
Proline as a Sparker Metabolite of Oxidative Metabolism during the Flight of the Bumblebee, Bombus impatiens


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
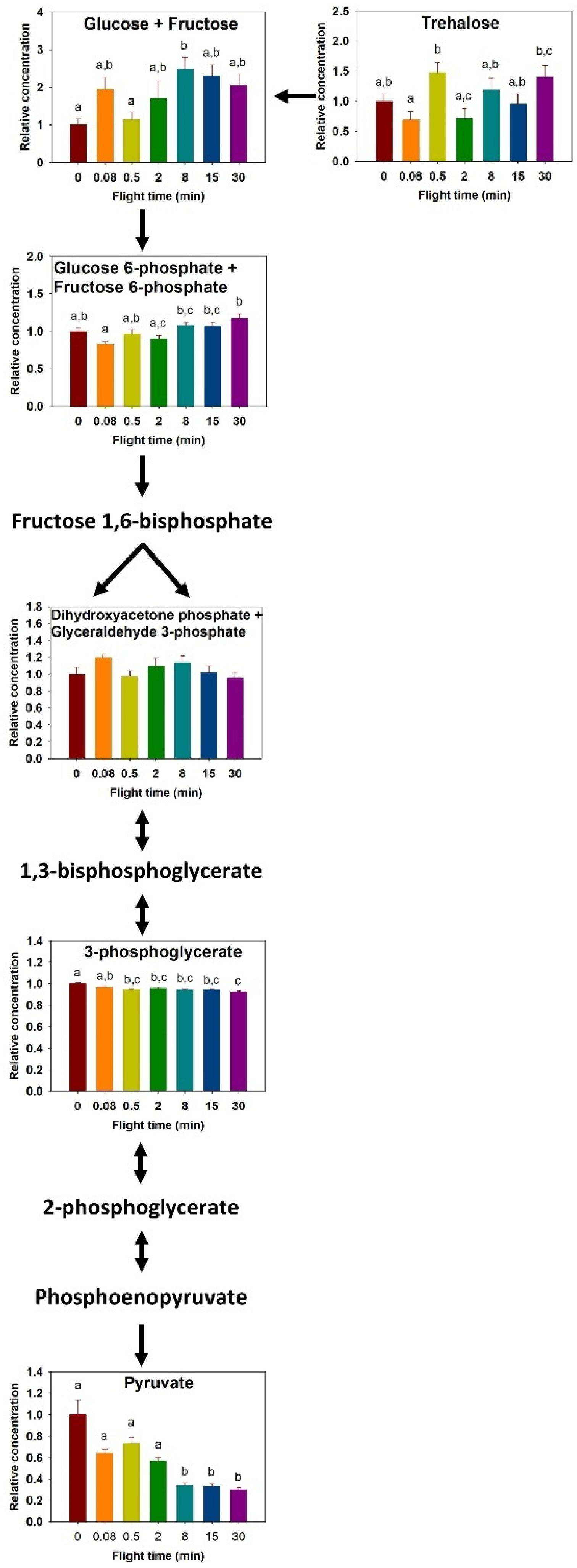
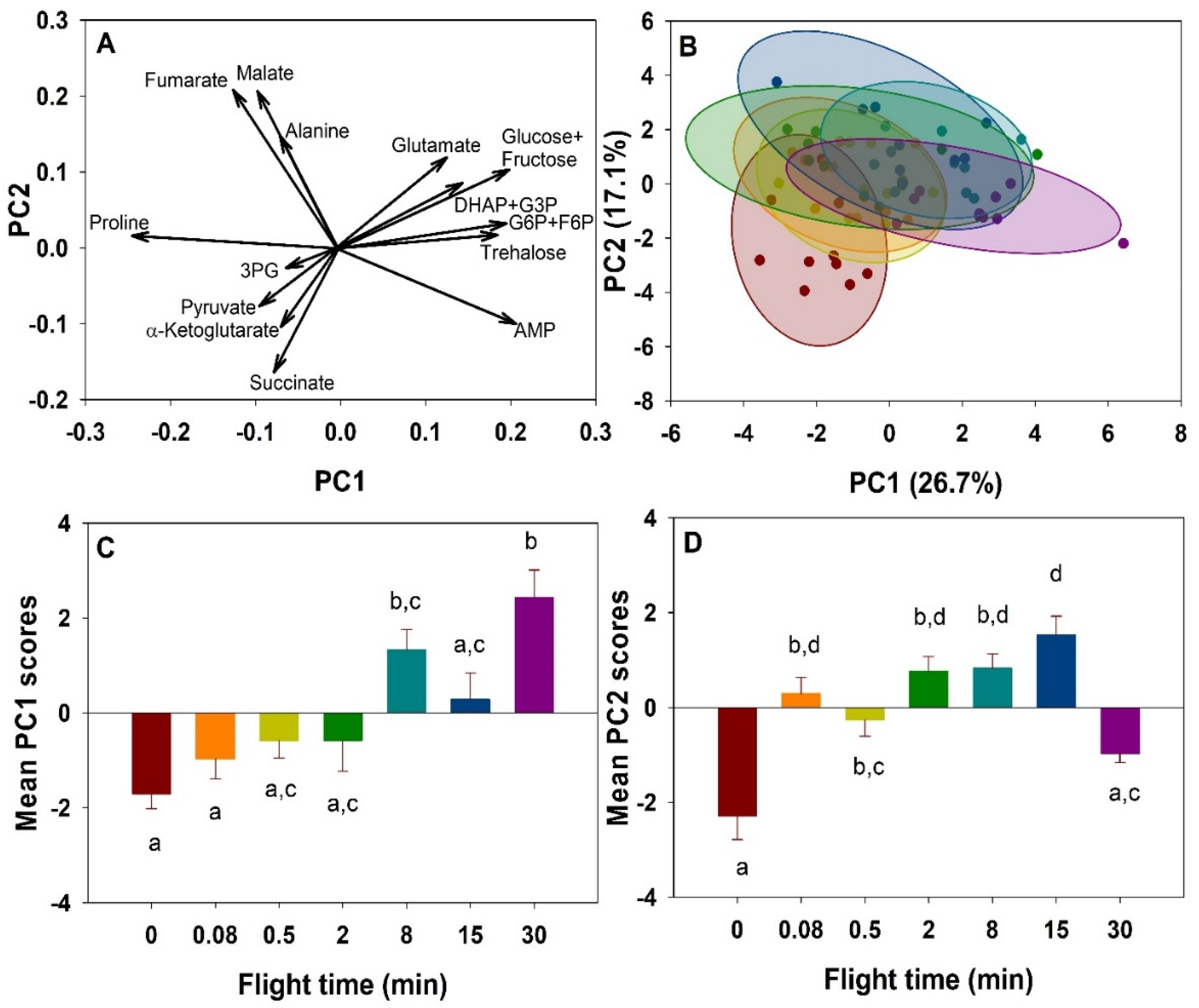
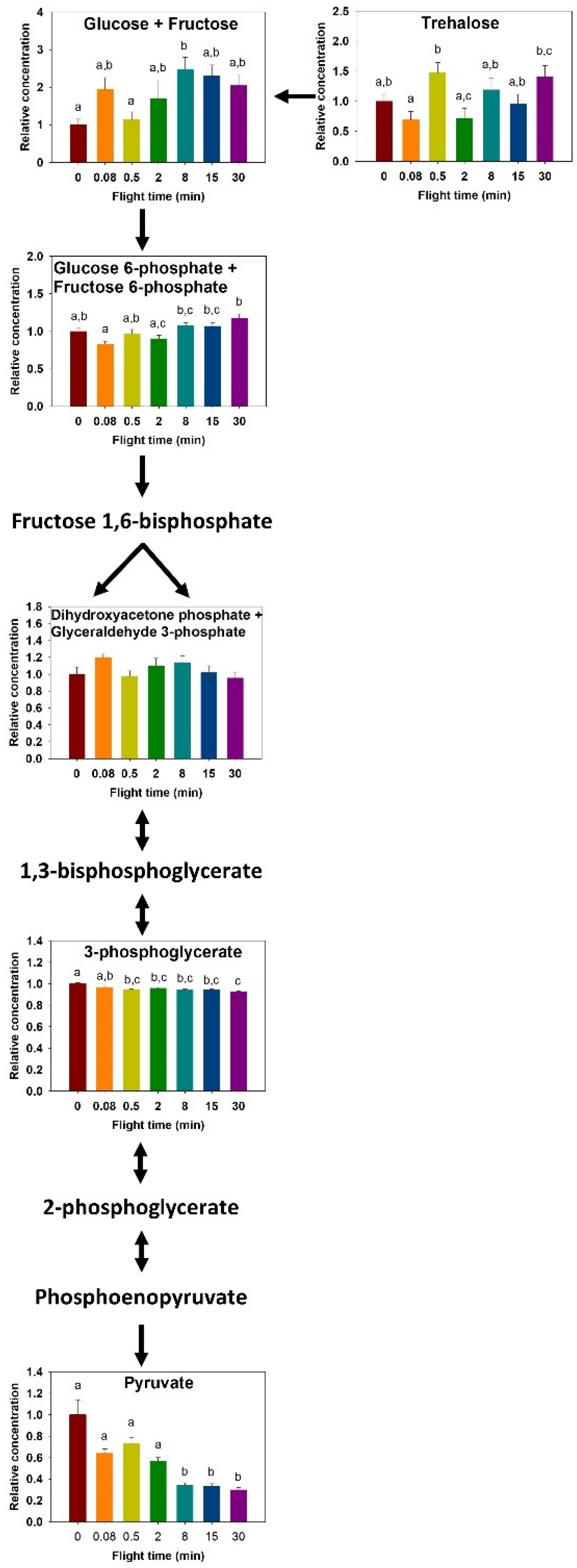
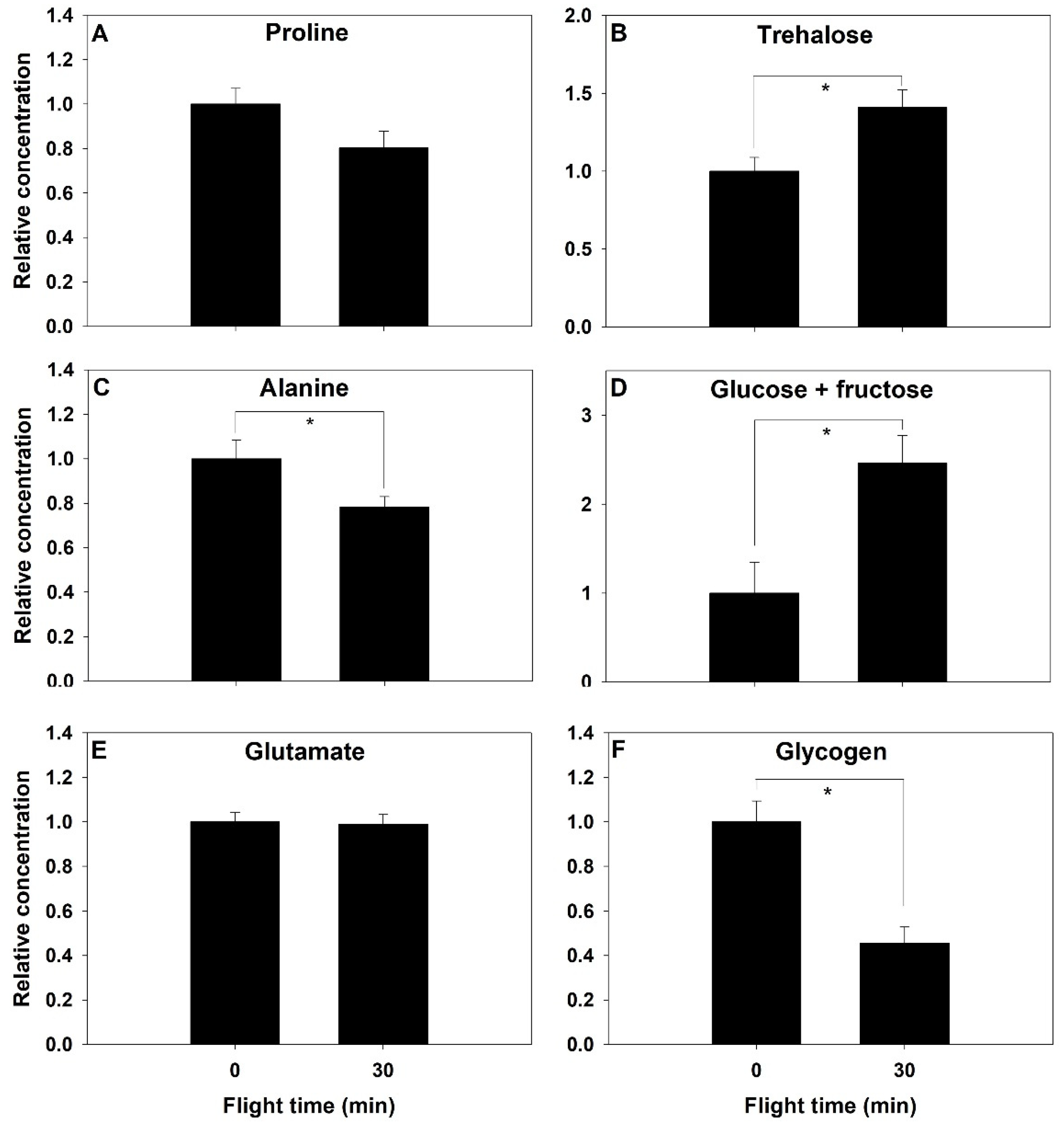
2. Results
2.1. Metabolites Profile Changes
2.2. Proline Metabolism
2.3. Carbohydrate Metabolism
2.4. TCA Cycle
2.5. Abdominal Metabolites Content
3. Discussion
4. Materials and Methods
4.1. Flight Experiments
4.2. Preparation and Detection of Analytical Standards
4.3. Preparation of Samples
4.4. Ultra-High Performance Liquid Chromatography-Electrospray Ionization-Quadrupole Time of Flight Mass Spectrometry (UPLC-ESI-QTOF MS)
4.5. Determination of Glycogen Content in Abdomens
4.6. Data Collection and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanderhorst, D.J.; Houben, N.M.D.; Beenakkers, A.M.T. Dynamics of energy substrates in the hemolymph of Locusta migratoria during flight. J. Insect Physiol. 1980, 26, 441–448. [Google Scholar] [CrossRef]
- Auerswald, L.; Gade, G. The fate of proline in the African fruit beetle Pachnoda sinuata. Insect Biochem. Mol. Biol. 1999, 29, 687–700. [Google Scholar] [CrossRef]
- Kammer, A.E.; Heinrich, B. Insect Flight Metabolism. In Advances in Insect Physiology; Treherne, J.E., Berridge, M.J., Wigglesworth, V.B., Eds.; Academic Press: New York, NY, USA, 1978; pp. 133–228. [Google Scholar]
- Bursell, E. Substrates of oxidative-metabolism in dipteran flight-muscle. Comp. Biochem. Physiol. B 1975, 52, 235–238. [Google Scholar] [CrossRef]
- Soares, J.; Gaviraghi, A.; Oliveira, M.F. Mitochondrial Physiology in the Major Arbovirus Vector Aedes aegypti: Substrate Preferences and Sexual Differences Define Respiratory Capacity and Superoxide Production. PLoS ONE 2015, 10, e0120600. [Google Scholar] [CrossRef] [Green Version]
