
Pharmacological and Genetic Evidence of Dopamine Receptor 3-Mediated Vasoconstriction in Isolated Mouse Aorta

 ,
,  , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Vessels and Analysis of Vascular Responses
2.3. In Vitro Lentiviral Delivery of Synthetic miRNA
2.4. Analysis of mRNA Expression by Real-Time Quantitative PCR
2.5. Drugs and Reagents
2.6. Statistical Analysis
3. Results
3.1. Vasomotor Responses in Aortic Segments Isolated from WT and D3R−/−
3.2. Vasomotor Effect of DR Agonist and Antagonists
3.3. Epression of DR mRNA in Aortic Segments in Basal Condition and Following Knockdown
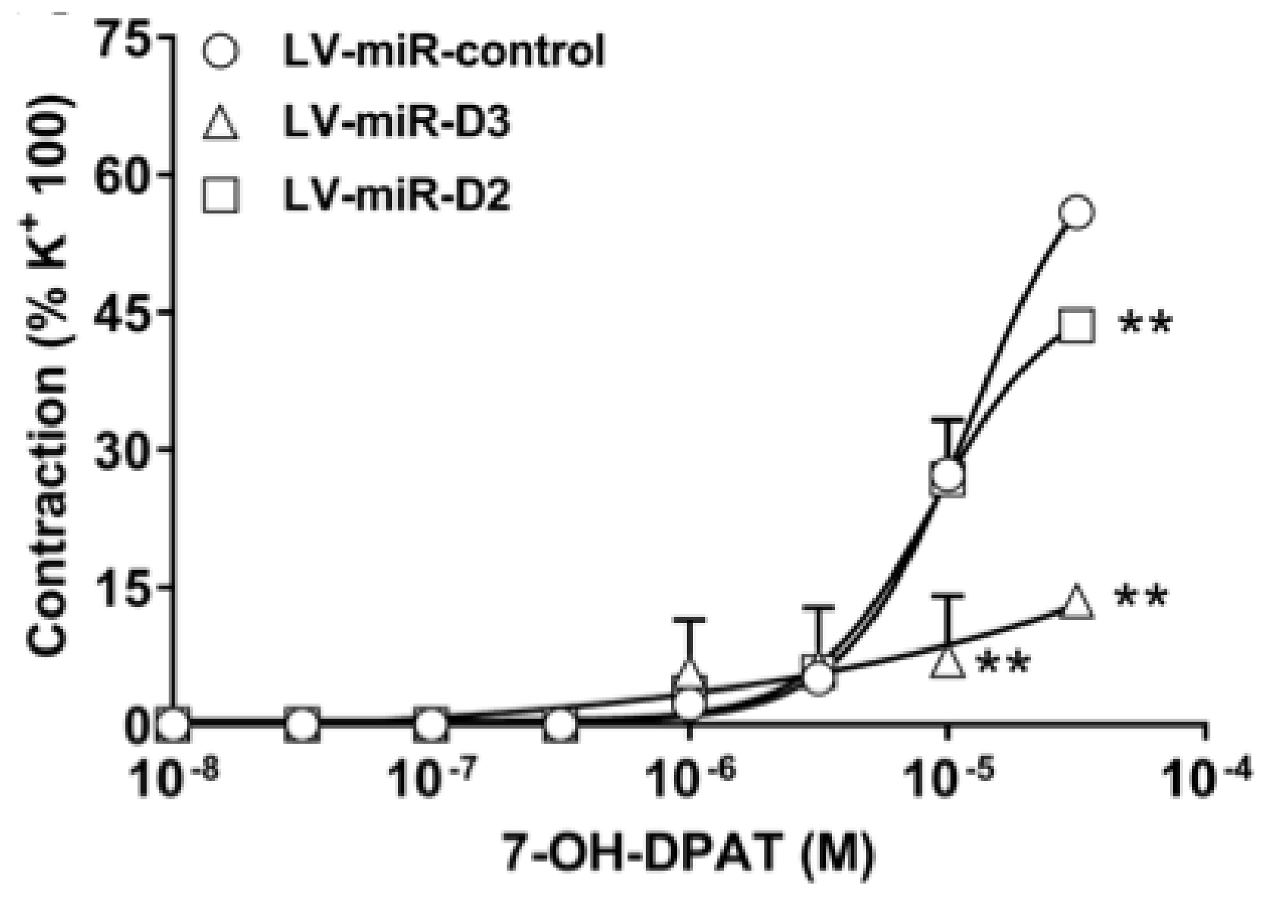
3.4. Effect of DR Knockdown on Vasomotor Responses to DR Agonist
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure to function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [CrossRef]
- Beaulieu, J.M.; Gainetdinov, R.R. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol. Rev. 2011, 63, 182–217. [Google Scholar] [CrossRef]
- Maggio, R.; Scarselli, M.; Capannolo, M.; Millan, M.J. Novel dimensions of D3 receptor function: Focus on heterodimerisation, transactivation and allosteric modulation. Eur. Neuropsychopharmacol. 2015, 25, 1470–1479. [Google Scholar] [CrossRef]
- Kapur, S.; Mamo, D. Half a century of antipsychotics and still a central role for dopamine D2 receptors. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Torrisi, S.A.; Salomone, S.; Geraci, F.; Caraci, F.; Bucolo, C.; Drago, F.; Leggio, G.M. Buspirone counteracts MK-801-induced schizophrenia-like phenotypes through dopamine D3 receptor blockade. Front. Pharmacol. 2017, 8, 710. [Google Scholar] [CrossRef]
- Leggio, G.M.; Torrisi, S.; Mastrogiacomo, R.; Mauro, D.; Chisari, M.; Manago, F.; Scheggia, D.; Nigro, M.; Giurdanella, G.; Costa, L.; et al. The epistatic interaction between the dopamine D3 receptor and dysbindin-1 modulates higher-order cognitive functions in mice and humans. Mol. Psychiatry 2019. [Google Scholar] [CrossRef]
- Seeman, P. Parkinson’s disease treatment may cause impulse-control disorder via dopamine D3 receptors. Synapse 2015, 69, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Heidbreder, C.A.; Gardner, E.L.; Xi, Z.X.; Thanos, P.K.; Mugnaini, M.; Hagan, J.J.; Ashby, C.R., Jr. The role of central dopamine D3 receptors in drug addiction: A review of pharmacological evidence. Brain Res. Brain Res. Rev. 2005, 49, 77–105. [Google Scholar] [CrossRef]
- Joyce, J.N.; Millan, M.J. Dopamine D3 receptor antagonists as therapeutic agents. Drug Discov. Today 2005, 10, 917–925. [Google Scholar] [CrossRef]
- Caine, S.B.; Koob, G.F. Modulation of cocaine self-administration in the rat through D-3 dopamine receptors. Science 1993, 260, 1814–1816. [Google Scholar] [CrossRef] [PubMed]
