Abstract
Natural ingredients have long been utilized to enhance human health. While Rosa rubiginosa L. has been relatively understudied, it is known for its antioxidant and anti-inflammatory properties. This study explores its diverse bioactivity on human primary keratinocytes and fibroblasts, highlighting its potential as a bio-functional agent in cosmeceuticals or nutraceuticals. The Rosa rubiginosa L. leaf extract demonstrated strong scavenging activity and high phenolic content protection while also preventing DNA breakage. At the molecular level, ATP determination revealed that the Rosa rubiginosa L. leaf confers cell viability even under oxidative stress. Gene expression analysis uncovered its protective effects on human primary keratinocytes and fibroblasts due to its strong antioxidant and anti-inflammatory response under oxidative stress induced by UVA irradiation. These findings suggest that Rosa rubiginosa L. leaf extract provides significant in vitro biological properties in cell-based models, offering new insights into its potential benefits and underscoring its bio-functional role.
1. Introduction
In recent years, several industries have witnessed a paradigm shift toward the development of bio-functional products, such as nutricosmetics or cosmeceuticals, providing beneficial effects on skin and body health. At the heart of this evolution are bio-functional ingredients, mainly naturally occurring derived compounds that provide targeted, biologically active effects on skin and overall appearance. These ingredients, such as peptides, antioxidants, polyphenols and vitamins, are critical for enhancing skin health, with a low toxicity profile [1,2,3]. One such promising bio-functional ingredient is Rosa rubiginosa L., commonly known as sweet briar rose or eglantine rose. Native to Europe and western Asia, Rosa rubiginosa L. has been traditionally valued for its medicinal and cosmetic properties, with historical use dating back centuries in wound healing and skin nourishment [4,5]. The plant is well known for its high concentrations of bioactive compounds, particularly in its seeds and leaves, and recent studies have shifted attention to its leaf extract due to its unique phytochemical profile [6].
Rosa rubiginosa leaf extract is rich in polyphenols, flavonoids, tannins and essential fatty acids, which are known for their antioxidant, anti-inflammatory, antimicrobial and collagen-promoting properties, making the leaf extract particularly attractive for nutricosmetic and cosmeceutical applications [3,4,6]. Polyphenols are especially potent in scavenging free radicals and protecting skin cells from oxidative damage, which is a key contributor to premature aging and pigmentation disorders [7]. In addition, the tannins present in the leaf extract provide astringent properties that can tighten pores and improve skin tone, while flavonoids contribute to skin barrier protection and overall dermal health [8].
Furthermore, in vitro studies have interestingly reported that other species of the Rosa genus, such as Rosa hybrida (white rose) petal extract, exhibited inhibitory effects on matrix metalloproteinase-1 (MMP-1), an enzyme responsible for collagen degradation, suggesting the potential for promoting collagen synthesis and skin health [9], while Rosa gallica extract exhibited skin anti-inflammatory activity through reduction of the MAPK signaling pathway [10]. In addition, another recent study suggested that Rosa rugosa extracts revealed significant antibacterial activity against various pathogens, indicating potential applications in treating skin conditions, such as acne [11]. Recently, the antioxidant activity of the rosehip of Rosa rubiginosa L. has been reported. However, there are no studies to highlight the potential antioxidant mechanism of action of Rosa rubiginosa L. leaf extract in human cells.
As the demand for plant-based, sustainable and science-backed bioactive ingredients continues to rise, Rosa rubiginosa L. leaf extract is poised to become a cornerstone ingredient in the next generation of bio-functional products, such as cosmeceuticals. Thus, in this work, we illuminate the biological properties of Rosa rubiginosa L. leaf extract in 2D skin-cell-based models under photo-oxidative stress, using an array of in vitro assays with the aim of providing an insight into specific skin-related biochemical mechanisms that may underscore its full potential as a bio-functional ingredient. For this purpose, we focused on cytotoxicity assessments, antioxidant assessments, as well as on providing an assembly of different gene expressions in 2D skin cell models, providing a critical insight into its long-term efficacy and safety.
2. Materials and Methods
2.1. Rosa rubiginosa L. Leaf Extract Preparation
Rosa rubiginosa was collected from Heraklion, Crete. The plant’s powdered material (10 g) was macerated in a 100 mL Erlenmeyer flask [12]. The extraction solvent consisted of 90% distilled water and 10% ethanol, a combination that is safe, cost-effective, eco-friendly and effective in preserving bioactive compounds. The mixture was left in the dark at room temperature for two weeks with frequent agitation. After filtration, the extract was centrifuged at 8000× g for 15 min, and the supernatant was collected. Solvent removal was carried out using a rotary evaporator at 50 °C for 2 h. The resulting solid residue was dissolved in an appropriate solvent and stored at 2 °C for further experiments [13]. Methanol was used for antioxidant capacity analysis, while water was employed to assess antimicrobial and cytotoxic effects.
Proper storage is essential for maintaining the stability and potency of plant extracts. Factors such as temperature, light exposure, air contact and solvent type affect their stability [14]. The extracts were kept at 2 °C to slow the degradation of heat-sensitive active compounds. They were also stored in darkness to prevent light-induced oxidative stress, which could reduce the effectiveness of phytochemicals. Additionally, airtight containers were used to minimize oxidation, thereby preserving bioactive components like polyphenols.
2.2. Antioxidant Assays
2.2.1. DPPH Method
The Rosa rubiginosa L. extract’s capacity to neutralize free radicals was evaluated using the 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical scavenging assay. DPPH loses its free radical characteristics and changes color from deep purple to pale yellow upon interacting with an antioxidant compound [15]. Using this method, various concentrations of the extract (8.44, 16.88, 33.75, 67.5, 135 μg/mL) (100 μL) were mixed with a 6 × 10−5 M methanolic solution of the DPPH radical (3400 μL) [16]. Absorbance of the samples was measured at 517 nm with a VIS spectrophotometer (Thermo Spectronic Helios Epsilon, Waltham, MA, USA) after being restored at room temperature for 45 min (Asample). A sample, which included 3400 μL DPPH solution and 100 μL methanol, was used as control to assess the maximum reduction in DPPH absorbance (Acontrol). The same procedure was repeated for five concentrations of the standard compound: gallic acid. The DPPH radical scavenging percentage was determined using the following equation:
2.2.2. Folin–Ciocalteu Method
Polyphenol compounds are essential in plants because their hydroxyl groups enable effective free radical scavenging. The total phenolic content of dried Rosa rubiginosa L. leaves was measured using the Folin–Ciocalteu method, as outlined by Huang et al. [17]. In summary, 200 μL of the different concentrations (8.44–135 μg/mL) of the tested extract was combined with 1 mL of Folin–Ciocalteu reagent and 800 μL of 7.5% w/v Na2CO3. The mixture was incubated for 1 h at room temperature in darkness, followed by absorbance measurement at 765 nm. Different concentrations (12.5, 25.0, 50.0, 100.0, 200.0 μg/mL) of the standard compound, gallic acid, were used to present the Folin–Ciocalteu results [18]. The equation describing the curve of gallic acid (12.5–200 μg/mL) was y = 0.0118x − 0.0819 and R2 = 0.9953, p < 0.05.
The findings were expressed as μg of gallic acid equivalents per mg of dry weight (μg GAE/mg DW).
