Implications of the Adiponectin System in Non-Small Cell Lung Cancer Patients: A Case-Control Study

 , ,
, ,  , and
, and 
Abstract
1. Introduction
- -
- Low Molecular Weight (LMW);
- -
- Medium Molecular Weight (MMW);
- -
- High Molecular Weight (HMW) oligomers [19].
2. Material and Methods
2.1. Subjects
2.2. Anthropometric and Biochemical Measurements
2.3. Western Blotting Analysis
2.4. RNA Extraction and Real-Time Quantitative PCR
2.5. Gel Filtration Analysis
2.6. Statistical Analysis
3. Results
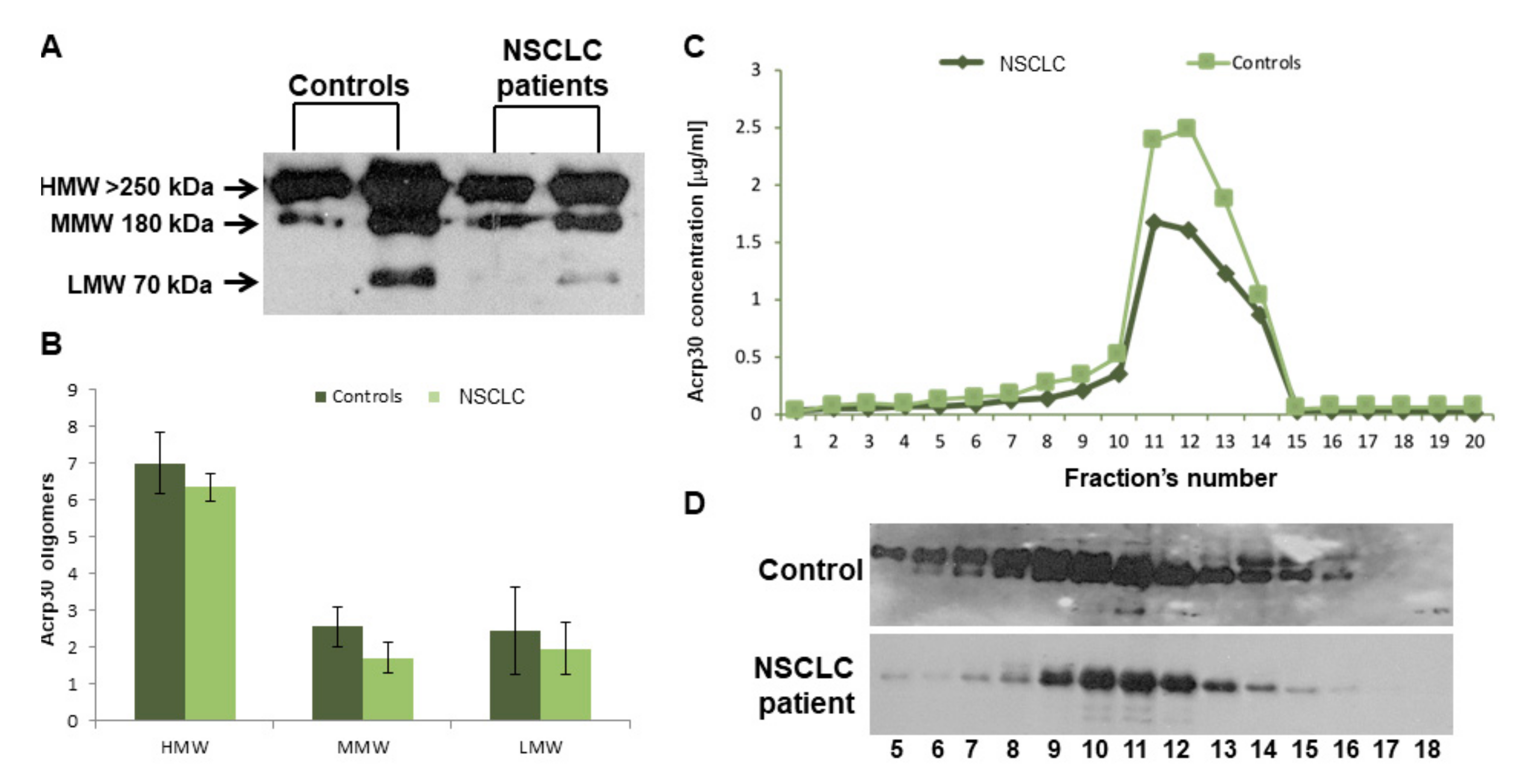
3.1. Baseline Features and Serum Levels of Total and HMW Acrp30
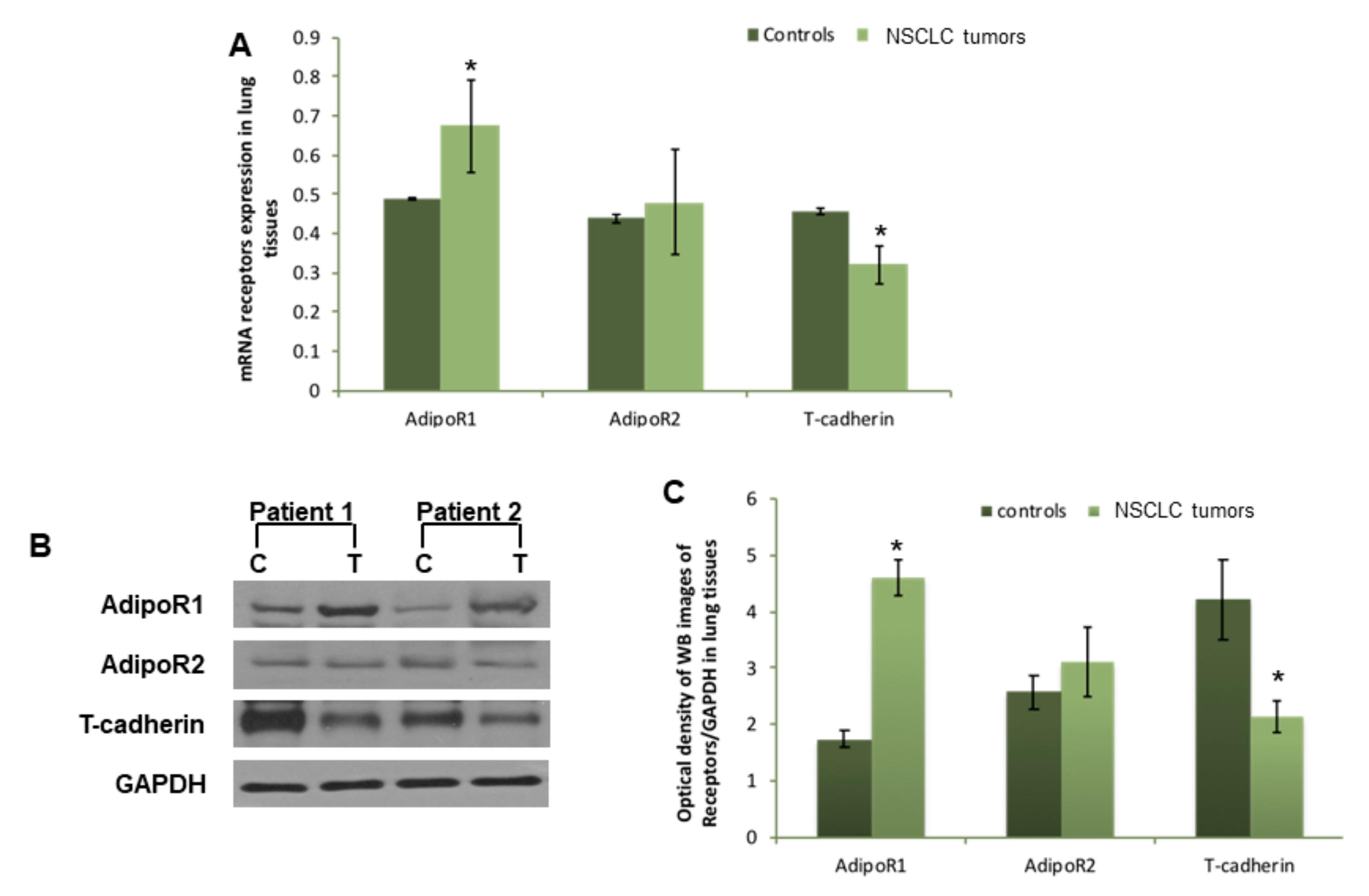
3.2. AdipoR1, AdipoR2, and T-Cadherin Expression in Lung Tissues
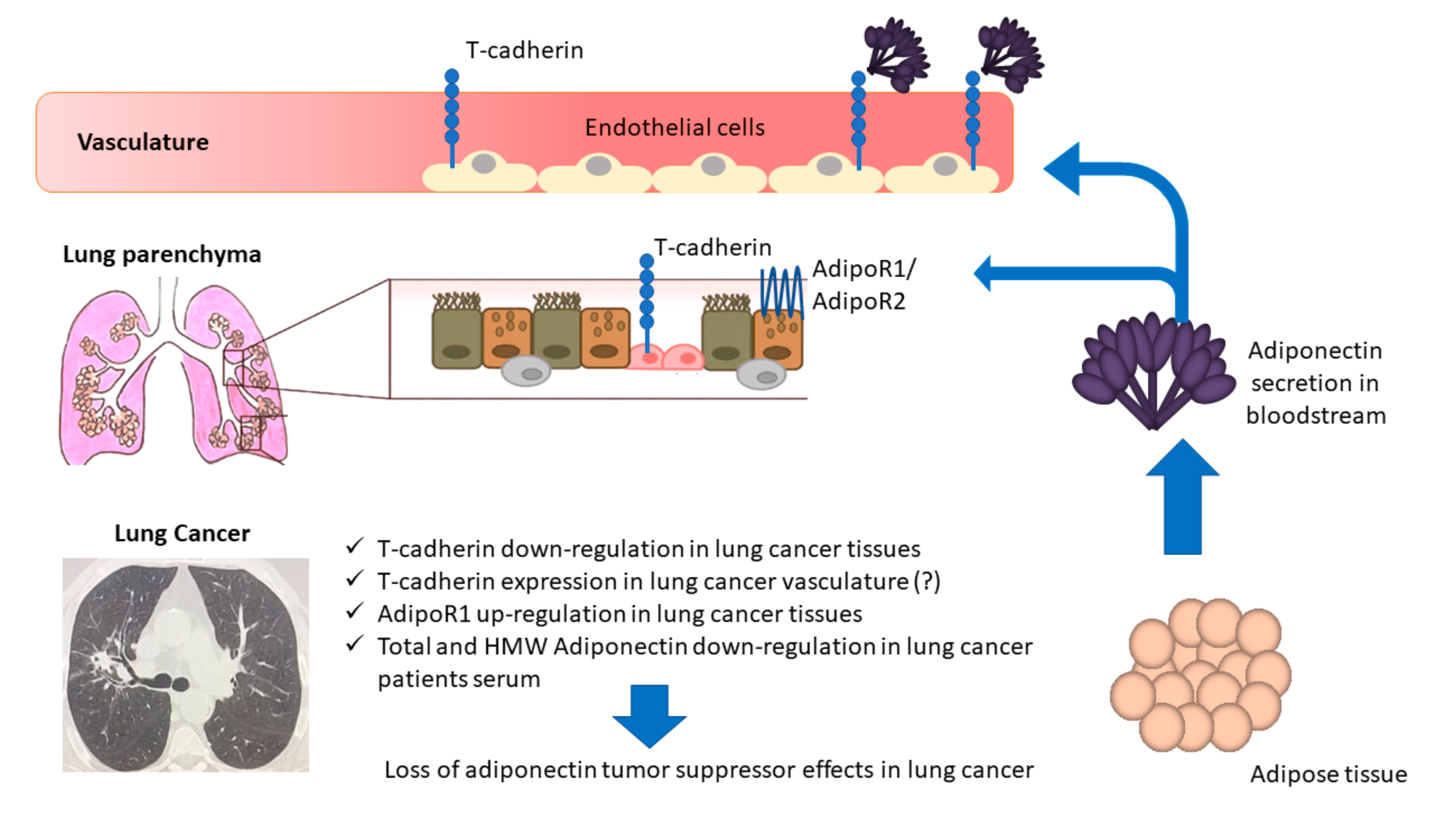
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nawaz, K.; Webster, R.M. The non-small-cell lung cancer drug market. Nat. Rev. Drug Discov. 2016, 15, 229–230. [Google Scholar] [CrossRef] [PubMed]
- Travis, W.D. Lung Cancer Pathology: Current Concepts. Clin. Chest Med. 2020, 41, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Relli, V.; Trerotola, M.; Guerra, E.; Alberti, S. Abandoning the Notion of Non-Small Cell Lung Cancer. Trends Mol. Med. 2019, 25, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bianco, A.; Malkinson, A.M.; Leoni, V.P.; Frau, G.; De Rosa, N.; André, P.-A.; Versace, R.; Boulvain, M.; Laurent, G.J.; et al. BARD1: An independent predictor of survival in non-small cell lung cancer. Int. J. Cancer 2011, 131, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.; Malapelle, U.; Rocco, D.; Perrotta, F.; Mazzarella, G. Targeting immune checkpoints in non small cell lung cancer. Curr. Opin. Pharmacol. 2018, 40, 46–50. [Google Scholar] [CrossRef]