- Storey, K.B. Metabolic Biochemistry of Insect Flight; Springer: Berlin/Heidelberg, Germany, 1985; pp. 193–207. [Google Scholar]
- Sacktor, B. Biochemistry of insect flight—Utilization of fuels by muscle. In Insect Biochemistry and Function; Candy, D.J., Kilby, B.A., Eds.; Chapman and Hall: London, UK, 1975; pp. 1–81. [Google Scholar]
- Johnson, R.N.; Hansford, R.G. The control of tricarboxylate-cycle oxidations in blowfly flight muscle—The steady-state concentrations of citrate, isocitrate 2-oxoglutarate and malate in flight muscle and isolated mitochondria. Biochem. J. 1975, 146, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Weeda, E.; Dekort, C.A.D.; Beenakkers, A.M.T. Fuels for energy metabolism in the Colorado potato beetle, Leptinotarsa decemlineata Say. J. Insect Physiol. 1979, 25, 951–955. [Google Scholar] [CrossRef]
- Zebe, E.; Gade, G. Flight metabolism in the African fruit beetle, Pachnoda sinuata. J. Comp. Physiol. B 1993, 163, 107–112. [Google Scholar] [CrossRef]
- Candy, D.J.; Kilby, B.A. Insect Biochemistry and Function; Springer: Boston, MA, USA, 1975; p. 314. [Google Scholar]
- Weber, J.M. Metabolic fuels: Regulating fluxes to select mix. J. Exp. Biol. 2011, 214, 286–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gade, G.; Auerswald, L. Beetles’ choice—Proline for energy output: Control by AKHs. Comp. Biochem. Physiol. B 2002, 132, 117–129. [Google Scholar] [CrossRef]
- Bursell, E. Synthesis of proline by fat body of the tsetse fly (Glossina morsitans): Metabolic pathways. Insect Biochem. 1977, 7, 427–434. [Google Scholar] [CrossRef]