2.2.3. Prevention of Supercoiled Plasmid DNA Strand Breakage
The inhibitory effect of Rosa rubiginosa L. extract on supercoiled DNA strand scission caused by peroxyl radicals was assessed using a method described by Chandrasekara and Shahidi [19]. Supercoiled plasmid DNA (pBR 322 from Escherichia coli RRI) was prepared at 50 μg·mL−1 in a 0.5 M phosphate-buffered solution (PBS). In a 100 μL Eppendorf tube, 2 μL of supercoiled plasmid DNA solution was mixed with 4 μL of 2,2′-Azobis(2-amidinopropane) dihydrochloride (AAHP), 2 μL of PBS and 2 μL of three different concentrations of the sample (135, IC50 and 2 μg/mL). The above mixture was kept for incubation at 37 °C for 20 min, protected from light. A control sample, containing only the plasmid DNA, and a blank, containing the plasmid DNA and the oxidant AAHP, were also prepared. At the end of the incubation, 2 μL of the loading dye was incorporated into the mixture. The prepared samples were electrophoresed utilizing a 0.8% agarose gel, made with Tris-borate-EDTA (TBE) buffer, at 100 V for 2 h. The bands were observed under UV light using the MiniBIS Pro device (DNR Bio-Imaging Systems Ltd., Neve Yamin, Israel) to evaluate the level of DNA scission [20]. The protective impact of the phenolic compounds included in the extract was determined according to the following formula:
2.3. Cell-Based Assessments
2.3.1. 2D Human Cell Cultures
Normal human dermal fibroblasts (NHDF) (Lonza, Walkersville, MA, USA) and normal human epidermal keratinocytes (NHEK) (Lonza, Walkersville, MA, USA) were cultured according to the manufacturer’s guidelines, as described previously [12]. For the RNA isolation, both cell lines were cultured in 6-well plates until they reached 95% confluence.
2.3.2. UVA Irradiation Treatment
Cell cultures were seeded at a density of 2.5 × 104 cells/well in 24-well plates and incubated with the extract (0.75 μg/mL) for 48 h. All cells were washed twice with phosphate-buffered saline (PBS) and covered with a thin layer of PBS before exposure to UVA light. The fibroblasts and keratinocytes were exposed to 10 J/cm2 and 25 J/cm2, respectively, of UVA light (lamp equipped with a 400 w ozone-free Philips HPA lamp), according to previous reports [14,15]. The range of UV type was between 300 and 400 nm. The 24-well plates were placed at a distance of 20 cm from UVA light for 30 min.
2.3.3. Cell Viability Assessments
ATP Determination
Cell viability was evaluated with the determination of intracellular levels of ATP utilizing an ATP kit (Thermo Fisher Scientific, Waltham, MA, USA). The determination of the intracellular levels of ATP was in accordance with the manufacturer’s directions, as previously described [16,17]. Specifically, cells were incubated for 48 h with different concentrations of the extract (10 μg/mL, 5 μg/mL, 1 μg/mL, 0.5 μg/mL) before ATP determination. The intracellular levels of ATP were determined under or without UVA irradiation. The outcome of this process was based on three independent experiments along with 8 technical repetitions in each experiment.
2.4. RNA Isolation and cDNA Synthesis
Total RNA was isolated and purified utilizing the Nucleospin RNA kit (Macherey-Nagel, Düren, Germany). The treatment with DNAse was performed on a column, following the manufacturer’s instruction. The RNA concentration and quality were determined by Nannodrop and agarose gel electrophoresis. For the cDNA synthesis, 50 ng of the total RNA was used, applying a PrimeScript™ RT kit (Takara Bio Inc., Kusatsu, Shiga, Japan). In this study, 20 genes related to the antioxidant and inflammation pathways were analyzed, but only those showing significant changes are presented here. The initial analysis targeted the following genes: GPX1 (glutathione peroxidase 1), GPX4 (glutathione peroxidase 4), SOD1 (superoxide dismutase 1), NRF2 (NFE2-like bZIP transcription factor 2), CAT (catalase), COL3A1 (collagen type III alpha 1 chain), IL1A (interleukin 1 alpha), IL1Β (interleukin 1 beta), IL6 (interleukin 6), IL8 (interleukin 8), TNFa (tumor necrosis alpha), DEFB4 (defensin beta 4A), occludin (OCLN), inhibin subunit beta A (INHBA), UDP-glucose ceramide glucosyltransferase (UGCG), b-glycocerebrosidase (GBA1), sirtuin1 (SIRT1), forkhead box O3 (FOXO3), CD44 molecule (CD44), matrix metallopeptidase 14 (MMP14).
Reverse-Transcription PCR and Real-Time PCR
The RT-qPCR reactions occurred in a Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) using SYBR Select Master Mix (Applied Biosystems, Foster City, CA, USA) and gene-specific primers at a final concentration of 0.5 µM each. The relative gene expression was based on the comparative Ct (2−∆∆Ct) method, while the normalization was based on two reference genes: ACTB and GAPDH [18,19]. Eight different experimental states were evaluated during the process: untreated NHEK cells under or without UVA irradiation; untreated NHDF cells under or without UVA irradiation; NHEK cells treated with the extract (0.5 µg/mL) under or without UVA irradiation; and NHDF cells treated with the extract (0.5 µg/mL) under or without UVA irradiation. In addition, Supplemental Table S1 depicts the relative expressions of gens in all samples, while primers of the PCR reactions are presented in Supplemental Table S2. The whole analysis was based on three biological replicates and eight technical repetitions.
2.5. Statistical Analysis
Statistical analysis was performed using the GraphPad Prism program version 5 (GraphPad Software Inc., San Diego, CA, USA). All outcomes of the biochemical assays describe the means of independent experiments (each individual experiment with three biological replicates and eight technical repeats). Additionally, the outcomes presented in the figures pertain to statistical analysis using analysis of variance (ANOVA) with Bonferroni multiple comparisons. The level of significance for the analysis was below 0.05 (p-value < 0.05).
