Impact of Heavy Metals on the Antioxidant Activity of Vitamin D: A Metabolic Perspective

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Measurement
2.2. Study-Group Classifications
2.3. UHPLC-QTOF/MS Analysis of Plasma Samples
2.4. Data Analysis
2.5. Putative Identification of Metabolites
2.6. Biomarker Analysis
2.7. Statistical Analysis
3. Results
3.1. Characteristics of the SVDD–LHM Group and the VDD–HHM Group
3.2. Correlation Analysis Between 25(OH)D and HMs
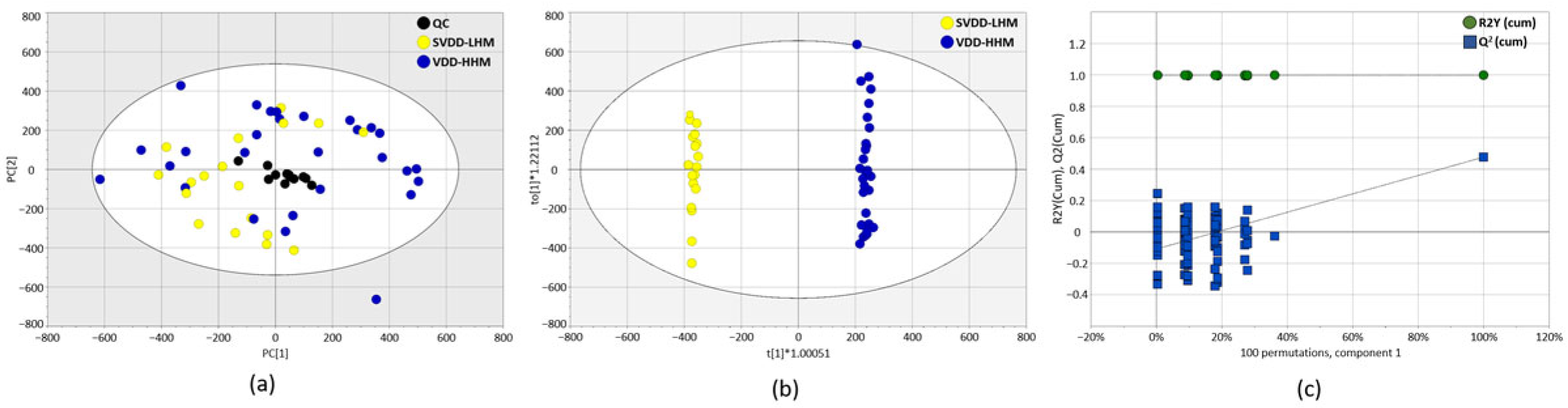
3.3. Untargeted Metabolic Analysis of Plasma Samples
3.4. Identification of Putative Metabolites
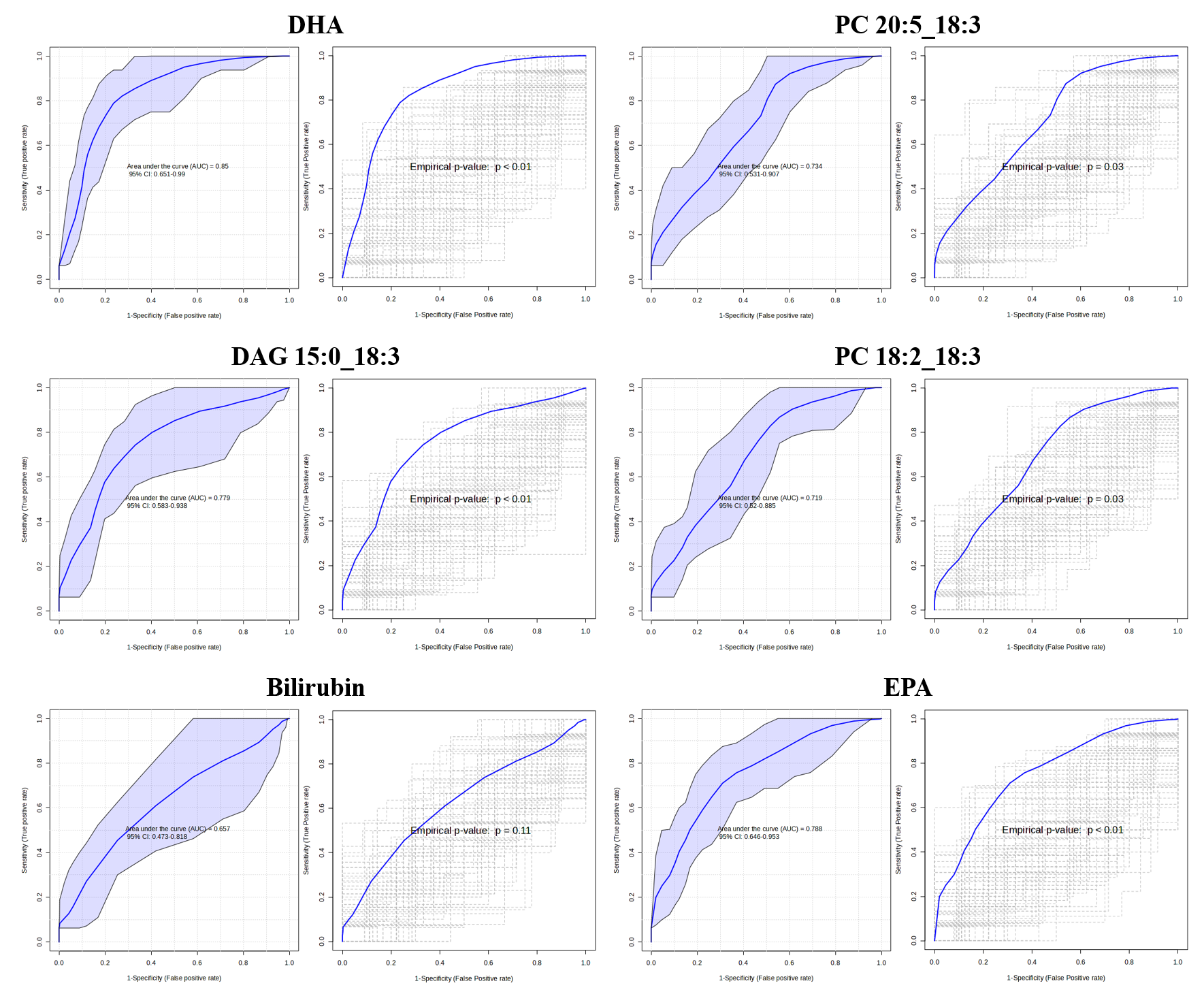
3.5. Identification of Potential Biomarker
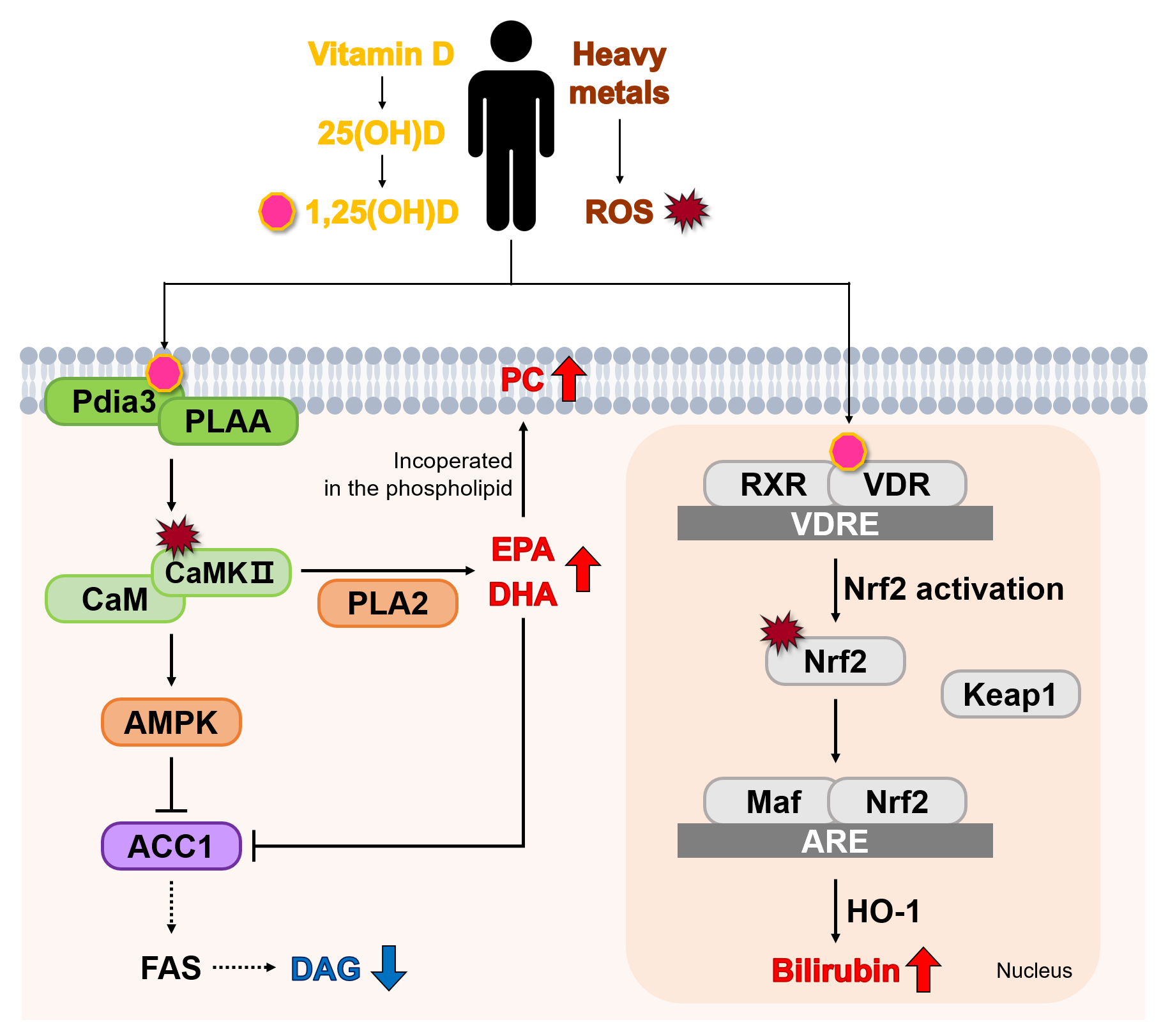
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berridge, M.J. Vitamin D cell signalling in health and disease. Biochem. Biophys. Res. Commun. 2015, 460, 53–71. [Google Scholar] [CrossRef]
- Grant, W.B.; Wimalawansa, S.J.; Pludowski, P.; Cheng, R.Z. Vitamin D: Evidence-Based Health Benefits and Recommendations for Population Guidelines. Nutrients 2025, 17, 277. [Google Scholar] [CrossRef]
- Giustina, A.; Bouillon, R.; Binkley, N.; Sempos, C.; Adler, R.A.; Bollerslev, J.; Dawson-Hughes, B.; Ebeling, P.R.; Feldman, D.; Heijboer, A.; et al. Controversies in Vitamin D: A Statement From the Third International Conference. JBMR Plus 2020, 4, e10417. [Google Scholar] [CrossRef]
- Holick, M.F.; Binkley, N.C.; Bischoff-Ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M. Evaluation, Treatment, and Prevention of Vitamin D Deficiency: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2011, 96, 1911–1930. [Google Scholar] [CrossRef] [PubMed]
- Amrein, K.; Scherkl, M.; Hoffmann, M.; Neuwersch-Sommeregger, S.; Köstenberger, M.; Tmava Berisha, A.; Martucci, G.; Pilz, S.; Malle, O. Vitamin D deficiency 2.0: An update on the current status worldwide. Eur. J. Clin. Nutr. 2020, 74, 1498–1513. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.J. Vitamin D Deficiency: Effects on Oxidative Stress, Epigenetics, Gene Regulation, and Aging. Biology 2019, 8, 30. [Google Scholar] [CrossRef]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic Mechanisms of Five Heavy Metals: Mercury, Lead, Chromium, Cadmium, and Arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhen, C.; Zeng, L.; Feng, H.; Wang, J.; Ai, Q.Y.H.; Ai, S.; Zhang, J.; Liang, Y.Y.; Xue, H.; et al. Association of blood cadmium and physical activity with mortality: A prospective cohort study. Ecotoxicol. Environ. Saf. 2025, 290, 117541. [Google Scholar] [CrossRef]
- Fu, Z.; Xi, S. The effects of heavy metals on human metabolism. Toxicol. Mech. Methods 2020, 30, 167–176. [Google Scholar] [CrossRef]
- Mitra, S.; Chakraborty, A.J.; Tareq, A.M.; Emran, T.B.; Nainu, F.; Khusro, A.; Idris, A.M.; Khandaker, M.U.; Osman, H.; Alhumaydhi, F.A.; et al. Impact of heavy metals on the environment and human health: Novel therapeutic insights to counter the toxicity. J. King Saud Univ. Sci. 2022, 34, 101865. [Google Scholar] [CrossRef]
- Chwalba, A.; Orłowska, J.; Słota, M.; Jeziorska, M.; Filipecka, K.; Bellanti, F.; Dobrakowski, M.; Kasperczyk, A.; Zalejska-Fiolka, J.; Kasperczyk, S. Effect of Cadmium on Oxidative Stress Indices and Vitamin D Concentrations in Children. J. Clin. Med. 2023, 12, 1572. [Google Scholar] [CrossRef] [PubMed]
- Flores-Bazán, T.; Izquierdo-Vega, J.A.; Guerrero-Solano, J.A.; Castañeda-Ovando, A.; Estrada-Luna, D.; Jiménez-Osorio, A.S. Interplay Between Vitamin D Levels and Heavy Metals Exposure in Pregnancy and Childbirth: A Systematic Review. Pathophysiology 2024, 31, 660–679. [Google Scholar] [CrossRef] [PubMed]
- Kemp Francis, W.; Neti Prasad, V.S.V.; Howell Roger, W.; Wenger, P.; Louria Donald, B.; Bogden John, D. Elevated Blood Lead Concentrations and Vitamin D Deficiency in Winter and Summer in Young Urban Children. Environ. Health Perspect. 2007, 115, 630–635. [Google Scholar] [CrossRef]
