Characterization of Pro- and Anti-Inflammatory Tissue Factors in Rosacea: A Pilot study


Abstract
:1. Introduction
2. Materials and Methods
2.1. Material Characteristics of Subjects
2.2. Immunohistochemical Analysis
2.3. Statistic Analysis
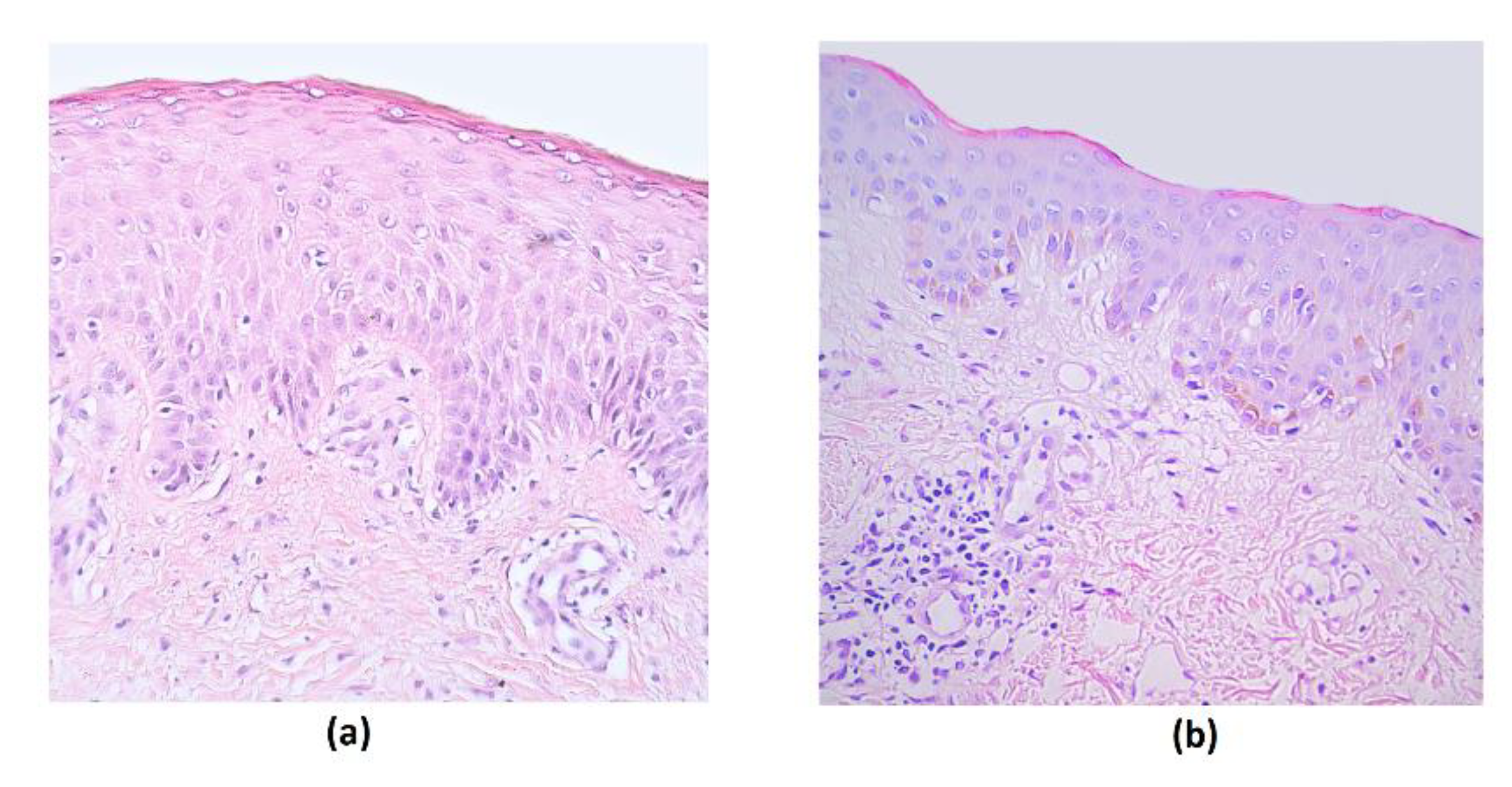
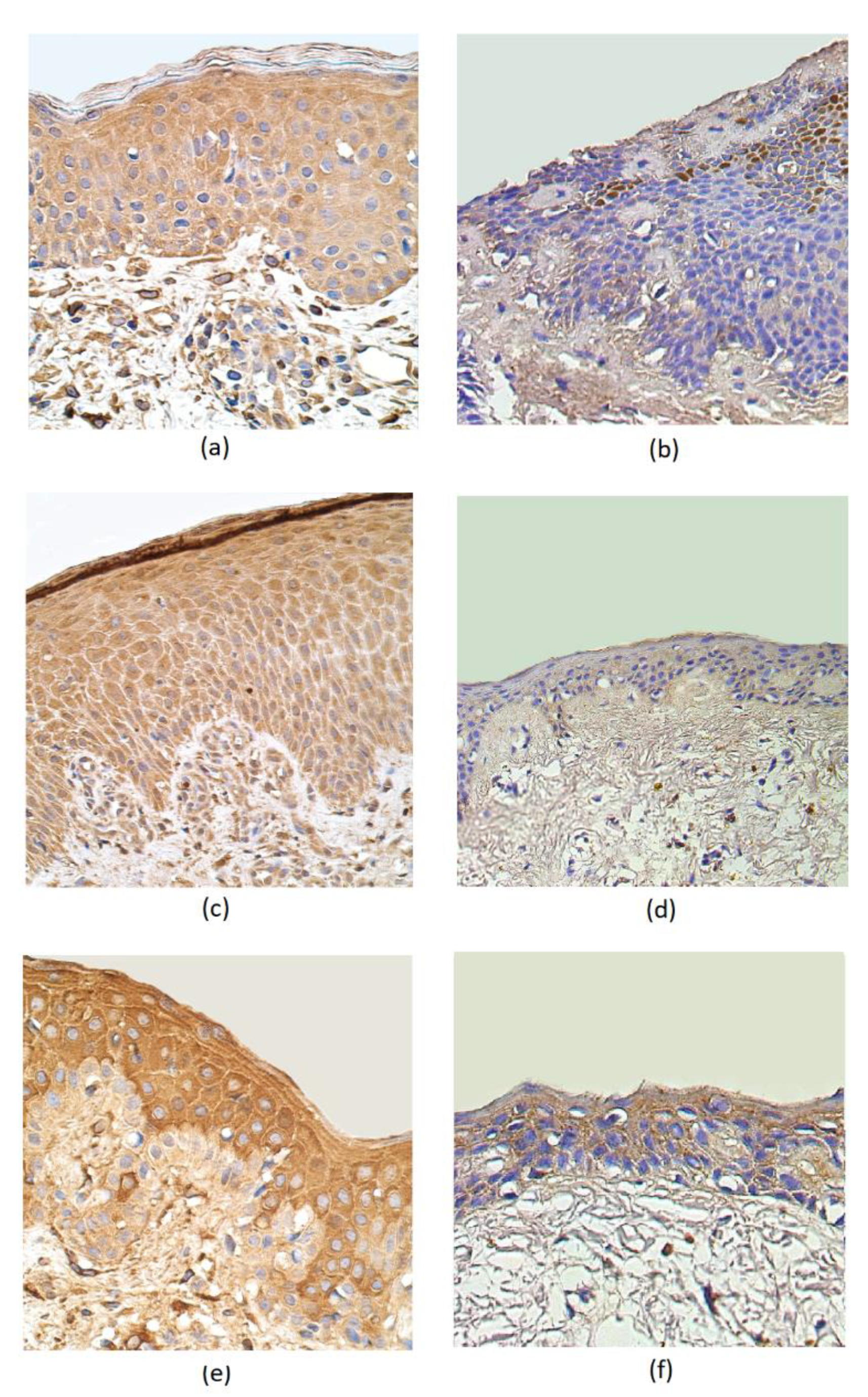
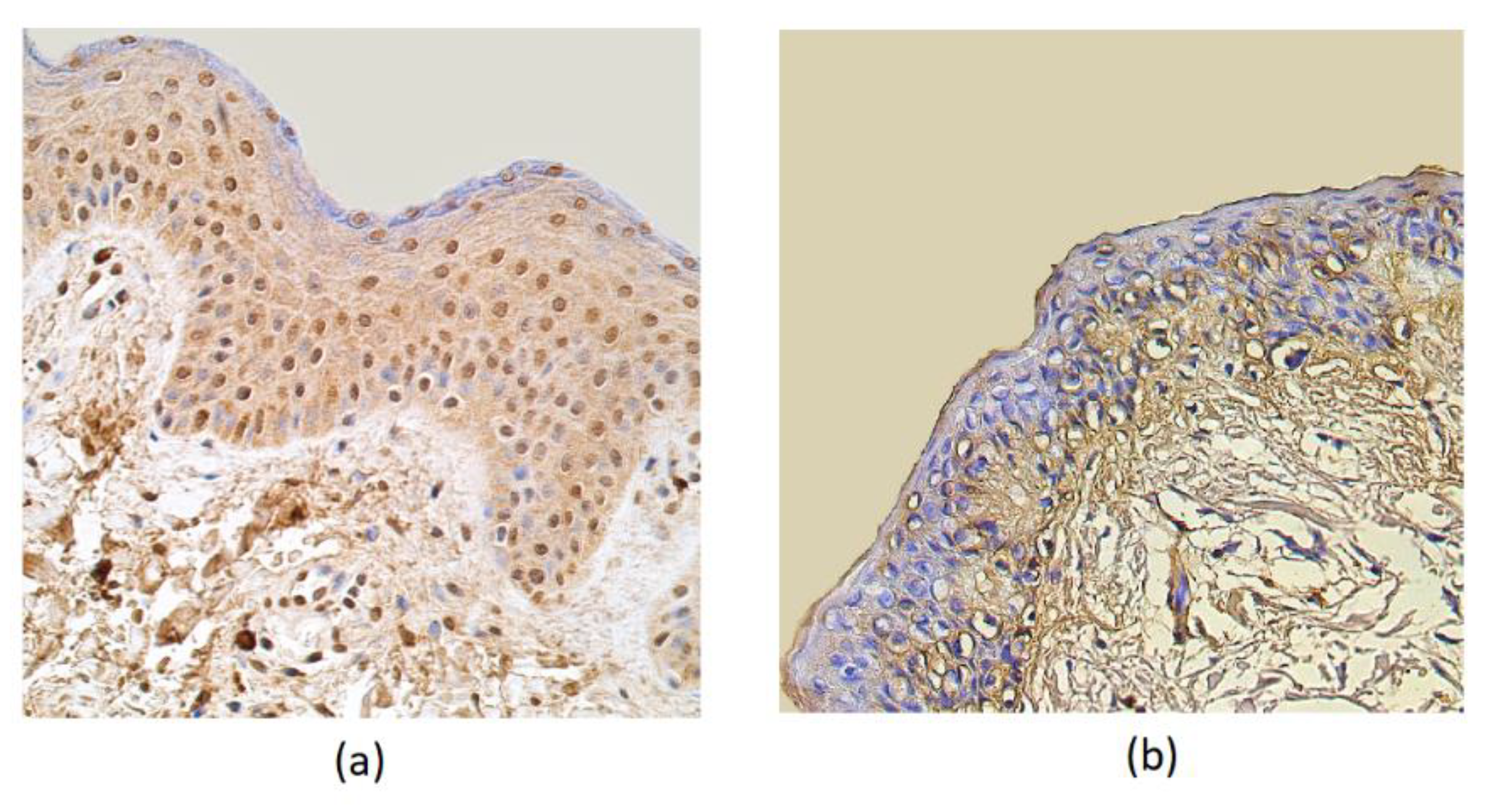
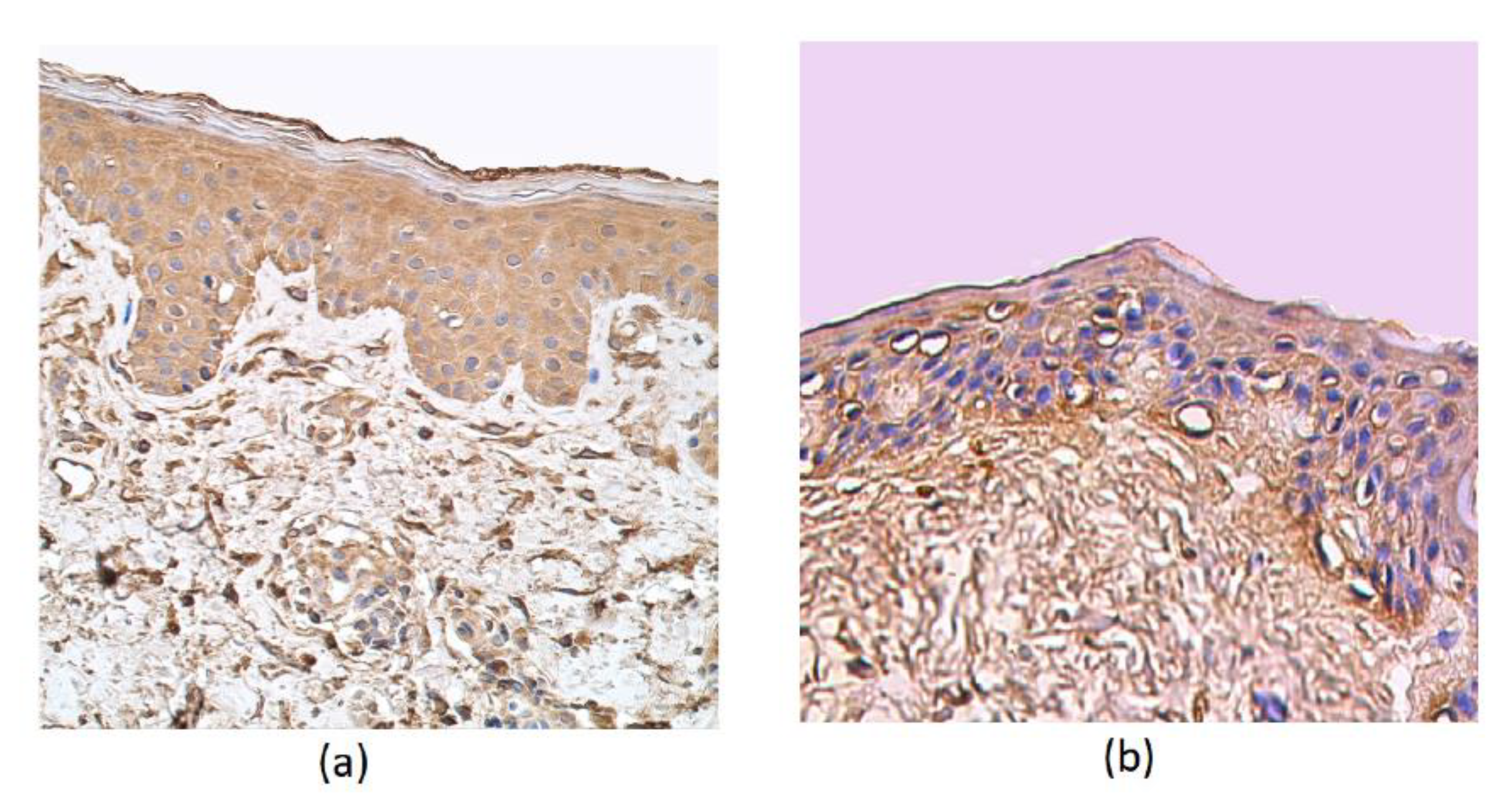
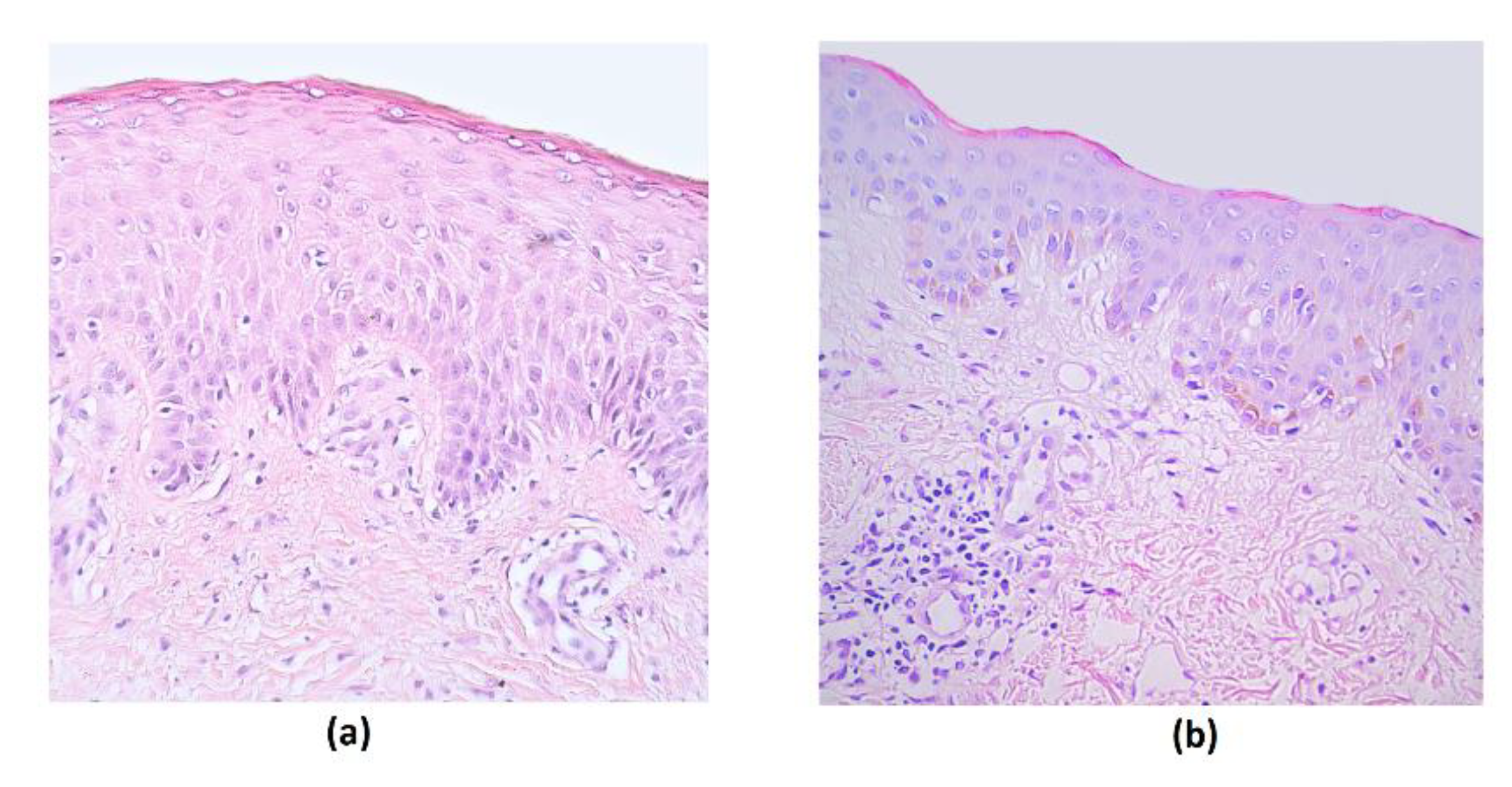
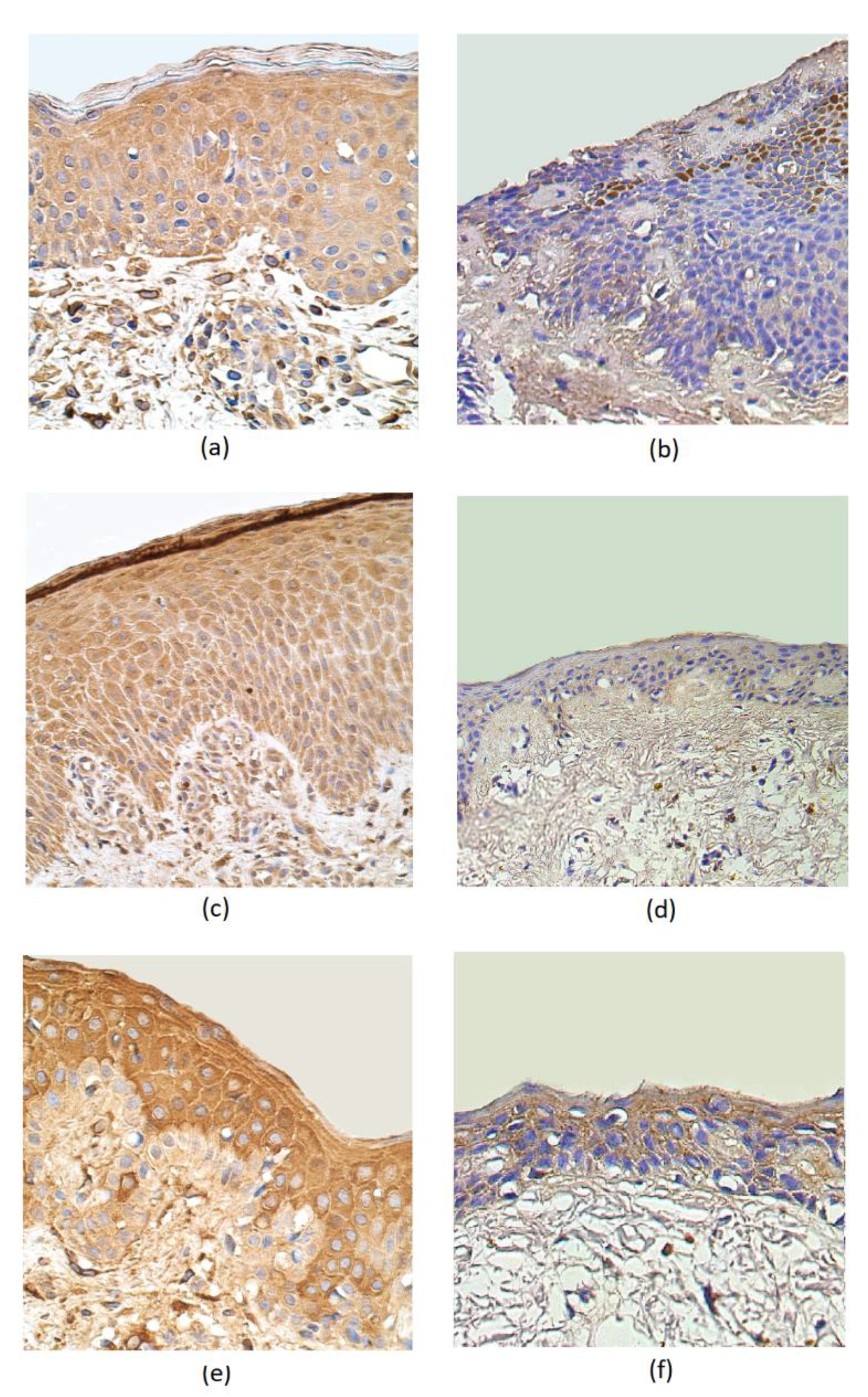
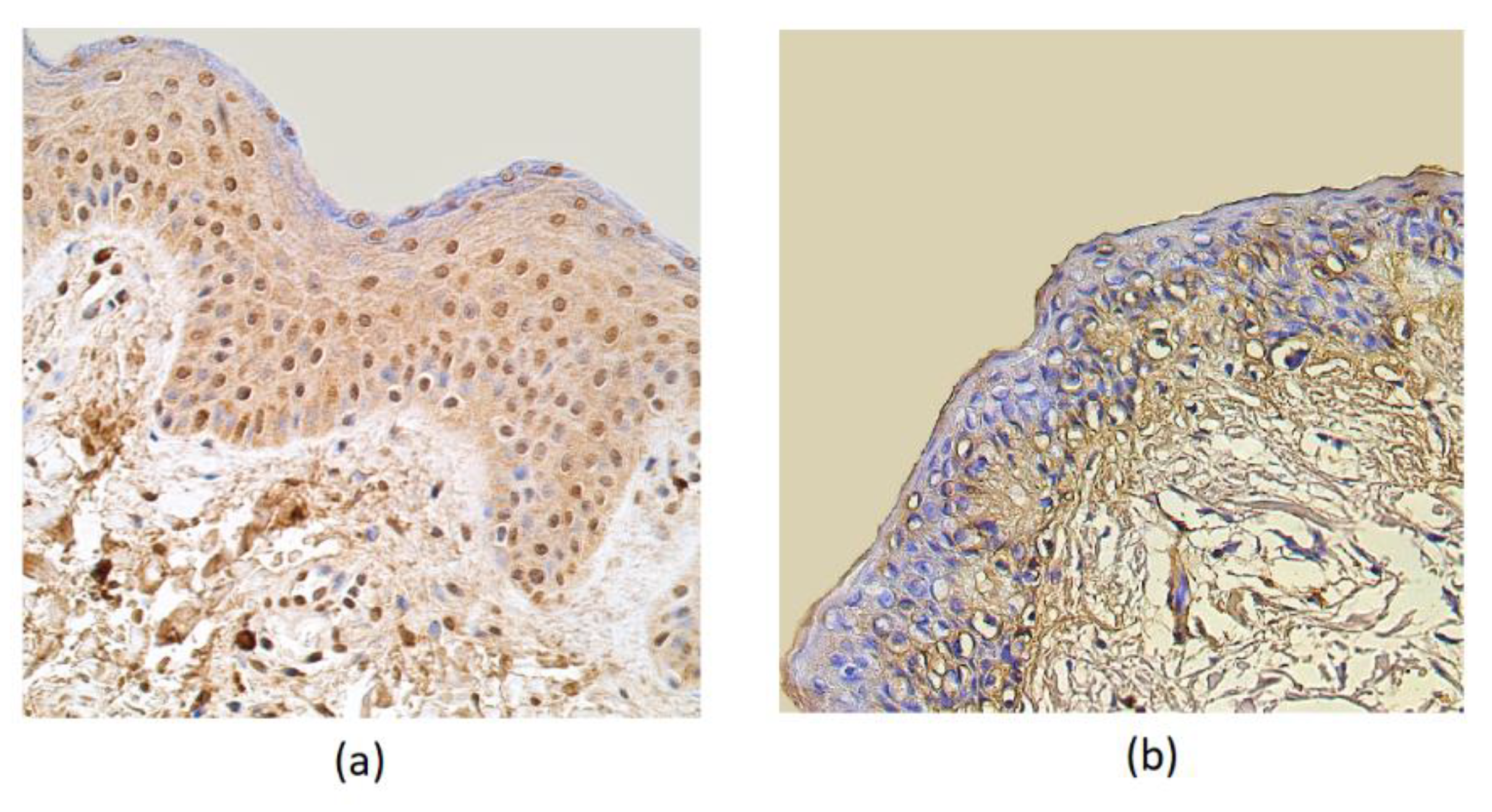
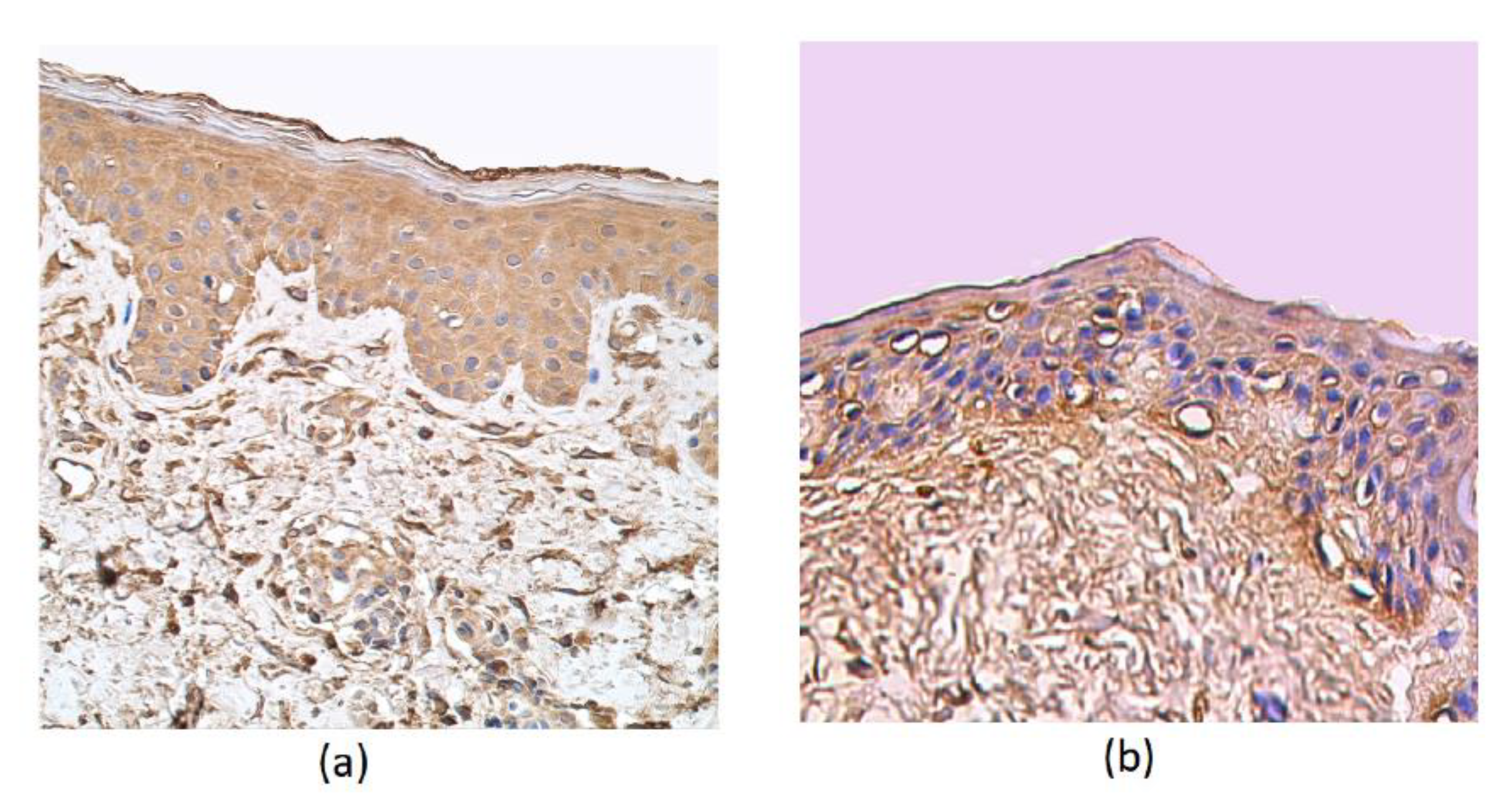
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Holmes, A.D.; Spoendlin, J.; Chien, A.L.; Baldwin, H.; Chang, A.L.S. Evidence-based update on rosacea comorbidities and their common physiologic pathways. J. Am. Acad. Dermatol. 2018, 78, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Abram, K.; Silm, H.; Oona, M. Prevalence of Rosacea in an Estonian Working Population Using a Standard Classification. Acta. Derm. Venereol. 2010, 90, 269–273. [Google Scholar] [CrossRef]
- Wilkin, J.; Dahl, M.; Detmar, M.; Drake, L.; Feinstein, A.; Odom, R.; Powell, F. Standard classification of rosacea: Report of the National Rosacea Society Expert Committee on the Classification and Staging of Rosacea. J. Am. Acad. Dermatol. 2002, 46, 584–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulk, M.; Seeliger, S.; Aubert, J.; Schwab, V.D.; Cevikbas, F.; Rivier, M.; Nowak, P.; Voegel, J.J.; Buddenkotte, J.; Steinhoff, M. Distribution and Expression of Non-Neuronal Transient Receptor Potential (TRPV) Ion Channels in Rosacea. J. Investig. Dermatol. 