- Carter, C.; Shafir, S.; Yehonatan, L.; Palmer, R.G.; Thornburg, R. A novel role for proline in plant floral nectars. Naturwissenschaften 2006, 93, 72–79. [Google Scholar] [CrossRef]
- Barker, R.J.; Lehner, Y. Free amino acids in thoraces of flown honey bees, Apis mellifera L. (Hymenoptera: Apidae). Comp. Biochem. Physiol. 1972, 43, 163–169. [Google Scholar] [CrossRef]
- Micheu, S.; Crailsheim, K.; Leonhard, B. Importance of proline and other amino acids during honeybee flight (Apis mellifera carnica POLLMANN). Amino Acids 2000, 18, 157–175. [Google Scholar] [CrossRef]
- Crailsheim, K.; Leonhard, B. Amino acids in honeybee worker haemolymph. Amino Acids 1997, 13, 141–153. [Google Scholar] [CrossRef]
- Crabtree, B.; Newsholme, E.A. The activities of proline dehydrogenase, glutamate dehydrogenase, aspartate-oxoglutarate aminotransferase and alanine-oxoglutarate aminotransferase in some insect flight muscles. Biochem. J. 1970, 117, 1019–1021. [Google Scholar] [CrossRef] [Green Version]
- Teulier, L.; Weber, J.M.; Crevier, J.; Darveau, C.A. Proline as a fuel for insect flight: Enhancing carbohydrate oxidation in hymenopterans. Proc. R. Soc. B Biol. Sci. 2016, 283, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenbergh, S.G. Pyruvate oxidation and the permeability of housefly sarcosomes. Biochem. J. 1964, 93, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosemer, R.W.; Veerabhadrappa, P.S. Pathway of proline oxidation in insect flight muscle. Biochim. Biophys. Acta 1965, 110, 102–112. [Google Scholar] [CrossRef]
- Sacktor, B.; Childress, C.C. Metabolism of proline in insect flight muscle and its significance in stimulating the oxidation of pyruvate. Arch. Biochem. Biophys. 1967, 120, 583–588. [Google Scholar] [CrossRef]