- Schwalfenberg, G.K.; Genuis, S.J. Vitamin D, Essential Minerals, and Toxic Elements: Exploring Interactions between Nutrients and Toxicants in Clinical Medicine. Sci. World J. 2015, 2015, 318595. [Google Scholar] [CrossRef] [PubMed]
- Jukic, A.M.Z.; Zuchniak, A.; Qamar, H.; Ahmed, T.; Mahmud, A.A.; Roth, D.E. Vitamin D Treatment during Pregnancy and Maternal and Neonatal Cord Blood Metal Concentrations at Delivery: Results of a Randomized Controlled Trial in Bangladesh. Environ. Health Perspect. 2020, 128, 117007. [Google Scholar] [CrossRef]
- Leung, R.Y.H.; Li, G.H.Y.; Cheung, B.M.Y.; Tan, K.C.B.; Kung, A.W.C.; Cheung, C.-L. Serum metabolomic profiling and its association with 25-hydroxyvitamin D. Clin. Nutr. 2020, 39, 1179–1187. [Google Scholar] [CrossRef]
- Kelly, R.S.; Bayne, H.; Spiro, A.; Vokonas, P.; Sparrow, D.; Weiss, S.T.; Schwartz, J.; Nassan, F.L.; Lee-Sarwar, K.; Huang, M.; et al. Metabolomic signatures of lead exposure in the VA Normative Aging Study. Environ. Res. 2020, 190, 110022. [Google Scholar] [CrossRef]
- Freedman, L.S.; Tasevska, N.; Kipnis, V.; Schatzkin, A.; Mares, J.; Tinker, L.; Potischman, N. Gains in statistical power from using a dietary biomarker in combination with self-reported intake to strengthen the analysis of a diet-disease association: An example from CAREDS. Am. J. Epidemiol. 2010, 172, 836–842. [Google Scholar] [CrossRef]
- Triba, M.N.; Le Moyec, L.; Amathieu, R.; Goossens, C.; Bouchemal, N.; Nahon, P.; Rutledge, D.N.; Savarin, P. PLS/OPLS models in metabolomics: The impact of permutation of dataset rows on the K-fold cross-validation quality parameters. Mol. Biosyst. 2015, 11, 13–19. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Broadhurst, D.I.; Wilson, M.; Wishart, D.S. Translational biomarker discovery in clinical metabolomics: An introductory tutorial. Metabolomics 2013, 9, 280–299. [Google Scholar] [CrossRef]
