
Neurons with Cat’s Eyes: A Synthetic Strain of α-Synuclein Fibrils Seeding Neuronal Intranuclear Inclusions

 , , , , ,
, , , , ,  , ,
, ,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. α-Syn Expression
2.2. α-Syn Purification
2.3. α-Syn Fibrillization
2.4. Sonication
2.5. Electron Microscopy
2.6. X-ray Diffraction
2.7. Magic-Angle Spinning NMR
2.8. Fourier-Transform Infra-Red (FTIR) Spectroscopy
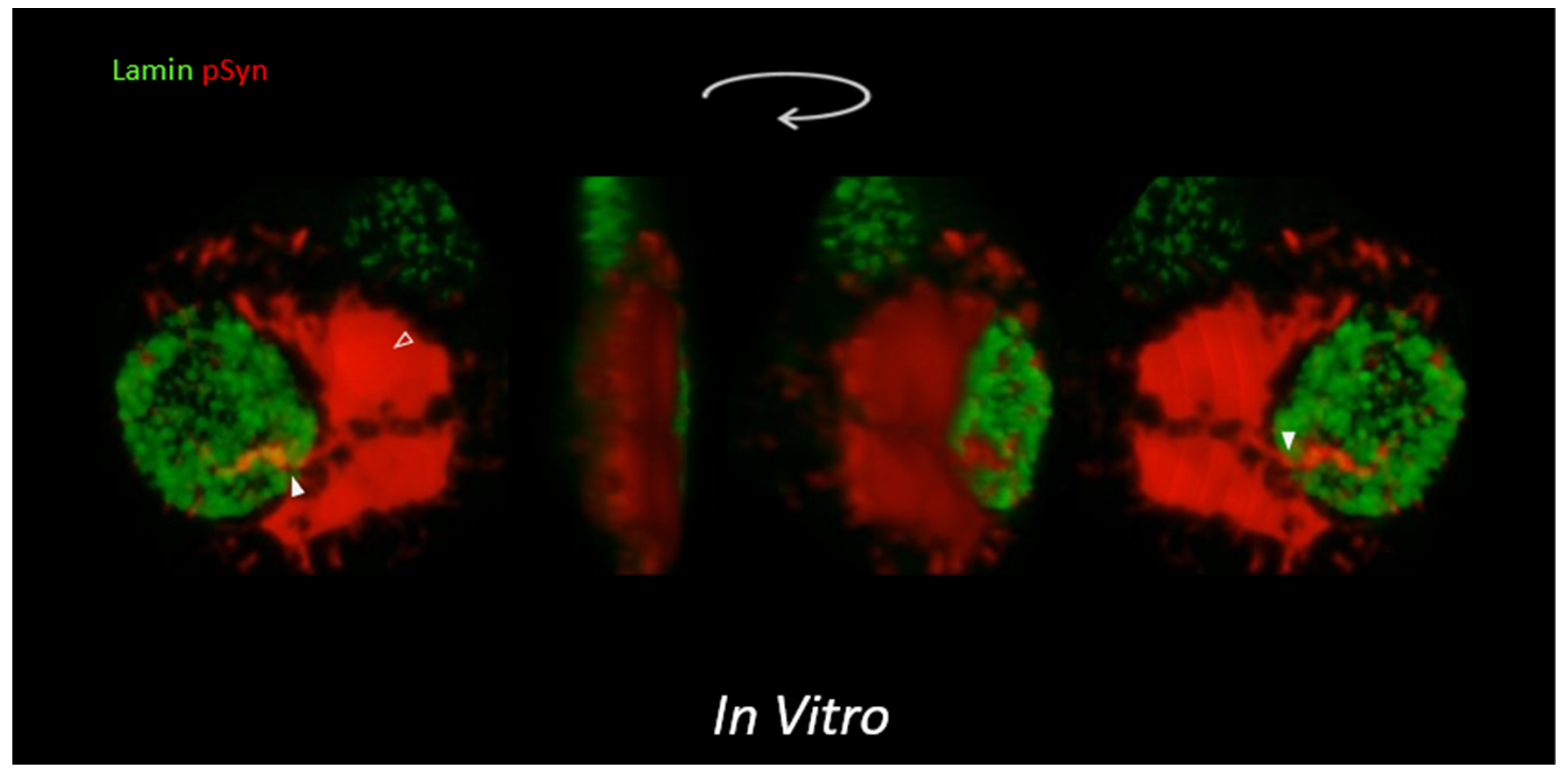