- Neisewander, J.L.; Fuchs, R.A.; Tran-Nguyen, L.T.; Weber, S.M.; Coffey, G.P.; Joyce, J.N. Increases in dopamine D3 receptor binding in rats receiving a cocaine challenge at various time points after cocaine self-administration: Implications for cocaine-seeking behavior. Neuropsychopharmacology 2004, 29, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Leggio, G.M.; Camillieri, G.; Platania, C.B.; Castorina, A.; Marrazzo, G.; Torrisi, S.A.; Nona, C.N.; D’Agata, V.; Nobrega, J.; Stark, H.; et al. Dopamine D3 receptor is necessary for ethanol consumption: An approach with buspirone. Neuropsychopharmacology 2014, 39, 2017–2028. [Google Scholar] [CrossRef]
- Leggio, G.M.; Di Marco, R.; Gulisano, W.; D’Ascenzo, M.; Torrisi, S.A.; Geraci, F.; Lavanco, G.; Dahl, K.; Giurdanella, G.; Castorina, A.; et al. Dopaminergic-GABAergic interplay and alcohol binge drinking. Pharmacol. Res. 2019, 141, 384–391. [Google Scholar] [CrossRef]
- Cao, J.L.; Covington, H.E., 3rd; Friedman, A.K.; Wilkinson, M.B.; Walsh, J.J.; Cooper, D.C.; Nestler, E.J.; Han, M.H. Mesolimbic dopamine neurons in the brain reward circuit mediate susceptibility to social defeat and antidepressant action. J. Neurosci. 2010, 30, 16453–16458. [Google Scholar] [CrossRef]
- Leggio, G.M.; Salomone, S.; Bucolo, C.; Platania, C.; Micale, V.; Caraci, F.; Drago, F. Dopamine D3 receptor as a new pharmacological target for the treatment of depression. Eur. J. Pharmacol. 2013, 719, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Leggio, G.M.; Micale, V.; Drago, F. Increased sensitivity to antidepressants of D3 dopamine receptor-deficient mice in the forced swim test (FST). Eur. Neuropsychopharmacol. 2008, 18, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Xing, B.; Liu, P.; Jiang, W.H.; Liu, F.; Zhang, H.; Cao, G.F.; Chen, T.; Dang, Y.H. Effects of immobilization stress on emotional behaviors in dopamine D3 receptor knockout mice. Behav. Brain Res. 2013, 243, 261–266. [Google Scholar] [CrossRef]
- Leggio, G.M.; Micale, V.; Le Foll, B.; Mazzola, C.; Nobrega, J.N.; Drago, F. Dopamine D3 receptor knock-out mice exhibit increased behavioral sensitivity to the anxiolytic drug diazepam. Eur. Neuropsychopharmacol. 2011, 21, 325–332. [Google Scholar] [CrossRef]
- Leggio, G.M.; Torrisi, S.A.; Castorina, A.; Platania, C.B.; Impellizzeri, A.A.; Fidilio, A.; Caraci, F.; Bucolo, C.; Drago, F.; Salomone, S. Dopamine D3 receptor-dependent changes in alpha6 GABAA subunit expression in striatum modulate anxiety-like behaviour: Responsiveness and tolerance to diazepam. Eur. Neuropsychopharmacol. 2015, 25, 1427–1436. [Google Scholar] [CrossRef]
- Ruffolo, R.R., Jr.; Messick, K.; Horng, J.S. Interactions of three inotropic agents, ASL-7022, dobutamine and dopamine, with alpha- and beta-adrenoceptors in vitro. Naunyn Schmiedeberg Arch. Pharmacol. 1984, 326, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, L.I.; Rajfer, S.I. Dopamine receptors: Applications in clinical cardiology. Circulation 1985, 72, 245–248. [Google Scholar] [CrossRef]