- Scaraffia, P.Y.; Wells, M.A. Proline can be utilized as an energy substrate during flight of Aedes aegypti females. J. Insect Physiol. 2003, 49, 591–601. [Google Scholar] [CrossRef]
- Weeda, E. Hormonal regulation of proline synthesis and glucose release in the fat body of the Colorado potato beetle, Leptinotarsa decemlineata. J. Insect Physiol. 1981, 27, 411–417. [Google Scholar] [CrossRef]
- Weeda, E.; Dekort, C.A.D.; Beenakkers, A.M.T. Oxidation of proline and pyruvate by flight muscle mitochondria of the Colorado beetle, Leptinotarsa decemlineata say. Insect Biochem. 1980, 10, 305–311. [Google Scholar] [CrossRef]
- Sacktor, B.; Hurlbut, E.C. Regulation of metabolism in working muscle in vivo. II. Concentrations of adenine nucleotides, arginine phosphate, and inorganic phosphate in insect flight muscle during flight. J. Biol. Chem. 1966, 241, 632–634. [Google Scholar] [CrossRef]
- Sacktor, B.; Wormser-Shavit, E. Regulation of metabolism in working muscle in vivo. I. Concentrations of some glycolytic, tricarboxylic acid cycle, and amino acid intermediates in insect flight muscle during flight. J. Biol. Chem. 1966, 241, 624–631. [Google Scholar] [CrossRef]
- Hansford, R.G.; Johnson, R.N. The nature and control of the tricarboxylate cycle in beetle flight muscle. Biochem. J. 1975, 148, 389–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beenakkers, A.M.T.; Vanderhorst, D.J.; Vanmarrewijk, W.J.A. Insect flight muscle metabolism. Insect Biochem. 1984, 14, 243–260. [Google Scholar] [CrossRef]
- Candy, D.J.; Becker, A.; Wegener, G. Coordination and integration of metabolism in insect flight. Comp. Biochem. Physiol. B 1997, 117, 497–512. [Google Scholar] [CrossRef]
- Elliott, J.; Hill, L.; Bailey, E. Changes in tissue carbohydrate content during flight of the fed and starved cockroach, Periplaneta americana L. Comp. Biochem. Physiol. A—Physiol. 1984, 78, 163–165. [Google Scholar] [CrossRef]