- Cattaneo, F.; Guerra, G.; Parisi, M.; Lucariello, A.; De Luca, A.; De Rosa, N.; Mazzarella, G.; Bianco, A.; Ammendola, R. Expression of Formyl-peptide Receptors in Human Lung Carcinoma. Anticancer. Res. 2015, 35, 2769–2774. [Google Scholar]
- Nigro, E.; Stiuso, P.; Matera, M.; Monaco, M.L.; Caraglia, M.; Maniscalco, M.; Perrotta, F.; Mazzarella, G.; Daniele, A.; Bianco, A. The anti-proliferative effects of adiponectin on human lung adenocarcinoma A549 cells and oxidative stress involvement. Pulm. Pharmacol. Ther. 2019, 55, 25–30. [Google Scholar] [CrossRef]
- Nigro, E.; Imperlini, E.; Scudiero, O.; Monaco, M.L.; Polito, R.; Mazzarella, G.; Orrù, S.; Bianco, A.; Daniele, A. Differentially expressed and activated proteins associated with non small cell lung cancer tissues. Respir. Res. 2015, 16, 74. [Google Scholar] [CrossRef]
- Illiano, M.; Nigro, E.; Sapio, L.; Caiafa, I.; Spina, A.; Scudiero, O.; Bianco, A.; Esposito, S.; Mazzeo, F.; Pedone, P.V.; et al. Adiponectin down-regulates CREB and inhibits proliferation of A549 lung cancer cells. Pulm. Pharmacol. Ther. 2017, 45, 114–120. [Google Scholar] [CrossRef]
- Bianco, A.; Perrotta, F.; Barra, G.; Malapelle, U.; Rocco, D.; De Palma, R. Prognostic Factors and Biomarkers of Responses to Immune Checkpoint Inhibitors in Lung Cancer. Int. J. Mol. Sci. 2019, 20, 4931–4946. [Google Scholar] [CrossRef]
- McKay, J.D.; SpiroMeta Consortium; Hung, R.J.; Han, Y.; Zong, X.; Carreras-Torres, R.; Christiani, D.C.; Caporaso, N.E.; Johansson, M.; Xiao, X.; et al. Large-scale association analysis identifies new lung cancer susceptibility loci and heterogeneity in genetic susceptibility across histological subtypes. Nat. Genet. 2017, 49, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Bossé, Y.; Li, Z.; Xia, J.; Manem, V.; Carreras-Torres, R.; Gabriel, A.; Gaudreault, N.; Albanes, D.; Aldrich, M.C.; Andrew, A.; et al. Transcriptome-wide association study reveals candidate causal genes for lung cancer. Int. J. Cancer 2020, 146, 1862–1878. [Google Scholar]