2012, 132, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buddenkotte, J.; Steinhoff, M. Recent advances in understanding and managing rosacea. F1000Research 2018, 7, 1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupper, T.S. Immune and inflammatory processes in cutaneous tissues: Mechanisms and speculations. J. Clin. Investig. 1990, 86, 1783–1789. [Google Scholar] [CrossRef]
- Palmer, G.; Talabot-Ayer, D.; Kaya, G.; Gabay, C. Type I IL-1 Receptor Mediates IL-1 and Intracellular IL-1 Receptor Antagonist Effects in Skin Inflammation. J. Investig. Dermatol. 2007, 127, 1938–1946. [Google Scholar] [CrossRef] [Green Version]
- Malik, A.; Kanneganti, T.D. Function and regulation of IL-1α in inflammatory diseases and cancer. Immunol. Rev. 2018, 281, 124–137. [Google Scholar] [CrossRef]
- Newby, C.S.; Barr, R.M.; Greaves, M.W.; Mallet, A.I. Cytokine release and cytotoxicity in human keratinocytes and fibroblasts induced by phenol and sodium dodecyl sulfate. J. Investig. Dermatol. 2000, 115, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Szabowski, A.; Maas-Szabowski, N.; Andrecht, S.; Kolbus, A.; Schorpp-Kistner, M.; Fusenig, N.E.; Angel, P. c-Jun and JunB antagonistically control cytokine-regulated mesenchymal-epidermal interaction in skin. Cell 2000, 103, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Harder, J.; Meyer-Hoffert, U.; Teran, L.M.; Schwichtenberg, L.; Bartels, J.; Maune, S.; Schröder, J.M. Mucoid Pseudomonas aeruginosa, TNF- α, and IL-1 β, but Not IL-6, Induce Human β -Defensin-2 in Respiratory Epithelia. ATS J. 1999, 22, 714–721. [Google Scholar] [CrossRef]
- Kubo, M.; Motomura, Y. Transcriptional regulation of the anti-inflammatory cytokine IL-10 in acquired immune cells. Front. Immunol. 2012, 3, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Ohshima, T.; Kondo, T. Regulatory role of endogenous interleukin-10 in cutaneous inflammatory response of murine wound healing. Biochem. Biophys. Res. Commun. 1999, 265, 194–199. [Google Scholar] [CrossRef]
