | MIC (µM) | |||||||
|---|---|---|---|---|---|---|---|---|
| Sa a | MRSA b | Pa c | Ec d | Kp e | Ab f | Ca g | Cn h | |
| 12a | 69.8 | 22.3 | >279 i | 139 | >45 j | >45 | >45 | n.t k |
| 12b | 268 | ≤0.34 | >268 | 134 | >43 | >43 | >43 | ≤0.34 |
| 12c | 132 | ≤0.33 | >263 | 65.9 | >42 | >42 | >42 | ≤0.33 |
| 12d | 64.7 | ≤0.32 | >259 | 32.4 | >41 | >41 | >41 | ≤0.32 |
| 12e | 31.2 | ≤0.31 | >250 | 125 | >40 | >40 | >40 | ≤0.31 |
| 12f | 3.77 | ≤0.30 | 121 | 15.1 | >39 | 4.82 | ≤0.30 | ≤0.30 |
| 3 | 4.8 i | 12.4 i | 200 i | 200 i | >50 i | >50 i | 49.5 i | 12.4 i |
| 13b | 2.2 | ≤0.28 | 71 | 6.92 | >35 | ≤0.28 | >35 | ≤0.28 |
| 13c | 13.6 | ≤0.27 | 220 | 110 | >35 | >35 | 35 | ≤0.27 |
| 13d | 3.36 | ≤0.27 | 107 | 53.7 | >34 | ≤0.27 | ≤0.27 | ≤0.27 |
| 13e | 3.26 | ≤0.26 | 104 | 52.2 | >34 | 16.7 | 4.17 | ≤0.26 |
| 13f | 3.17 | ≤0.25 | 203 | >203 | ≤0.25 | 2.03 | ≤0.25 | ≤0.25 |
| 14a | 257 | 10.3 | >257 | 257 | >41 | >41 | >41 | n.t k |
| 14b | 248 | ≤0.31 | >248 | 248 | >40 | >40 | >40 | ≤0.31 |
| 14c | 122 | >39 | >244 | >244 | >39 | >39 | >39 | n.t k |
| 14d | 60.0 | 19.2 | >240 | 120 | >38 | >38 | >38 | ≤0.30 |
| 14e | 29.0 | ≤0.29 | 232 | 58.1 | >37 | >37 | >37 | ≤0.29 |
| 14f | 3.52 | ≤0.28 | 112 | 14.1 | >36 | >36 | ≤0.28 | ≤0.28 |
| 15a | 67.1 | ≤0.34 | >269 | >269 | >43 | >43 | >43 | ≤0.34 |
| 15b | 259 | 5.18 | >259 | 129 | >41 | >41 | >41 | ≤0.32 |
| 15c | 63.5 | ≤0.32 | >254 | 254 | >41 | >41 | >41 | ≤0.32 |
| 15d | 31.2 | ≤0.31 | 250 | 125 | >40 | >40 | >40 | ≤0.31 |
| 15e | 3.77 | ≤0.30 | 60.3 | 30.2 | >39 | 19.3 | ≤0.30 | ≤0.30 |
| 15f | 3.65 | ≤0.29 | 58.3 | 29.2 | >37.3 | ≤0.29 | ≤0.29 | ≤0.29 |
| 16a | >266 | >43 | 266 | >266 | >43 | >43 | >43 | n.t k |
| 16b | 256 | >41 | >256 | 256 | >41 | >41 | >41 | n.t k |
| 16c | >251 | ≤0.31 | 251 | >251 | >40 | >40 | >40 | ≤0.31 |
| 16d | 247 | 9.89 | 247 | 247 | >40 | >40 | >40 | ≤0.31 |
| 16e | 239 | ≤0.30 | 239 | 239 | >38 | >38 | ≤0.30 | ≤0.30 |
| 16f | 231 | ≤0.29 | 116 | 231 | >37 | >37 | >37 | ≤0.29 |
| 17a | 129 | 20.6 | >257 | >257 | >41 | >41 | >41 | n.t k |
| 17b | 124 | >40 | >248 | 248 | >40 | >40 | >40 | n.t k |
| 17c | 122 | 2.44 | >244 | 244 | >39 | >39 | >39 | ≤0.31 |
| 17d | 120 | ≤0.30 | >240 | 240 | >38 | >38 | >38 | ≤0.30 |
| 17e | 232 | ≤0.29 | >232 | >232 | >37 | >37 | >37 | ≤0.29 |
| 17f | 7.03 | ≤0.28 | 225 | 28.1 | >36 | >36 | 4.50 | ≤0.28 |
| 18a | >269 | 21.5 | 269 | >269 | >43 | >43 | >43 | n.t k |
| 18b | 129 | >41 | >259 | 129 | >41 | >41 | >41 | ≤0.32 |
| 18c | >254 | ≤0.32 | 254 | >254 | >41 | >41 | >41 | ≤0.32 |
| 18d | >250 | ≤0.31 | 250 | >250 | >40 | >40 | >40 | ≤0.31 |
| 18e | 30.2 | ≤0.30 | 60.3 | 30.2 | >39 | >39 | 4.83 | ≤0.30 |
| 18f | 117 | ≤0.29 | 117 | 117 | >37 | 9.33 | ≤0.29 | ≤0.29 |
| Streptomycin | 21.5 | 21.5 | 21.5 | |||||
| Chloramphenicol | 1.5–3 | |||||||
| Vancomycin | 0.7 | |||||||
| Colistin | 1 | 2 | 0.2 | 0.2 | ||||
| Fluconazole | 0.4 | 26 | ||||||
| Compound | Dox/Pa a | Erythro/Ec b | Compound | Dox/Pa a | Erythro/Ec b |
|---|---|---|---|---|---|
| 12a | 34.9 (>8) | 34.9 (4) | 15d | 31.2 (8) | 31.2 (4) |
| 12b | 33.6 (>8) | 33.6 (4) | 15e | 15.1 (4) | 7.54 (4) |
| 12c | 65.9 (>4) | 65.9 (1) | 15f | 117 (0.5) | 29.2 (1) |
