
Ex Vivo Assessment of Heat and Humidity Effects on Human Skin and Potential Protection by Kombucha Tea Extract

 , , ,
, , , 

Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Human Skin Samples
2.2. Kombucha Tea Extract
2.3. Environmental Stress Exposure
2.4. Immunofluorescence Assays
2.5. Image Acquisition and Analysis
2.6. Atomic Force Microscopy (AFM) Measurements
2.7. Proteome Analysis
2.8. Statistical Analysis
3. Results
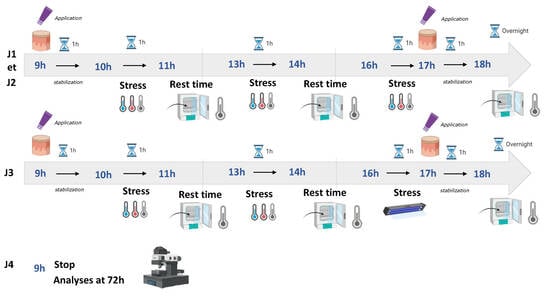
3.1. Experimental Setup for Exposing Human Skin Explants to Challenging Environmental Conditions and Evaluating the Effects of Kombucha Tea Extract
3.2. Epidermal Morphometry Is Altered Under Tropical Conditions and Preserved by Kombucha Tea Extract
3.3. AFM Analysis Reveals Environmental Stress–Induced Loss of Mechanical Contrast at the DEJ, Mitigated by Kombucha Tea Extract
3.4. Environmental Stress Alters MSCP and Collagen VII Expression, Normalized by Kombucha Tea Extract
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blanpain, C.; Fuchs, E. Epidermal homeostasis: A balancing act of stem cells in the skin. Nat. Rev. Mol. Cell Biol. 2009, 10, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Watt, F.M. Epidermal stem cells: Markers, patterning and the control of stem cell fate. Philos. Trans. R. Soc. B Biol. Sci. 1998, 353, 831–837. [Google Scholar] [CrossRef]
- Lawlor, K.T.; Kaur, P. Dermal Contributions to Human Interfollicular Epidermal Architecture and Self-Renewal. Int. J. Mol. Sci. 2015, 16, 28098–28107. [Google Scholar] [CrossRef] [PubMed]
- Roig-Rosello, E.; Rousselle, P. The Human Epidermal Basement Membrane: A Shaped and Cell Instructive Platform That Aging Slowly Alters. Biomolecules 2020, 10, 1607. [Google Scholar] [CrossRef]
- Jensen, K.B.; Watt, F.M. Single-cell expression profiling of human epidermal stem and transit-amplifying cells: Lrig1 is a regulator of stem cell quiescence. Proc. Natl. Acad. Sci. USA 2006, 103, 11958–11963. [Google Scholar] [CrossRef]
- Jones, P.H.; Harper, S.; Watt, F.M. Stem cell patterning and fate in human epidermis. Cell 1995, 80, 83–93. [Google Scholar] [CrossRef]
- Legg, J.; Jensen, U.B.; Broad, S.; Leigh, I.; Watt, F.M. Role of melanoma chondroitin sulphate proteoglycan in patterning stem cells in human interfollicular epidermis. Development 2003, 130, 6049–6063. [Google Scholar] [CrossRef]
- Inoue, K.; Aoi, N.; Sato, T.; Yamauchi, Y.; Suga, H.; Eto, H.; Kato, H.; Araki, J.; Yoshimura, K. Differential expression of stem-cell-associated markers in human hair follicle epithelial cells. Lab. Investig. 2009, 89, 844–856. [Google Scholar] [CrossRef]
- Webb, A.; Li, A.; Kaur, P. Location and phenotype of human adult keratinocyte stem cells of the skin. Differentiation 2004, 72, 387–395. [Google Scholar] [CrossRef]
- Wang, S.; Drummond, M.L.; Guerrero-Juarez, C.F.; Tarapore, E.; MacLean, A.L.; Stabell, A.R.; Wu, S.C.; Gutierrez, G.; That, B.T.; Benavente, C.A.; et al. Single cell transcriptomics of human epidermis identifies basal stem cell transition states. Nat. Commun. 2020, 11, 4239. [Google Scholar] [CrossRef] [PubMed]