- Abdoli, A.; Maspi, N.; Ghaffarifar, F. Wound healing in cutaneous leishmaniasis: A double edged sword of IL-10 and TGF-β. Comp. Immunol. Microbiol. Infect. Dis. 2017, 51, 15–26. [Google Scholar] [CrossRef]
- Taefehshokr, N.; Isazadeh, A.; Oveisi, A.; Key, Y.A.; Taefehshokr, S. Reciprocal role of hBD2 and hBD3 on the adaptive immune response by measuring T lymphocyte proliferation in terms of CD4 and CCR6 expression. Horm. Mol. Biol. Clin. Investig. 2018, 35. [Google Scholar] [CrossRef]
- Johansen, C.; Bertelsen, T.; Ljungberg, C.; Mose, M.; Iversen, L. Characterization of TNF-α– and IL-17A–Mediated Synergistic Induction of DEFB4 Gene Expression in Human Keratinocytes through IκBζ. J. Investig. Dermatol. 2016, 136, 1608–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehrer, R.I.; Ganz, T. Defensins of vertebrate animals. Curr. Opin. Immunol. 2002, 14, 96–102. [Google Scholar] [CrossRef]
- Schröder, J.M.; Harder, J. Human beta-defensin-2. Int. J. Biochem. Cell. Biol. 1999, 31, 645–651. [Google Scholar] [CrossRef]
- Baroni, A.; Donnarumma, G.; Paoletti, I.; Longanesi-Cattani, I.; Bifulco, K.; Tufano, M.A.; Carriero, M.V. Antimicrobial human beta-defensin-2 stimulates migration, proliferation and tube formation of human umbilical vein endothelial cells. Peptides 2009, 30, 267–272. [Google Scholar] [CrossRef]
- Lehrer, R.I.; Lu, W. α-Defensins in human innate immunity. Immunol. Rev. 2012, 245, 4–112. [Google Scholar] [CrossRef]