- Thériault, S.; Gaudreault, N.; Lamontagne, M.; Rosa, M.; Boulanger, M.-C.; Messika-Zeitoun, D.; Clavel, M.-A.; Capoulade, R.; Dagenais, F.; Pibarot, P.; et al. A transcriptome-wide association study identifies PALMD as a susceptibility gene for calcific aortic valve stenosis. Nat. Commun. 2018, 9, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Lengyel, E.; Makowski, L.; DiGiovanni, J.; Kolonin, M.G. Cancer as a Matter of Fat: The Crosstalk between Adipose Tissue and Tumors. Trends Cancer 2018, 4, 374–384. [Google Scholar] [CrossRef]
- Chu, D.-T.; Phuong, T.N.T.; Tien, N.L.B.; Tran, D.-K.; Nguyen, T.; Van Thanh, V.; Quang, T.L.; Minh, L.B.; Pham, V.H.; Ngoc, V.T.N.; et al. The Effects of Adipocytes on the Regulation of Breast Cancer in the Tumor Microenvironment: An Update. Cells 2019, 8, 857–876. [Google Scholar] [CrossRef]
- Son, B.; Lee, S.; Youn, H.; Kim, E.; Kim, W.; Youn, B. The role of tumor microenvironment in therapeutic resistance. Oncotarget 2016, 8, 3933–3945. [Google Scholar] [CrossRef]
- Ntikoudi, E.; Kiagia, M.; Boura, P.; Syrigos, K. Hormones of adipose tissue and their biologic role in lung cancer. Cancer Treat. Rev. 2014, 40, 22–30. [Google Scholar] [CrossRef]
- Di Zazzo, E.; Polito, R.; Bartollino, S.; Nigro, E.; Porcile, C.; Bianco, A.; Daniele, A.; Moncharmont, B. Adiponectin as Link Factor between Adipose Tissue and Cancer. Int. J. Mol. Sci. 2019, 20, 839–852. [Google Scholar] [CrossRef]
- Fang, H.; Judd, R.L. Adiponectin Regulation and Function. Comprehensive Physiology 2018, 8, 1031–1063. [Google Scholar] [CrossRef]