- Ghali, L.; Wong, S.-T.; Tidman, N.; Quinn, A.; Philpott, M.P.; Leigh, I.M. Epidermal and hair follicle progenitor cells express melanoma-associated chondroitin sulfate proteoglycan core protein. J. Investig. Dermatol. 2004, 122, 433–442. [Google Scholar] [CrossRef]
- Jones, P.H.; Watt, F.M. Separation of human epidermal stem cells from transit amplifying cells on the basis of differences in integrin function and expression. Cell 1993, 73, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Baqar, A. Dermal papillae; the histomorphological changes in the height of dermal papillae of human skin, in different age groups. Prof. Med. J. 2019, 26, 208–212. [Google Scholar] [CrossRef]
- Giangreco, A.; Goldie, S.J.; Failla, V.; Saintigny, G.; Watt, F.M. Human skin aging is associated with reduced expression of the stem cell markers beta1 integrin and MCSP. J. Investig. Dermatol. 2010, 130, 604–608. [Google Scholar] [CrossRef]
- Zobiri, O.; Deshayes, N.; Rathman-Josserand, M. Evolution of the clonogenic potential of human epidermal stem/progenitor cells with age. Stem Cells Cloning 2012, 5, 1–4. [Google Scholar] [CrossRef][Green Version]
- Jang, S.I.; Han, J.; Lee, M.; Seo, J.; Kim, B.J.; Kim, E. A study of skin characteristics according to humidity during sleep. Ski. Res. Technol. 2019, 25, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Tsukahara, K.; Hotta, M.; Fujimura, T.; Haketa, K.; Kitahara, T. Effect of room humidity on the formation of fine wrinkles in the facial skin of Japanese. Ski. Res. Technol. 2007, 13, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Bouwstra, J.A.; de Graaff, A.; Gooris, G.S.; Nijsse, J.; Wiechers, J.W.; van Aelst, A.C. Water distribution and related morphology in human stratum corneum at different hydration levels. J. Investig. Dermatol. 2003, 120, 750–758. [Google Scholar] [CrossRef]
- Cau, L.; Pendaries, V.; Lhuillier, E.; Thompson, P.R.; Serre, G.; Takahara, H.; Méchin, M.-C.; Simon, M. Lowering relative humidity level increases epidermal protein deimination and drives human filaggrin breakdown. J. Dermatol. Sci. 2017, 86, 106–113. [Google Scholar] [CrossRef]
- Denda, M.; Sato, J.; Masuda, Y.; Tsuchiya, T.; Koyama, J.; Kuramoto, M.; Elias, P.M.; Feingold, K.R. Exposure to a dry environment enhances epidermal permeability barrier function. J. Investig. Dermatol. 1998, 111, 858–863. [Google Scholar] [CrossRef]
- Denda, M.; Sato, J.; Tsuchiya, T.; Elias, P.M.; Feingold, K.R. Low humidity stimulates epidermal DNA synthesis and amplifies the hyperproliferative response to barrier disruption: Implication for seasonal exacerbations of inflammatory dermatoses. J. Investig. Dermatol. 1998, 111, 873–878. [Google Scholar] [CrossRef]
- Shinohara, K.; Hara-Chikuma, M. Low humidity altered the gene expression profile of keratinocytes in a three-dimensional skin model. Mol. Biol. Rep. 2022, 49, 7465–7474. [Google Scholar] [CrossRef]
- Seo, J.Y.; Chung, J.H. Thermal aging: A new concept of skin aging. J. Dermatol. Sci. Suppl. 2006, 2, S13–S22. [Google Scholar] [CrossRef]
- Chen, Z.; Seo, J.Y.; Kim, Y.K.; Lee, S.R.; Kim, K.H.; Cho, K.H.; Eun, H.C.; Chung, J.H. Heat modulation of tropoelastin, fibrillin-1, and matrix metalloproteinase-12 in human skin in vivo. J. Investig. Dermatol. 2005, 124, 70–78. [Google Scholar] [CrossRef]