- Nayar, J.K.; Van Handel, E. Fuel for sustained mosquito flight. J. Insect Physiol. 1971, 17, 471–481. [Google Scholar] [CrossRef]
- Blatt, J.; Roces, F. Haemolymph sugar levels in foraging honeybees (Apis mellifera carnica): Dependence on metabolic rate and in vivo measurement of maximal rates of trehalose synthesis. J. Exp. Biol. 2001, 204, 2709–2716. [Google Scholar] [CrossRef]
- Rowan, A.N.; Newsholme, E.A. Changes in the contents of adenine nucleotides and intermediates of glycolysis and the citric acid cycle in flight muscle of the locust upon flight and their relationship to the control of the cycle. Biochem. J. 1979, 178, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Wone, B.W.M.; Kinchen, J.M.; Kaup, E.R.; Wone, B. A procession of metabolic alterations accompanying muscle senescence in Manduca sexta. Sci. Rep. 2018, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Van Vranken, J.G.; Bricker, D.K.; Dephoure, N.; Gygi, S.P.; Cox, J.E.; Thummel, C.S.; Rutter, J. SDHAF4 Promotes Mitochondrial Succinate Dehydrogenase Activity and Prevents Neurodegeneration. Cell Metab. 2014, 20, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Vanhandel, E. Rapid determination of glycogen and sugars in mosquitoes. J. Am. Mosq. Control Assoc. 1985, 1, 299–301. [Google Scholar]
- Kaufmann, C.; Brown, M.R. Regulation of carbohydrate metabolism and flight performance by a hypertrehalosaemic hormone in the mosquito Anopheles gambiae. J. Insect Physiol. 2008, 54, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, M.W. Adipokinetic hormone inhibits the formation of energy stores and egg production in the cricket Gryllus bimaculatus. Comp. Biochem. Physiol. B 2003, 136, 197–206. [Google Scholar] [CrossRef]