- Van Andel, M.; Heijboer, A.C.; Drent, M.L. Adiponectin and Its Isoforms in Pathophysiology. Adv. Clin. Chem. 2018, 85, 115–147. [Google Scholar] [CrossRef]
- Buechler, C.; Wanninger, J.; Neumeier, M. Adiponectin receptor binding proteins - recent advances in elucidating adiponectin signalling pathways. FEBS Lett. 2010, 584, 4280–4286. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T. Adiponectin and Adiponectin Receptors. Endocr. Rev. 2005, 26, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Lang, K.; Ratke, J. Leptin and Adiponectin: New players in the field of tumor cell and leukocyte migration. Cell Commun. Signal. 2009, 7, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Kita, S.; Fukuda, S.; Maeda, N.; Shimomura, I. Native adiponectin in serum binds to mammalian cells expressing T-cadherin, but not AdipoRs or calreticulin. eLife 2019, 8, 1–13. [Google Scholar] [CrossRef]
- Maeda, N.; Funahashi, T.; Matsuzawa, Y.; Shimomura, I. Adiponectin, a unique adipocyte-derived factor beyond hormones. Atherosclerosis 2019, 292, 1–9. [Google Scholar] [CrossRef]
- Perrotta, F.; Nigro, E.; Mollica, M.; Costigliola, A.; D’Agnano, V.; Daniele, A.; Bianco, A.; Guerra, G. Pulmonary Hypertension and Obesity: Focus on Adiponectin. Int. J. Mol. Sci. 2019, 20, 912–925. [Google Scholar] [CrossRef]
- Khan, S.; Shukla, S.; Sinha, S.; Meeran, S.M. Role of adipokines and cytokines in obesity-associated breast cancer: Therapeutic targets. Cytokine Growth Factor Rev. 2013, 24, 503–513. [Google Scholar] [CrossRef]
- Li, F.; Cao, Y.; Li, J.; Gao, C.; Dong, X.; Ren, P.; Meng, C.; Chen, C. The clinical significance of serum adipocytokines level in patients with lung cancer. J. Thorac. Dis. 2019, 11, 3547–3555. [Google Scholar] [CrossRef]