| 12d | 32.3 (>8) | n.t c | 16a | 266 (1) | >266 (1) |
| 12e | 62.4 (>4) | 62.4 (2) | 16b | 64.0 (>4) | 128 (2) |
| 12f | 15.1 (8) | 7.54 (2) | 16c | 62.9 (>4) | 252 (1) |
| 3 | 3.125 c (64) | 100 (2) | 16d | 30.9 (>8) | 247 (1) |
| 13b | 3.46 (21) | 6.92 (1) | 16e | 29.9 (8) | 239 (1) |
| 13c | 13.6 (16) | 6.82 (16) | 16f | 116 (1) | 231 (1) |
| 13d | 13.4 (8) | 6.72 (8) | 17a | 129 (>2) | >257 (1) |
| 13e | 26.1 (4) | 6.52 (8) | 17b | >248 (1) | >248 (1) |
| 13f | 50.7 (4) | 25.3 (>8) | 17c | 30.5 (>8) | 244 (1) |
| 14a | 64.4 (>4) | 257 (1) | 17d | 60.0 (>4) | 120 (2) |
| 14b | 31.1 (>8) | 62.1 (4) | 17e | 58.1 (>4) | >232 (1) |
| 14c | 61.1 (>4) | 244 (>1) | 17f | 56.2 (4) | 56.2 (0.5) |
| 14d | 30.0 (>8) | 240 (0.5) | 18a | 269 (1) | >269 (1) |
| 14e | 29.0 (8) | 116 (0.5) | 18b | 32.4 (>8) | 32.4 (4) |
| 14f | 56.2 (2) | 14.1 (1) | 18c | 15.9 (16) | >254 (1) |
| 15a | 33.6 (>8) | 67.1 (>4) | 18d | 31.2 (8) | 250 (1) |
| 15b | 32.4 (>8) | 32.4 (4) | 18e | 15.1 (4) | 15.1(2) |
| 15c | 63.5 (>4) | 63.5 (4) | 18f | 29.2 (4) | 116.7 (1) |
| Compound | Cytotoxicity a | Hemolysis b | Compound | Cytotoxicity a | Hemolysis b |
|---|---|---|---|---|---|
| 12a | >45 | >45 | 15d | >40 | >40 |
| 12b | >43 | >43 | 15e | >39 | >39 |
| 12c | >42 | >42 | 15f | 0.55 | >37 |
| 12d | >41 | >41 | 16a | >43 | >43 |
| 12e | >40 | >40 | 16b | >41 | >41 |
| 12f | >39 | >39 | 16c | >40 | >40 |
| 3 | >50 c | >50 c | 16d | >40 | >40 |
| 13b | >35 | >35 | 16e | >38 | >38 |
| 13c | >35 | >35 | 16f | >37 | >37 |
| 13d | >34 | >34 | 17a | >41 | >41 |
| 13e | >33 | >33 | 17b | >40 | >40 |
| 13f | ≤0.25 | 14.2 | 17c | >39 | >39 |
| 14a | >41 | >41 | 17d | >38 | >38 |
| 14b | >40 | >40 | 17e | >37 | >37 |
| 14c | >39 | >39 | 17f | >36 | >36 |
| 14d | >38 | >38 | 18a | >43 | >43 |
| 14e | >37 | >37 | 18b | >41 | >41 |
| 14f | >36 | >36 | 18c | >41 | >41 |
| 15a | >43 | >43 | 18d | >40 | >40 |
| 15b | >41 | >41 | 18e | >39 | >39 |
| 15c | >41 | >41 | 18f | >37 | >37 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sue, K.; Cadelis, M.M.; Rouvier, F.; Bourguet-Kondracki, M.-L.; Brunel, J.M.; Copp, B.R. Antimicrobial Indole-3-Carboxamido-Polyamine Conjugates Target Bacterial Membranes and Are Antibiotic Potentiators. Biomolecules 2024, 14, 261. https://doi.org/10.3390/biom14030261
Sue K, Cadelis MM, Rouvier F, Bourguet-Kondracki M-L, Brunel JM, Copp BR. Antimicrobial Indole-3-Carboxamido-Polyamine Conjugates Target Bacterial Membranes and Are Antibiotic Potentiators. Biomolecules. 2024; 14(3):261. https://doi.org/10.3390/biom14030261
Chicago/Turabian StyleSue, Kenneth, Melissa M. Cadelis, Florent Rouvier, Marie-Lise Bourguet-Kondracki, Jean Michel Brunel, and Brent R. Copp. 2024. "Antimicrobial Indole-3-Carboxamido-Polyamine Conjugates Target Bacterial Membranes and Are Antibiotic Potentiators" Biomolecules 14, no. 3: 261. https://doi.org/10.3390/biom14030261
APA StyleSue, K., Cadelis, M. M., Rouvier, F., Bourguet-Kondracki, M.-L., Brunel, J. M., & Copp, B. R. (2024). Antimicrobial Indole-3-Carboxamido-Polyamine Conjugates Target Bacterial Membranes and Are Antibiotic Potentiators. Biomolecules, 14(3), 261. https://doi.org/10.3390/biom14030261