- Souza, A.L.; Patti, G.J. A Protocol for Untargeted Metabolomic Analysis: From Sample Preparation to Data Processing. Methods Mol. Biol. 2021, 2276, 357–382. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Lim, Y.H.; Park, J.B.; Rhie, J.; Lee, S.J. Environmental and Occupation Factors Associated with Vitamin D Deficiency in Korean Adults: The Korea National Health and Nutrition Examination Survey (KNHANES) 2010–2014. Int. J. Environ. Res. Public Health 2020, 17, 9166. [Google Scholar] [CrossRef]
- Park, J.H.; Hong, I.Y.; Chung, J.W.; Choi, H.S. Vitamin D status in South Korean population: Seven-year trend from the KNHANES. Medicine 2018, 97, e11032. [Google Scholar] [CrossRef]
- Sallsten, G.; Ellingsen, D.G.; Berlinger, B.; Weinbruch, S.; Barregard, L. Variability of lead in urine and blood in healthy individuals. Environ. Res. 2022, 212, 113412. [Google Scholar] [CrossRef] [PubMed]
- Zafar, S.; Fatima, S.; Asad, F.; Nazir, M.M.; Batool, S.; Ashraf, A. Combating Lead (Pb) Contamination: Integrating Biomonitoring, Advanced Detection, and Remediation for Environmental and Public Health. Water Air Soil Pollut. 2024, 236, 8. [Google Scholar] [CrossRef]
- Sun, H.; Wang, D.; Zhou, Z.; Ding, Z.; Chen, X.; Xu, Y.; Huang, L.; Tang, D. Association of cadmium in urine and blood with age in a general population with low environmental exposure. Chemosphere 2016, 156, 392–397. [Google Scholar] [CrossRef]
- Kim, S.A.; Kwon, Y.; Kim, S.; Joung, H. Assessment of Dietary Mercury Intake and Blood Mercury Levels in the Korean Population: Results from the Korean National Environmental Health Survey 2012-2014. Int. J. Environ. Res. Public Health 2016, 13, 877. [Google Scholar] [CrossRef]
- Orloff, K.; Mistry, K.; Metcalf, S. Biomonitoring for Environmental Exposures to Arsenic. J. Toxicol. Environ. Health B Crit. Rev. 2009, 12, 509–524. [Google Scholar] [CrossRef]
- Pan, Z.; Gong, T.; Liang, P. Heavy Metal Exposure and Cardiovascular Disease. Circ. Res. 2024, 134, 1160–1178. [Google Scholar] [CrossRef] [PubMed]
- Karadeniz, Y.; Özpamuk-Karadeniz, F.; Ahbab, S.; Ataoğlu, E.; Can, G. Vitamin D Deficiency Is a Potential Risk for Blood Pressure Elevation and the Development of Hypertension. Medicina 2021, 57, 1297. [Google Scholar] [CrossRef] [PubMed]