- Nichols, A.J.; Ruffolo, R.R., Jr.; Brooks, D.P. The pharmacology of fenoldopam. Am. J. Hypertens. 1990, 3, 116S–119S. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.W.; Broadley, K.J. Renal vasodilatation by dopexamine and fenoldopam due to alpha 1-adrenoceptor blockade. Br. J. Pharmacol. 1995, 115, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Amenta, F.; Collier, W.L.; Ricci, A. Autoradiographic localization of vascular dopamine receptors. Am. J. Hypertens. 1990, 3, 34S–36S. [Google Scholar] [CrossRef]
- Salomone, S.; Waeber, C. Selectivity and specificity of sphingosine-1-phosphate receptor ligands: Caveats and critical thinking in characterizing receptor-mediated effects. Front. Pharmacol. 2011, 2, 9. [Google Scholar] [CrossRef]
- Pfeil, U.; Kuncova, J.; Bruggmann, D.; Paddenberg, R.; Rafiq, A.; Henrich, M.; Weigand, M.A.; Schluter, K.D.; Mewe, M.; Middendorff, R.; et al. Intrinsic vascular dopamine—A key modulator of hypoxia-induced vasodilatation in splanchnic vessels. J. Physiol. 2014, 592, 1745–1756. [Google Scholar] [CrossRef]
- Li, J.; Gu, J.; Wang, B.; Xie, M.; Huang, L.; Liu, Y.; Zhang, L.; Xue, J.; Guo, F. Activation of Dopamine D1 Receptors Regulates Dendritic Morphogenesis Through Rac1 and RhoA in Prefrontal Cortex Neurons. Mol. Neurobiol. 2015, 51, 1024–1037. [Google Scholar] [CrossRef]
- Tu, G.; Ying, L.; Ye, L.; Zhao, J.; Liu, N.; Li, J.; Liu, Y.; Zhu, M.; Wu, Y.; Xiao, B.; et al. Dopamine D1 and D2 Receptors Differentially Regulate Rac1 and Cdc42 Signaling in the Nucleus Accumbens to Modulate Behavioral and Structural Plasticity After Repeated Methamphetamine Treatment. Biol. Psychiatry 2019, 86, 820–835. [Google Scholar] [CrossRef]
- Sawada, N.; Salomone, S.; Kim, H.H.; Kwiatkowski, D.J.; Liao, J.K. Regulation of endothelial nitric oxide synthase and postnatal angiogenesis by Rac1. Circ. Res. 2008, 103, 360–368. [Google Scholar] [CrossRef]
- Wang, W.Z.; Zhao, R.R.; Qin, F.Z. Comparison of the effects of dopamine1- and dopamine2-receptor agonists on the cAMP generating system in canine coronary and renal arteries. Methods Find. Exp. Clin. Pharmacol. 1994, 16, 691–696. [Google Scholar]
- Zhu, L.; Zhao, R.; Zhang, W.F. Effects of dopamine receptor agonists on the cAMP content in arteries of the rabbit. Sheng Li Xue Bao 2000, 52, 247–251. [Google Scholar] [PubMed]
- Haynes, J.; Robinson, J., Jr.; Saunders, L.; Taylor, A.E.; Strada, S.J. Role of cAMP-dependent protein kinase in cAMP-mediated vasodilation. Am. J. Physiol. 1992, 262, H511–H516. [Google Scholar] [CrossRef] [PubMed]