2.9. In Vitro α-Syn Pathology, High Content Analysis (HCA) and Laser-Scanning Confocal Microscopy (LSCM)
2.10. In Vivo α-Syn Pathology and Histology
3. Results
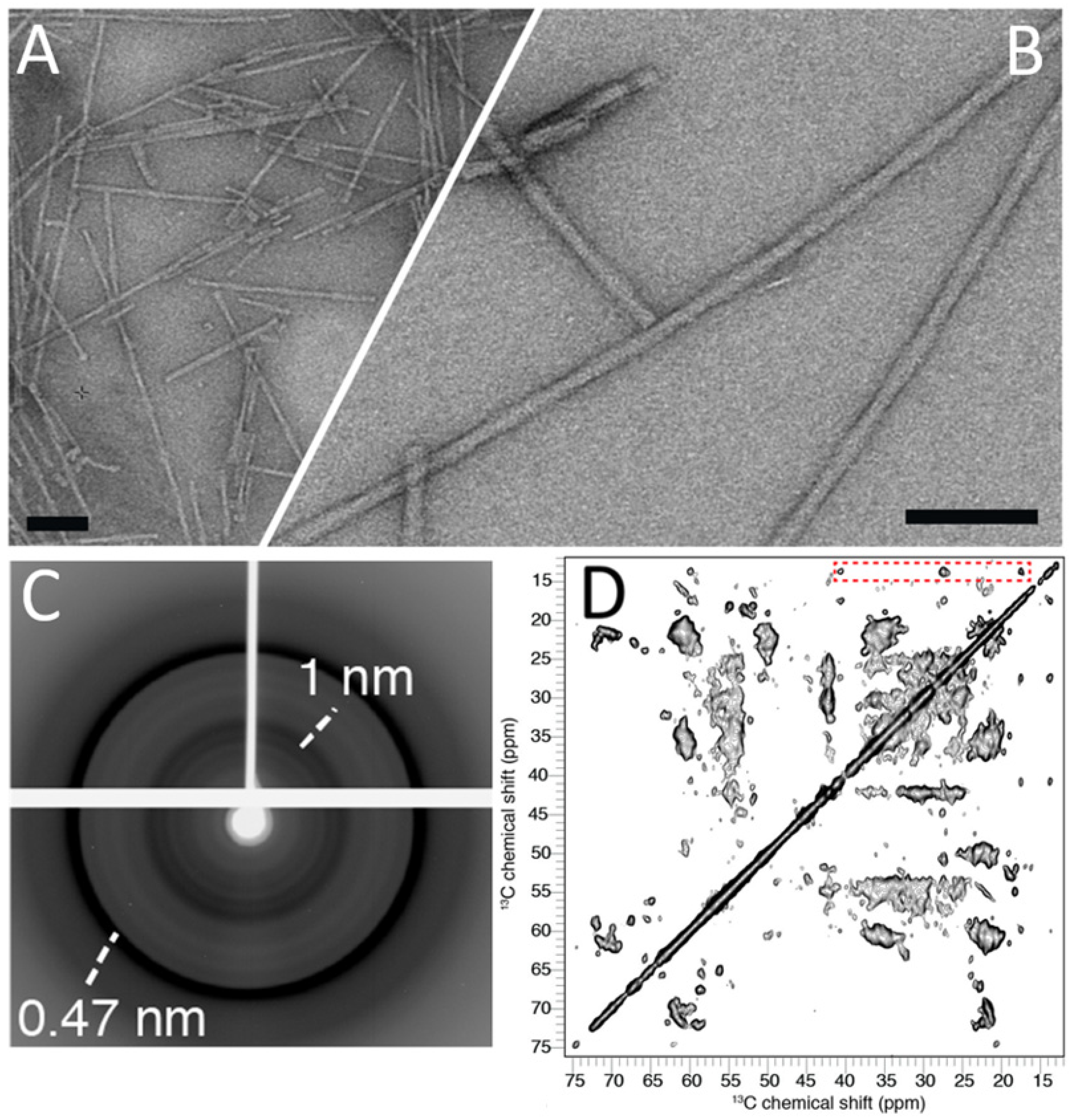
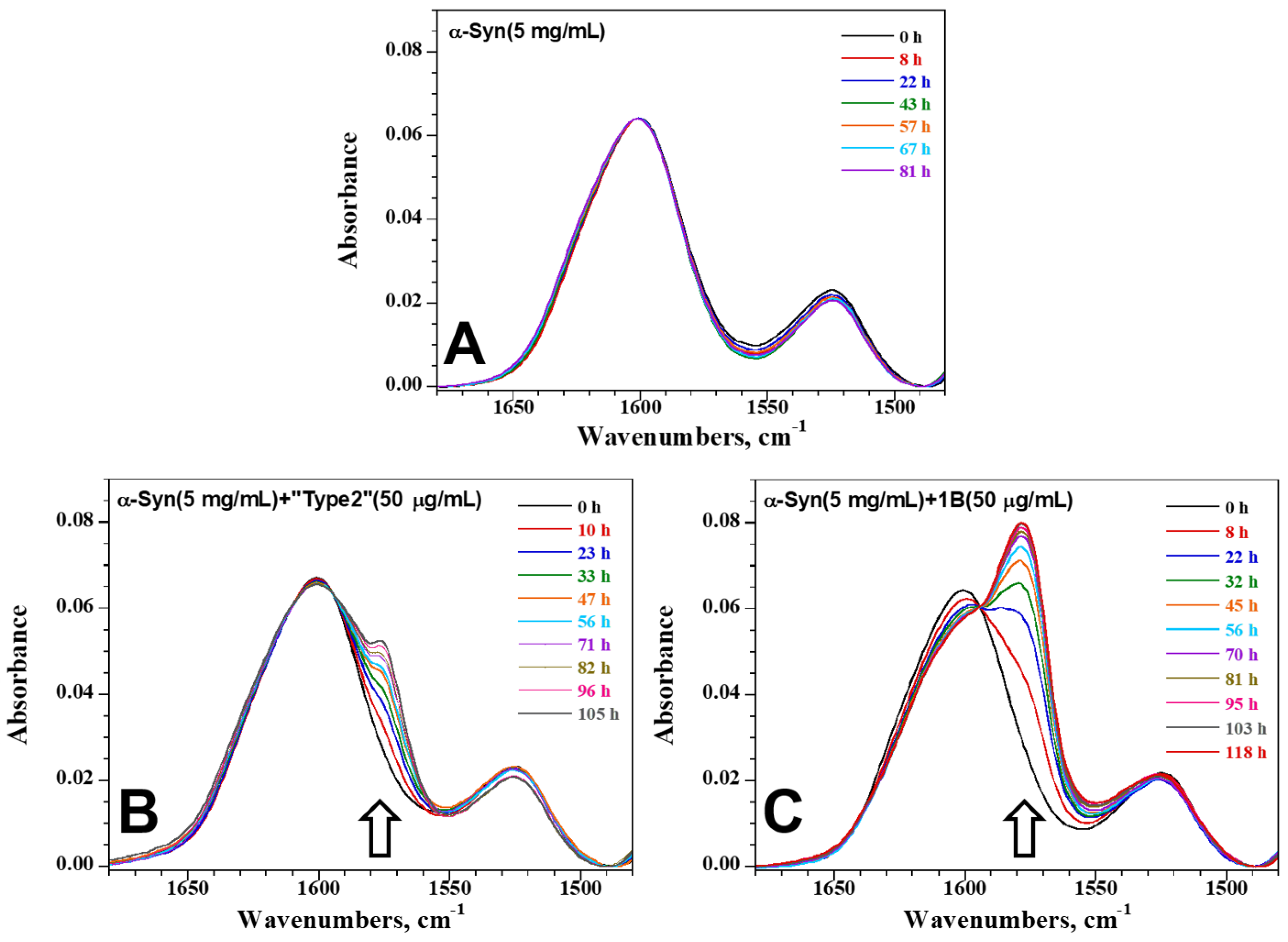
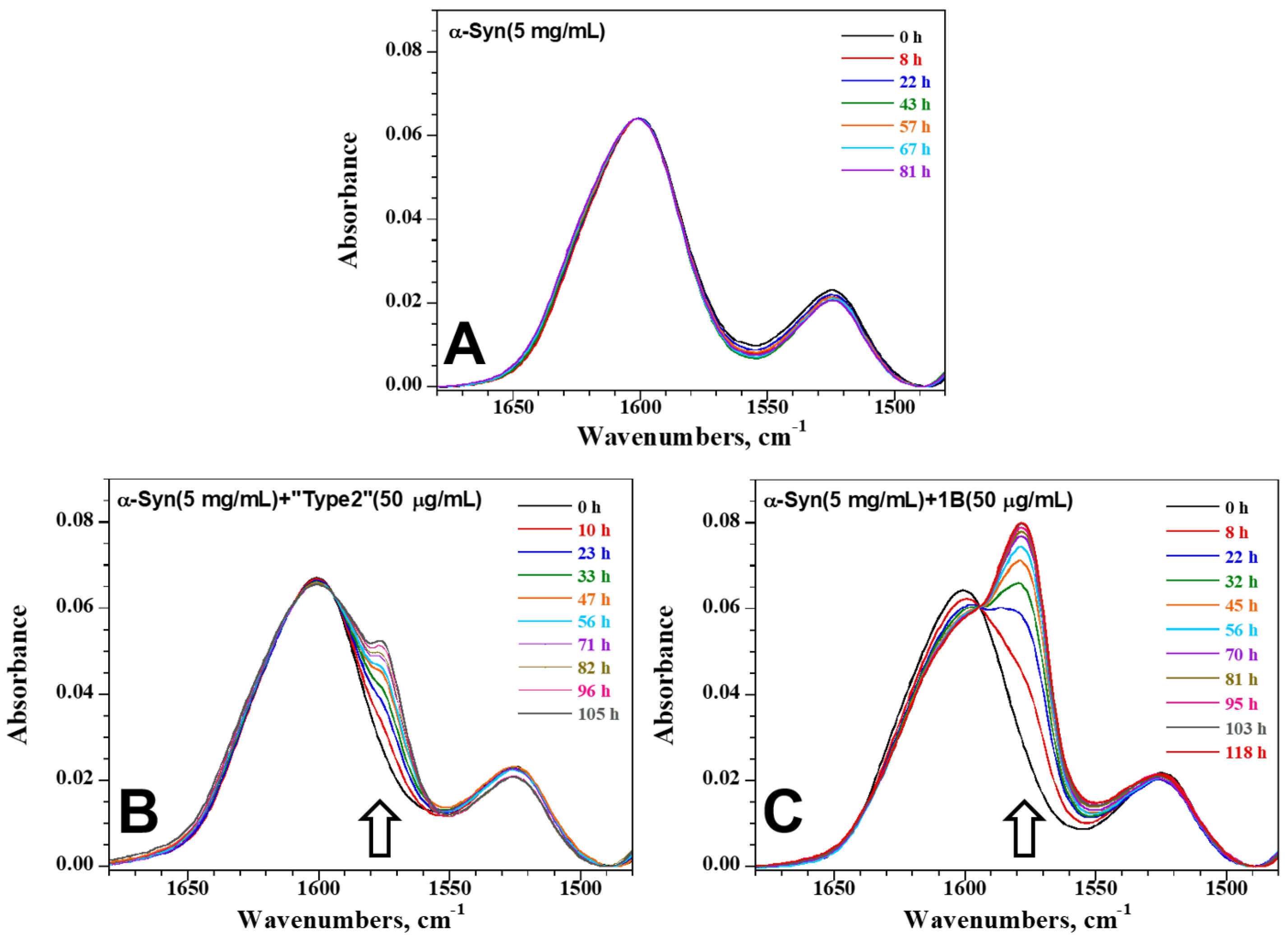
3.1. Biophysical Characterization of the Synthetic Human α-Syn Fibril Strain 1B
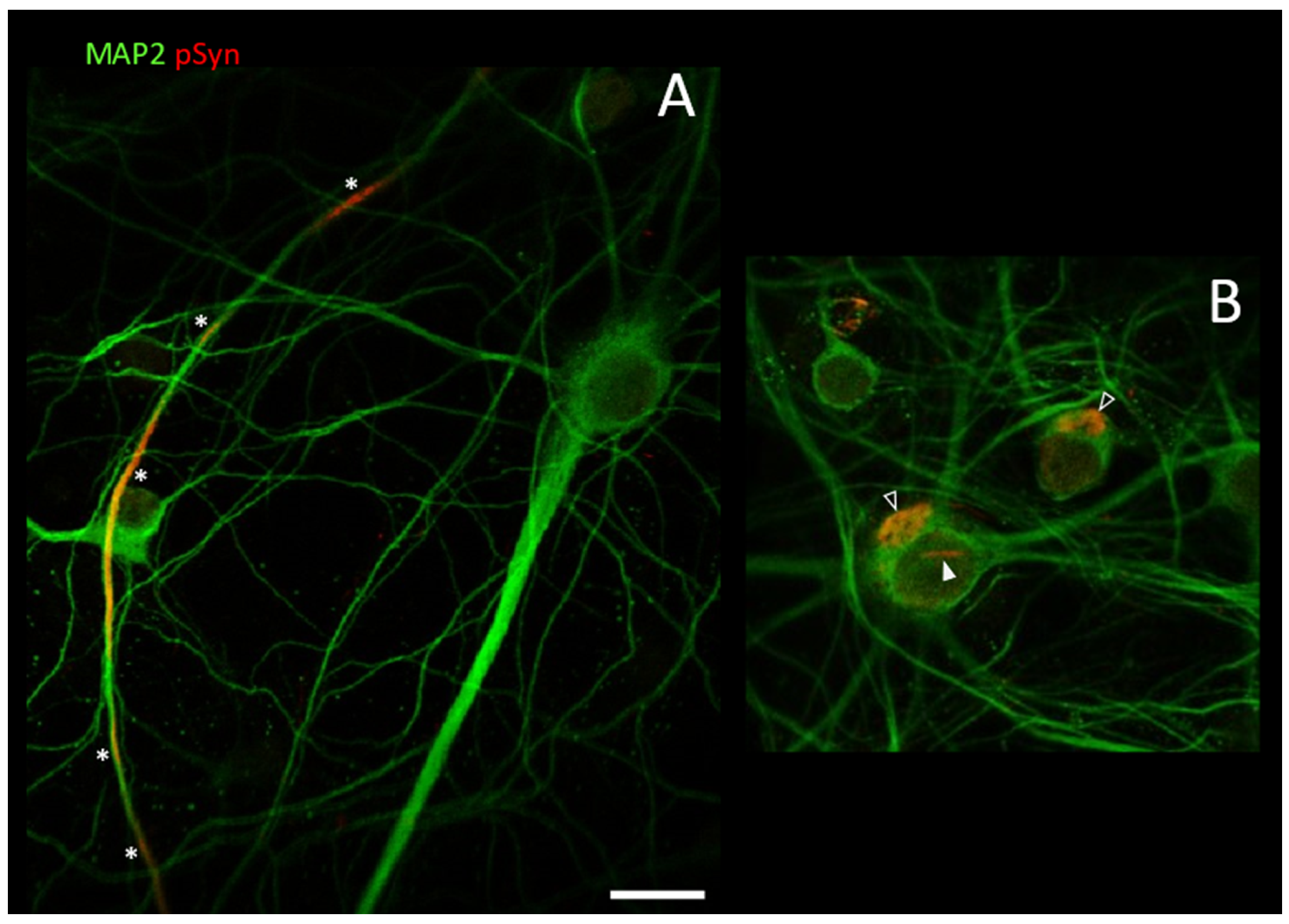
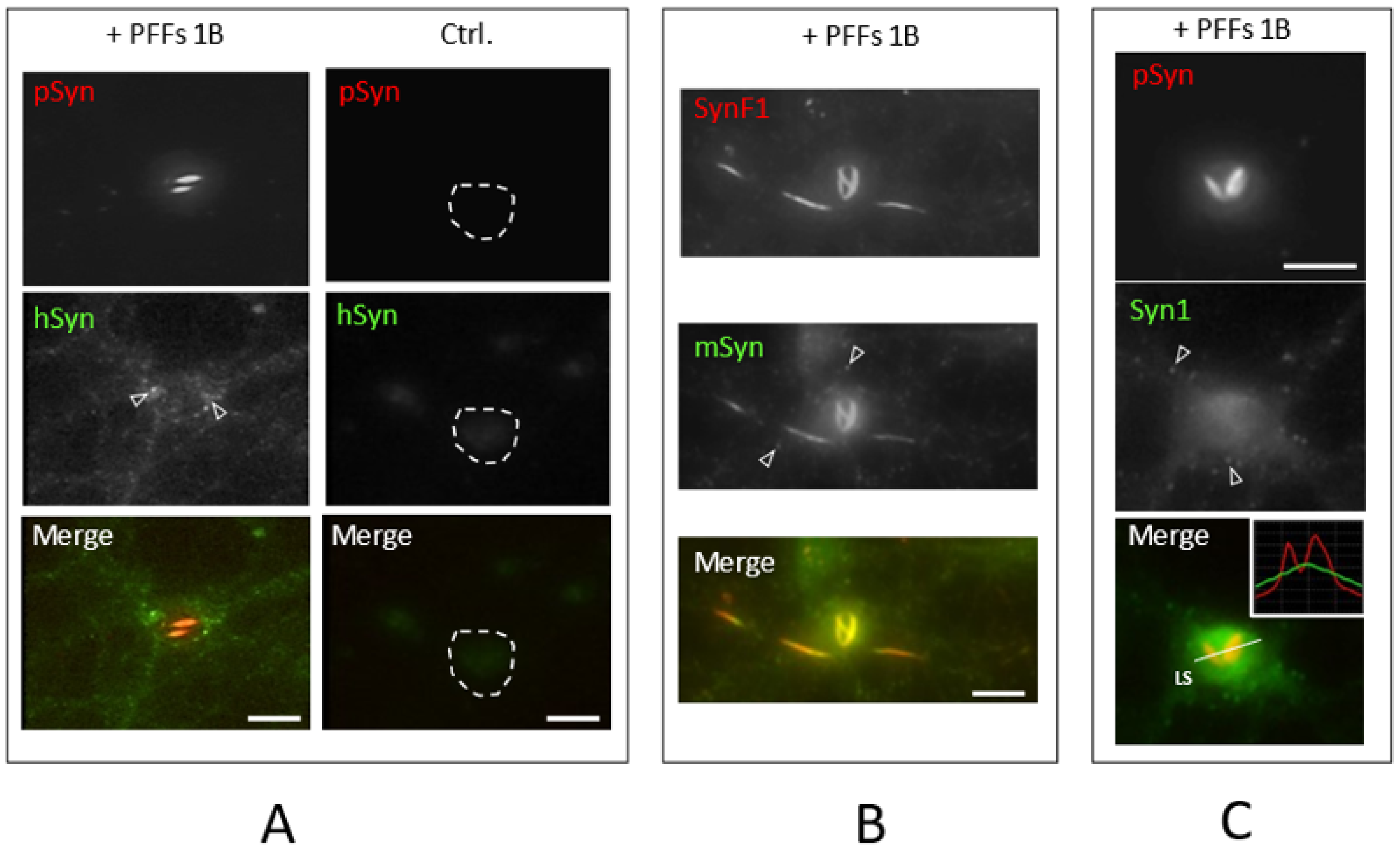