- Alcalá-Santiago, Á.; García-Villanova, B.; Ruíz-López, M.D.; Gil, Á.; Rodriguez-Barranco, M.; Sánchez, M.J.; Molina-Montes, E. Dietary and lifestyle determinants of vitamin D status in the UK Biobank Cohort study for predictive modeling. J. Nutr. Biochem. 2025, 142, 109919. [Google Scholar] [CrossRef]
- Li, T.; Yu, L.; Yang, Z.; Shen, P.; Lin, H.; Shui, L.; Tang, M.; Jin, M.; Chen, K.; Wang, J. Associations of Diet Quality and Heavy Metals with Obesity in Adults: A Cross-Sectional Study from National Health and Nutrition Examination Survey (NHANES). Nutrients 2022, 14, 4038. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.-Y.; Lee, Y.-S.; Lee, S.-G.; Seo, M.-N.; Choi, B.-S.; Kim, Y.-D.; Lim, J.-A.; Hwang, M.-S.; Kwon, H.-J.; Kim, Y.-M.; et al. Lead, Mercury, and Cadmium Exposure in the Korean General Population. J. Korean Med. Sci. 2018, 33, e9. [Google Scholar] [CrossRef]
- Zuo, J.; Huesker, K.; Liu, Y.; Hocher, J.-G.; Zhang, X.; von Baehr, V.; Krämer, B.K.; Hocher, B. Association of whole blood heavy metal concentrations with kidney function. Sci. Rep. 2025, 15, 8370. [Google Scholar] [CrossRef]
- Pawlukianiec, C.; Lauko, K.K.; Michalak, D.; Żendzian-Piotrowska, M.; Zalewska, A.; Maciejczyk, M. A comparative study on the antioxidant and antiglycation properties of different vitamin D forms. Eur. J. Med. Chem. 2025, 285, 117263. [Google Scholar] [CrossRef]
- Doroudi, M.; Schwartz, Z.; Boyan, B.D. Membrane-mediated actions of 1,25-dihydroxy vitamin D3: A review of the roles of phospholipase A2 activating protein and Ca2+/calmodulin-dependent protein kinase II. J. Steroid Biochem. Mol. Biol. 2015, 147, 81–84. [Google Scholar] [CrossRef]
- Chen, J.; Olivares-Navarrete, R.; Wang, Y.; Herman, T.R.; Boyan, B.D.; Schwartz, Z. Protein-disulfide isomerase-associated 3 (Pdia3) mediates the membrane response to 1,25-dihydroxyvitamin D3 in osteoblasts. J. Biol. Chem. 2010, 285, 37041–37050. [Google Scholar] [CrossRef]