3. Results
3.1. Antioxidant Assessments
3.1.1. DPPH and Folin–Ciocalteu Method
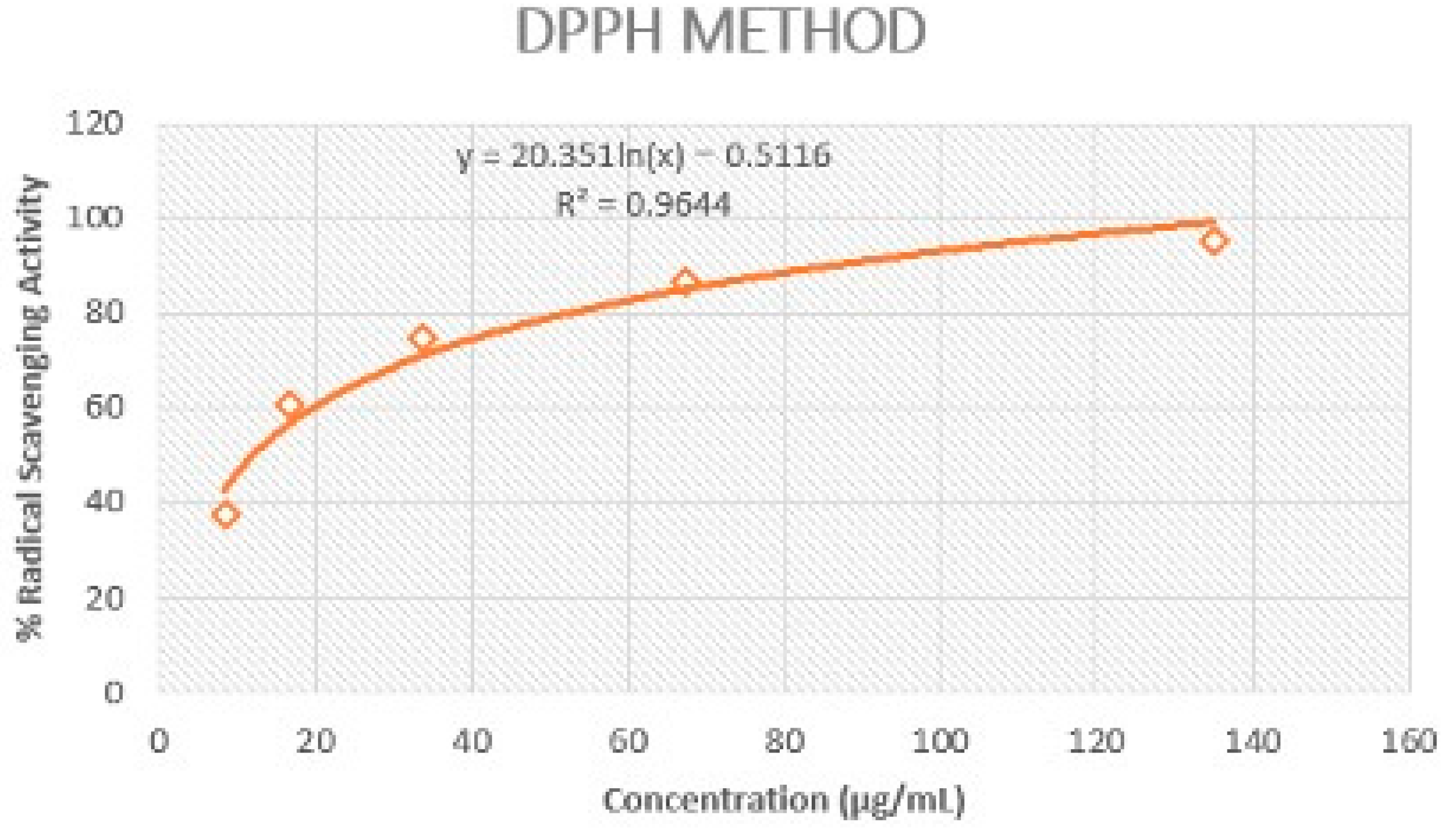
The extract of Rosa rubiginosa L. exhibited strong DPPH scavenging activity (IC50 = 11.97 ± 0.42 μg/mL) in comparison to the value recorded for gallic acid (IC50 = 6.96 ± 0.25 μg/mL) (Figure 1). The strong antioxidant potential of this plant indicated its effective protection against oxidative damage.
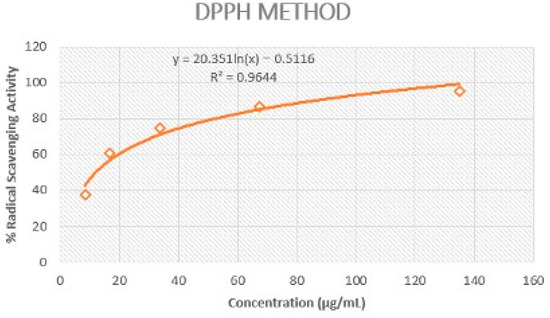
Figure 1.
Representation of Rosa rubiginosa L. extract’s ability to scavenge DPPH radical.
In addition, phenolic compounds are essential plant components, possessing redox characteristics that contribute to their capacity to scavenge free radicals due to their hydroxyl groups [6]. The total phenolic content in the tested sample was evaluated using the Folin–Ciocalteu method. The amount of phenolic compounds present in the extract was 633.93 ± 0.14 μg GAE/mg DW.
3.1.2. Prevention of Supercoiled Plasmid DNA Strand Breakage
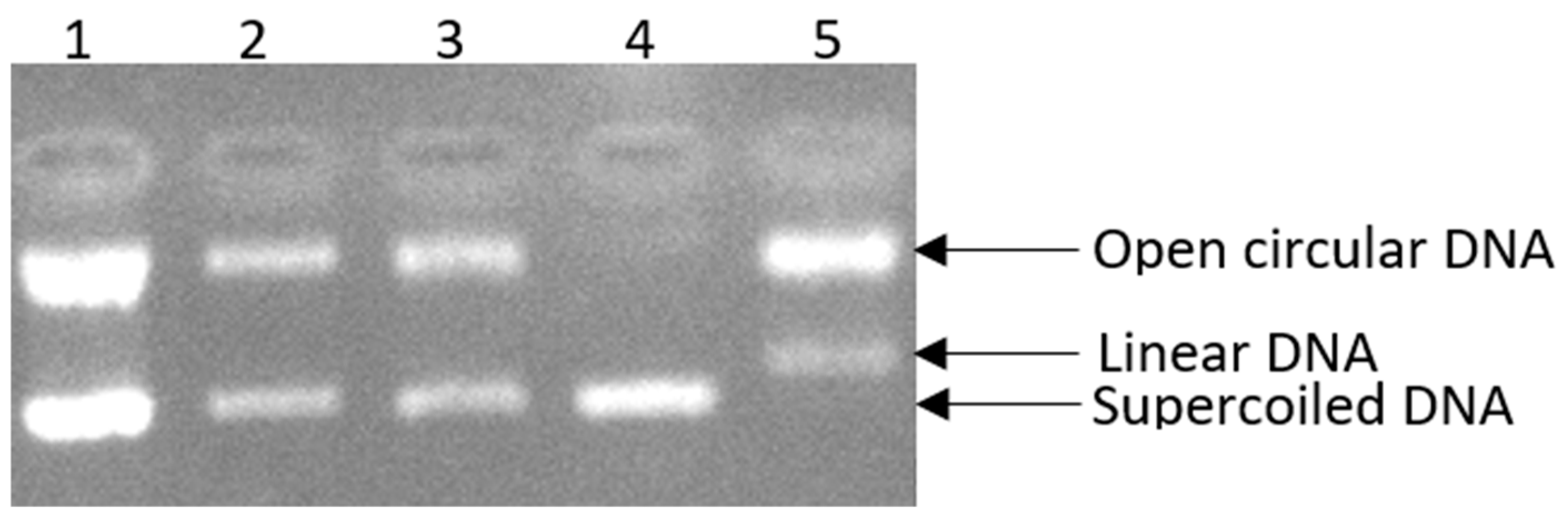
Here, we assessed the ability of the Rosa rubiginosa L. extract to inhibit DNA supercoiled strand breakage induced by peroxyl radicals. This extract, which belongs to the Rosaceae family, was examined at three different concentrations (135, 11.97 (IC50), 2 μg/mL) to assess the proportion of supercoiled DNA retained (Figure 2, Lane 2, Lane 3). In particular, the highest tested concentration (135 μg∙mL−1) of the extract was shown to be the most efficient in protecting against oxidative damage, inhibiting DNA scission by 98.39% (Figure 2, Lane 1). Furthermore, the extract at a concentration of 11.97 μg∙mL−1 exhibited strong activity, achieving 85.15% inhibition (Figure 2, Lane 2), while the extract at a lower concentration (2 μg∙mL−1) provided protection against DNA damage induced by peroxyl radicals, with an inhibition rate of 75.60% (Figure 2, Lane 3).