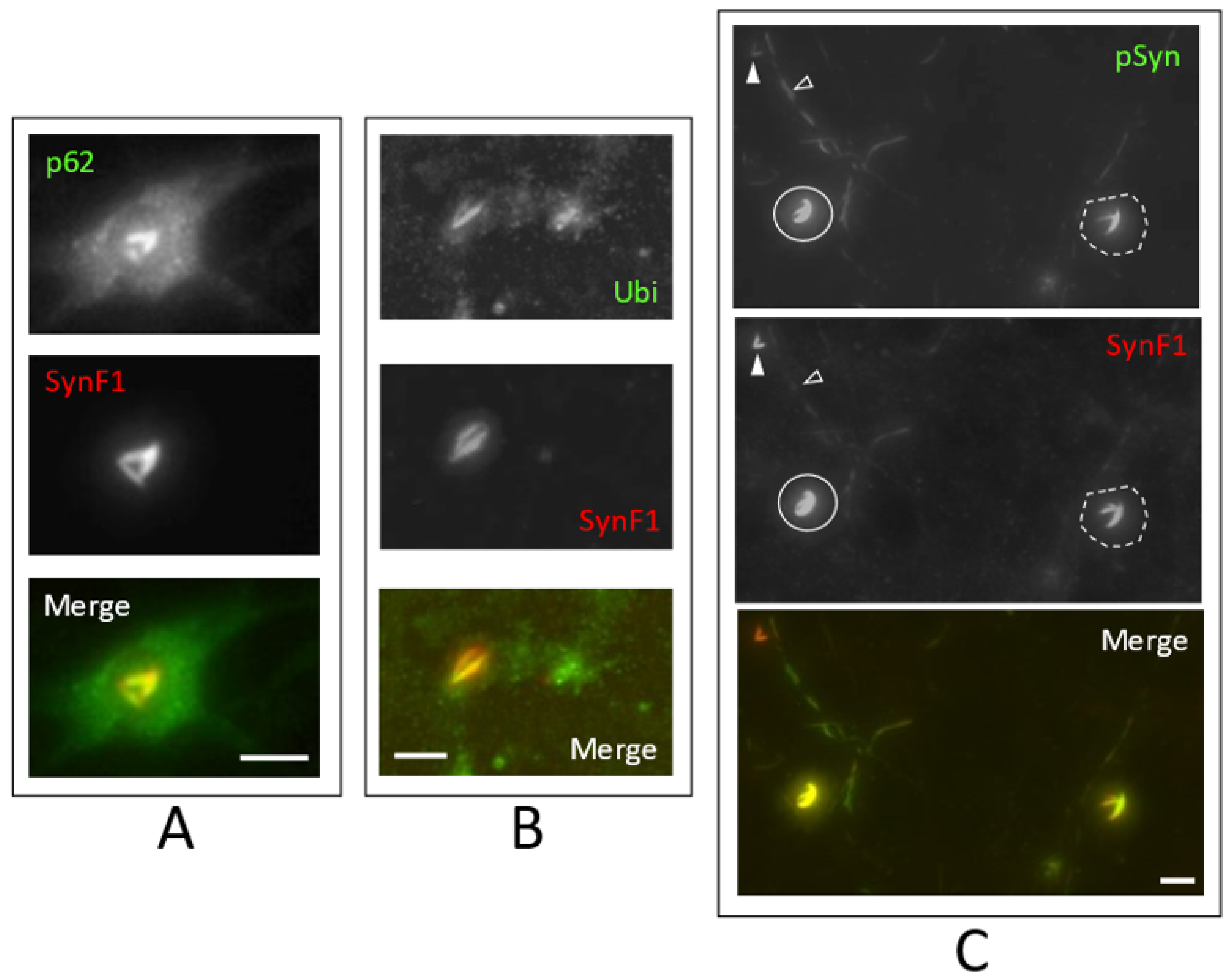
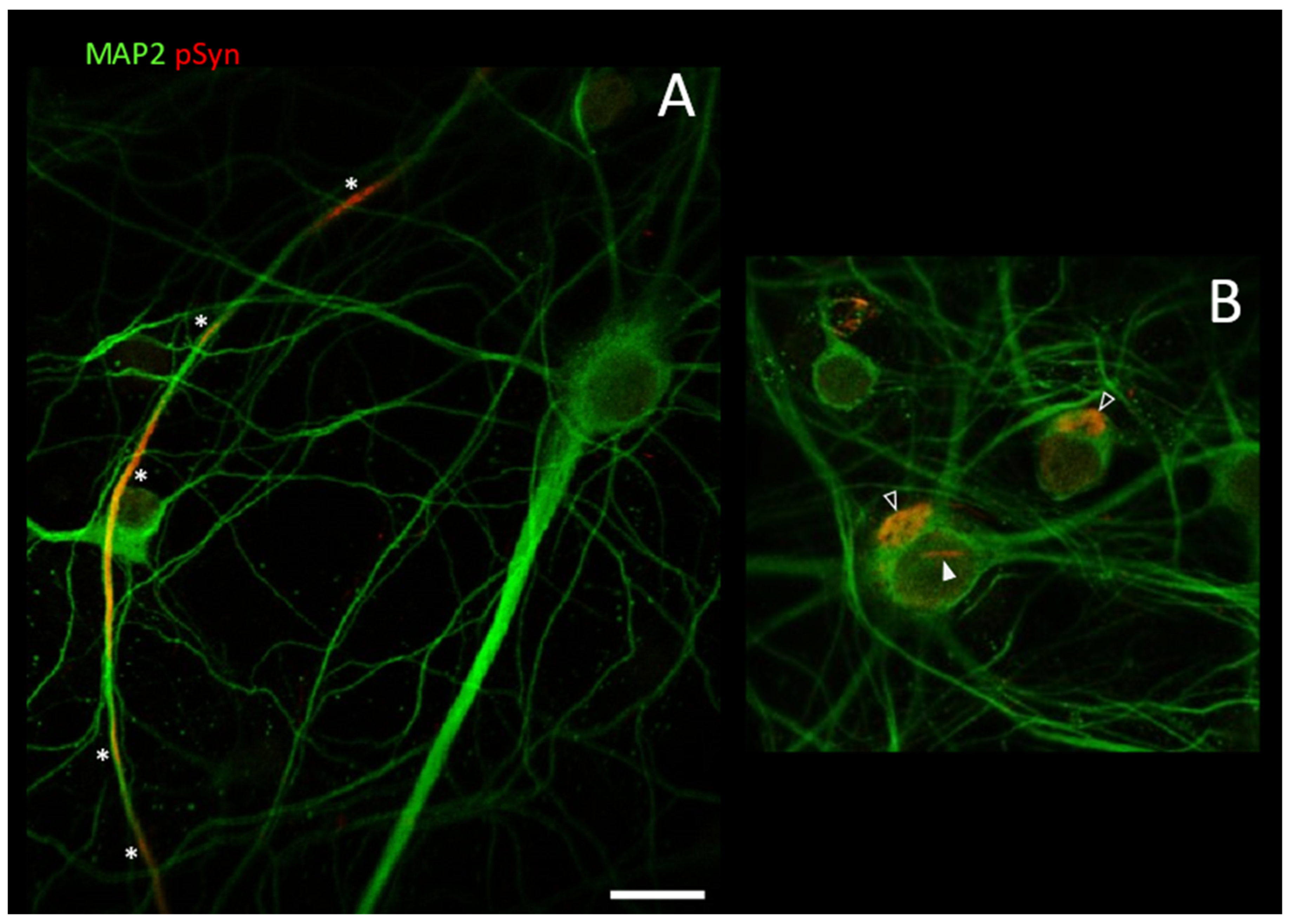
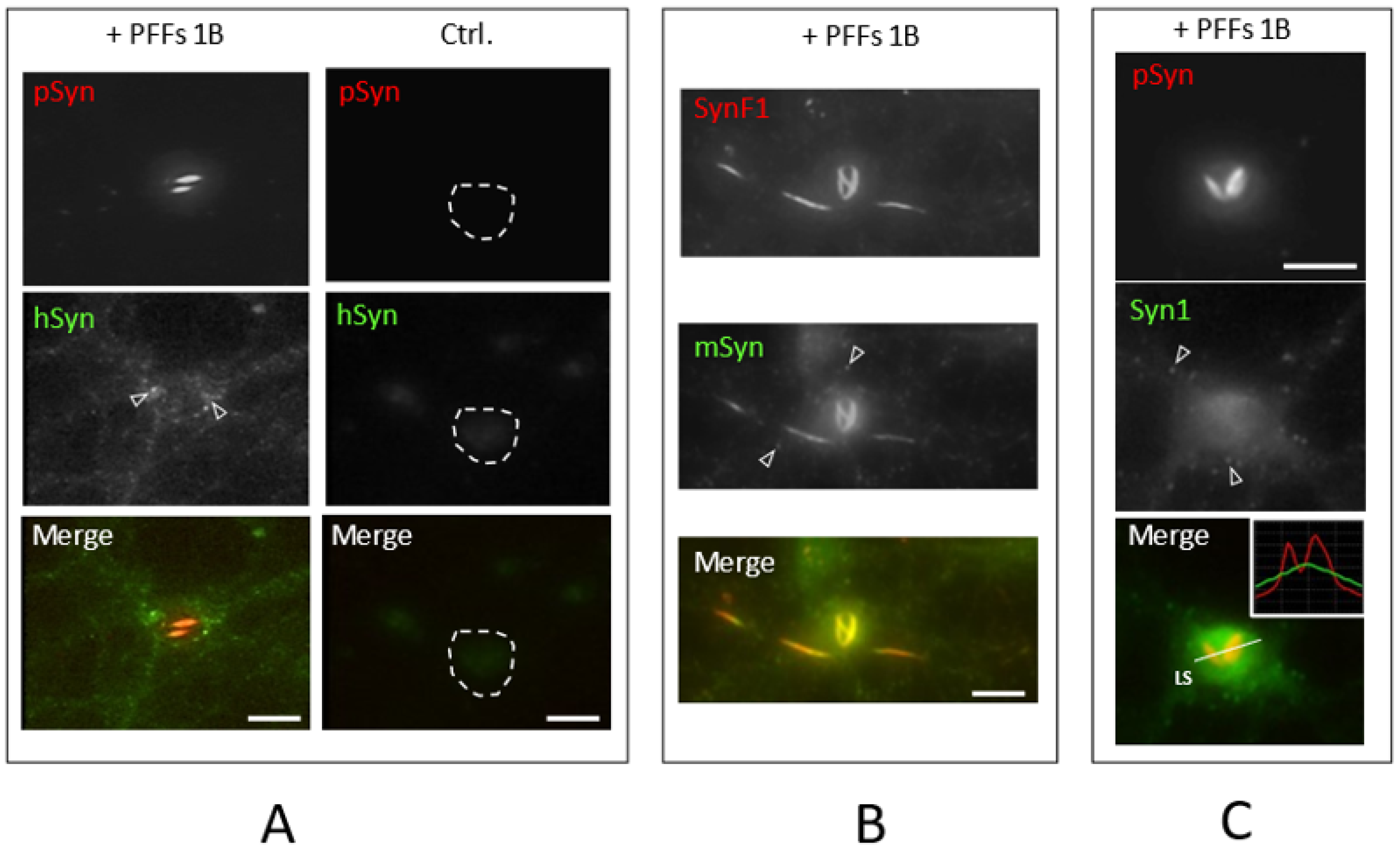
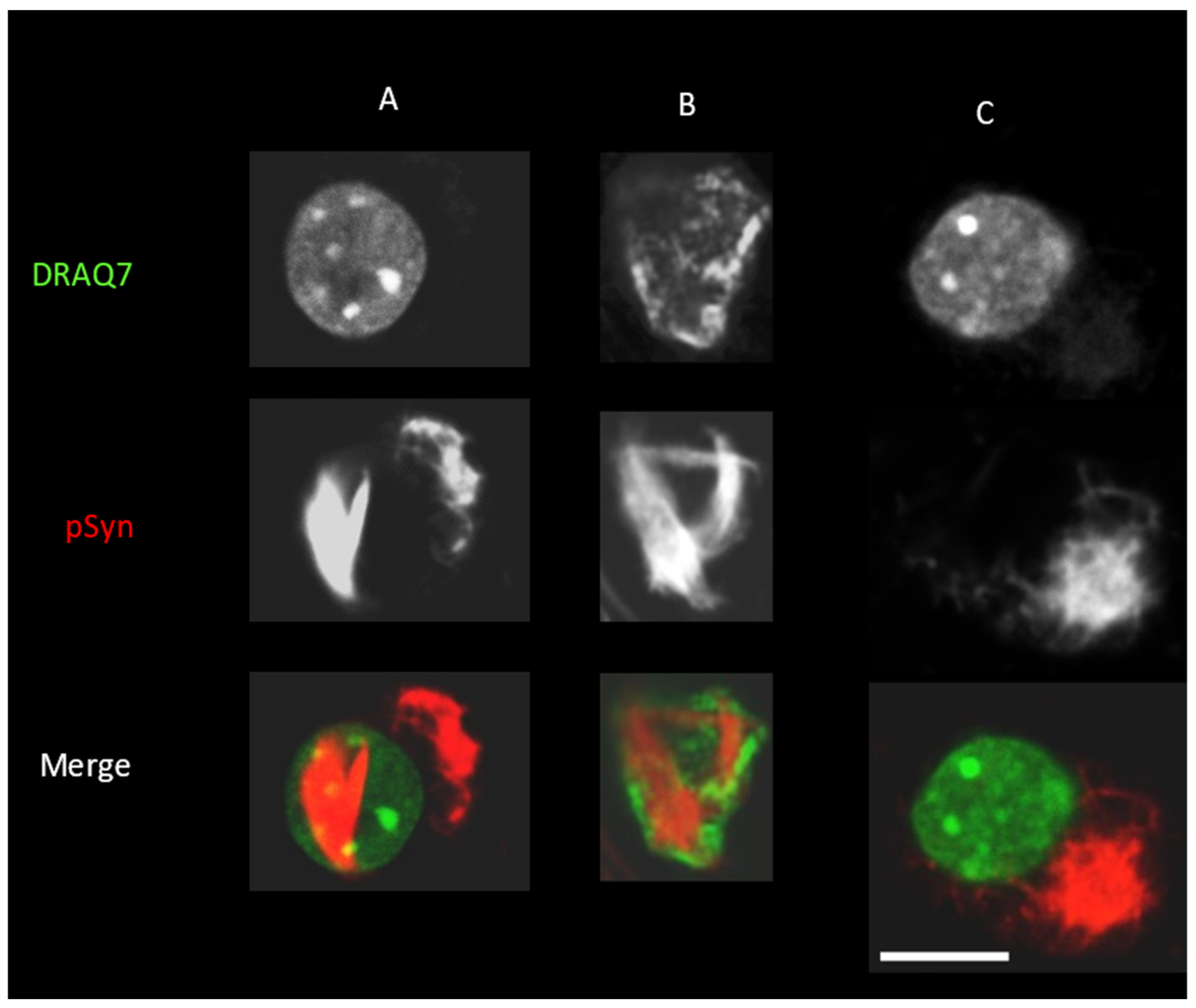
3.2. Neuronal Inclusions Seeded by 1B α-Syn Fibrils
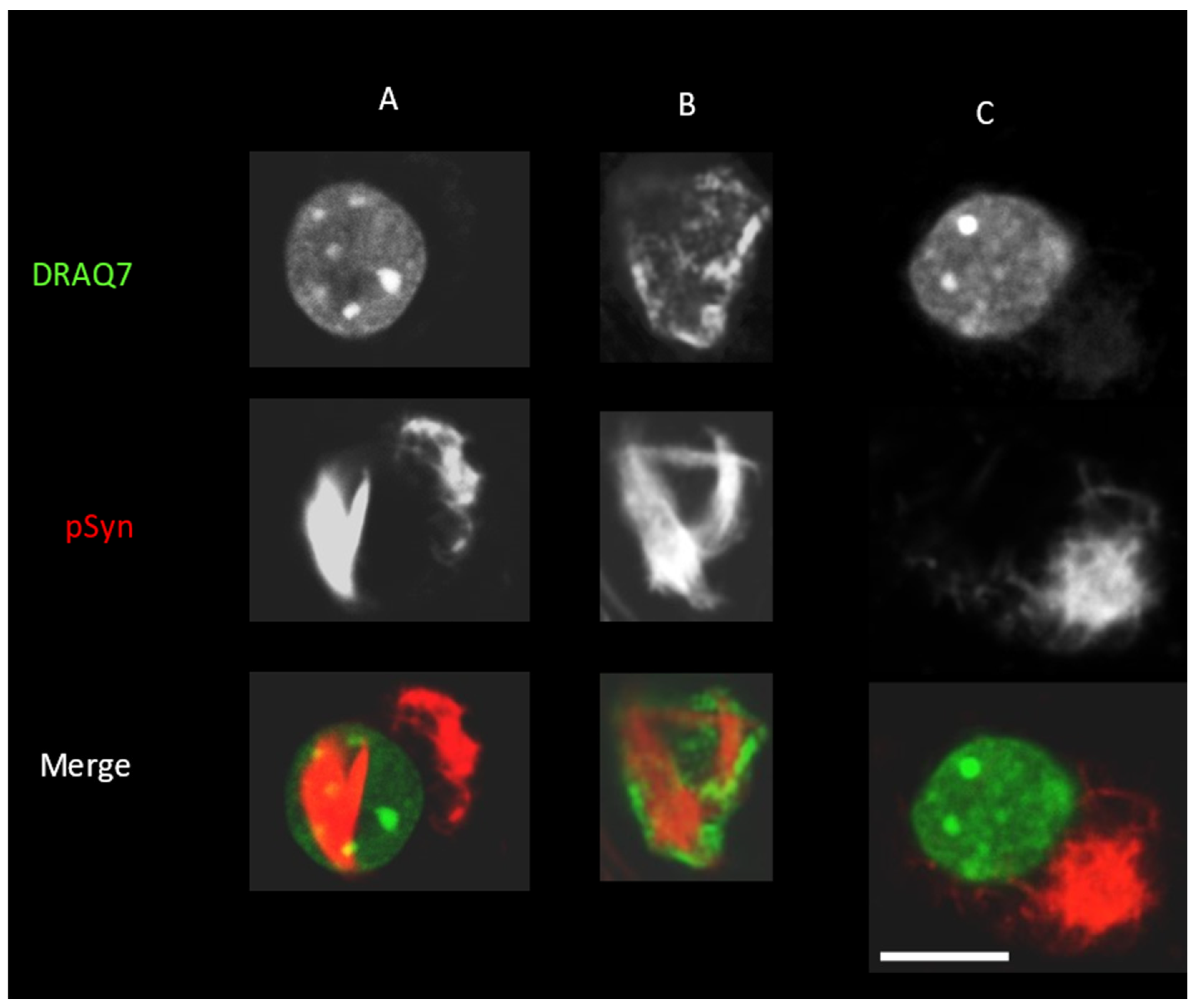
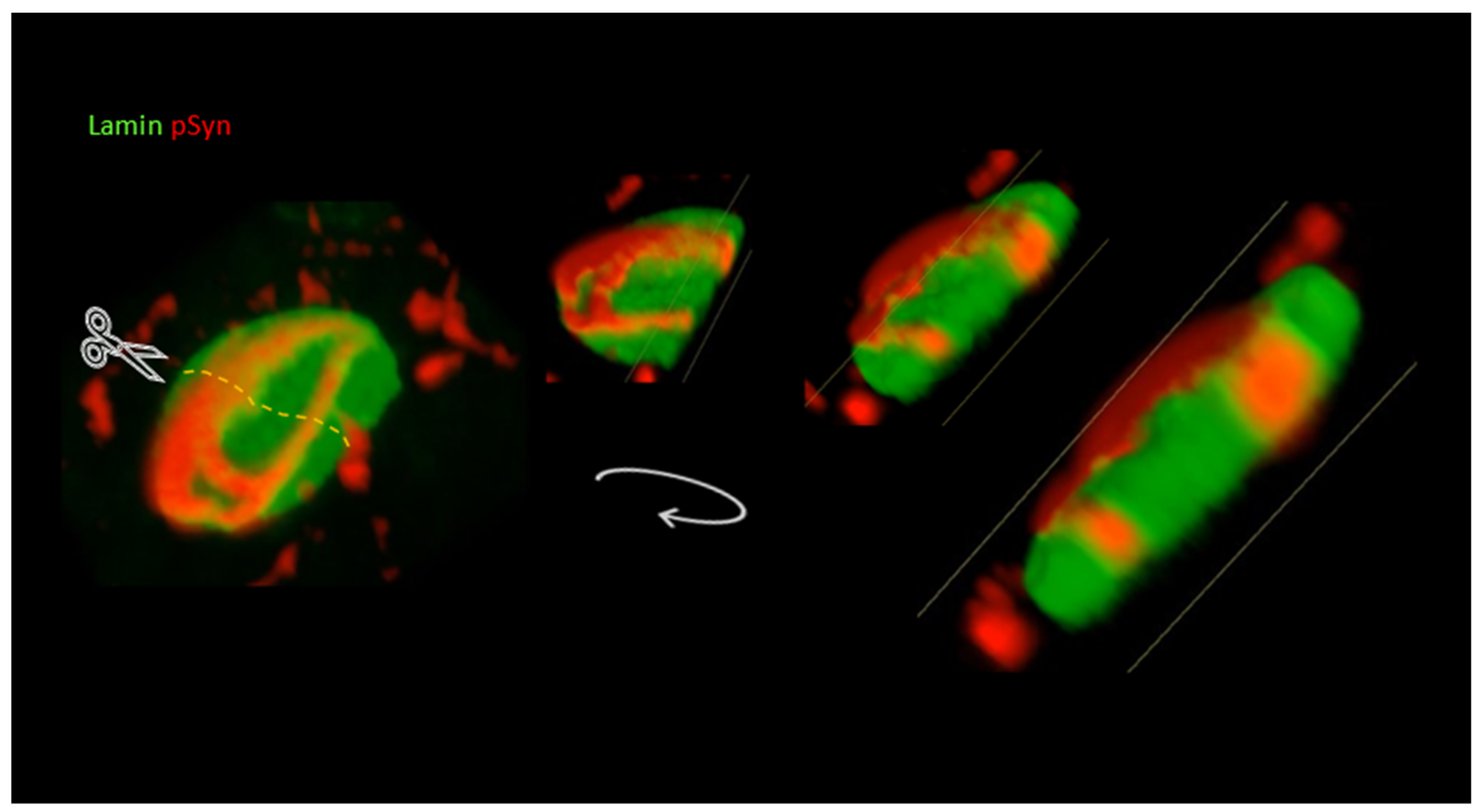
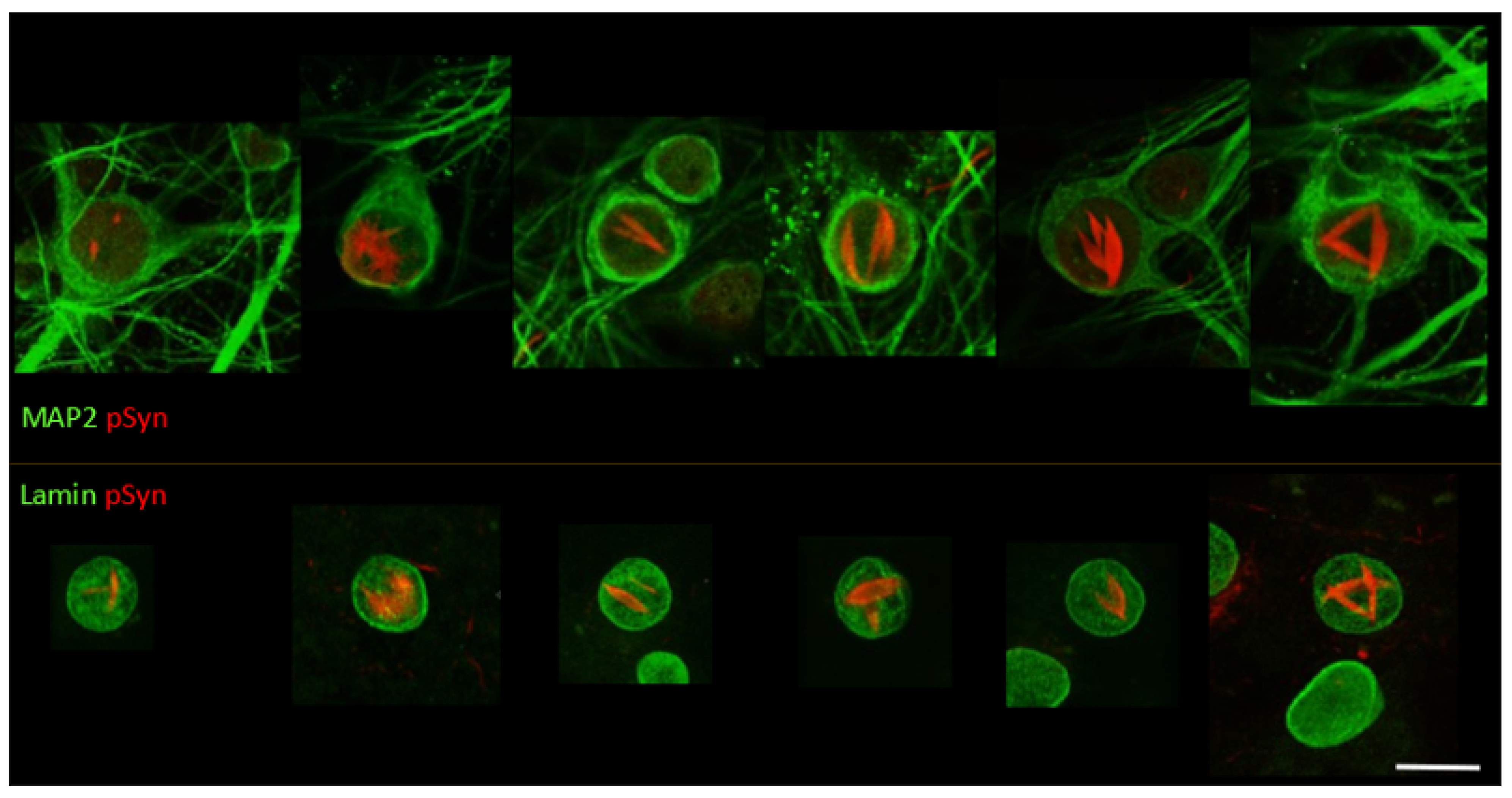
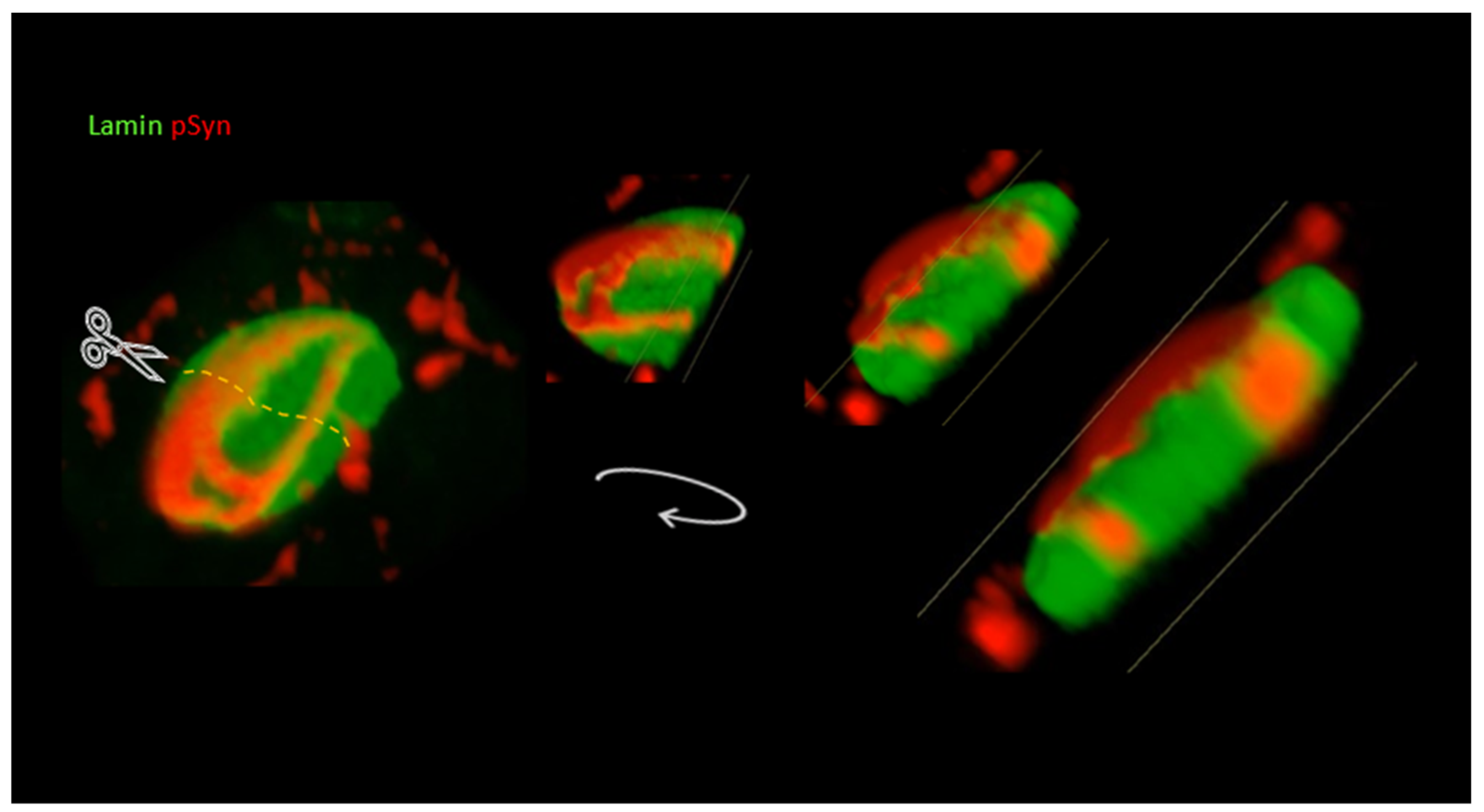
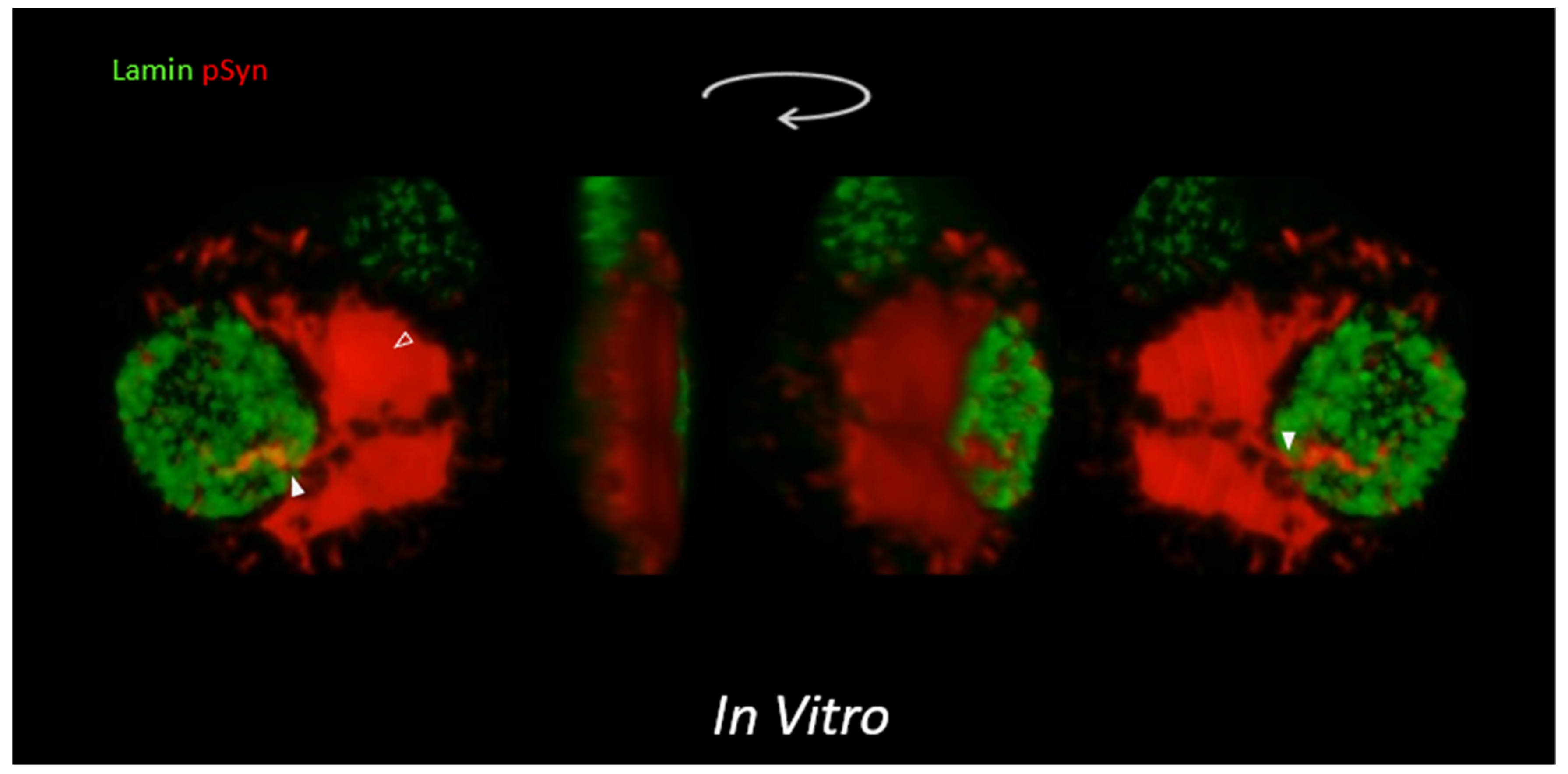
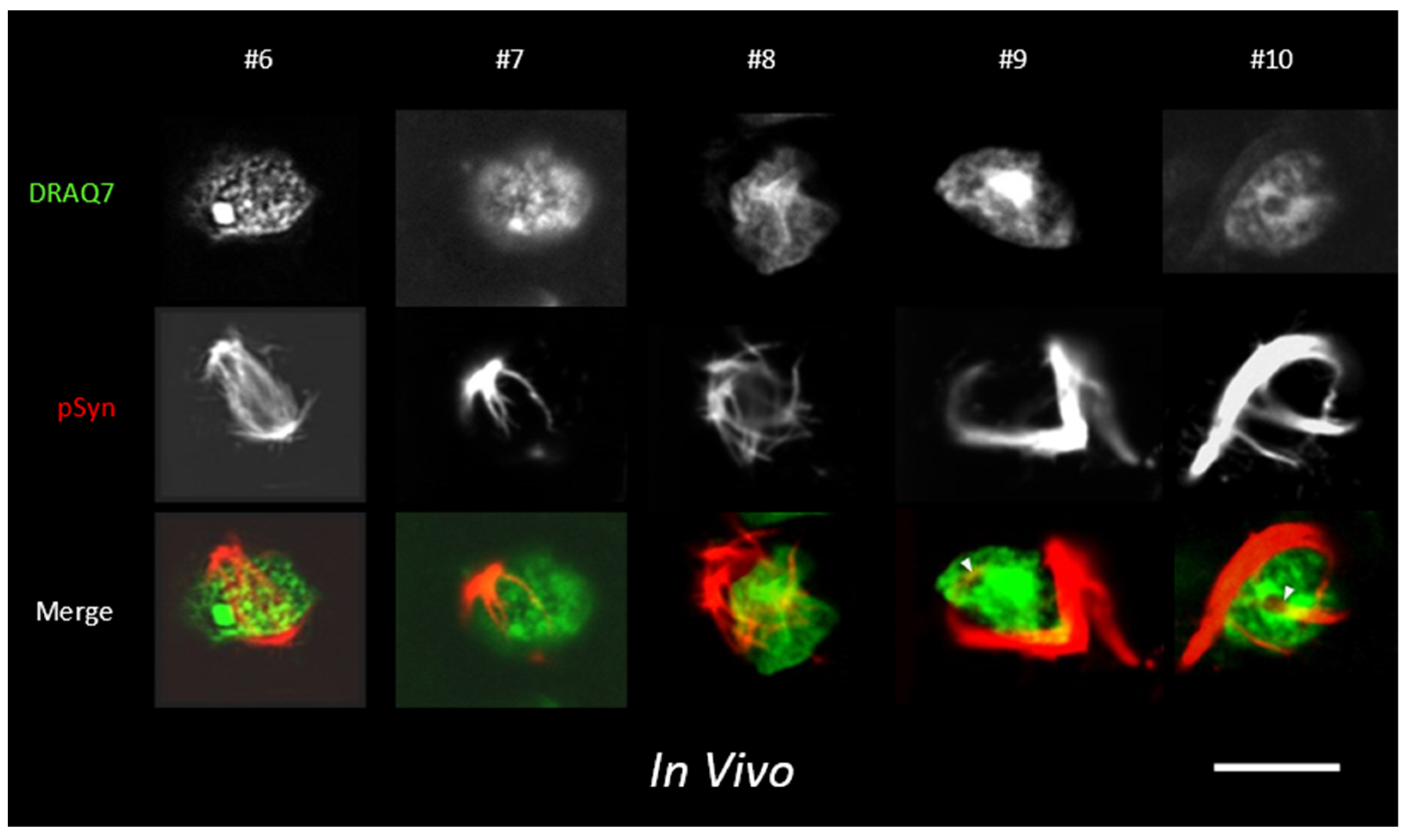