- Schneider, J.J.; Unholzer, A.; Schaller, M.; Schäfer-Korting, M.; Korting, H.C. Human defensins. J. Mol. Med. 2005, 83, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Smiley, A.K.; Gardner, J.; Klingenberg, J.M.; Neely, A.N.; Supp, D.M. Expression of human beta defensin 4 in genetically modified keratinocytes enhances antimicrobial activity. J. Burn. Care. Res. 2007, 28, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Harder, J.; Meyer-Hoffert, U.; Wehkamp, K.; Schwichtenberg, L.; Schröder, J.M. Differential Gene Induction of Human β-Defensins (hBD-1, -2, -3, and -4) in Keratinocytes Is Inhibited by Retinoic Acid. J. Investig. Dermatol. 2004, 123, 522–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabisiak, A.; Murawska, N.; Fichna, J. LL-37: Cathelicidin-related antimicrobial peptide with pleiotropic activity. Pharmacol. Rep. 2016, 68, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Bandurska, K.; Berdowska, A.; Barczyńska-Felusiak, R.; Krupa, P. Unique features of human cathelicidin LL-37. Biofactors 2015, 41, 289–300. [Google Scholar] [CrossRef]
- da Silva, P.F.; Machado, M.C. The dual role of cathelicidins in systemic inflammation. Immunol. Lett. 2017, 182, 57–60. [Google Scholar] [CrossRef]
- Di Nardo, A.; Vitiello, A.; Gallo, R.L. Cutting edge: Mast Cell antimicrobial activity is mediated by expression of cathelicidin antimicrobial peptide. J. Immunol. 2003, 170, 2274–2278. [Google Scholar] [CrossRef]
- Peric, M.; Lehmann, B.; Vashina, G.; Dombrowski, Y.; Koglin, S.; Meurer, M.; Ruzicka, T.; Schauber, J. UV-B—Triggered induction of vitamin D3 metabolism differentially affects antimicrobial peptide expression in keratinocytes. J. Allergy Clin. Immunol. 2010, 125, 746–749. [Google Scholar] [CrossRef]