- Accili, D.; Fishburn, C.S.; Drago, J.; Steiner, H.; Lachowicz, J.E.; Park, B.H.; Gauda, E.B.; Lee, E.J.; Cool, M.H.; Sibley, D.R.; et al. A targeted mutation of the D3 dopamine receptor gene is associated with hyperactivity in mice. Proc. Natl. Acad. Sci. USA 1996, 93, 1945–1949. [Google Scholar] [CrossRef]
- Salomone, S.; Soydan, G.; Moskowitz, M.A.; Sims, J.R. Inhibition of cerebral vasoconstriction by dantrolene and nimodipine. Neurocrit. Care 2009, 10, 93–102. [Google Scholar] [CrossRef]
- Salomone, S.; Silva, C.L.; Morel, N.; Godfraind, T. Facilitation of the vasorelaxant action of calcium antagonists by basal nitric oxide in depolarized artery. Naunyn Schmiedeberg Arch. Pharmacol. 1996, 354, 505–512. [Google Scholar] [CrossRef]
- Scheggia, D.; Mastrogiacomo, R.; Mereu, M.; Sannino, S.; Straub, R.E.; Armando, M.; Managò, F.; Guadagna, S.; Piras, F.; Zhang, F.; et al. Variations in Dysbindin-1 are associated with cognitive response to antipsychotic drug treatment. Nat. Commun. 2018, 9, 2265. [Google Scholar] [CrossRef] [PubMed]
- Platania, C.B.; Salomone, S.; Leggio, G.M.; Drago, F.; Bucolo, C. Homology modeling of dopamine D2 and D3 receptors: Molecular dynamics refinement and docking evaluation. PLoS ONE 2012, 7, e44316. [Google Scholar] [CrossRef]
- Bucolo, C.; Leggio, G.M.; Drago, F.; Salomone, S. Dopamine outside the brain: The eye, cardiovascular system and endocrine Pancreas. Pharmacol. Ther. 2019, 203, 107392. [Google Scholar] [CrossRef]
- Vass, M.; Kooistra, A.J.; Yang, D.; Stevens, R.C.; Wang, M.W.; de Graaf, C. Chemical diversity in the G protein-coupled receptor superfamily. Trends Pharmacol. Sci. 2018, 39, 494–512. [Google Scholar] [CrossRef]
- Burris, K.D.; Pacheco, M.A.; Filtz, T.M.; Kung, M.P.; Kung, H.F.; Molinoff, P.B. Lack of discrimination by agonists for D2 and D3 dopamine receptors. Neuropsychopharmacology 1995, 12, 335–345. [Google Scholar] [CrossRef]
- Chumpradit, S.; Kung, M.P.; Vessotskie, J.; Foulon, C.; Mu, M.; Kung, H.F. Iodinated 2-aminotetralins and 3-amino-1-benzopyrans: Ligands for dopamine D2 and D3 receptor. J. Med. Chem. 1994, 37, 4245–4250. [Google Scholar] [CrossRef] [PubMed]
- Freedman, S.B.; Patel, S.; Marwood, R.; Emms, F.; Seabrook, G.R.; Knowles, M.R.; McAllister, G. Expression and pharmacological characterization of the human D3 dopamine receptor. J. Pharmacol. Exp. Ther. 1994, 268, 417–426. [Google Scholar] [PubMed]
- Sautel, F.; Griffon, N.; Lévesque, D.; Pilon, C.; Schwartz, J.C.; Sokoloff, P. A functional test identifies dopamine agonists selective for D3 versus D2 receptors. Neuroreport 1995, 6, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Grundt, P.; Husband, S.L.; Luedtke, R.R.; Taylor, M.; Newman, A.H. Analogues of the dopamine D2 receptor antagonist L741,626: Binding, function, and SAR. Bioorg. Med. Chem. Lett. 2007, 17, 745–749. [Google Scholar] [CrossRef]