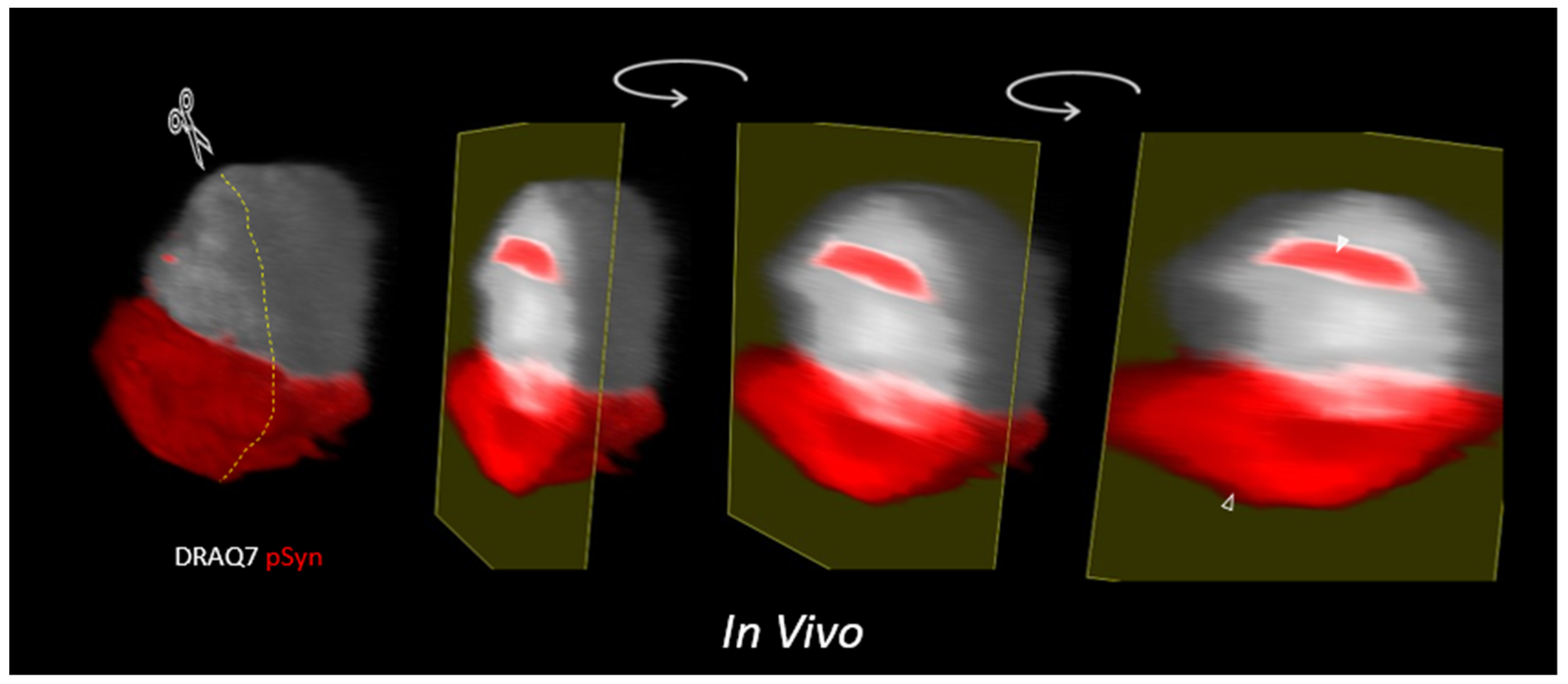
3.3. NIIs Seeded by 1B
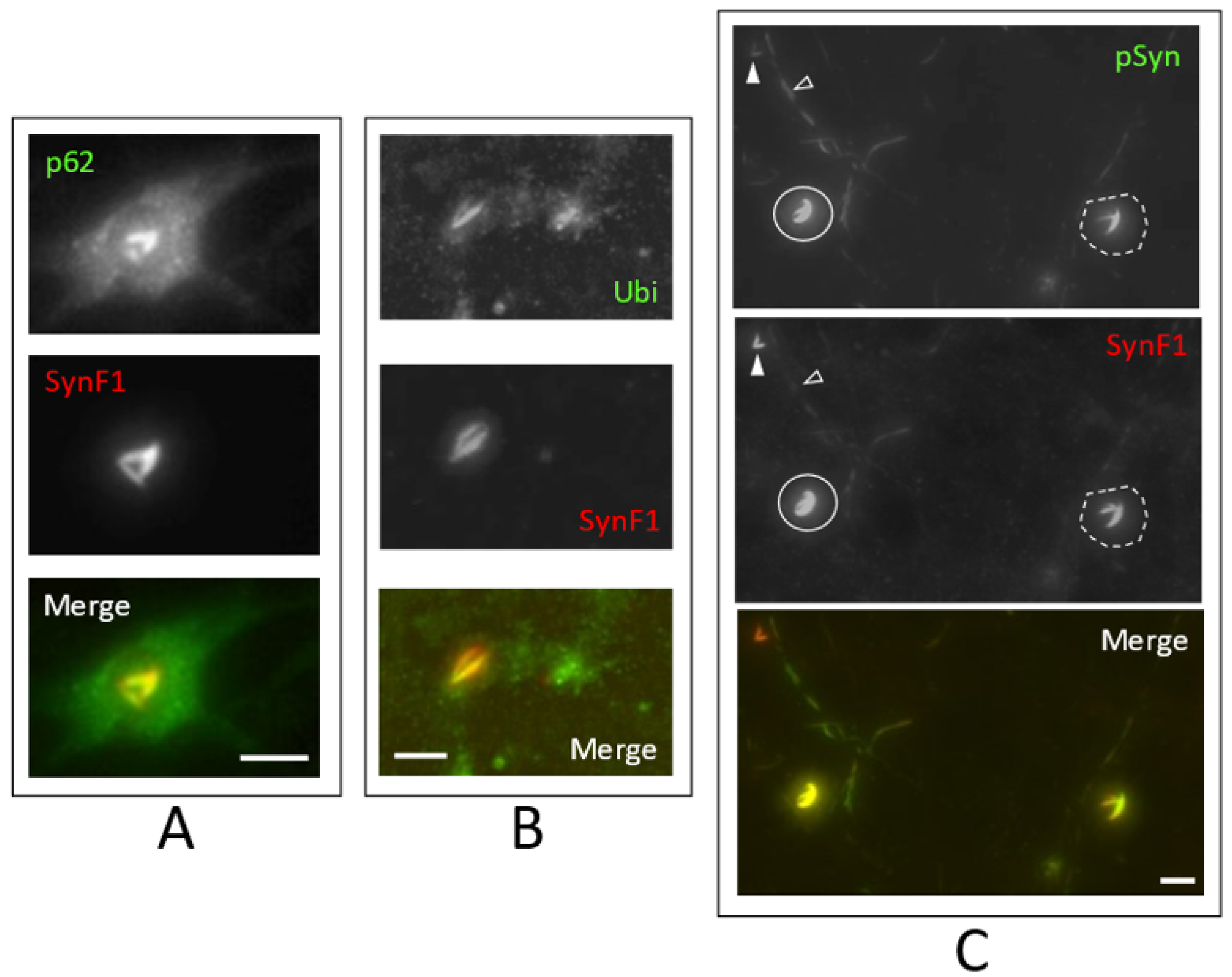
3.4. NIIs Are Amyloid Assemblies of Endogenous α-Syn
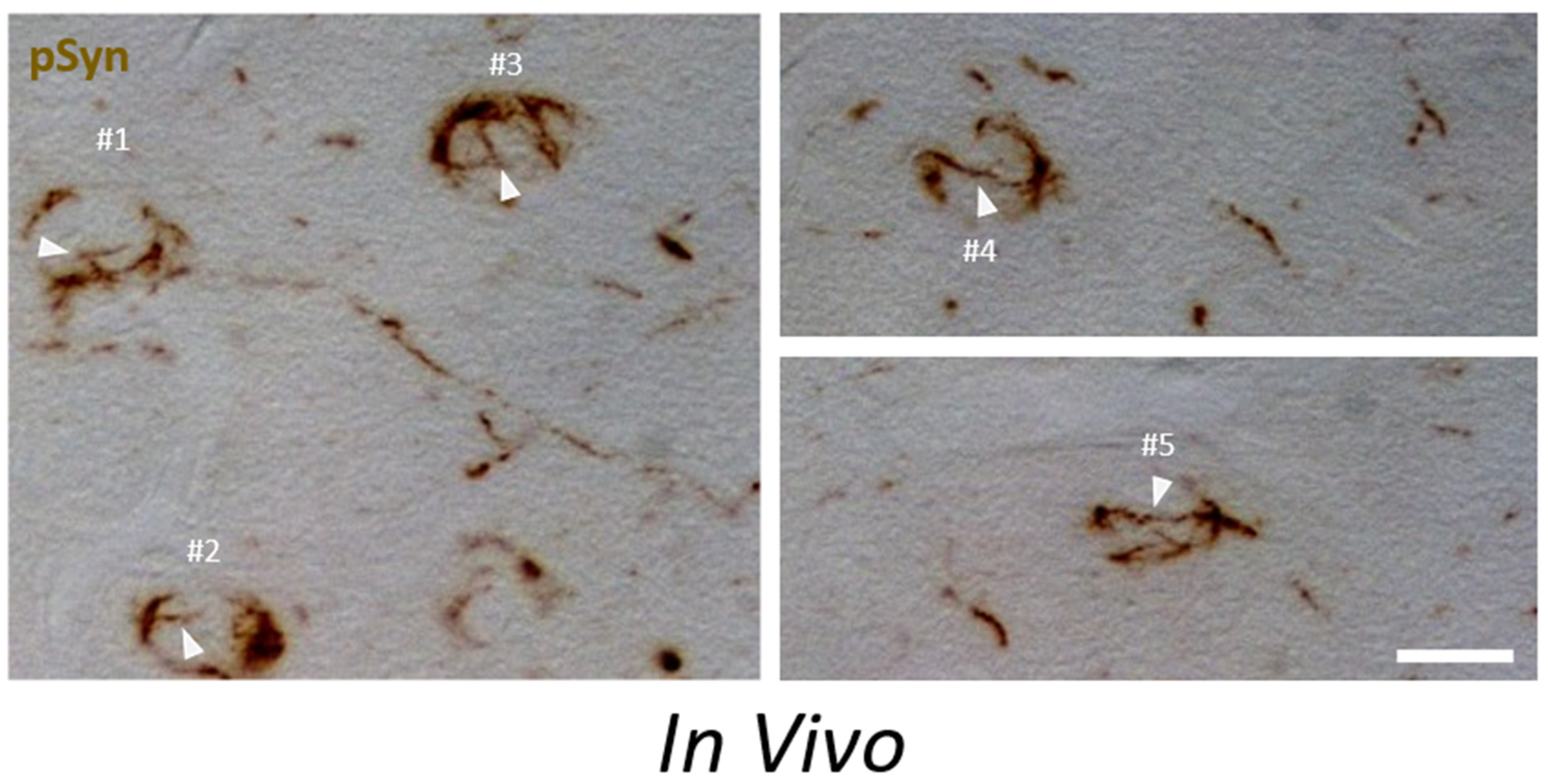
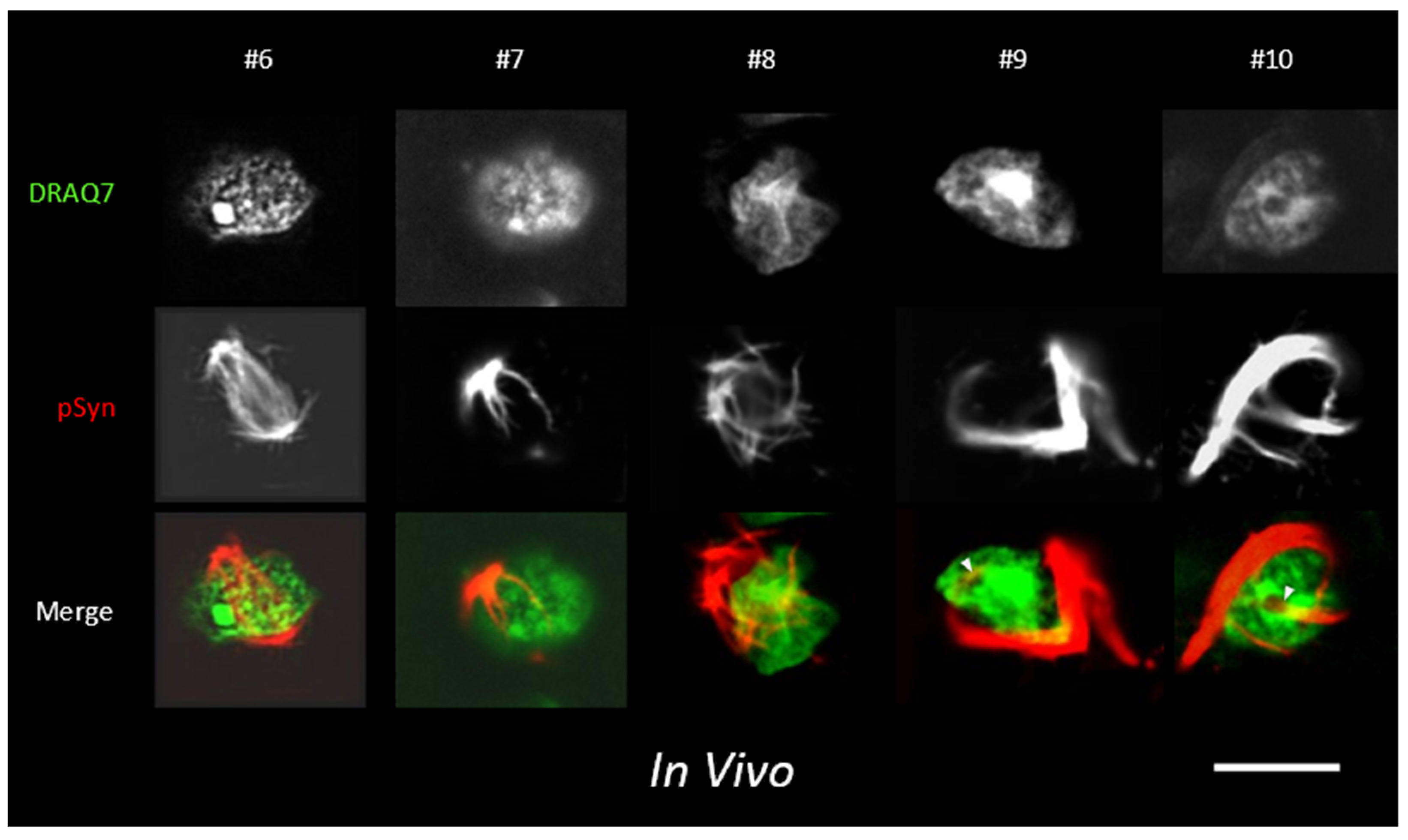
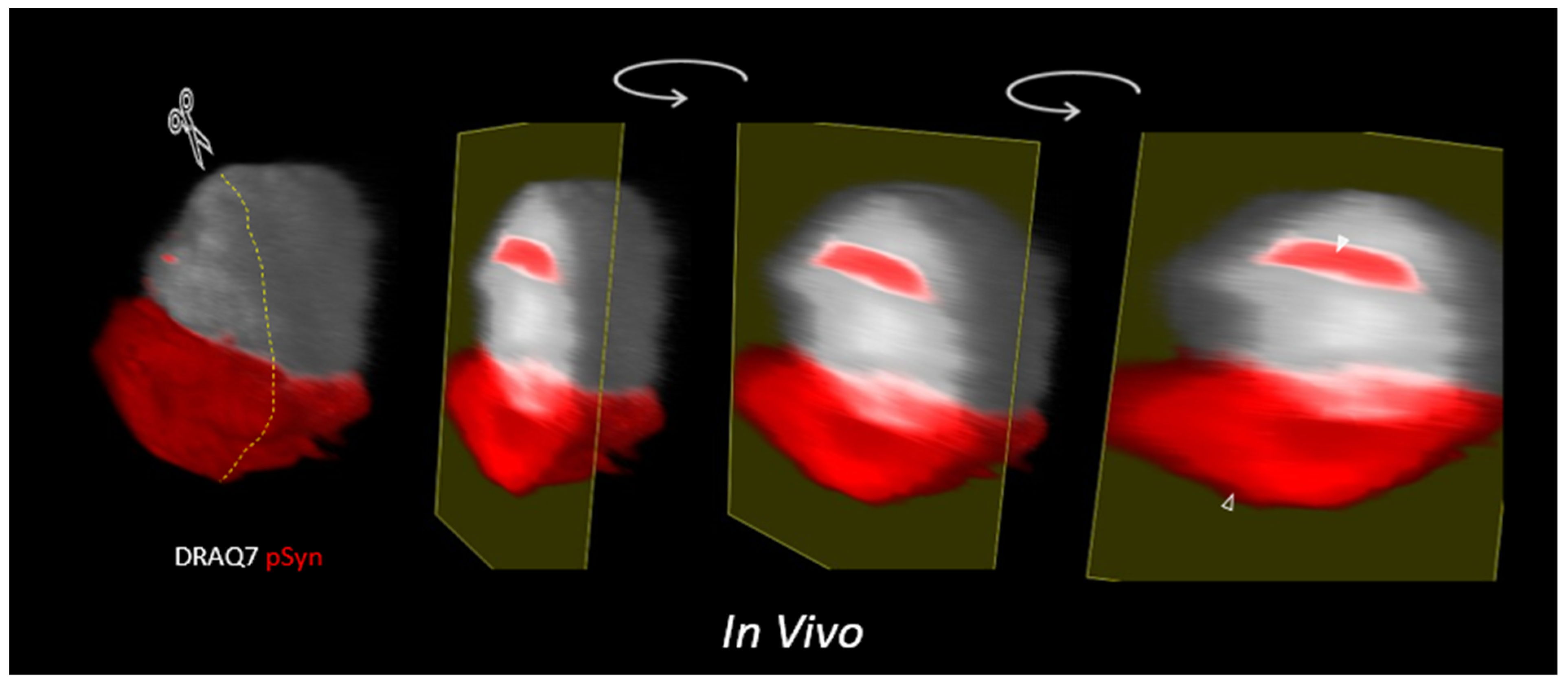
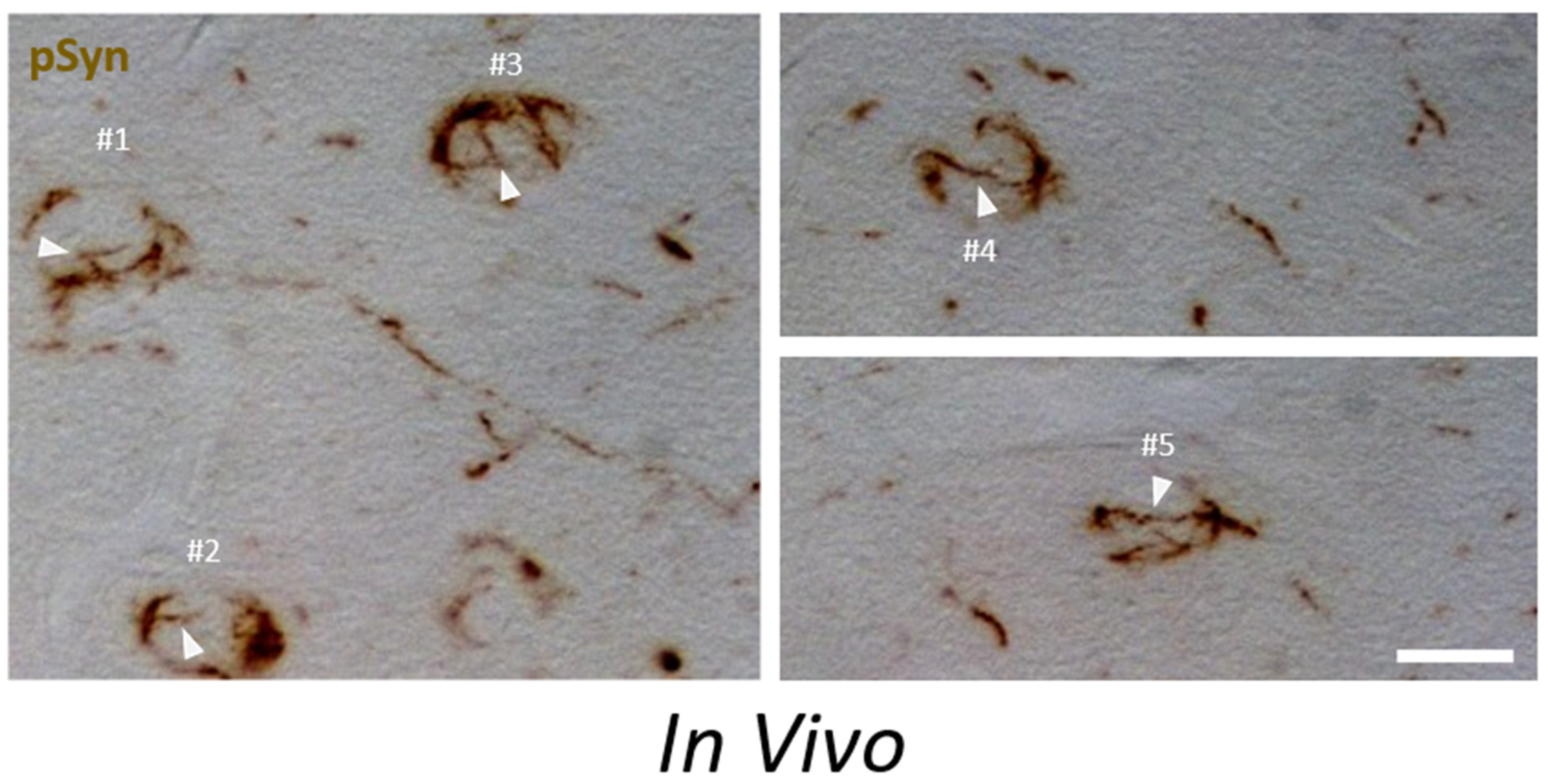
3.5. α-Syn NIIs Seeded In Vivo in Mice after Stereotaxic Injections of 1B Fibrils
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galvin, J.E.; Lee, V.M.Y.; Trojanowski, J.Q. Synucleinopathies: Clinical and Pathological Implications. Arch. Neurol. 2001, 58, 186–190. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Goedert, M. The α-Synucleinopathies: Parkinson’s Disease, Dementia with Lewy Bodies, and Multiple System Atrophy. Ann. N. Y. Acad. Sci. 2006, 920, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Espay, A.J. Movement Disorders Research in 2021: Cracking the Paradigm. Lancet. Neurol. 2022, 21, 10–11. [Google Scholar] [CrossRef]
- Espay, A.J. Our Winter Narrative—If Neurologists Had Discovered Snow: Water Monomers Misfold, Aggregated Ice Oligomers Replicate, Spread of Certain Ice Strains Cause Worse Winters, Anti-Ice Aggregation Strategies Will Restore Summer. Available online: https://twitter.com/AlbertoEspay/status/1490342015388864514?t=2H7j5GVZHxaIB21UM2KlDw&s=03 (accessed on 8 February 2022).