- Junga, A.; Pilmane, M.; Ābola, Z.; Volrāts, O. The Distribution of Vascular Endothelial Growth Factor (VEGF), Human Beta-Defensin-2 (HBD-2), and Hepatocyte Growth Factor (HGF) in Intra-Abdominal Adhesions in Children under One Year of Age. Sci. World J. 2018, 2018, 5953095. [Google Scholar] [CrossRef]
- Gerber, P.A.; Buhren, B.A.; Steinhoff, M.; Homey, B. Rosacea: The Cytokine and Chemokine Network. J. Investig. Dermatol. Symp. Proc. 2011, 15, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Li, J.; Li, Y.; Deng, Z.; Zhou, L.; Long, J.; Tang, Y.; Zuo, Z.; Zhang, Y.; Xie, H. Artemisinin, a potential option to inhibit inflammation and angiogenesis in rosacea. Biomed. Pharmacother. 2019, 117, 109181. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, J.; Li, D.; Li, P.; Zhou, X.; Li, Y.; He, Z.; Qin, L.; Liang, L.; Luo, X. Interleukin-10 inhibits interleukin-1β production and inflammasome activation of microglia in epileptic seizures. J. Neuroinflamm. 2019, 16, 66. [Google Scholar] [CrossRef] [PubMed]
- de Waal Malefyt, R.; Abrams, J.; Bennett, B.; Figdor, C.G.; de Vries, J.E. Interleukin 10(IL-10) inhibits cytokine synthesis by human monocytes: An autoregulatory role of IL-10 produced by monocytes. J. Exp. Med. 1991, 174, 1209–1220. [Google Scholar] [CrossRef] [Green Version]
- Commins, S.; Steinke, J.W.; Borish, L. The extended IL-10 superfamily: IL-10, IL-19, IL-20, IL-22, IL-24, IL-26, IL-28, and IL-29. J. Allergy Clin. Immunol. 2008, 121, 1108–1111. [Google Scholar] [CrossRef] [PubMed]
- Weiss, E.; Mamelak, A.J.; La Morgia, S.; Wang, B.; Feliciani, C.; Tulli, A.; Sauder, D.N. The role of interleukin 10 in the pathogenesis and potential treatment of skin diseases. J. Am. Acad. Dermatol. 2004, 50, 657–675. [Google Scholar] [CrossRef]