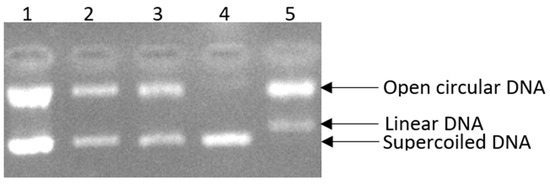
Figure 2.
The positive influence of Rosa rubiginosa L. extract at three different concentrations on pBR322 plasmid DNA damage. Lane 1: DNA + 30 mM AAHP + 135 μg/mL sample; Lane 2: DNA + 30 mM AAHP + 11.97 μg/mL (IC50) sample; Lane 3: DNA + 30 mM AAHP + 2 μg/mL sample; Lane 4: DNA only; Lane 5: DNA + 30 mM AAHP.
3.2. In Vitro Assessments for Two-Dimensional Skin Models
3.2.1. ATP Assessments
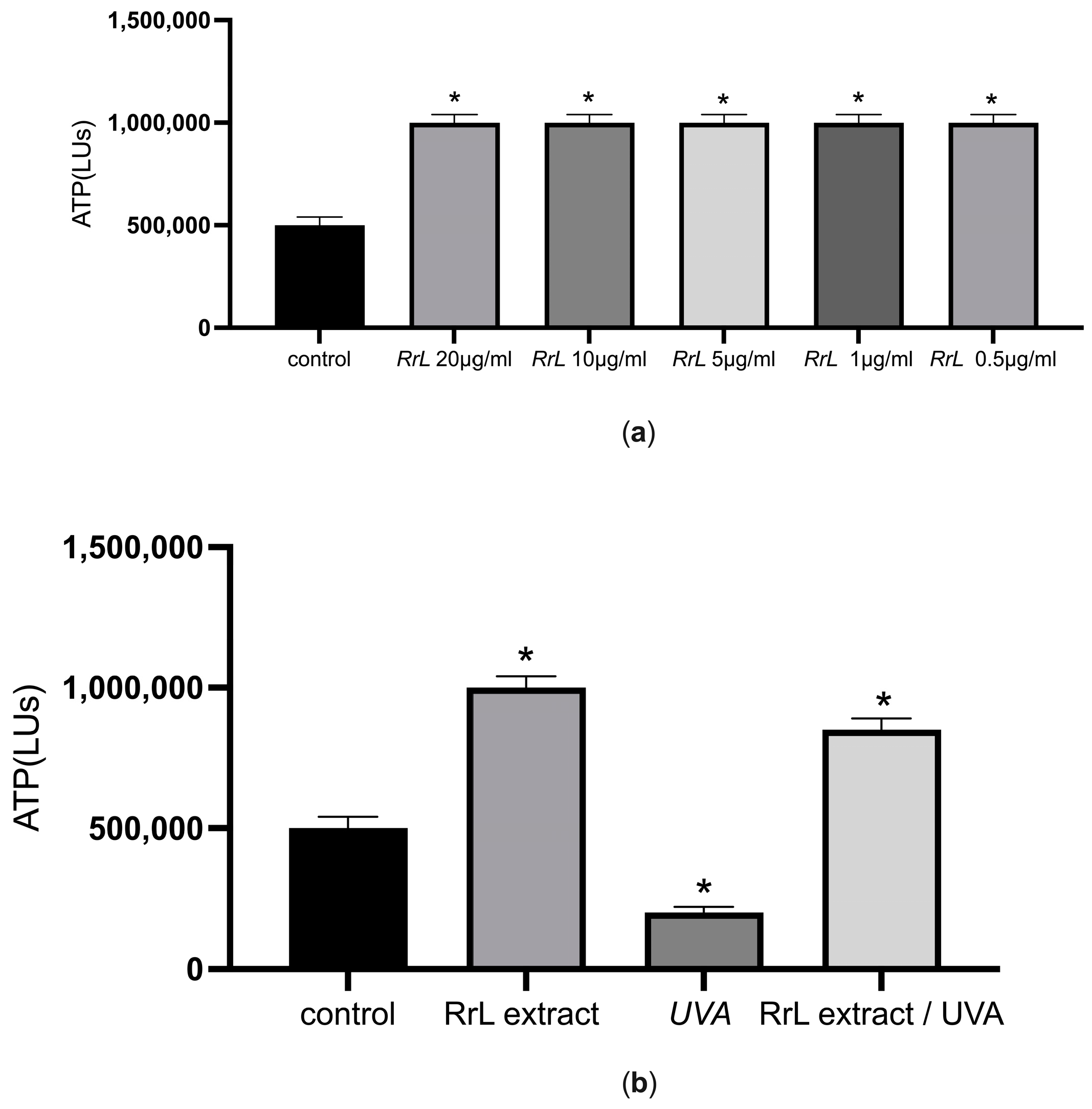
Initially, cell viability was evaluated in order to gain an insight into the potential cytotoxicity of the extracts in human fibroblasts and keratinocytes. For this purpose, we determined the intracellular levels of ATP after treatment with different concentrations of the extract (10–0.5 μg/mL) in 2D cell cultures. Figure 3a,b present the cell viability assessments in NHDF cells under or without UVA irradiation. According to Figure 3a, the intracellular levels of ATP increased after treatment of NHDF cells with different concentrations of the extract (p < 0.05) compared to the control (untreated NHDF cells). In addition, we explored cell viability under UVA irradiation after treatment of NHDF cells with the lowest concentration for 48 h (Figure 3b). According to Figure 3b, the extract maintained the same intracellular levels of ATP as the control after treatment with UVA irradiation (p < 0.05). Interestingly, the intracellular levels of ATP cells treated with the extract after treatment with UVA irradiation were significantly higher compared to the intracellular levels of ATP in cells under UVA irradiation without the extract (p < 0.05), suggesting the energy-promoting effect of the extract under UVA irradiation. The same profile applies for NHEK after the same treatment.
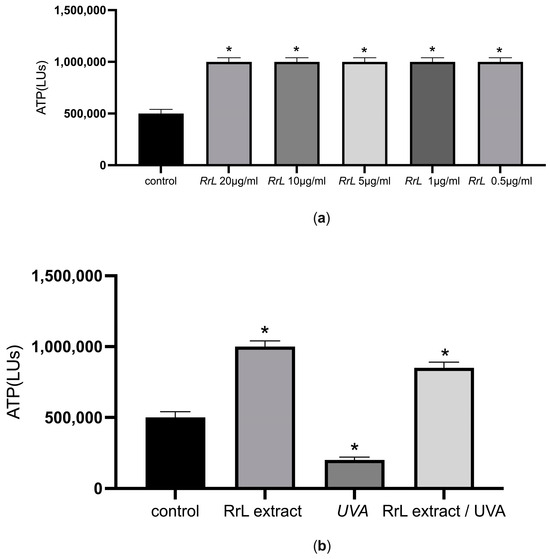
Figure 3.
(a) ATP intracellular levels (LUs) indicated as mean ± SEM for control and NHDF cells treated with the Rosa rubiginosa L. leaf extract (10–0.5 μg/mL). * p < 0.05 significant difference from the control (ANOVA test); (b) ATP intracellular levels (LUs) indicated as mean ± SEM for control: untreated NHDF cells, RrL: NHDF cells treated with 0.5 μg/mL of the Rosa rubiginosa L. leaf extract, UVA: NHDF cells under UVA irradiation and RrL//UVA: NHDF cells treated with 0.5 μg/mL of the Rosa rubiginosa L. leaf extract under UVA irradiation. * p < 0.05 significant difference from the control (ANOVA test).
