
Metabolic Profiling of SH-SY5Y and Neuro2A Cells in Relation to Fetal Calf Serum (FCS) Concentration in Culture Media

Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Quantification of Glucose, Pyruvate, L-Glutamine, and Lactate in Culture Media Using Amplex Red Assays
2.3. Assessment of Cellular Energy Metabolism Utilizing the Seahorse XF Real-Time ATP Rate Assay
2.4. Determination of the Cellular L-Glutamine Demand Utilizing the Seahorse XF Substrate Oxidation Stress Test
2.5. In-Situ Cell Counting Using a BioTek Cytation 5
2.6. Calculation of Glucose Utilization, Pyruvate, and L-Glutamine Consumption, and Lactate Production Rates
2.7. Calculation of Total Aerobic and Anaerobic ATP Rate
2.8. Calculation of the Cellular L-Glutamine Demand
2.9. Statistical Analysis
3. Results
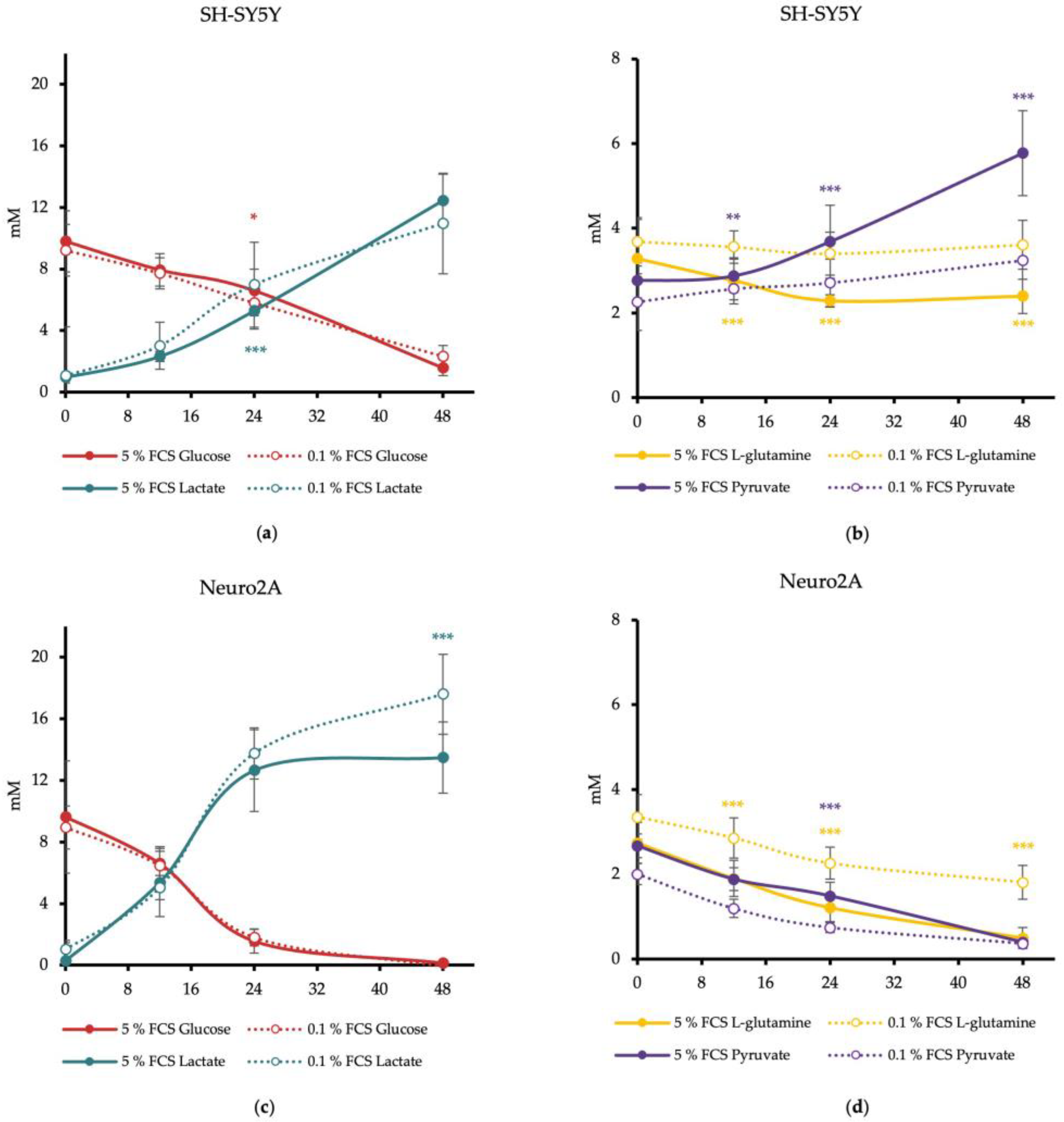
3.1. Energy Substrate Consumption of Neuro2A and SH-SY5Y Cells Depends on the FCS Concentration in the Cell Culture Medium
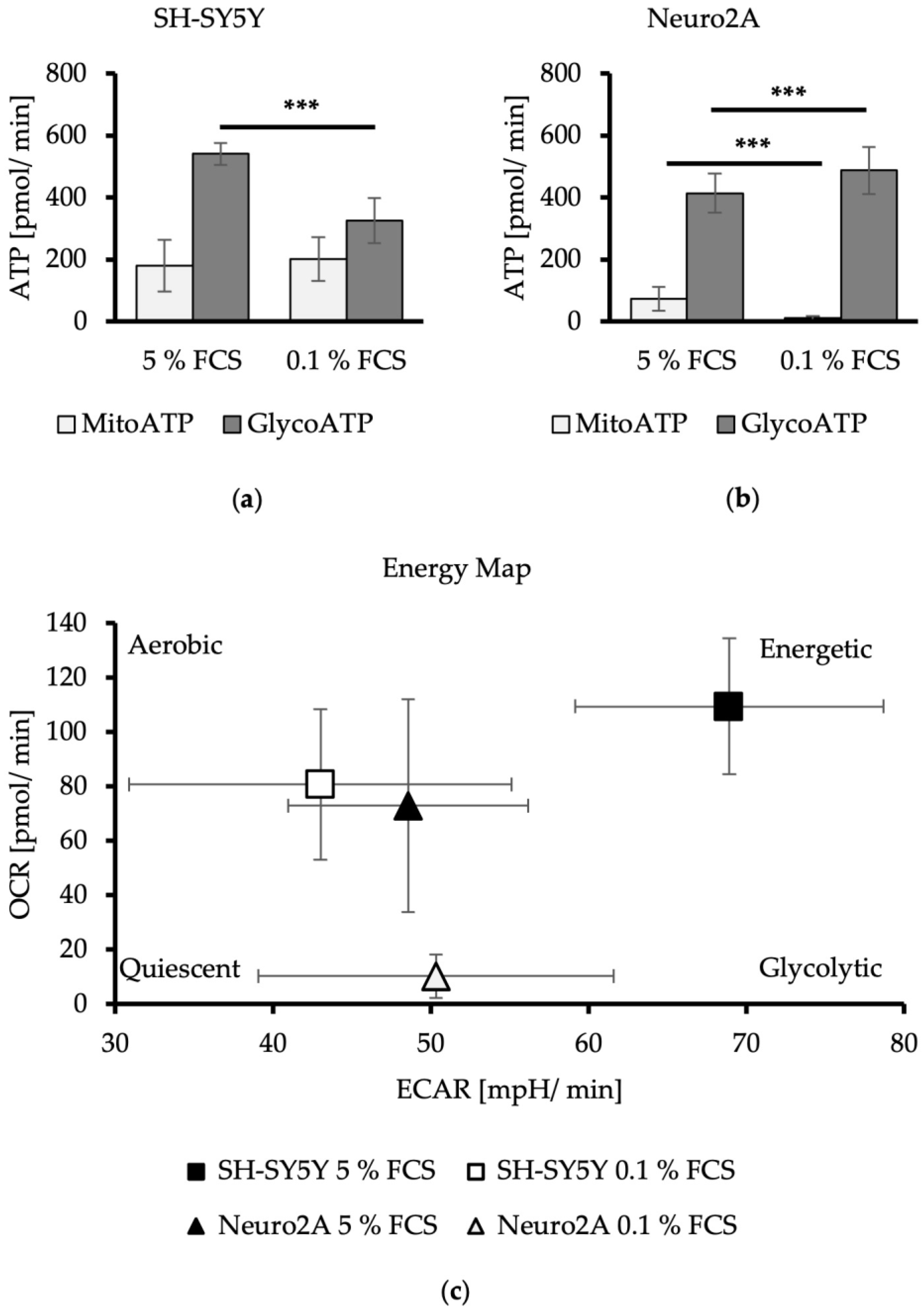
3.2. FCS Reduction Leads to a Switch in the Energy Production Strategy of Neuroblastoma Cell Lines
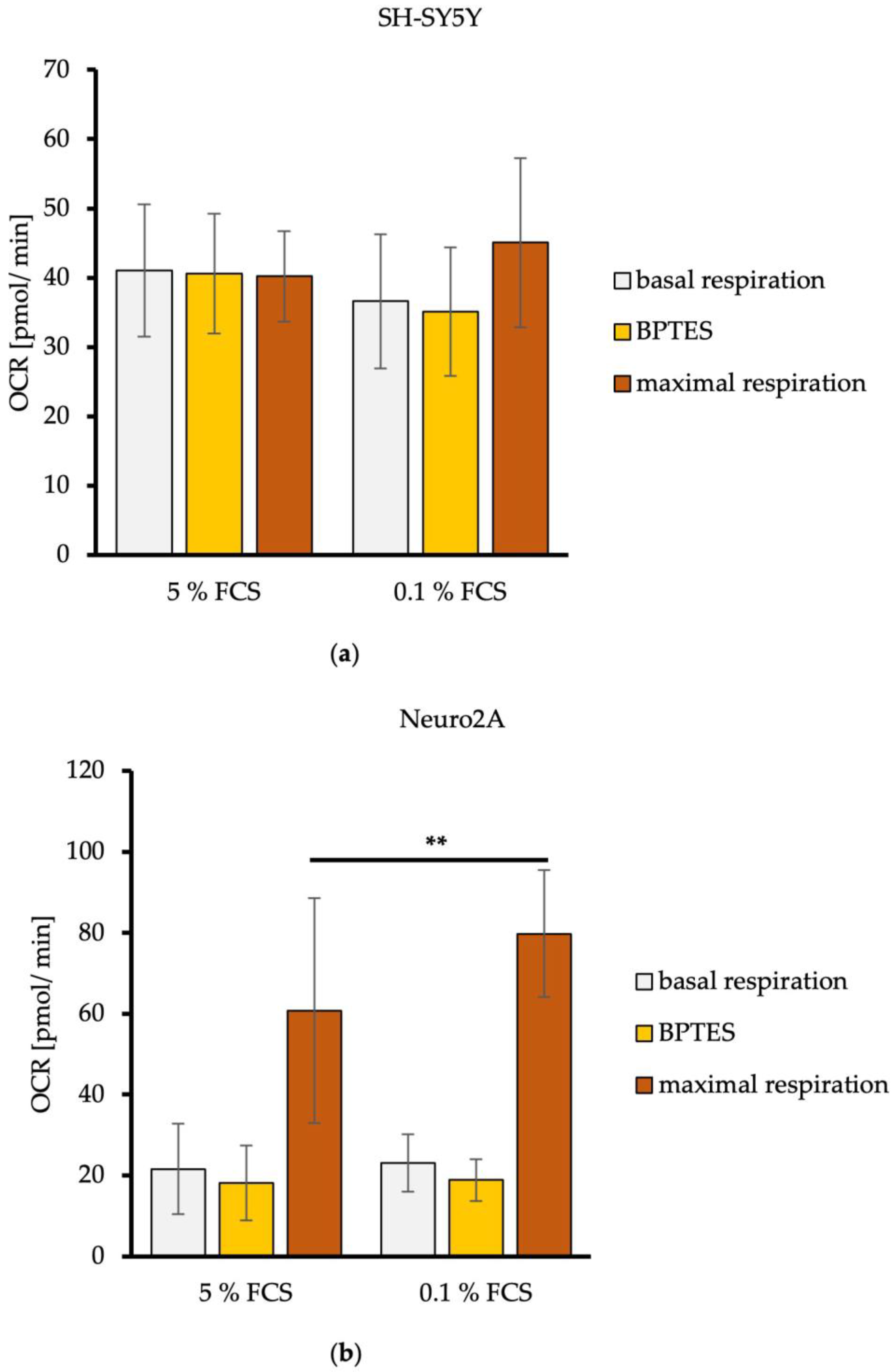
3.3. Neuro2A Cells Prefer L-Glutamine as an Alternative Energy Substrate after Reducing FCS in the Cell Culture Medium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McKee, T.J.; Komarova, S.V. Is It Time to Reinvent Basic Cell Culture Medium? Am. J. Physiol. Cell Physiol. 2017, 312, C624–C626. [Google Scholar] [CrossRef] [PubMed]
- Camandola, S.; Mattson, M.P. Brain Metabolism in Health, Aging, and Neurodegeneration. EMBO J. 2017, 36, 1474–1492. [Google Scholar] [CrossRef] [PubMed]