- Erickson, J.R.; Joiner, M.L.; Guan, X.; Kutschke, W.; Yang, J.; Oddis, C.V.; Bartlett, R.K.; Lowe, J.S.; O’Donnell, S.E.; Aykin-Burns, N.; et al. A dynamic pathway for calcium-independent activation of CaMKII by methionine oxidation. Cell 2008, 133, 462–474. [Google Scholar] [CrossRef]
- Junho, C.V.C.; Caio-Silva, W.; Trentin-Sonoda, M.; Carneiro-Ramos, M.S. An Overview of the Role of Calcium/Calmodulin-Dependent Protein Kinase in Cardiorenal Syndrome. Front. Physiol. 2020, 11, 735. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, D.; Mouchlis, V.D.; Dennis, E.A. Omega-3 versus Omega-6 fatty acid availability is controlled by hydrophobic site geometries of phospholipase A2s. J. Lipid Res. 2021, 62, 100113. [Google Scholar] [CrossRef]
- Aldhafiri, F.K. Investigating the Role of EPA and DHA on Cellular Oxidative Stress; Profiling Antidiabetic and Antihypertensive Potential. J. Pharm. Bioallied Sci. 2022, 14, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Mei, Q.; Niu, R. Oxidative CaMKII as a potential target for inflammatory disease (Review). Mol. Med. Rep. 2019, 20, 863–870. [Google Scholar] [CrossRef]
- Meng, Y.; Ding, P.; Wang, H.; Yang, X.; Wang, Z.; Nie, D.; Liu, J.; Huang, Y.; Su, G.; Hu, J.; et al. Ca2+/calmodulin-dependent protein kinase II inhibition reduces myocardial fatty acid uptake and oxidation after myocardial infarction. Biochim. Biophys. Acta 2022, 1867, 159120. [Google Scholar] [CrossRef] [PubMed]
- Shibabaw, T. Omega-3 polyunsaturated fatty acids: Anti-inflammatory and anti-hypertriglyceridemia mechanisms in cardiovascular disease. Mol. Cell. Biochem. 2021, 476, 993–1003. [Google Scholar] [CrossRef]
- Jeon, S.-M. Regulation and function of AMPK in physiology and diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef]