- Boura, P.; Grapsa, D.; Loukides, S.; Angelidou, M.; Tsakanika, K.; Syrigos, N.; Gkiozos, I. The prognostic value of serum and bronchoalveolar lavage levels of adiponectin in advanced non-small-cell lung cancer. Lung Cancer Manag. 2017, 6, 55–65. [Google Scholar] [CrossRef]
- Pecoraro, A.; Nigro, E.; Polito, R.; Monaco, M.L.; Scudiero, O.; Mormile, I.; Marcelli, A.C.; Capasso, M.; Habetswallner, F.; Genovese, A.; et al. Total and High Molecular Weight Adiponectin Expression Is Decreased in Patients with Common Variable Immunodeficiency: Correlation with Ig Replacement Therapy. Front. Immunol. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Nigro, E.; Scudiero, O.; Monaco, M.L.; Polito, R.; Schettino, P.; Grandone, A.; Perrone, L.; Del Giudice, E.M.; Daniele, A. Adiponectin profile and Irisin expression in Italian obese children: Association with insulin-resistance. Cytokine 2017, 94, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Dias, A.S.; Almeida, C.R.; Helguero, L.A.; Duarte, I.F. Metabolic crosstalk in the breast cancer microenvironment. Eur. J. Cancer 2019, 121, 154–171. [Google Scholar] [CrossRef] [PubMed]
- Vona-Davis, L.; Gibson, L.F. Adipocytes as a critical component of the tumor microenvironment. Leuk. Res. 2013, 37, 483–484. [Google Scholar] [CrossRef][Green Version]
- Bianco, A.; Mazzarella, G.; Turchiarelli, V.; Nigro, E.; Corbi, G.; Scudiero, O.; Sofia, M.; Daniele, A. Adiponectin: An Attractive Marker for Metabolic Disorders in Chronic Obstructive Pulmonary Disease (COPD). Nutrients 2013, 5, 4115–4125. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.; Nigro, E.; Monaco, M.L.; Matera, M.G.; Scudiero, O.; Mazzarella, G.; Daniele, A. The burden of obesity in asthma and COPD: Role of adiponectin. Pulm. Pharmacol. Ther. 2017, 43, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Petridou, E.T.; Mitsiades, N.; Gialamas, S.; Angelopoulos, M.; Skalkidou, A.; Dessypris, N.; Hsi, A.; Lazaris, N.; Polyzos, A.; Syrigos, C.; et al. Circulating Adiponectin Levels and Expression of Adiponectin Receptors in Relation to Lung Cancer: Two Case-Control Studies. Oncology 2007, 73, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Karapanagiotou, E.M.; Tsochatzis, E.A.; Dilana, K.D.; Tourkantonis, I.; Gratsias, I.; Syrigos, K.N.; Tsochatzis, E.A. The significance of leptin, adiponectin, and resistin serum levels in non-small cell lung cancer (NSCLC). Lung Cancer 2008, 61, 391–397. [Google Scholar] [CrossRef]
- Kerenidi, T.; Lada, M.; Tsaroucha, A.; Georgoulias, P.; Mystridou, P.; Gourgoulianis, K.I. Clinical significance of serum adipokines levels in lung cancer. Med Oncol. 2013, 30, 1–8. [Google Scholar] [CrossRef]
- Wang, H.; Wu, J.; Gu, W.; Wang, B.; Wan, F.; Dai, B.; Zhang, H.; Shi, G.; Shen, Y.; Zhu, Y.; et al. Serum Adiponectin Level May be an Independent Predictor of Clear Cell Renal Cell Carcinoma. J. Cancer 2016, 7, 1340–1346. [Google Scholar] [CrossRef]
- Hebbard, L.; Ranscht, B. Multifaceted roles of adiponectin in cancer. Best Pr. Res. Clin. Endocrinol. Metab. 2013, 28, 59–69. [Google Scholar] [CrossRef]
- Zhong, Y.; Delgado, Y.; Gomez, J.; Lee, S.W.; Perez-Soler, R. Loss of H-cadherin protein expression in human non-small cell lung cancer is associated with tumorigenicity. Clin. Cancer Res. 2001, 7, 1683–1687. [Google Scholar] [PubMed]
- Tanaka, Y.; Kita, S.; Nishizawa, H.; Fukuda, S.; Fujishima, Y.; Obata, Y.; Nagao, H.; Masuda, S.; Nakamura, Y.; Shimizu, Y.; et al. Adiponectin promotes muscle regeneration through binding to T-cadherin. Sci. Rep. 2019, 9, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Denzel, M.S.; Scimia, M.-C.; Zumstein, P.M.; Walsh, K.; Ruiz-Lozano, P.; Ranscht, B. T-cadherin is critical for adiponectin-mediated cardioprotection in mice. J. Clin. Investig. 2010, 120, 4342–4352. [Google Scholar] [CrossRef] [PubMed]