- Mergenthaler, P.; Lindauer, U.; Dienel, G.A.; Meisel, A. Sugar for the Brain: The Role of Glucose in Physiological and Pathological Brain Function. Trends Neurosci. 2013, 36, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Yako, H.; Niimi, N.; Kato, A.; Takaku, S.; Tatsumi, Y.; Nishito, Y.; Kato, K.; Sango, K. Role of Pyruvate in Maintaining Cell Viability and Energy Production under High-Glucose Conditions. Sci. Rep. 2021, 11, 18910. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Ko, B.; Hensley, C.T.; Jiang, L.; Wasti, A.T.; Kim, J.; Sudderth, J.; Calvaruso, M.A.; Lumata, L.; Mitsche, M.; et al. Glutamine Oxidation Maintains the TCA Cycle and Cell Survival during Impaired Mitochondrial Pyruvate Transport. Mol. Cell 2014, 56, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Klebe, R.J.; Chen, T.-R.; Ruddle, F.H. Controlled production of proliferating somatic cell hybrids. J. Cell Biol. 1970, 45, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.A.; Spengler, B.A.; Biedler, J.L. Coordinate Morphological and Biochemical Interconversion of Human Neuroblastoma Cells. J. Natl. Cancer Inst. 1983, 71, 741–747. [Google Scholar] [PubMed]
- Biedler, J.L.; Helson, L.; Spengler, B.A. Morphology and Growth, Tumorigenicity, and Cytogenetics of Human Neuroblastoma Cells in Continuous Culture. Cancer Res. 1973, 33, 2643–2652. [Google Scholar]
- Biedler, J.L.; Roffler-Tarlov, S.; Schachner, M.; Freedman, L.S. Multiple Neurotransmitter Synthesis by Human Neuroblastoma Cell Lines and Clones. Cancer Res. 1978, 38 Pt 1, 3751–3757. [Google Scholar] [PubMed]