- Panzenbock, U.; Crailsheim, K. Glycogen in honeybee queens, workers and drones (Apis mellifera carnica Pollm). J. Insect Physiol. 1997, 43, 155–165. [Google Scholar] [CrossRef]
- Kapoore, R.V.; Vaidyanathan, S. Towards quantitative mass spectrometry-based metabolomics in microbial and mammalian systems. Philos. Trans. A Math. Phys. Eng. Sci. 2016, 374, 20150363. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Su, X.; Klein, M.S.; Lewis, I.A.; Fiehn, O.; Rabinowitz, J.D. Metabolite measurement: Pitfalls to avoid and practices to follow. Annu. Rev. Biochem. 2017, 86, 277–304. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Metabolomic data processing, analysis, and interpretation using MetaboAnalyst. Curr. Protoc. Bioinform. 2011, 34, 14.10.1–14.10.48. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stec, N.; Saleem, A.; Darveau, C.-A. Proline as a Sparker Metabolite of Oxidative Metabolism during the Flight of the Bumblebee, Bombus impatiens. Metabolites 2021, 11, 511. https://doi.org/10.3390/metabo11080511
Stec N, Saleem A, Darveau C-A. Proline as a Sparker Metabolite of Oxidative Metabolism during the Flight of the Bumblebee, Bombus impatiens. Metabolites. 2021; 11(8):511. https://doi.org/10.3390/metabo11080511
Chicago/Turabian StyleStec, Nadia, Ammar Saleem, and Charles-A. Darveau. 2021. "Proline as a Sparker Metabolite of Oxidative Metabolism during the Flight of the Bumblebee, Bombus impatiens" Metabolites 11, no. 8: 511. https://doi.org/10.3390/metabo11080511
APA StyleStec, N., Saleem, A., & Darveau, C.-A. (2021). Proline as a Sparker Metabolite of Oxidative Metabolism during the Flight of the Bumblebee, Bombus impatiens. Metabolites, 11(8), 511. https://doi.org/10.3390/metabo11080511