- Park, C.-H.; Lee, M.J.; Ahn, J.; Kim, S.; Kim, H.H.; Kim, K.H.; Eun, H.C.; Chung, J.H. Heat shock-induced matrix metalloproteinase (MMP)-1 and MMP-3 are mediated through ERK and JNK activation and via an autocrine interleukin-6 loop. J. Investig. Dermatol. 2004, 123, 1012–1019. [Google Scholar] [CrossRef]
- Shin, M.H.; Seo, J.-E.; Kim, Y.K.; Kim, K.H.; Chung, J.H. Chronic heat treatment causes skin wrinkle formation and oxidative damage in hairless mice. Mech. Ageing Dev. 2012, 133, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Park, G.; Oh, M.S. Acceleration of heat shock-induced collagen breakdown in human dermal fibroblasts with knockdown of NF-E2-related factor 2. BMB Rep. 2015, 48, 467–472. [Google Scholar] [CrossRef]
- Shin, M.H.; Moon, Y.J.; Seo, J.-E.; Lee, Y.; Kim, K.H.; Chung, J.H. Reactive oxygen species produced by NADPH oxidase, xanthine oxidase, and mitochondrial electron transport system mediate heat shock-induced MMP-1 and MMP-9 expression. Free Radic. Biol. Med. 2008, 44, 635–645. [Google Scholar] [CrossRef]
- Cho, S.; Shin, M.H.; Kim, Y.K.; Seo, J.-E.; Lee, Y.M.; Park, C.-H.; Chung, J.H. Effects of infrared radiation and heat on human skin aging in vivo. J. Investig. Dermatol. Symp. Proc. 2009, 14, 15–19. [Google Scholar] [CrossRef]
- Shimoni, C.; Goldstein, M.; Ribarski-Chorev, I.; Schauten, I.; Nir, D.; Strauss, C.; Schlesinger, S. Heat Shock Alters Mesenchymal Stem Cell Identity and Induces Premature Senescence. Front. Cell Dev. Biol. 2020, 8, 565970. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, B.Y. Changes in major components of tea fungus metabolites during prolonged fermentation. J. Appl. Microbiol. 2000, 89, 834–839. [Google Scholar] [CrossRef]
- Battikh, H.; Chaieb, K.; Bakhrouf, A.; Ammar, E. Antibacterial and Antifungal Activities of Black and Green Kombucha Teas. J. Food Biochem. 2013, 37, 231–236. [Google Scholar] [CrossRef]
- Conney, A.H.; Lu, Y.-P.; Lou, Y.-R.; Huang, M.-T. Inhibitory effects of tea and caffeine on UV-induced carcinogenesis: Relationship to enhanced apoptosis and decreased tissue fat. Eur. J. Cancer Prev. 2002, 11, S28–S36. [Google Scholar] [PubMed]
- Ioannides, C.; Yoxall, V. Antimutagenic activity of tea: Role of polyphenols. Curr. Opin. Clin. Nutr. Metab. Care 2003, 6, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Park, A.M.; Dong, Z. Signal transduction pathways: Targets for green and black tea polyphenols. J. Biochem. Mol. Biol. 2003, 36, 66–77. [Google Scholar]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.-P.; Renard, T.; Rollan, S.; Taillandier, P. Impact of fermentation conditions on the production of bioactive compounds with anticancer, anti-inflammatory and antioxidant properties in kombucha tea extracts. Process Biochem. 2019, 83, 44–54. [Google Scholar] [CrossRef]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.-P.; Taillandier, P. Understanding Kombucha Tea Fermentation: A Review. J. Food Sci. 2018, 83, 580–588. [Google Scholar] [CrossRef]
- Wang, Y.; Ji, B.; Wu, W.; Wang, R.; Yang, Z.; Zhang, D.; Tian, W. Hepatoprotective effects of kombucha tea: Identification of functional strains and quantification of functional components. J. Sci. Food Agric. 2014, 94, 265–272. [Google Scholar] [CrossRef]