- Jeromson, S.; Mackenzie, I.; Doherty, M.K.; Whitfield, P.D.; Bell, G.; Dick, J.; Shaw, A.; Rao, F.V.; Ashcroft, S.P.; Philp, A.; et al. Lipid remodeling and an altered membrane-associated proteome may drive the differential effects of EPA and DHA treatment on skeletal muscle glucose uptake and protein accretion. Am. J. Physiol. Endocrinol. Metab. 2018, 314, e605–e619. [Google Scholar] [CrossRef]
- Levental, K.R.; Lorent, J.H.; Lin, X.; Skinkle, A.D.; Surma, M.A.; Stockenbojer, E.A.; Gorfe, A.A.; Levental, I. Polyunsaturated Lipids Regulate Membrane Domain Stability by Tuning Membrane Order. Biophys. J. 2016, 110, 1800–1810. [Google Scholar] [CrossRef]
- Bikle, D.D. Vitamin D metabolism, mechanism of action, and clinical applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef]
- Sepidarkish, M.; Farsi, F.; Akbari-Fakhrabadi, M.; Namazi, N.; Almasi-Hashiani, A.; Maleki Hagiagha, A.; Heshmati, J. The effect of vitamin D supplementation on oxidative stress parameters: A systematic review and meta-analysis of clinical trials. Pharmacol. Res. 2019, 139, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liao, Q.; Li, D.; Chen, L.; Zhang, H.; Yi, B. Vitamin D receptor alleviates lipid peroxidation in diabetic nephropathy by regulating ACLY/Nrf2/Keap1 pathway. FASEB J. 2024, 38, e70060. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | SVDD–LHM | VDD–HHM | p-Value * |
|---|---|---|---|
| n | 18 | 28 | |
| 25(OH)D, ng/mL | 8.4 [5.110–9.8] | 13.0 [10.5–26.9] | 0.000 |
| Heavy metals | 127.5 [37.0–199.0] | 255.50 [208.0–330.0] | 0.000 |
| Age, year | 42.0 [32.0–60.0] | 53.0 [32.0–65.0] | 0.001 |
| Male, n | 6 | 11 | 0.761 |
| Smoking, n | 1 | 6 | 0.417 |
| Drinking, n | 11 | 13 | 0.378 |
| Hypertension, n | 5 | 7 | 0.834 |
| BMI | 24.1 [17.9–37.4] | 25.4 [21.5–32.0] | 0.862 |
| ALT | 20.0 [12.0–103.0] | 20.5 [12.0–128.0] | 0.397 |
| AST | 23.0 [14.0–69.0] | 25.5 [16.0–105.0] | 0.295 |
| γ-GTP | 19.0 [9.0–105.0] | 24.5 [9.0–129.0] | 0.710 |
| Uric acid | 4.6 [2.3–7.4] | 5.1 [2.1–7.5] | 0.178 |
| hs-CRP | 0.6 [0.2–2.6] | 0.4 [0.1–4.3] | 0.362 |
| HbA1c | 5.4 [5.0–5.7] | 5.5 [4.8–6.1] | 0.260 |
| Total cholesterol | 184.5 [134.0–313.0] | 195.0 [151.0–236.0] | 0.831 |