- Kovalevich, J.; Langford, D. Considerations for the Use of SH-SY5Y Neuroblastoma Cells in Neurobiology. Methods Mol. Biol. 2013, 1078, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Heiden, M.G.V.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of Metabolism and Mitochondrial Homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef]
- Barone, E.; Di Domenico, F.; Perluigi, M.; Butterfield, D.A. The Interplay among Oxidative Stress, Brain Insulin Resistance and AMPK Dysfunction Contribute to Neurodegeneration in Type 2 Diabetes and Alzheimer Disease. Free Radic. Biol. Med. 2021, 176, 16–33. [Google Scholar] [CrossRef] [PubMed]
- Sakagami, H.; Suzuki, R.; Shirataki, Y.; Iwama, S.; Nakagawa, M.; Suzuki, H.; Tanaka, K.; Tamura, N.; Takeshima, H. Re-Evaluation of Culture Condition of PC12 and SH-SY5Y Cells Based on Growth Rate and Amino Acid Consumption. In Vivo 2017, 31, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Gardner, G.L.; Moradi, F.; Moffatt, C.; Cliche, M.; Garlisi, B.; Gratton, J.; Mehmood, F.; Stuart, J.A. Rapid Nutrient Depletion to below the Physiological Range by Cancer Cells Cultured in Plasmax. Am. J. Physiol. Cell Physiol. 2022, 323, C823–C834. [Google Scholar] [CrossRef] [PubMed]
- Xun, Z.; Lee, D.Y.; Lim, J.; Canaria, C.A.; Barnebey, A.; Yanonne, S.M.; McMurray, C.T. Retinoic Acid-Induced Differentiation Increases the Rate of Oxygen Consumption and Enhances the Spare Respiratory Capacity of Mitochondria in SH-SY5Y Cells. Mech. Ageing Dev. 2012, 133, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Lee, S.Y.; Yun, S.H.; Jeong, J.W.; Kim, J.H.; Kim, H.W.; Choi, J.S.; Kim, G.D.; Joo, S.T.; Choi, I.; et al. Review of the Current Research on Fetal Bovine Serum and the Development of Cultured Meat. Food Sci. Anim. Resour. 2022, 42, 775–799. [Google Scholar] [CrossRef] [PubMed]
- Mo, W.; Zhang, Z.; Liu, Y.; Bartlett, P.F.; He, R. Magnetic Shielding Accelerates the Proliferation of Human Neuroblastoma Cell by Promoting G1-Phase Progression. PLoS ONE 2013, 8, e54775. [Google Scholar] [CrossRef] [PubMed]
- Viallon, J.; Chinain, M.; Darius, H.T. Revisiting the Neuroblastoma Cell-Based Assay (CBA-N2a) for the Improved Detection of Marine Toxins Active on Voltage Gated Sodium Channels (VGSCs). Toxins 2020, 12, 281. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.; Romero, R.; Petty, H.R. A Sensitive Fluorimetric Assay for Pyruvate. Anal. Biochem. 2010, 396, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Watabe, S.; Sakamoto, Y.; Morikawa, M.; Okada, R.; Miura, T.; Ito, E. Highly Sensitive Determination of Hydrogen Peroxide and Glucose by Fluorescence Correlation Spectroscopy. PLoS ONE 2011, 6, e22955. [Google Scholar] [CrossRef] [PubMed]
- Muller, B.; Lewis, N.; Adeniyi, T.; Leese, H.J.; Brison, D.R.; Sturmey, R.G. Application of Extracellular Flux Analysis for Determining Mitochondrial Function in Mammalian Oocytes and Early Embryos. Sci. Rep. 2019, 9, 16778. [Google Scholar] [CrossRef] [PubMed]
- Sprague, J.E.; María Arbeláez, A. Glucose Counterregulatory Responses to Hypoglycemia. Pediatr. Endocrinol. Rev. 2011, 9, 463–475. [Google Scholar] [PubMed]