3.2.2. RT-PCR Analysis
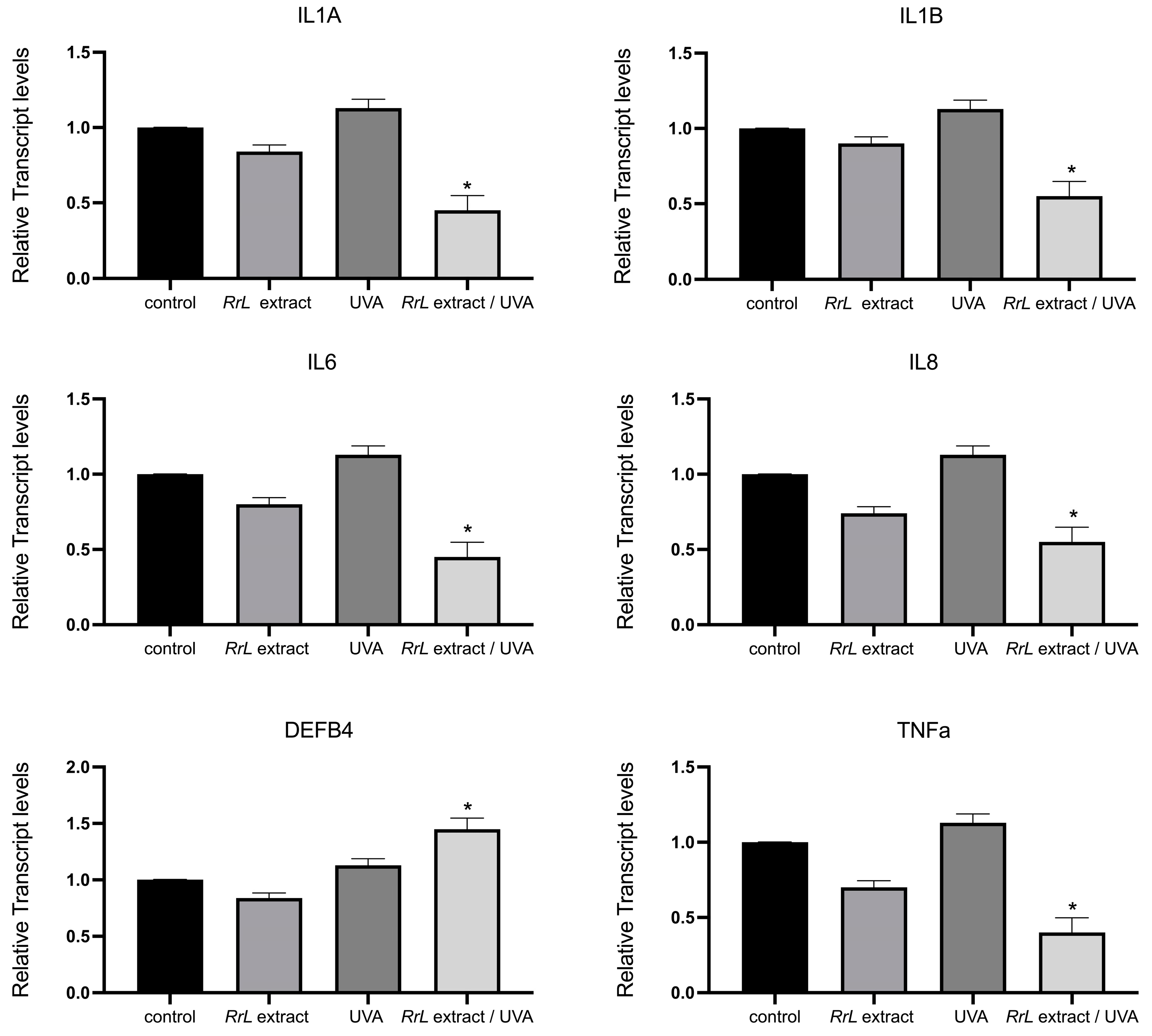
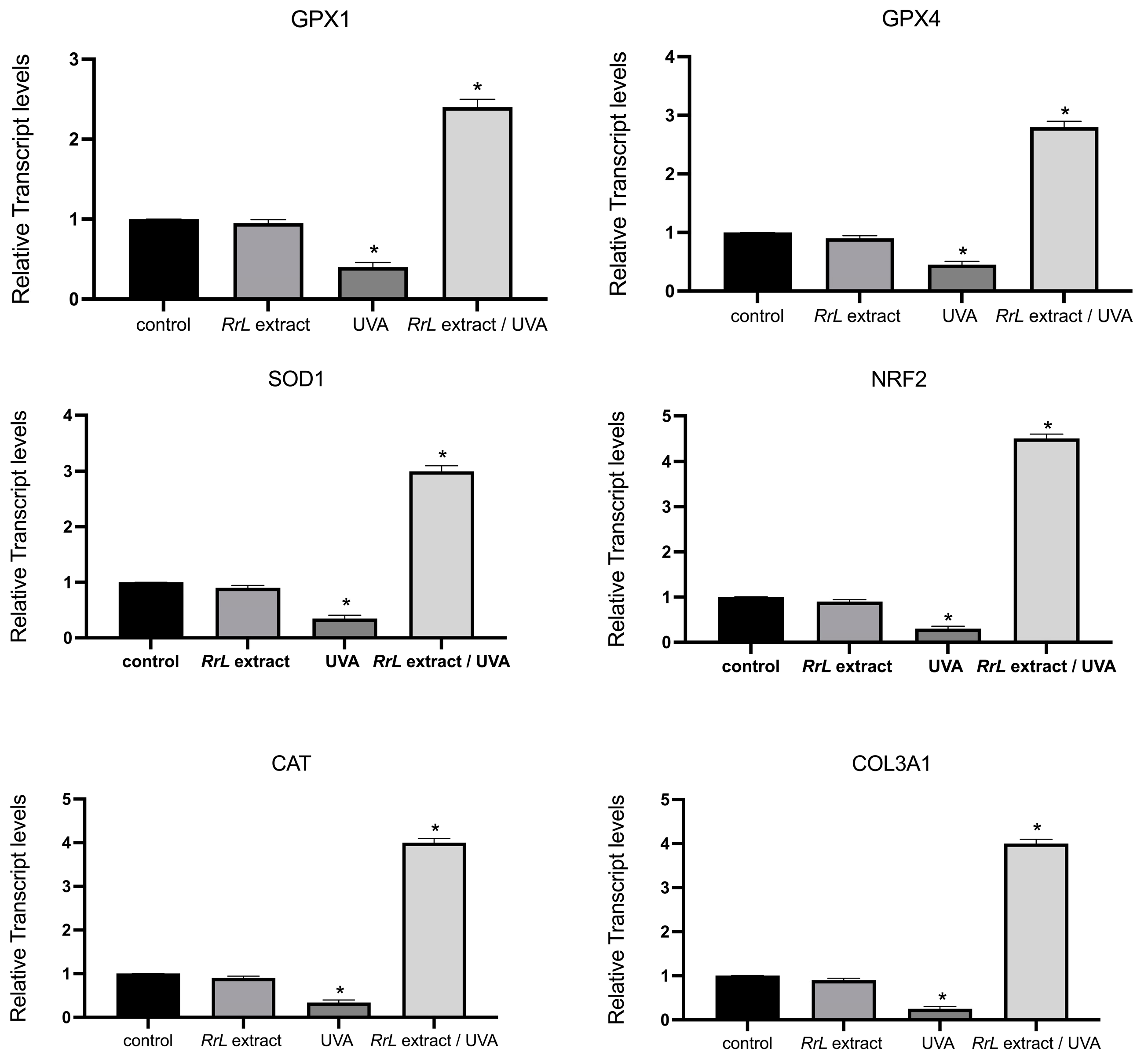
In order to gain an insight into the biological function of Rosa rubiginosa L. leaf extract in human fibroblasts and keratinocytes under UVA irradiation, we studied an array of transcript accumulations of genes involved in antioxidant response, inflammation, cell proliferation and DNA damage. However, in this study, we only present the significant outcome of our investigation. Figure 4 and Figure 5 present the transcript accumulation of genes that were modified with the addition of Rosa rubiginosa L. leaf extract in NHEK and NHDF cells, respectively, under UVA or without irradiation. Specifically, in Figure 4, the transcript accumulation of the genes IL1A (interleukin 1 alpha), IL1B (interleukin 1 beta), IL6 (interleukin 6), IL8 (interleukin 8), TNFa (tumor necrosis alpha) was downregulated, while the transcript accumulation of the gene DEFB4 (defensin beta 4A) was upregulated in NHEK cells treated with the Rosa rubiginosa L. leaf extract (0.5 μg/mL) under UVA irradiation as compared to all experimental conditions (p < 0.05). In addition, Figure 5 demonstrates that the transcript accumulation of the genes GPX1 (glutathione peroxidase 1), GPX4 (glutathione peroxidase 4), SOD1 (superoxide dismutase 1), NRF2 (NFE2-like bZIP transcription factor 2), CAT (catalase), COL3A1 (collagen type III alpha 1 chain) was upregulated in NHDF cells treated with the Rosa rubiginosa L. leaf extract (0.5 μg/mL) under UVA irradiation as compared to all experimental states (p < 0.05). Of note, the transcript accumulation of these genes was severely downregulated under UVA irradiation compared to all experimental states (p < 0.05).