- Margalit, A.; Kowalczyk, M.J.; Żaba, R.; Kavanagh, K. The role of altered cutaneous immune responses in the induction and persistence of rosacea. J. Dermatol. Sci. 2016, 82, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Marcinkiewicz, M.; Majewski, S. The role of antimicrobial peptides in chronic inflammatory skin diseases. Postepy Dermatol. Alergol. 2016, 33, 6–12. [Google Scholar] [CrossRef]
- Duits, L.A.; Ravensbergen, B.; Rademaker, M.; Hiemstra, P.S.; Nibbering, P.H. Expression of β-defensin 1 and 2 mRNA by human monocytes, macrophages and dendritic cells. Immunology 2002, 106, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Witthöft, T.; Pilz, C.S.; Fellermann, K.; Nitschke, M.; Stange, E.F.; Ludwig, D. Enhanced human beta-defensin-2 (HBD-2) expression by corticosteroids is independent of NF-kappaB in colonic epithelial cells (CaCo2). Dig. Dis. Sci. 2005, 50, 1252–1259. [Google Scholar] [CrossRef]
- Wang, B.; McHugh, B.J.; Qureshi, A.; Campopiano, D.J.; Clarke, D.J.; Fitzgerald, J.R.; Dorin, J.R.; Weller, R.; Davidson, D.J. IL-1β–Induced Protection of Keratinocytes against Staphylococcus aureus-Secreted Proteases Is Mediated by Human β-Defensin 2. J. Investig. Dermatol. 2017, 137, 95–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Chertov, O.; Bykovskaia, S.N.; Chen, Q.; Buffo, M.J.; Shogan, J.; Anderson, M.; Schröder, J.M.; Wang, J.M.; Howard, O.M.Z.; et al. β-Defensins: Linking Innate and Adaptive Immunity Through Dendritic and T Cell CCR6. Science 1999, 286, 525–528. [Google Scholar] [CrossRef]