- Han, H.; Weinreb, P.H.; Lansbury, P.T. The Core Alzheimer’s Peptide NAC Forms Amyloid Fibrils Which Seed and Are Seeded by Beta-Amyloid: Is NAC a Common Trigger or Target in Neurodegenerative Disease? Chem. Biol. 1995, 2, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Hardy, J. Expression of Normal Sequence Pathogenic Proteins for Neurodegenerative Disease Contributes to Disease Risk: “permissive Templating” as a General Mechanism Underlying Neurodegeneration. Biochem. Soc. Trans. 2005, 33, 578–581. [Google Scholar] [CrossRef] [PubMed]
- Lansbury, P.T.; Caughey, B. The Chemistry of Scrapie Infection: Implications of the “ice 9” Metaphor. Chem. Biol. 1995, 2, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Luk, K.C.; Song, C.; O’Brien, P.; Stieber, A.; Branch, J.R.; Brunden, K.R.; Trojanowski, J.Q.; Lee, V.M.Y. Exogenous Alpha-Synuclein Fibrils Seed the Formation of Lewy Body-like Intracellular Inclusions in Cultured Cells. Proc. Natl. Acad. Sci. USA 2009, 106, 20051–20056. [Google Scholar] [CrossRef] [Green Version]
- Desplats, P.; Lee, H.J.; Bae, E.J.; Patrick, C.; Rockenstein, E.; Crews, L.; Spencer, B.; Masliah, E.; Lee, S.J. Inclusion Formation and Neuronal Cell Death through Neuron-to-Neuron Transmission of Alpha-Synuclein. Proc. Natl. Acad. Sci. USA 2009, 106, 13010–13015. [Google Scholar] [CrossRef] [Green Version]
- Volpicelli-Daley, L.A.; Luk, K.C.; Patel, T.P.; Tanik, S.A.; Riddle, D.M.; Stieber, A.; Meaney, D.F.; Trojanowski, J.Q.; Lee, V.M.Y. Exogenous α-Synuclein Fibrils Induce Lewy Body Pathology Leading to Synaptic Dysfunction and Neuron Death. Neuron 2011, 72, 57–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luk, K.C.; Kehm, V.; Carroll, J.; Zhang, B.; O’Brien, P.; Trojanowski, J.Q.; Lee, V.M.Y. Pathological α-Synuclein Transmission Initiates Parkinson-like Neurodegeneration in Nontransgenic Mice. Science 2012, 338, 949–953. [Google Scholar] [CrossRef] [Green Version]
- Mahul-Mellier, A.L.; Burtscher, J.; Maharjan, N.; Weerens, L.; Croisier, M.; Kuttler, F.; Leleu, M.; Knott, G.W.; Lashuel, H.A. The Process of Lewy Body Formation, Rather than Simply α-Synuclein Fibrillization, Is One of the Major Drivers of Neurodegeneration. Proc. Natl. Acad. Sci. USA 2020, 117, 4971–4982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, S.J.; Wypych, J.; Steavenson, S.; Louis, J.C.; Citron, M.; Biere, A.L. α-Synuclein Fibrillogenesis Is Nucleation-Dependent: Implications for the pathogenesis of Parkinson’S disease *. J. Biol. Chem. 1999, 274, 19509–19512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda-Suzukake, M.; Nonaka, T.; Hosokawa, M.; Oikawa, T.; Arai, T.; Akiyama, H.; Mann, D.M.A.; Hasegawa, M. Prion-like Spreading of Pathological α-Synuclein in Brain. Brain J. Neurol. 2013, 136, 1128–1138. [Google Scholar] [CrossRef] [PubMed]
- De Giorgi, F.; Laferrière, F.; Zinghirino, F.; Faggiani, E.; Lends, A.; Bertoni, M.; Yu, X.; Grélard, A.; Morvan, E.; Habenstein, B.; et al. Novel Self-Replicating α-Synuclein Polymorphs That Escape ThT Monitoring Can Spontaneously Emerge and Acutely Spread in Neurons. Sci. Adv. 2020, 6, eabc4364. [Google Scholar] [CrossRef] [PubMed]
- Sacino, A.N.; Thomas, M.A.; Ceballos-Diaz, C.; Cruz, P.E.; Rosario, A.M.; Lewis, J.; Giasson, B.I.; Golde, T.E. Conformational Templating of α-Synuclein Aggregates in Neuronal-Glial Cultures. Mol. Neurodegener. 2013, 8, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kordower, J.H.; Chu, Y.; Hauser, R.A.; Freeman, T.B.; Olanow, C.W. Lewy Body-like Pathology in Long-Term Embryonic Nigral Transplants in Parkinson’s Disease. Nat. Med. 2008, 14, 504–506. [Google Scholar] [CrossRef]
- Braak, H.; del Tredici, K.; Rüb, U.; de Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of Brain Pathology Related to Sporadic Parkinson’s Disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Li, J.Y.; Englund, E.; Holton, J.L.; Soulet, D.; Hagell, P.; Lees, A.J.; Lashley, T.; Quinn, N.P.; Rehncrona, S.; Björklund, A.; et al. Lewy Bodies in Grafted Neurons in Subjects with Parkinson’s Disease Suggest Host-to-Graft Disease Propagation. Nat. Med. 2008, 14, 501–503. [Google Scholar] [CrossRef]
- Greffard, S.; Verny, M.; Bonnet, A.M.; Seilhean, D.; Hauw, J.J.; Duyckaerts, C. A Stable Proportion of Lewy Body Bearing Neurons in the Substantia Nigra Suggests a Model in Which the Lewy Body Causes Neuronal Death. Neurobiol. Aging 2010, 31, 99–103. [Google Scholar] [CrossRef]
- Singleton, A.B.; Farrer, M.; Johnson, J.; Singleton, A.; Hague, S.; Kachergus, J.; Hulihan, M.; Peuralinna, T.; Dutra, A.; Nussbaum, R.; et al. Alpha-Synuclein Locus Triplication Causes Parkinson’s Disease. Science 2003, 302, 841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, A.; Myers, A.; Hardy, J. The Law of Mass Action Applied to Neurodegenerative Disease: A Hypothesis Concerning the Etiology and Pathogenesis of Complex Diseases. Hum. Mol. Genet. 2004, 13, R123–R126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bousset, L.; Pieri, L.; Ruiz-Arlandis, G.; Gath, J.; Jensen, P.H.; Habenstein, B.; Madiona, K.; Olieric, V.; Böckmann, A.; Meier, B.H.; et al. Structural and Functional Characterization of Two Alpha-Synuclein Strains. Nat. Commun. 2013, 4, 2575. [Google Scholar] [CrossRef] [Green Version]
- Barclay, A.M.; Dhavale, D.D.; Courtney, J.M.; Kotzbauer, P.T.; Rienstra, C.M. Resonance Assignments of an α-Synuclein Fibril Prepared in Tris Buffer at Moderate Ionic Strength. Biomol. NMR Assign. 2018, 12, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, M.D.; Comellas, G.; Nieuwkoop, A.J.; Covell, D.J.; Berthold, D.A.; Kloepper, K.D.; Courtney, J.M.; Kim, J.K.; Barclay, A.M.; Kendall, A.; et al. Solid-State NMR Structure of a Pathogenic Fibril of Full-Length Human α-Synuclein. Nat. Struct. Mol. Biol. 2016, 23, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Lövestam, S.; Schweighauser, M.; Matsubara, T.; Murayama, S.; Tomita, T.; Ando, T.; Hasegawa, K.; Yoshida, M.; Tarutani, A.; Hasegawa, M.; et al. Seeded Assembly in Vitro Does Not Replicate the Structures of α-Synuclein Filaments from Multiple System Atrophy. FEBS Open Bio. 2021, 11, 999–1013. [Google Scholar] [CrossRef] [PubMed]
- Schweighauser, M.; Shi, Y.; Tarutani, A.; Kametani, F.; Murzin, A.G.; Ghetti, B.; Matsubara, T.; Tomita, T.; Ando, T.; Hasegawa, K.; et al. Structures of α-Synuclein Filaments from Multiple System Atrophy. Nature 2020, 585, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Burger, D.; Fenyi, A.; Bousset, L.; Stahlberg, H.; Melki, R. Cryo-EM Structure of Alpha-Synuclein Fibrils Amplified by PMCA from PD and MSA Patient Brains. bioRxiv 2021. [Google Scholar] [CrossRef]
- Suzuki, G.; Imura, S.; Hosokawa, M.; Katsumata, R.; Nonaka, T.; Hisanaga, S.I.; Saeki, Y.; Hasegawa, M. α-Synuclein Strains That Cause Distinct Pathologies Differentially Inhibit Proteasome. eLife 2020, 9, e56825. [Google Scholar] [CrossRef]
- Shrivastava, A.N.; Bousset, L.; Renner, M.; Redeker, V.; Savistchenko, J.; Triller, A.; Melki, R. Differential Membrane Binding and Seeding of Distinct α-Synuclein Fibrillar Polymorphs. Biophys. J. 2020, 118, 1301–1320. [Google Scholar] [CrossRef] [Green Version]
- Pascuzzo, R.; Oxtoby, N.P.; Young, A.L.; Blevins, J.; Castelli, G.; Garbarino, S.; Cohen, M.L.; Schonberger, L.B.; Gambetti, P.; Appleby, B.S.; et al. Prion Propagation Estimated from Brain Diffusion MRI Is Subtype Dependent in Sporadic Creutzfeldt-Jakob Disease. Acta Neuropathol. 2020, 140, 169–181. [Google Scholar] [CrossRef]
- Peelaerts, W.; Bousset, L.; van der Perren, A.; Moskalyuk, A.; Pulizzi, R.; Giugliano, M.; van den Haute, C.; Melki, R.; Baekelandt, V. α-Synuclein Strains Cause Distinct Synucleinopathies after Local and Systemic Administration. Nature 2015, 522, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.-Y.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. Alpha-Synuclein in Lewy Bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef]
- Grazia Spillantini, M.; Anthony Crowther, R.; Jakes, R.; Cairns, N.J.; Lantos, P.L.; Goedert, M. Filamentous Alpha-Synuclein Inclusions Link Multiple System Atrophy with Parkinson’s Disease and Dementia with Lewy Bodies. Neurosci. Lett. 1998, 251, 205–208. [Google Scholar] [CrossRef]
- Dhillon, J.K.S.; Trejo-Lopez, J.A.; Riffe, C.; Levites, Y.; Sacino, A.N.; Borchelt, D.R.; Yachnis, A.Y.; Giasson, B.I. Comparative Analyses of the in Vivo Induction and Transmission of α-Synuclein Pathology in Transgenic Mice by MSA Brain Lysate and Recombinant α-Synuclein Fibrils. Acta Neuropathol. Commun. 2019, 7, 80. [Google Scholar] [CrossRef] [PubMed]
- van der Perren, A.; Gelders, G.; Fenyi, A.; Bousset, L.; Brito, F.; Peelaerts, W.; van den Haute, C.; Gentleman, S.; Melki, R.; Baekelandt, V. The Structural Differences between Patient-Derived α-Synuclein Strains Dictate Characteristics of Parkinson’s Disease, Multiple System Atrophy and Dementia with Lewy Bodies. Acta Neuropathol. 2020, 139, 977–1000. [Google Scholar] [CrossRef] [PubMed]
- Rey, N.L.; Bousset, L.; George, S.; Madaj, Z.; Meyerdirk, L.; Schulz, E.; Steiner, J.A.; Melki, R.; Brundin, P. α-Synuclein Conformational Strains Spread, Seed and Target Neuronal Cells Differentially after Injection into the Olfactory Bulb. Acta Neuropathol. Commun. 2019, 7, 221. [Google Scholar] [CrossRef] [Green Version]
- Sorrentino, Z.A.; Xia, Y.; Gorion, K.M.; Hass, E.; Giasson, B.I. Carboxy-Terminal Truncations of Mouse α-Synuclein Alter Aggregation and Prion-like Seeding. FEBS Lett. 2020, 594, 1271–1283. [Google Scholar] [CrossRef] [PubMed]
- Lau, A.; So, R.W.L.; Lau, H.H.C.; Sang, J.C.; Ruiz-Riquelme, A.; Fleck, S.C.; Stuart, E.; Menon, S.; Visanji, N.P.; Meisl, G.; et al. α-Synuclein Strains Target Distinct Brain Regions and Cell Types. Nat. Neurosci. 2020, 23, 21–31. [Google Scholar] [CrossRef]
- Chung, H.K.; Ho, H.-A.; Pérez-Acuña, D.; Lee, S.-J. Modeling α-Synuclein Propagation with Preformed Fibril Injections. J. Mov. Disord. 2019, 12, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Mehra, S.; Gadhe, L.; Bera, R.; Sawner, A.S.; Maji, S.K. Structural and Functional Insights into α-Synuclein Fibril Polymorphism. Biomolecules 2021, 11, 419. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, S.O.; Uzunoğlu, G.; Nussbaum-Krammer, C. α-Synuclein Strains: Does Amyloid Conformation Explain the Heterogeneity of Synucleinopathies? Biomolecules 2021, 11, 931. [Google Scholar] [CrossRef] [PubMed]
- Laferrière, F.; Claverol, S.; Bezard, E.; De Giorgi, F.; Ichas, F. Similar Neuronal Imprint and No Cross-Seeded Fibrils in α-Synuclein Aggregates from MSA and Parkinson’s Disease. NPJ Parkinson’s Dis. 2022, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.L.; Delucia, M.W.; Dickson, D.W. Alpha-Synuclein Immunoreactivity in Neuronal Nuclear Inclusions and Neurites in Multiple System Atrophy. Neurosci. Lett. 2004, 354, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Nishie, M.; Mori, F.; Yoshimoto, M.; Takahashi, H.; Wakabayashi, K. A Quantitative Investigation of Neuronal Cytoplasmic and Intranuclear Inclusions in the Pontine and Inferior Olivary Nuclei in Multiple System Atrophy. Neuropathol. Appl. Neurobiol. 2004, 30, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Laferrière, F.; He, X.; Zinghirino, F.; Doudnikoff, E.; Faggiani, E.; Meissner, W.G.; Bezard, E.; De Giorgi, F.; Ichas, F. Overexpression of α-Synuclein by Oligodendrocytes in Transgenic Mice Does Not Recapitulate the Fibrillar Aggregation Seen in Multiple System Atrophy. Cells 2020, 9, 2371. [Google Scholar] [CrossRef] [PubMed]