- Msaouel, P.; Malouf, G.G.; Tannir, N.M. Metabolic Derangements in Succinate Dehydrogenase B–Mutated Renal-Cell Carcinomas: More Than Meets the Eye? JCO Precis. Oncol. 2017, 1, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Fendt, S.M.; Verstreken, P. Neurons Eat Glutamate to Stay Alive. J. Cell Biol. 2017, 216, 863–865. [Google Scholar] [CrossRef] [PubMed]
- Mazat, J.P.; Ransac, S. The Fate of Glutamine in Human Metabolism. The Interplay with Glucose in Proliferating Cells. Metabolites 2019, 9, 81. [Google Scholar] [CrossRef] [PubMed]
- Divakaruni, A.S.; Jastroch, M. A Practical Guide for the Analysis, Standardization and Interpretation of Oxygen Consumption Measurements. Nat. Metab. 2022, 4, 978–994. [Google Scholar] [CrossRef] [PubMed]
- Deberardinis, R.J.; Mancuso, A.; Daikhin, E.; Nissim, I.; Yudkoff, M.; Wehrli, S.; Thompson, C.B. Beyond Aerobic Glycolysis: Transformed Cells Can Engage in Glutamine Metabolism That Exceeds the Requirement for Protein and Nucleotide Synthesis. Proc. Natl. Acad. Sci. USA 2007, 104, 19345–19350. [Google Scholar] [CrossRef] [PubMed]
- Voorde, J.V.; Ackermann, T.; Pfetzer, N.; Sumpton, D.; Mackay, G.; Kalna, G.; Nixon, C.; Blyth, K.; Gottlieb, E.; Tardito, S. CANCER. Improving the Metabolic Fidelity of Cancer Models with a Physiological Cell Culture Medium. Sci. Adv. 2019, 5, eaau7314. [Google Scholar] [CrossRef] [PubMed]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does It Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef]