- Yamasaki, K.; Schauber, J.; Coda, A.; Lin, H.; Dorschner, R.A.; Schechter, N.M.; Bonnart, C.; Descargues, P.; Hovnanian, A.; Gallo, R.L. Kallikrein-mediated proteolysis regulates the antimicrobial effects of cathelicidins in skin. FASEB J. 2006, 20, 2068–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaiou, M.; Gallo, R.L. Cathelicidins, essential gene-encoded mammalian antibiotics. J. Mol. Med. 2002, 80, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Schauber, J.; Gallo, R.L. The vitamin D pathway: A new target for control of the skin’s immune response. Exp. Dermatol. 2008, 17, 633–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, K.; Di Nardo, A.; Bardan, A.; Murakami, M.; Ohtake, T.; Coda, A.; Dorschner, R.A.; Bonnart, C.; Descargues, P.; Hovnanian, A.; et al. Increased serine protease activity and cathelicidin promotes skin inflammation in rosacea. Nat. Med. 2000, 13, 975–980. [Google Scholar] [CrossRef]
- Elssner, A.; Duncan, M.; Gavrilin, M.; Wewers, M.D. A Novel P2 × 7 Receptor Activator, the Human Cathelicidin-Derived Peptide LL37, Induces IL-1β Processing and Release. J. Immunol. 2004, 172, 4987–4994. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nr | Age | Diagnosis | IL-1α | IL-10 | HBD-2 | HBD-4 | LL-37 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E | CT | E | CT | E | CT | E | CT | E | CT | |||
| 1 | 32 | Papulopustular rosacea | +++ | +++ | +++/++++ | +++ | +++ | ++/+++ | ++/+++ | ++ | +++ | ++ |
| 2 | 35 | Papulopustular rosacea | ++/+++ | ++/+++ | +++/++++ | ++/+++ | ++ | ++ | +++ | ++ | ++/+++ | ++/+++ |
| 3 | 36 | Papulopustular rosacea | +++ | ++/+++ | +++ | +++ | ++ | +++ | ++ | +/++ | +++ | +++ |
| 4 | 36 | Papulopustular rosacea | ++ | ++ | +++ | ++/+++ | +/++ | +++ | +++ | ++/+++ | +/++ | ++ |
| 5 | 44 | Papulopustular rosacea | ++/+++ | ++ | ++/+++ | +++ | ++ | ++/+++ | +++ | ++/+++ | +++ | ++ |
| 6 | 45 | Papulopustular rosacea | ++/+++ | +++ | +++ | +++ | ++/+++ | ++/+++ | ++/+++ | ++ | ++ | ++/+++ |