- Bag, S.; Anbarasu, A. Revealing the Strong Functional Association of adipor2 and cdh13 with adipoq: A Gene Network Study. Cell Biophys. 2014, 71, 1445–1456. [Google Scholar] [CrossRef]
- Berx, G.; Van Roy, F. Involvement of Members of the Cadherin Superfamily in Cancer. Cold Spring Harb. Perspect. Boil. 2009, 1, 1–28. [Google Scholar] [CrossRef]
- Bosetti, C.; Rosato, V.; Buniato, D.; Zambon, A.; La Vecchia, C.; Corrao, G. Cancer Risk for Patients Using Thiazolidinediones for Type 2 Diabetes: A Meta-Analysis. Oncologist 2013, 18, 148–156. [Google Scholar] [CrossRef]
- Rispoli, M.; Salvi, R.; Cennamo, A.; Di Natale, D.; Natale, G.; Meoli, I.; Gioia, M.R.; Esposito, M.; Nespoli, M.R.; De Finis, M.; et al. Effectiveness of home-based preoperative pulmonary rehabilitation in COPD patients undergoing lung cancer resection. Tumori J. 2020. [Google Scholar] [CrossRef]
- Perrotta, F.; Cennamo, A.; Cerqua, F.S.; Stefanelli, F.; Bianco, A.; Musella, S.; Rispoli, M.; Salvi, R.; Meoli, I. Effects of a high-intensity pulmonary rehabilitation program on the minute ventilation/carbon dioxide output slope during exercise in a cohort of patients with COPD undergoing lung resection for non-small cell lung cancer. J. Bras. de Pneumol publicacao Of da Soc Bras Pneumol e Tisilogia. 2019, 45, 1–5. [Google Scholar] [CrossRef]
- The Effects of Nanocurcumin on Serum Oxidative Stress,Inflammation,Adiponectin and NF-kB in Blood Mononuclear Cells in Metabolic Syndrome Patients (Nuclear Factor-κB). Available online: https://clinicaltrials.gov/ct2/show/NCT03514667 (accessed on 27 April 2020).