- Currie, E.; Schulze, A.; Zechner, R.; Walther, T.C.; Farese, R.V. Cellular Fatty Acid Metabolism and Cancer. Cell Metab. 2013, 18, 153–161. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| SH-SY5Y | Neuro2A | |||
|---|---|---|---|---|
| [fmol/cell/h] | 5% FCS | 0.1% FCS | 5% FCS | 0.1% FCS |
| Glucose Consumption Rate | 836.60 | 563.02 | 2093.72 | 2290.65 |
| Pyruvate Consumption Rate | −239.75 | −74.14 | 348.24 | 405.42 |
| L-Glutamine Consumption Rate | 152.00 | 46.69 | 239.85 | 180.18 |
| Lactate Production Rate | 1130.72 | 1349.63 | 3629.42 | 4137.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kronenberger, L.; Mett, J.; Hoppstädter, J.; Müller, U. Metabolic Profiling of SH-SY5Y and Neuro2A Cells in Relation to Fetal Calf Serum (FCS) Concentration in Culture Media. Metabolites 2024, 14, 188. https://doi.org/10.3390/metabo14040188
Kronenberger L, Mett J, Hoppstädter J, Müller U. Metabolic Profiling of SH-SY5Y and Neuro2A Cells in Relation to Fetal Calf Serum (FCS) Concentration in Culture Media. Metabolites. 2024; 14(4):188. https://doi.org/10.3390/metabo14040188
Chicago/Turabian StyleKronenberger, Lys, Janine Mett, Jessica Hoppstädter, and Uli Müller. 2024. "Metabolic Profiling of SH-SY5Y and Neuro2A Cells in Relation to Fetal Calf Serum (FCS) Concentration in Culture Media" Metabolites 14, no. 4: 188. https://doi.org/10.3390/metabo14040188
APA StyleKronenberger, L., Mett, J., Hoppstädter, J., & Müller, U. (2024). Metabolic Profiling of SH-SY5Y and Neuro2A Cells in Relation to Fetal Calf Serum (FCS) Concentration in Culture Media. Metabolites, 14(4), 188. https://doi.org/10.3390/metabo14040188