- Kulagowski, J.J.; Broughton, H.B.; Curtis, N.R.; Mawer, I.M.; Ridgill, M.P.; Baker, R.; Emms, F.; Freedman, S.B.; Marwood, R.; Patel, S.; et al. 3-((4-(4-Chlorophenyl)piperazin-1-yl)-methyl)-1H-pyrrolo-2,3-b-pyridine: An antagonist with high affinity and selectivity for the human dopamine D4 receptor. J. Med. Chem. 1996, 39, 1941–1942. [Google Scholar] [CrossRef]
- Millan, M.J.; Gobert, A.; Newman-Tancredi, A.; Lejeune, F.; Cussac, D.; Rivet, J.M.; Audinot, V.; Dubuffet, T.; Lavielle, G. S33084, a novel, potent, selective, and competitive antagonist at dopamine D(3)-receptors: I. Receptorial, electrophysiological and neurochemical profile compared with GR218,231 and L741,626. J. Pharmacol. Exp. Ther. 2000, 293, 1048–1062. [Google Scholar]
- Reavill, C.; Taylor, S.G.; Wood, M.D.; Ashmeade, T.; Austin, N.E.; Avenell, K.Y.; Boyfield, I.; Branch, C.L.; Cilia, J.; Coldwell, M.C.; et al. Pharmacological actions of a novel, high-affinity, and selective human dopamine D(3) receptor antagonist, SB-277011-A. J. Pharmacol. Exp. Ther. 2000, 294, 1154–1165. [Google Scholar] [PubMed]
- Arunlakshana, O.; Schild, H.O. Some quantitative uses of drug antagonists. Br. J. Pharmacol. Chemother. 1959, 14, 48–58. [Google Scholar] [CrossRef]
- Scigliano, G.; Ronchetti, G. Antipsychotic-induced metabolic and cardiovascular side effects in schizophrenia: A novel mechanistic hypothesis. CNS Drug 2013, 27, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Han, Y.; Wang, X.; Chen, C.; Yu, C.; He, D.; Wang, H.; Zhou, L.; Asico, L.D.; Jose, P.A.; et al. Inhibitory effect of the D(3) dopamine receptor on insulin receptor expression and function in vascular smooth muscle cells. Am. J. Hypertens. 2011, 24, 654–660. [Google Scholar] [CrossRef]
- Kok, P.; Roelfsema, F.; Frolich, M.; van Pelt, J.; Stokkel, M.P.; Meinders, A.E.; Pijl, H. Activation of dopamine D2 receptors simultaneously ameliorates various metabolic features of obese women. Am. J. Physiol. 2006, 291, E1038–E1043. [Google Scholar] [CrossRef] [PubMed]
- Uvnas-Moberg, K.; Ahlenius, S.; Alster, P.; Hillegaart, V. Effects of selective serotonin and dopamine agonists on plasma levels of glucose, insulin and glucagon in the rat. Neuroendocrinology 1996, 63, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Han, Y.; Wang, H.; Wang, Z.; Liu, Y.; Chen, X.; Cai, Y.; Guan, W.; Yang, D.; Asico, L.D.; et al. Impaired dopamine D1 receptor-mediated vasorelaxation of mesenteric arteries in obese Zucker rats. Cardiovasc. Diabetol. 2014, 13, 50. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Amenta, F.; Bronzetti, E.; Felici, L.; Hussain, T.; Lokhandwala, M.F. Age-related changes of dopamine receptor protein immunoreactivity in the rat mesenteric vascular tree. Mech. Ageing Dev. 2002, 123, 537–546. [Google Scholar] [CrossRef]