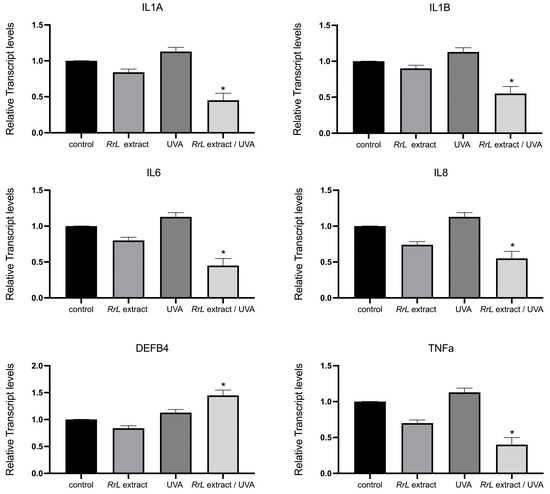
Figure 4.
Gene expression levels of IL1A, IL1B, IL6, IL8, TNFa, DEFB4 in NHEK cells, indicated as a fold change ± SEM. Experimental conditions: control: untreated NHEK cells, RrL: NHEK cells treated with the Rosa rubiginosa L. leaf extract (0.5 μg/mL), UVA: NHEK cells under UVA irradiation, RrL/UVA: NHEK cells treated with the Rosa rubiginosa L. leaf extract (0.5 μg/mL) under UVA irradiation. For the transcriptomic analysis, ACTB and GADPH were used as internal reference genes. * p < 0.05 indicates groups significantly different from the control (ANOVA test).
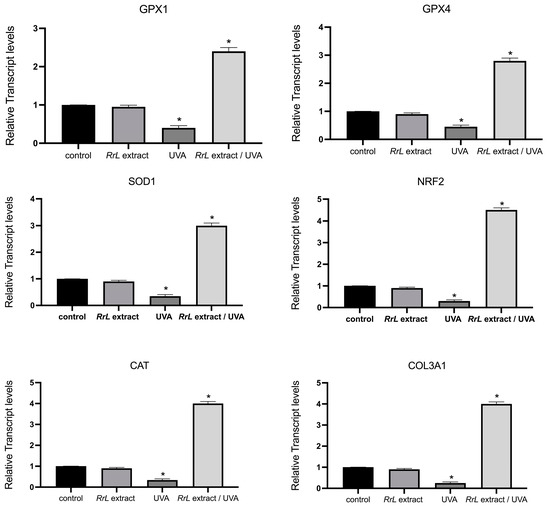
Figure 5.
Gene expression levels of GPX1, GPX4, SOD1, NRF2, CAT, COL3A1 in NHDF cells, indicated as a fold change ± SEM. Experimental conditions: control: untreated NHDF cells, RrL: NHDF cells treated with the Rosa rubiginosa L. leaf extract (0.5 μg/mL), UVA: NHDF cells under UVA irradiation, RrL/UVA: NHDF cells treated with the Rosa rubiginosa L. leaf extract (0.5 μg/mL) under UVA irradiation. For the transcriptomic analysis, ACTB and GADPH were used as internal reference genes. * p < 0.05 indicates groups significantly different from the control (ANOVA test).
4. Discussion
In this study, we explored the biological effects of Rosa rubiginosa L. leaf extract on 2D keratinocyte- and fibroblast-based skin models to shed light on the use of Rosa rubiginosa L. leaf extract as a bio-functional ingredient in cosmeceuticals or nutraceuticals. To elucidate its antioxidant role in vitro, we focused on the oxidative stress induced by UVA irradiation. Oxidative stress is a condition, which occurs when the generation and aggregation of free radicals in the living organisms exceed a biological system’s capacity to neutralize these reactive substances. These free radicals, reactive oxygen species (ROS) and reactive nitrogen species (RNS), can potentially damage all three main types of biological molecules—nucleic acids, proteins and lipids—due to their high reactivity. As a result, the overproduction of free radicals can lead to various pathological disorders, as well as aging [21]. In addition, UVA irradiation can penetrate the skin to a depth of 3–4 mm, directly harming cellular components and potentially playing a role in skin aging and cancer development [22,23]. The action of both enzymatic and non-enzymatic antioxidants balances the detrimental effects of free radicals on the human body [24]. There are several bioanalytical techniques available for measuring the antioxidant activity of the extracts. Among these, the DPPH scavenging assay is the most widely used and preferred method for evaluating antioxidant potential. The IC50 value is a commonly used and effective way of assessing the DPPH radical scavenging activity [25]. This value describes the quantity of the antioxidant required to reduce the initial DPPH absorption by 50% [26]. According to our results, the extract of Rosa rubiginosa L. exhibited strong DPPH scavenging activity (IC50 = 11.97 ± 0.42 μg/mL) in comparison to the value recorded for gallic acid (IC50 = 6.96 ± 0.25 μg/mL), indicating its effective protection against oxidative damage. This outcome is in accordance with a previous report [6] but quite different from other studies based on Rosa species that present a range of IC50 values of the extracts between 30.12 and 156 μg/mL, depending on the extraction methods [27,28,29]. The variation in IC50 values may be attributed to multiple factors, including the cultivation area, the extraction process, climatic factors, application method and harvest time [29]. The amount of phenolic compounds present in the extract was 633.93 ± 0.14 μgGAE/mgDW, lower than in previous limited studies [6,30] but quite comparable with studies based on other Rosa species (326.3–575.1 mgGAE/100 gDW) [27,31]. Similarly, as previously mentioned, the values of total phenolic content of a similar extract may differ depending on the extraction techniques, climate, harvest time, soil, etc.