| 7 | 47 | Papulopustular rosacea | ++ | 0/+ | ++/+++ | 0/+ | ++ | + | ++ | + | ++ | +/++ |
| 8 | 50 | Papulopustular rosacea | +/++ | ++/+++ | +++ | ++/+++ | +++ | ++/+++ | +++ | ++ | ++/+++ | ++/+++ |
| Subjects common | ++ | ++ | +++ | ++/+++ | ++ | ++ | ++ | ++ | ++/+++ | ++ | ||
| Control common | + | 0/+ | + | + | +/++* | 0/+ | + | 0 | + | + | ||
| Detected Factors | Mann-Whitney U | Z-score | p-Value |
|---|---|---|---|
| HBD-2 in epithelium | 6 | −1.820 | 0.049 |
| Strength of Correlation | Marker 1 | Marker 2 | Rho (ρ) | p-Value |
|---|---|---|---|---|
| Strong positive correlation | HBD-4 in connective tissue | HBD-4 in epithelium | 0.828 | 0.011 |
| IL-1α in epithelium | IL-10 in connective tissue | 0.771 | 0.025 | |
| IL-1α in connective tissue | IL-10 in epithelium | 0.721 | 0.044 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lohova, E.; Pilmane, M.; Rone-Kupfere, M.; Kisis, J. Characterization of Pro- and Anti-Inflammatory Tissue Factors in Rosacea: A Pilot study. Cosmetics 2020, 7, 82. https://doi.org/10.3390/cosmetics7040082
Lohova E, Pilmane M, Rone-Kupfere M, Kisis J. Characterization of Pro- and Anti-Inflammatory Tissue Factors in Rosacea: A Pilot study. Cosmetics. 2020; 7(4):82. https://doi.org/10.3390/cosmetics7040082
Chicago/Turabian StyleLohova, Elizabeta, Mara Pilmane, Mara Rone-Kupfere, and Janis Kisis. 2020. "Characterization of Pro- and Anti-Inflammatory Tissue Factors in Rosacea: A Pilot study" Cosmetics 7, no. 4: 82. https://doi.org/10.3390/cosmetics7040082
APA StyleLohova, E., Pilmane, M., Rone-Kupfere, M., & Kisis, J. (2020). Characterization of Pro- and Anti-Inflammatory Tissue Factors in Rosacea: A Pilot study. Cosmetics, 7(4), 82. https://doi.org/10.3390/cosmetics7040082