{kind=link}
{kind=link}
{kind=link}
| Parameters | NSCLC Patients (n = 72) | Controls (n = 60) | p |
|---|---|---|---|
| Sex, n (%) | 0.300 | ||
| M | 46 (63.9) | 33 (55) | |
| F | 26 (36.1) | 27 (45) | |
| Age (yrs.), median [IQR] | 65 [58.5–70.5] | 63 [51–76.8] | 0.757 |
| Weight (kg), median [IQR] | 70 [64–79.5] | 70.5 [64.3–81.8] | 0.626 |
| BMI (kg/m2), median [IQR] | 25.3 [22.4–26.6] | 24.9 [23.7–25.7] | 0.696 |
| Histology, n (%) | n.a. | ||
| Adenocarcinoma | 32 (44.4) | - | |
| Squamous Cell Carcinoma | 27 (37.5) | - | |
| Other * | 13 (18.1) | - | |
| Stage, n (%) | - | ||
| 1/2 | 22 (30.6) | ||
| 3/4 | 50 (69.4) | - | |
| Performance status (ECOG), n (%) | - | ||
| 0/1 | 48 (66.7) | ||
| 2/3/4 | 24 (33.3) | - | |
| Lung Resection, n (%) | 20 (27.8) | - | n.a. |
| Brain Metastases, n (%) | 12 (16.6) | - | n.a. |
| Total Cholesterol (mg/dL), median [IQR] | 180 (165–195) | 179 (54.3–194.8) | 0.874 |
| Triglycerides (mg/dL), median [IQR] | 110 (88.3–135.8) | 98.5 (69–133.8) | 0.167 |
| Fasting Glucose (mg/dL), median [IQR] | 99 (89.3–108.8) | 89 (80.8–98.3) | < 0.001 |
| AST, median [IQR] | 20 (17–23.8) | 17.5 (15–21.8) | 0.039 |
| ALT, median [IQR] | 21 (17.3–27.8) | 14 (11–22) | < 0.001 |
| GGT, median [IQR] | 30.5 (21.3–41) | 16 (11–26.3) | < 0.001 |
| Acpr30 (μg/mL), median [IQR] | 10.8 (9.3–13.7) | 15.5 (12.6–19) | < 0.001 |
| Parameters | Coefficient | 95% | CI | p |
|---|---|---|---|---|
| Sex (M/F) | 0.132 | - 0.700 | 2.561 | 0.258 |
| Age (years) | 0.141 | - 0.036 | 0.151 | 0.223 |
| Stage (1/2 or 3/4) | 0.010 | - 1.849 | 2.000 | 0.938 |
| Histology (adenocarcinoma, SCC or Other*) | 0.347 | 0.566 | 2.606 | 0.003 |
| BMI (kg/m2) | 0.011 | - 0.340 | 0.370 | 0.933 |
| Cholesterol (mg/dL) | - 0.181 | - 0.055 | 0.008 | 0.147 |
| Triglycerides (mg/dL) | 0.225 | 0.000 | 0.047 | 0.048 |
| Fasting Glucose (mg/dL) | - 0.086 | - 0.041 | 0.019 | 0.462 |
| AST (U/L) | - 0.020 | - 0.124 | 0.105 | 0.870 |
| ALT (U/L) | 0.019 | - 0.072 | 0.083 | 0.886 |
| GGT (U/L) | 0.135 | - 0.015 | 0.062 | 0.231 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nigro, E.; Perrotta, F.; Monaco, M.L.; Polito, R.; Pafundi, P.C.; Matera, M.G.; Daniele, A.; Bianco, A. Implications of the Adiponectin System in Non-Small Cell Lung Cancer Patients: A Case-Control Study. Biomolecules 2020, 10, 926. https://doi.org/10.3390/biom10060926
Nigro E, Perrotta F, Monaco ML, Polito R, Pafundi PC, Matera MG, Daniele A, Bianco A. Implications of the Adiponectin System in Non-Small Cell Lung Cancer Patients: A Case-Control Study. Biomolecules. 2020; 10(6):926. https://doi.org/10.3390/biom10060926
Chicago/Turabian StyleNigro, Ersilia, Fabio Perrotta, Maria Ludovica Monaco, Rita Polito, Pia Clara Pafundi, Maria Gabriella Matera, Aurora Daniele, and Andrea Bianco. 2020. "Implications of the Adiponectin System in Non-Small Cell Lung Cancer Patients: A Case-Control Study" Biomolecules 10, no. 6: 926. https://doi.org/10.3390/biom10060926
APA StyleNigro, E., Perrotta, F., Monaco, M. L., Polito, R., Pafundi, P. C., Matera, M. G., Daniele, A., & Bianco, A. (2020). Implications of the Adiponectin System in Non-Small Cell Lung Cancer Patients: A Case-Control Study. Biomolecules, 10(6), 926. https://doi.org/10.3390/biom10060926