The beneficial biological activities of the Rosa rubiginosa L. extract include antioxidant, antimicrobial, antifungal and anti-inflammatory properties [5]. In addition, Jiménez-López J. et al. reported that the Rosa rubiginosa L. extract was particularly abundant in phenolic acids and flavonoids, particularly quercetin derivatives [6]. These components exhibit antibacterial, anti-inflammatory, antiviral, anticancer properties, influencing every stage of carcinogenesis, and the ability to regulate lipid peroxidation [32]. The irreparable alteration of DNA caused by oxidative damage has been established as the initial stage in various pathological disorders. Free-radical-induced DNA damage can differ and involves base modification, breakage in the DNA strand, DNA–protein linkages and irregular chromosomal structures [19]. The extract’s ability to inhibit DNA strand scission reflects its positive effect in combating numerous diseases within biological systems [33]. Peroxyl radicals are recognized for causing oxidative damage in biological molecules because of their relatively extended half-life and hence increased tendency to permeate into biological fluids within cells [34]. As shown in our study, supercoiled DNA strands were preserved by the three concentrations of the tested extract, which is rich in polyphenols, as confirmed by the Folin–Ciocalteu method. It has been proven that the phenolic hydroxyl groups in the structures of phenolic compounds can donate electrons or hydrogen atoms [35]. This method allows the flavonoids to inhibit the cleavage of DNA strands and, as a result, the oxidative damage caused by ROS, hence helping retain the supercoiled structure of DNA. The findings of the present study agree with previous research, as it is shown that the Rosa species demonstrate protection against DNA damage induced by ROO• [36].
We also examined the biological effects of the Rosa rubiginosa L. extract on human primary keratinocytes and fibroblasts in vitro to explore its potential applications in skin-related research. Notably, our findings revealed that the extract enhanced the viability of both human epidermal keratinocytes and dermal fibroblasts even under oxidative stress induced by UVA irradiation. The observed rise in intracellular ATP levels in the cells treated with the extract compared to other experimental stages, particularly compared with the untreated cells under UVA irradiation, indicates that, at least in vitro, the extract promotes cell viability, proliferation and energy metabolism. Previous studies suggest that this effect is linked to increased mitochondrial activity, energy metabolism and cell growth, further supporting the extract’s non-toxic nature [37,38]. The Rosa rubiginosa L. extract consistently aids mitochondrial function under oxidative stress, which may contribute to delaying the aging process and sustaining essential cellular functions [39,40].
To gain a broader understanding of the effects of the Rosa rubiginosa L. extract on NHEK cells, we analyzed the genes associated with various pathways involved in epidermal cell functions. Our results revealed that several genes were transcriptionally regulated under the study’s experimental conditions. Specifically, in the 2D keratinocyte-based model, we observed that the genes IL1A, IL1Β, IL6, IL8, TNFa, DEFB4 were notably modified under UVA irradiation in the presence of the extract. Specifically, IL1 plays a role in triggering local inflammation and coagulation [41], while IL6 promotes fibrinogen synthesis, contributing to the acute inflammatory phase [42]. IL8 is involved in various inflammation-related processes, including basophil chemotaxis and angiogenesis [42], whereas TNFa serves as a key mediator of inflammation [43].
In this study, we found that the gene expression of IL1, IL6, IL8 and TNFa was significantly downregulated in NHEK cells treated with the Rosa rubiginosa L. extract under UVA irradiation compared to all other experimental conditions, which potentially means that the presence of the extract unlocks the anti-inflammatory response of the cells. It is worth noting that the presence of the extract enhances this anti-inflammatory response, as untreated cells under UVA irradiation showed higher levels for these transcripts compared to the control. While additional studies may be needed to confirm this intriguing outcome, this reduction aligns with previous studies highlighting the extract’s anti-inflammatory properties [44,45,46,47]. This outcome reveals an interesting role of the Rosa rubiginosa L. extract as an anti-inflammatory agent under UVA irradiation, as exposure to UV radiation, environmental pollutants and other external factors can initiate skin inflammation, accelerate inflammaging and contribute to a harmful cycle of chronic inflammation and cellular senescence [42,43].
Additionally, we examined the genes involved in immune responses within the epidermal layer. Human β-defensin 2 (hBD2), encoded by the DEFB4 gene, is an inducible antimicrobial peptide with a molecular mass of 4–6 kDa. It functions as an endogenous antibiotic, protecting the host against Gram-positive and Gram-negative bacteria, fungi and certain enveloped viruses. It plays a key role in innate immunity, as its release is triggered by pro-inflammatory cytokines, endogenous signals, infections or wounds [48,49]. Notably, our study revealed an upregulation of DEFB4 only when NHEK cells were treated with the Rosa rubiginosa L. extract under UVA irradiation, suggesting that the extract may trigger antimicrobial processes, although further research is needed to confirm this.
In the 2D fibroblast-based model, we observed that the genes GPX1, GPX4, SOD1, NRF2, CAT, COL3A1 were modified under UVA irradiation. Specifically, GPX1, GPX4, SOD1, CAT and NRF2 serve as essential antioxidant cellular markers [50,51,52] and are upregulated in response to stress [53,54]. In addition, COL3A1 plays a significant role in regulating the extracellular matrix (ECM) during cellular aging [55]. In our study, we observed that the expression of these gene was upregulated with the addition of Rosa rubiginosa L. extract in UVA-treated NHDF cells compared to the control as well as to cells untreated with the extract under UVA irradiation, suggesting that the presence of the Rosa rubiginosa L. extract in cells induces a potential antioxidant response of NHDF under oxidative stress. Consistent with these findings, the addition of Rosa rubiginosa L. extract under UVA exposure led to an increase in cell viability, confirming its protective effect. This activation of genes GPX1, GPX4, SOD1 and NRF2 under oxidative stress has previously been documented in studies on fibroblasts treated with various extracts during photo-oxidation [56,57,58,59]. In particular, other in vitro studies on different rosa species have shown that dietary Rosa damascena helps prevent UVB-induced skin aging by enhancing collagen production through the inhibition of MMPs in mice, highlighting its use a functional food [60]. Another study highlights the important antioxidant role of 18 Rosa L. cultivars in the melanoma cell line A375 and the normal skin cell line Hs27 due to their high anthocyanic content [61]. In addition, Rosa gallica petal extracts demonstrate anti-aging properties [62]. Additional research indicates that Rosa rugosa extracts and their isolated compounds, rosarugosides A and D, may help prevent and protect against skin aging and damage [63]. Finally, a recent study on a fermented rosa extract exhibited antioxidant, anti-aging properties by inducing the upregulation of COL3A1 and COL1A1 and the downregulation of MMPs in human fibroblasts [64].
In summary, the present study showed that the Rosa rubiginosa L. leaf extract has a high phenolic content, exhibits strong scavenging activity and inhibits the DNA supercoiled strand breakage. Additionally, based on 2D cell-based models, the study revealed that the Rosa rubiginosa L. leaf extract confers cell viability and modulates the expression levels of several genes involved in the inflammation pathway in human keratinocytes and in the antioxidant pathway in human fibroblasts under oxidative stress induced by UVA irradiation.
These findings suggest that the Rosa rubiginosa L. leaf extract is a promising agent to be incorporated in cosmeceuticals or nutraceuticals. However, further research is needed to better understand the molecular mechanisms behind the cytoprotective effects of Rosa rubiginosa L. leaf extract and to evaluate their effectiveness in clinical settings.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cosmetics12020062/s1, Table S1: Relative mRNA expression of genes; Table S2: Gene name, Accesion No, Kegg pathway, Primer.