- Uemura, N.; Uemura, M.T.; Lo, A.; Bassil, F.; Zhang, B.; Luk, K.C.; Lee, V.M.Y.; Takahashi, R.; Trojanowski, J.Q. Slow Progressive Accumulation of Oligodendroglial Alpha-Synuclein (a-Syn) Pathology in Synthetic a-Syn Fibril-Induced Mouse Models of Synucleinopathy. J. Neuropathol. Exp. Neurol. 2019, 78, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Papp, M.I.; Lantos, P.L. Accumulation of Tubular Structures in Oligodendroglial and Neuronal Cells as the Basic Alteration in Multiple System Atrophy. J. Neurol. Sci. 1992, 107, 172–182. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Yoshimoto, M.; Tsuji, S.; Takahashi, H. Alpha-Synuclein Immunoreactivity in Glial Cytoplasmic Inclusions in Multiple System Atrophy. Neurosci. Lett. 1998, 249, 180–182. [Google Scholar] [CrossRef]
- Woulfe, J.M. Abnormalities of the Nucleus and Nuclear Inclusions in Neurodegenerative Disease: A Work in Progress. Neuropathol. Appl. Neurobiol. 2007, 33, 2–42. [Google Scholar] [CrossRef]
- Shahnawaz, M.; Mukherjee, A.; Pritzkow, S.; Mendez, N.; Rabadia, P.; Liu, X.; Hu, B.; Schmeichel, A.; Singer, W.; Wu, G.; et al. Discriminating α-Synuclein Strains in Parkinson’s Disease and Multiple System Atrophy. Nature 2020, 578, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Vranken, W.F.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, M.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN Data Model for NMR Spectroscopy: Development of a Software Pipeline. Proteins 2005, 59, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Buffeteau, T.; Lagugné-Labarthet, F.; Sourisseau, C. Vibrational Circular Dichroism in General Anisotropic Thin Solid Films: Measurement and Theoretical Approach. Appl. Spectrosc. 2005, 59, 732–745. [Google Scholar] [CrossRef]
- de Chaumont, F.; Dallongeville, S.; Chenouard, N.; Hervé, N.; Pop, S.; Provoost, T.; Meas-Yedid, V.; Pankajakshan, P.; Lecomte, T.; le Montagner, Y.; et al. Icy: An Open Bioimage Informatics Platform for Extended Reproducible Research. Nat. Methods 2012, 9, 690–696. [Google Scholar] [CrossRef]
- Serpell, L.C.; Berriman, J.; Jakes, R.; Goedert, M.; Crowther, R.A. Fiber Diffraction of Synthetic α-Synuclein Filaments Shows Amyloid-like Cross-β Conformation. Proc. Natl. Acad. Sci. USA 2000, 97, 4897–4902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Ge, P.; Murray, K.A.; Sheth, P.; Zhang, M.; Nair, G.; Sawaya, M.R.; Shin, W.S.; Boyer, D.R.; Ye, S.; et al. Cryo-EM of Full-Length α-Synuclein Reveals Fibril Polymorphs with a Common Structural Kernel. Nat. Commun. 2018, 9, 3609. [Google Scholar] [CrossRef] [PubMed]
- Heise, H.; Hoyer, W.; Becker, S.; Andronesi, O.C.; Riedel, D.; Baldus, M. Molecular-Level Secondary Structure, Polymorphism, and Dynamics of Full-Length α-Synuclein Fibrils Studied by Solid-State NMR. Proc. Natl. Acad. Sci. USA 2005, 102, 15871–15876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gath, J.; Bousset, L.; Habenstein, B.; Melki, R.; Bö Ckmann, A. Unlike Twins: An NMR Comparison of Two a-Synuclein Polymorphs Featuring Different Toxicity. PLoS ONE 2014, 9, 90659. [Google Scholar] [CrossRef] [PubMed]
- Loquet, A.; Giller, K.; Becker, S.; Lange, A. Supramolecular Interactions Probed by 13C-13C Solid-State NMR Spectroscopy. J. Am. Chem. Soc. 2010, 132, 15164–15166. [Google Scholar] [CrossRef] [PubMed]
- Gath, J.; Bousset, L.; Habenstein, B.; Melki, R.; Meier, B.H.; Böckmann, A. Yet Another Polymorph of α-Synuclein: Solid-State Sequential Assignments. Biomol. NMR Assign. 2014, 8, 395–404. [Google Scholar] [CrossRef] [Green Version]
- Gath, J.; Habenstein, B.; Bousset, L.; Melki, R.; Meier, B.H.; Böckmann, A. Solid-State NMR Sequential Assignments of α-Synuclein. Biomol. NMR Assign. 2012, 6, 51–55. [Google Scholar] [CrossRef]
- Guerrero-Ferreira, R.; Taylor, N.M.I.; Arteni, A.A.; Kumari, P.; Mona, D.; Ringler, P.; Britschgi, M.; Lauer, M.E.; Makky, A.; Verasdock, J.; et al. Two New Polymorphic Structures of Human Full-Length Alpha-Synuclein Fibrils Solved by Cryo-Electron Microscopy. eLife 2019, 8, e48907. [Google Scholar] [CrossRef] [PubMed]
- Verasdonck, J.; Bousset, L.; Gath, J.; Melki, R.; Böckmann, A.; Meier, B.H. Further Exploration of the Conformational Space of α-Synuclein Fibrils: Solid-State NMR Assignment of a High-PH Polymorph. Biomol. NMR Assign. 2016, 10, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N.; Li, J.; Fink, A.L. Evidence for a Partially Folded Intermediate in α-Synuclein Fibril Formation. J. Biol. Chem. 2001, 276, 10737–10744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, R.A.G.D.; Kubelka, J.; Bour, P.; Decatur, S.M.; Keiderling, T.A. Site-Specific Conformational Determination in Thermal Unfolding Studies of Helical Peptides Using Vibrational Circular Dichroism with Isotopic Substitution. Proc. Natl. Acad. Sci. USA 2000, 97, 8318–8323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterlini, M.G.; Freedman, T.B.; Nafie, L.A. Vibrational Circular Dichroism Spectra of Three Conformationally Distinct States and an Unordered State of Poly(L-Lysine) in Deuterated Aqueous Solution. Biopolymers 1986, 25, 1751–1765. [Google Scholar] [CrossRef] [PubMed]
- Volpicelli-Daley, L.A.; Luk, K.C.; Lee, V.M.Y. Addition of Exogenous α-Synuclein Preformed Fibrils to Primary Neuronal Cultures to Seed Recruitment of Endogenous α-Synuclein to Lewy Body and Lewy Neurite-like Aggregates. Nat. Protoc. 2014, 9, 2135–2146. [Google Scholar] [CrossRef] [Green Version]
- Gribaudo, S.; Tixador, P.; Bousset, L.; Fenyi, A.; Lino, P.; Melki, R.; Peyrin, J.M.; Perrier, A.L. Propagation of α-Synuclein Strains within Human Reconstructed Neuronal Network. Stem Cell Rep. 2019, 12, 230–244. [Google Scholar] [CrossRef] [Green Version]
- Gray, M.T.; Munoz, D.G.; Gray, D.A.; Schlossmacher, M.G.; Woulfe, J.M. Alpha-Synuclein in the Appendiceal Mucosa of Neurologically Intact Subjects. Mov. Disord. Off. J. Mov. Disord. Soc. 2014, 29, 991–998. [Google Scholar] [CrossRef]
- Teixeira, M.; Sheta, R.; Idi, W.; Oueslati, A. Alpha-Synuclein and the Endolysosomal System in Parkinson’s Disease: Guilty by Association. Biomolecules 2021, 11, 1333. [Google Scholar] [CrossRef]
- Vaikath, N.N.; Majbour, N.K.; Paleologou, K.E.; Ardah, M.T.; van Dam, E.; van de Berg, W.D.J.; Forrest, S.L.; Parkkinen, L.; Gai, W.P.; Hattori, N.; et al. Generation and Characterization of Novel Conformation-Specific Monoclonal Antibodies for α-Synuclein Pathology. Neurobiol. Dis. 2015, 79, 81–99. [Google Scholar] [CrossRef]
- Dominguez-Meijide, A.; Parrales, V.; Vasili, E.; González-Lizárraga, F.; König, A.; Lázaro, D.F.; Lannuzel, A.; Haik, S.; del Bel, E.; Chehín, R.; et al. Doxycycline Inhibits α-Synuclein-Associated Pathologies in Vitro and in Vivo. Neurobiol. Dis. 2021, 151, 105256. [Google Scholar] [CrossRef] [PubMed]
- Kuzuhara, S.; Mori, H.; Izumiyama, N.; Yoshimura, M.; Ihara, Y. Lewy Bodies Are Ubiquitinated. Acta Neuropathol. 1988, 75, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Kuusisto, E.; Salminen, A.; Alafuzoff, I. Ubiquitin-Binding Protein P62 Is Present in Neuronal and Glial Inclusions in Human Tauopathies and Synucleinopathies. Neuroreport 2001, 12, 2085–2090. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, T.; Iwatsubo, T.; Hasegawa, M. Ubiquitination of α-Synuclein. Biochemistry 2005, 44, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Fu, A.; Cohen-Kaplan, V.; Avni, N.; Livneh, I.; Ciechanover, A. P62-Containing, Proteolytically Active Nuclear Condensates, Increase the Efficiency of the Ubiquitin-Proteasome System. Proc. Natl. Acad. Sci. USA 2021, 118, e2107321118. [Google Scholar] [CrossRef] [PubMed]
- Awa, S.; Suzuki, G.; Masuda-Suzukake, M.; Nonaka, T.; Saito, M.; Hasegawa, M. Phosphorylation of Endogenous α-Synuclein Induced by Extracellular Seeds Initiates at the Pre-Synaptic Region and Spreads to the Cell Body. Sci. Rep. 2022, 12, 1163. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.B.; Porta, S.; Michael Baer, G.; Xu, Y.; Suh, E.R.; Kwong, L.K.; Elman, L.; Grossman, M.; Lee, V.M.Y.; Irwin, D.J.; et al. Expansion of the Classification of FTLD-TDP: Distinct Pathology Associated with Rapidly Progressive Frontotemporal Degeneration. Acta Neuropathol. 2017, 134, 65–78. [Google Scholar] [CrossRef]
- Chen, A.K.H.; Lin, R.Y.Y.; Hsieh, E.Z.J.; Tu, P.H.; Chen, R.P.Y.; Liao, T.Y.; Chen, W.; Wang, C.H.; Huang, J.J.T. Induction of Amyloid Fibrils by the C-Terminal Fragments of TDP-43 in Amyotrophic Lateral Sclerosis. J. Am. Chem. Soc. 2010, 132, 1186–1187. [Google Scholar] [CrossRef]
- Goers, J.; Manning-Bog, A.B.; McCormack, A.L.; Millett, I.S.; Doniach, S.; di Monte, D.A.; Uversky, V.N.; Fink, A.L. Nuclear Localization of Alpha-Synuclein and Its Interaction with Histones. Biochemistry 2003, 42, 8465–8471. [Google Scholar] [CrossRef]
- Koss, D.J.; Erskine, D.; Porter, A.; Palmoski, P.; Leite, M.; Attems, J.; Outeiro, T.F. Nuclear Alpha-Synuclein Is Present in the Human Brain and Is Modified in Dementia with Lewy Bodies. bioRxiv 2021. [Google Scholar] [CrossRef]
- Pasha, T.; Zatorska, A.; Sharipov, D.; Rogelj, B.; Hortobágyi, T.; Hirth, F. Karyopherin Abnormalities in Neurodegenerative Proteinopathies. Brain J. Neurol. 2021, 144, 2915–2932. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.L.; Song, L.K.; Yuan, Y.H.; Zhang, Y.; Han, N.; Gao, K.; Chen, N.H. The Nuclear Accumulation of Alpha-Synuclein Is Mediated by Importin Alpha and Promotes Neurotoxicity by Accelerating the Cell Cycle. Neuropharmacology 2014, 82, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Woulfe, J.M.; Prichett-Pejic, W.; Rippstein, P.; Munoz, D.G. Promyelocytic Leukaemia-Immunoreactive Neuronal Intranuclear Rodlets in the Human Brain. Neuropathol. Appl. Neurobiol. 2007, 33, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Singh, N.; Kumar, R.; Patel, K.; Pandey, S.; Datta, D.; Mahato, J.; Panigrahi, R.; Navalkar, A.; Mehra, S.; et al. α-Synuclein Aggregation Nucleates through Liquid–Liquid Phase Separation. Nat. Chem. 2020, 12, 705–716. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Giorgi, F.; Abdul-Shukkoor, M.B.; Kashyrina, M.; Largitte, L.-A.; De Nuccio, F.; Kauffmann, B.; Lends, A.; Laferrière, F.; Bonhommeau, S.; Lofrumento, D.D.; et al. Neurons with Cat’s Eyes: A Synthetic Strain of α-Synuclein Fibrils Seeding Neuronal Intranuclear Inclusions. Biomolecules 2022, 12, 436. https://doi.org/10.3390/biom12030436
De Giorgi F, Abdul-Shukkoor MB, Kashyrina M, Largitte L-A, De Nuccio F, Kauffmann B, Lends A, Laferrière F, Bonhommeau S, Lofrumento DD, et al. Neurons with Cat’s Eyes: A Synthetic Strain of α-Synuclein Fibrils Seeding Neuronal Intranuclear Inclusions. Biomolecules. 2022; 12(3):436. https://doi.org/10.3390/biom12030436
Chicago/Turabian StyleDe Giorgi, Francesca, Muhammed Bilal Abdul-Shukkoor, Marianna Kashyrina, Leslie-Ann Largitte, Francesco De Nuccio, Brice Kauffmann, Alons Lends, Florent Laferrière, Sébastien Bonhommeau, Dario Domenico Lofrumento, and et al. 2022. "Neurons with Cat’s Eyes: A Synthetic Strain of α-Synuclein Fibrils Seeding Neuronal Intranuclear Inclusions" Biomolecules 12, no. 3: 436. https://doi.org/10.3390/biom12030436
APA StyleDe Giorgi, F., Abdul-Shukkoor, M. B., Kashyrina, M., Largitte, L.-A., De Nuccio, F., Kauffmann, B., Lends, A., Laferrière, F., Bonhommeau, S., Lofrumento, D. D., Bousset, L., Bezard, E., Buffeteau, T., Loquet, A., & Ichas, F. (2022). Neurons with Cat’s Eyes: A Synthetic Strain of α-Synuclein Fibrils Seeding Neuronal Intranuclear Inclusions. Biomolecules, 12(3), 436. https://doi.org/10.3390/biom12030436