- Abe, Y.; Tanaka, N. Roles of the Hedgehog Signaling Pathway in Epidermal and Hair Follicle Development, Homeostasis, and Cancer. J. Dev. Biol. 2017, 5, 12. [Google Scholar] [CrossRef]
- Chuang, L.S.H.; Matsuo, J.; Douchi, D.; Bte Mawan, N.A.; Ito, Y. RUNX3 in Stem Cell and Cancer Biology. Cells 2023, 12, 408. [Google Scholar] [CrossRef]
- Rozen, E.J.; Ozeroff, C.D.; Allen, M.A. RUN(X) out of blood: Emerging RUNX1 functions beyond hematopoiesis and links to Down syndrome. Hum. Genom. 2023, 17, 83. [Google Scholar] [CrossRef] [PubMed]
- Borrell, V.; Cárdenas, A.; Ciceri, G.; Galcerán, J.; Flames, N.; Pla, R.; Nóbrega-Pereira, S.; García-Frigola, C.; Peregrín, S.; Zhao, Z.; et al. Slit/Robo signaling modulates the proliferation of central nervous system progenitors. Neuron 2012, 76, 338–352. [Google Scholar] [CrossRef]
- Harburg, G.; Compton, J.; Liu, W.; Iwai, N.; Zada, S.; Marlow, R.; Strickland, P.; Zeng, Y.A.; Hinck, L. SLIT/ROBO2 signaling promotes mammary stem cell senescence by inhibiting Wnt signaling. Stem Cell Rep. 2014, 3, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-S.; Cho, R.; Kang, E.-Y.; Oh, Y.-M. Effect of Slit/Robo signaling on regeneration in lung emphysema. Exp. Mol. Med. 2021, 53, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Miny, S.; Runel, G.; Chlasta, J.; Lapart, J.-A.; Bonod, C. Putative interfollicular stem cells of skin epidermis possess a specific mechanical signature that evolves during aging. bioRxiv 2025. [Google Scholar] [CrossRef]
- Ji, Y.; Kumar, R.; Gokhale, A.; Chao, H.-P.; Rycaj, K.; Chen, X.; Li, Q.; Tang, D.G. LRIG1, a regulator of stem cell quiescence and a pleiotropic feedback tumor suppressor. Semin. Cancer Biol. 2022, 82, 120–133. [Google Scholar] [CrossRef]
- Breitkreutz, D.; Koxholt, I.; Thiemann, K.; Nischt, R. Skin basement membrane: The foundation of epidermal integrity—BM functions and diverse roles of bridging molecules nidogen and perlecan. Biomed. Res. Int. 2013, 2013, 179784. [Google Scholar] [CrossRef]
- Izutsu-Matsumoto, Y.; Okano, Y.; Masaki, H.; Tokudome, Y.; Iwabuchi, T. Effect of low humidity on the barrier functions of keratinocytes in a reconstructed human epidermal model. Int. J. Cosmet. Sci. 2025, 47, 523–534. [Google Scholar] [CrossRef]
- Rawlings, A.V.; Matts, P.J. Stratum corneum moisturization at the molecular level: An update in relation to the dry skin cycle. J. Investig. Dermatol. 2005, 124, 1099–1110. [Google Scholar] [CrossRef]
- Warner, R.R.; Stone, K.J.; Boissy, Y.L. Hydration disrupts human stratum corneum ultrastructure. J. Investig. Dermatol. 2003, 120, 275–284. [Google Scholar] [CrossRef]
- Kim, S.; Park, J.W.; Yeon, Y.; Han, J.Y.; Kim, E. Influence of exposure to summer environments on skin properties. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 2192–2196. [Google Scholar] [CrossRef]