- Zeng, C.; Wang, D.; Asico, L.D.; Welch, W.J.; Wilcox, C.S.; Hopfer, U.; Eisner, G.M.; Felder, R.A.; Jose, P.A. Aberrant D1 and D3 dopamine receptor transregulation in hypertension. Hypertension 2004, 43, 654–660. [Google Scholar] [CrossRef]
- Hennekens, C.H.; Hennekens, A.R.; Hollar, D.; Casey, D.E. Schizophrenia and increased risks of cardiovascular disease. Am. Heart J. 2005, 150, 1115–1121. [Google Scholar] [CrossRef] [PubMed]
- Osborn, D.P.; Levy, G.; Nazareth, I.; Petersen, I.; Islam, A.; King, M.B. Relative risk of cardiovascular and cancer mortality in people with severe mental illness from the United Kingdom’s General Practice Research Database. Arch. Gen. Psychiatry 2007, 64, 242–249. [Google Scholar] [CrossRef]
- Pratt, L.A.; Ford, D.E.; Crum, R.M.; Armenian, H.K.; Gallo, J.J.; Eaton, W.W. Depression, psychotropic medication, and risk of myocardial infarction: Prospective data from the Baltimore ECA follow-up. Circulation 1996, 94, 3123–3129. [Google Scholar] [CrossRef]
- Lin, S.T.; Chen, C.C.; Tsang, H.Y.; Lee, C.S.; Yang, P.; Cheng, K.D.; Li, D.J.; Wang, C.J.; Hsieh, Y.C.; Yang, W.C. Association between antipsychotic use and risk of acute myocardial infarction: A nationwide case-crossover study. Circulation 2014, 130, 235–243. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| D2R | D3R | 5-HT1A | 5-HT1D | 5-HT2A | 5-HT2B | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 7-OH-DPAT | 2.6–165 [40] | 142 [41] | 60 [42] | 103 [43] | 0.4–1.2 [40] | 2.90 [41] | 1.6 [42] | 2.1 [43] | 72.7 [41] | |||
| L741,626 | 11.2 [44] | 2.4 [45] | 4.0–6.3 [46] | 163 [44] | 100 [45] | 63 [46] | 316 [46] | 631 [46] | ||||
| SB-277011-A | 1047 [47] | 11.0 [47] | 1621 [47] | 1288 [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zingales, V.; Torrisi, S.A.; Leggio, G.M.; Bucolo, C.; Drago, F.; Salomone, S. Pharmacological and Genetic Evidence of Dopamine Receptor 3-Mediated Vasoconstriction in Isolated Mouse Aorta. Biomolecules 2021, 11, 418. https://doi.org/10.3390/biom11030418
Zingales V, Torrisi SA, Leggio GM, Bucolo C, Drago F, Salomone S. Pharmacological and Genetic Evidence of Dopamine Receptor 3-Mediated Vasoconstriction in Isolated Mouse Aorta. Biomolecules. 2021; 11(3):418. https://doi.org/10.3390/biom11030418
Chicago/Turabian StyleZingales, Veronica, Sebastiano Alfio Torrisi, Gian Marco Leggio, Claudio Bucolo, Filippo Drago, and Salvatore Salomone. 2021. "Pharmacological and Genetic Evidence of Dopamine Receptor 3-Mediated Vasoconstriction in Isolated Mouse Aorta" Biomolecules 11, no. 3: 418. https://doi.org/10.3390/biom11030418
APA StyleZingales, V., Torrisi, S. A., Leggio, G. M., Bucolo, C., Drago, F., & Salomone, S. (2021). Pharmacological and Genetic Evidence of Dopamine Receptor 3-Mediated Vasoconstriction in Isolated Mouse Aorta. Biomolecules, 11(3), 418. https://doi.org/10.3390/biom11030418