| HDL-cholesterol | 57.5 [34.0–85.0] | 53.5 [29.0–94.0] | 0.860 |
| TGs | 96.0 [34.0–536.0] | 108.5 [3.9–348.0] | 0.770 |
| VIP Score | RT | m/z | Name | HMDB ID | Related Pathway | p-Value * | Trend |
|---|---|---|---|---|---|---|---|
| 2.47 | 10.27 | 329.2478 | Docosahexaenoic acid | HMDB0002183 | Biosynthesis of unsaturated fatty acids | 0.000 | ↑ |
| 2.29 | 16.62 | 802.5380 | PC 20:5_18:3 | HMDB0008501 | Glycerophospholipid metabolism | 0.002 | ↑ |
| 2.25 | 13.02 | 577.4838 | DAG 15:0_18:3 | HMDB0007075 | Phosphatidylcholine biosynthesis | 0.002 | ↓ |
| 2.02 | 16.13 | 780.5552 | PC 18:2_18:3 | HMDB0008624 | Glycerophospholipid metabolism | 0.004 | ↑ |
| 1.91 | 6.01 | 585.2719 | Bilirubin | HMDB0000054 | Porphyrin metabolism | 0.049 | ↑ |
| 1.55 | 9.83 | 303.2309 | Eicosapentaenoic acid | HMDB0001999 | Biosynthesis of unsaturated fatty acids | 0.001 | ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.S.; Gwon, M.-R.; Lee, J.H.; Park, J.J.; Lee, H.W.; Lee, D.-H.; Seong, S.J.; Yoon, Y.-R. Impact of Heavy Metals on the Antioxidant Activity of Vitamin D: A Metabolic Perspective. Metabolites 2025, 15, 440. https://doi.org/10.3390/metabo15070440
Park JS, Gwon M-R, Lee JH, Park JJ, Lee HW, Lee D-H, Seong SJ, Yoon Y-R. Impact of Heavy Metals on the Antioxidant Activity of Vitamin D: A Metabolic Perspective. Metabolites. 2025; 15(7):440. https://doi.org/10.3390/metabo15070440
Chicago/Turabian StylePark, Ji Seo, Mi-Ri Gwon, Jae Hwa Lee, Jin Ju Park, Hae Won Lee, Duk-Hee Lee, Sook Jin Seong, and Young-Ran Yoon. 2025. "Impact of Heavy Metals on the Antioxidant Activity of Vitamin D: A Metabolic Perspective" Metabolites 15, no. 7: 440. https://doi.org/10.3390/metabo15070440
APA StylePark, J. S., Gwon, M.-R., Lee, J. H., Park, J. J., Lee, H. W., Lee, D.-H., Seong, S. J., & Yoon, Y.-R. (2025). Impact of Heavy Metals on the Antioxidant Activity of Vitamin D: A Metabolic Perspective. Metabolites, 15(7), 440. https://doi.org/10.3390/metabo15070440