- Song, E.J.; Lee, J.A.; Park, J.J.; Kim, H.J.; Kim, N.S.; Byun, K.S.; Choi, G.S.; Moon, T.K. A study on seasonal variation of skin parameters in Korean males. Int. J. Cosmet. Sci. 2015, 37, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Dams, S.D.; de Liefde-van Beest, M.; Nuijs, A.M.; Oomens, C.W.J.; Baaijens, F.P.T. Heat shocks enhance procollagen type I and III expression in fibroblasts in ex vivo human skin. Skin Res. Technol. 2011, 17, 167–180. [Google Scholar] [CrossRef]
- Guarnieri, G.; Olivieri, B.; Senna, G.; Vianello, A. Relative Humidity and Its Impact on the Immune System and Infections. Int. J. Mol. Sci. 2023, 24, 9456. [Google Scholar] [CrossRef]
- Sonnenberg, A.; Calafat, J.; Janssen, H.; Daams, H.; van der Raaij-Helmer, L.M.; Falcioni, R.; Kennel, S.J.; Aplin, J.D.; Baker, J.; Loizidou, M. Integrin alpha 6/beta 4 complex is located in hemidesmosomes, suggesting a major role in epidermal cell-basement membrane adhesion. J. Cell Biol. 1991, 113, 907–917. [Google Scholar] [CrossRef]
- Yu, X.; Miyamoto, S.; Mekada, E. Integrin alpha 2 beta 1-dependent EGF receptor activation at cell-cell contact sites. J. Cell Sci. 2000, 113, 2139–2147. [Google Scholar] [CrossRef]
- Broussard, J.A.; Yang, R.; Huang, C.; Nathamgari, S.S.P.; Beese, A.M.; Godsel, L.M.; Hegazy, M.H.; Lee, S.; Zhou, F.; Sniadecki, N.J.; et al. The desmoplakin-intermediate filament linkage regulates cell mechanics. Mol. Biol. Cell 2017, 28, 3156–3164. [Google Scholar] [CrossRef] [PubMed]
- Efremov, Y.M.; Dokrunova, A.A.; Efremenko, A.V.; Kirpichnikov, M.P.; Shaitan, K.V.; Sokolova, O.S. Distinct impact of targeted actin cytoskeleton reorganization on mechanical properties of normal and malignant cells. Biochim. Biophys. Acta 2015, 1853, 3117–3125. [Google Scholar] [CrossRef]
- Kasza, K.E.; Rowat, A.C.; Liu, J.; Angelini, T.E.; Brangwynne, C.P.; Koenderink, G.H.; Weitz, D.A. The cell as a material. Curr. Opin. Cell Biol. 2007, 19, 101–107. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, C.; Jiang, N.; Zheng, L.; Zeng, J.; Qiu, C.; Yang, H.; Xie, S. Quantitative analysis of the cell-surface roughness and viscoelasticity for breast cancer cells discrimination using atomic force microscopy. Scanning 2016, 38, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Mezencev, R.; Kim, B.; Wang, L.; McDonald, J.; Sulchek, T. Cell stiffness is a biomarker of the metastatic potential of ovarian cancer cells. PLoS ONE 2012, 7, e46609. [Google Scholar] [CrossRef]
- Homberg, M.; Ramms, L.; Schwarz, N.; Dreissen, G.; Leube, R.E.; Merkel, R.; Hoffmann, B.; Magin, T.M. Distinct Impact of Two Keratin Mutations Causing Epidermolysis Bullosa Simplex on Keratinocyte Adhesion and Stiffness. J. Investig. Dermatol. 2015, 135, 2437–2445. [Google Scholar] [CrossRef] [PubMed]
- Ramms, L.; Fabris, G.; Windoffer, R.; Schwarz, N.; Springer, R.; Zhou, C.; Lazar, J.; Stiefel, S.; Hersch, N.; Schnakenberg, U.; et al. Keratins as the main component for the mechanical integrity of keratinocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 18513–18518. [Google Scholar] [CrossRef]
- Seltmann, K.; Fritsch, A.W.; Käs, J.A.; Magin, T.M. Keratins significantly contribute to cell stiffness and impact invasive behavior. Proc. Natl. Acad. Sci. USA 2013, 110, 18507–18512. [Google Scholar] [CrossRef]
- Byfield, F.J.; Reen, R.K.; Shentu, T.-P.; Levitan, I.; Gooch, K.J. Endothelial actin and cell stiffness is modulated by substrate stiffness in 2D and 3D. J. Biomech. 2009, 42, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Discher, D.E.; Mooney, D.J.; Zandstra, P.W. Growth factors, matrices, and forces combine and control stem cells. Science 2009, 324, 1673–1677. [Google Scholar] [CrossRef]
- Emig, R.; Knodt, W.; Krussig, M.J.; Zgierski-Johnston, C.M.; Gorka, O.; Groß, O.; Kohl, P.; Ravens, U.; Peyronnet, R. Piezo1 Channels Contribute to the Regulation of Human Atrial Fibroblast Mechanical Properties and Matrix Stiffness Sensing. Cells 2021, 10, 663. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef]
- Solon, J.; Levental, I.; Sengupta, K.; Georges, P.C.; Janmey, P.A. Fibroblast adaptation and stiffness matching to soft elastic substrates. Biophys. J. 2007, 93, 4453–4461. [Google Scholar] [CrossRef]
- Zhang, L.; Zeng, H.; Jiang, L.; Fu, C.; Zhang, Y.; Hu, Y.; Zhang, X.; Zhu, L.; Zhang, F.; Huang, J.; et al. Heat promotes melanogenesis by increasing the paracrine effects in keratinocytes via the TRPV3/Ca2+/Hh signaling pathway. iScience 2023, 26, 106749. [Google Scholar] [CrossRef] [PubMed]
- Joo, J.-H.; Seok, J.H.; Hong, I.-k.; Kim, N.K.; Choi, E. Inhibitory Effects of Dunaliella salina Extracts on Thermally-Induced Skin Aging. J. Soc. Cosmet. Sci. Korea 2016, 42, 57–64. [Google Scholar] [CrossRef]
- Kim, H.-S.; Montana, V.; Jang, H.-J.; Parpura, V.; Kim, J.-a. Epigallocatechin gallate (EGCG) stimulates autophagy in vascular endothelial cells: A potential role for reducing lipid accumulation. J. Biol. Chem. 2013, 288, 22693–22705. [Google Scholar] [CrossRef] [PubMed]
- Son, D.; Kim, M.; Woo, H.; Park, D.; Jung, E. Anti-Thermal Skin Aging Activity of Aqueous Extracts Derived from Apple Mint (Mentha suaveolens Ehrh.) in Human Dermal Fibroblasts. Evid. Based Complement. Altern. Med. 2018, 2018, 4595982. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Chlasta, J.; Runel, G.; Boussard, M.; Pele-Joly, T.; Lazou, K.; Pays, K.; Nizard, C.; Amoedo, N.D.; Rossignol, R.; Bulteau, A.-L. Ex Vivo Assessment of Heat and Humidity Effects on Human Skin and Potential Protection by Kombucha Tea Extract. Cosmetics 2026, 13, 114. https://doi.org/10.3390/cosmetics13030114
Chlasta J, Runel G, Boussard M, Pele-Joly T, Lazou K, Pays K, Nizard C, Amoedo ND, Rossignol R, Bulteau A-L. Ex Vivo Assessment of Heat and Humidity Effects on Human Skin and Potential Protection by Kombucha Tea Extract. Cosmetics. 2026; 13(3):114. https://doi.org/10.3390/cosmetics13030114
Chicago/Turabian StyleChlasta, Julien, Gaël Runel, Manon Boussard, Tiphaine Pele-Joly, Kristell Lazou, Karl Pays, Carine Nizard, Nivea Dias Amoedo, Rodrigue Rossignol, and Anne-Laure Bulteau. 2026. "Ex Vivo Assessment of Heat and Humidity Effects on Human Skin and Potential Protection by Kombucha Tea Extract" Cosmetics 13, no. 3: 114. https://doi.org/10.3390/cosmetics13030114
APA StyleChlasta, J., Runel, G., Boussard, M., Pele-Joly, T., Lazou, K., Pays, K., Nizard, C., Amoedo, N. D., Rossignol, R., & Bulteau, A.-L. (2026). Ex Vivo Assessment of Heat and Humidity Effects on Human Skin and Potential Protection by Kombucha Tea Extract. Cosmetics, 13(3), 114. https://doi.org/10.3390/cosmetics13030114