Author Contributions
Conceptualization, S.L.; methodology, S.L. and A.T. (Aliki Tsakni) formal analysis, S.L., A.T. (Aliki Tsakni), D.A. and D.H.; investigation, S.L., A.T. (Aliki Tsakni) and D.H.; writing—original draft preparation, S.L. and A.T. (Aliki Tsakni); writing—review and editing, A.T. (Alexandros Tsoupras). and D.H.; supervision, S.L. and D.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Michalak, M. Plant-Derived Antioxidants: Significance in Skin Health and the Ageing Process. Int. J. Mol. Sci. 2022, 23, 585. [Google Scholar] [CrossRef] [PubMed]
- Olivero-Verbel, J.; Quintero-Rincón, P.; Caballero-Gallardo, K. Aromatic Plants as Cosmeceuticals: Benefits and Applications for Skin Health. Planta 2024, 260, 132. [Google Scholar] [CrossRef]
- Alves, A.; Sousa, E.; Kijjoa, A.; Pinto, M. Marine-Derived Compounds with Potential Use as Cosmeceuticals and Nutricosmetics. Molecules 2020, 25, 2536. [Google Scholar] [CrossRef] [PubMed]
- Cunja, V.; Mikulic-Petkovsek, M.; Stampar, F.; Schmitzer, V. Compound Identification of Selected Rose Species and Cultivars: An Insight to Petal and Leaf Phenolic Profiles. J. Am. Soc. Hortic. Sci. 2014, 139, 157–166. [Google Scholar] [CrossRef]
- de Santana, F.B.; Gontijo, L.C.; Mitsutake, H.; Mazivila, S.J.; de Souza, L.M.; Borges Neto, W. Non-Destructive Fraud Detection in Rosehip Oil by MIR Spectroscopy and Chemometrics. Food Chem. 2016, 209, 228–233. [Google Scholar] [CrossRef]
- Jiménez-López, J.; Ruiz-Medina, A.; Ortega-Barrales, P.; Llorent-Martínez, E.J. Rosa rubiginosa and Fraxinus oxycarpa Herbal Teas: Characterization of Phytochemical Profiles by Liquid Chromatography-Mass Spectrometry, and Evaluation of the Antioxidant Activity. New J. Chem. 2017, 41, 7681–7688. [Google Scholar] [CrossRef]
- Farhan, M. The Promising Role of Polyphenols in Skin Disorders. Molecules 2024, 29, 865. [Google Scholar] [CrossRef]
- Son, D.-H.; Nam, M.-H.; Hong, C.-O.; Seol, H.-M.; Yang, J.-E.; Kim, Y.-B.; Kim, C.-T.; Lee, K.-W. 5-α Reductase Inhibitory Effect and Astringent Activity of Green Apple Rind Extract on Human Keratinocytes and Fibroblast Cells. Biosci. Biotechnol. Biochem. 2013, 77, 714–721. [Google Scholar] [CrossRef]
- Choi, E.-K.; Guo, H.; Choi, J.-K.; Jang, S.-K.; Shin, K.; Cha, Y.-S.; Choi, Y.; Seo, D.-W.; Lee, Y.-B.; Joo, S.-S.; et al. Extraction Conditions of White Rose Petals for the Inhibition of Enzymes Related to Skin Aging. Lab. Anim. Res. 2015, 31, 148. [Google Scholar] [CrossRef]
- Lee, M.; Nam, T.G.; Lee, I.; Shin, E.J.; Han, A.; Lee, P.; Lee, S.; Lim, T. Skin Anti-inflammatory Activity of Rose Petal Extract (Rosa gallica) through Reduction of MAPK Signaling Pathway. Food Sci. Nutr. 2018, 6, 2560–2567. [Google Scholar] [CrossRef]
- Maželienė, Ž.; Kirvaitienė, J.; Kaklauskienė, K.; Volskienė, R.; Aleksandravičienė, A. Antifungal and Antibacterial Activity of Aqueous and Ethanolic Extracts of Different Rosa rugosa Parts. Microbiol. Res. 2025, 16, 26. [Google Scholar] [CrossRef]
- Ćujić, N.; Šavikin, K.; Janković, T.; Pljevljakušić, D.; Zdunić, G.; Ibrić, S. Optimization of Polyphenols Extraction from Dried Chokeberry Using Maceration as Traditional Technique. Food Chem. 2016, 194, 135–142. [Google Scholar] [CrossRef]
- Velickovic, D.; Nikolova, M.; Ivancheva, S.; Stojanovic, J.; Veljkovic, V. Extraction of Flavonoids from Garden (Salvia officinalis L.) and Glutinous (Salvia glutinosa L.) Sage by Ultrasonic and Classical Maceration. J. Serbian Chem. Soc. 2007, 72, 73–80. [Google Scholar] [CrossRef]
- Thakur, L.; Ghodasra, U.; Patel, N.; Dabhi, M. Novel Approaches for Stability Improvement in Natural Medicines. Pharmacogn. Rev. 2011, 5, 48. [Google Scholar] [CrossRef]
- Pinheiro, A.C.; Nunes, I.J.; Ferreira, W.V.; Tomasini, P.P.; Trindade, C.; Martins, C.C.; Wilhelm, E.A.; Oliboni, R.d.S.; Netz, P.A.; Stieler, R.; et al. Antioxidant and Anticancer Potential of the New Cu(II) Complexes Bearing Imine-Phenolate Ligands with Pendant Amine N-Donor Groups. Pharmaceutics 2023, 15, 376. [Google Scholar] [CrossRef] [PubMed]
- Tsakni, A.; Kyriakopoulou, E.; Letsiou, S.; Halvatsiotis, P.; Rigopoulos, H.; Vassilaki, N.; Houhoula, D. In Vitro Determination of Antimicrobial, Antioxidant and Antiviral Properties of Greek Plant Extracts. Microorganisms 2025, 13, 177. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Prior, R.L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Pérez, M.; Dominguez-López, I.; Lamuela-Raventós, R.M. The Chemistry Behind the Folin–Ciocalteu Method for the Estimation of (Poly)Phenol Content in Food: Total Phenolic Intake in a Mediterranean Dietary Pattern. J. Agric. Food Chem. 2023, 71, 17543–17553. [Google Scholar] [CrossRef]
- Chandrasekara, A.; Shahidi, F. Bioactivities and Antiradical Properties of Millet Grains and Hulls. J. Agric. Food Chem. 2011, 59, 9563–9571. [Google Scholar] [CrossRef]
- Tsakni, A.; Chatzilazarou, A.; Tsakali, E.; Tsantes, A.G.; Van Impe, J.; Houhoula, D. Identification of Bioactive Compounds in Plant Extracts of Greek Flora and Their Antimicrobial and Antioxidant Activity. Separations 2023, 10, 373. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Wang, X.; Heraud, S.; Thépot, A.; Dos Santos, M.; Luo, Z. The Whitening Properties of the Mixture Composed of Pomegranate, Osmanthus and Olive and the Protective Effects Against Ultraviolet Deleterious Effects. Clin. Cosmet. Investig. Dermatol. 2021, 14, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Letsiou, S.; Koldiri, E.; Beloukas, A.; Rallis, E.; Kefala, V. Deciphering the Effects of Different Types of Sunlight Radiation on Skin Function: A Review. Cosmetics 2024, 11, 80. [Google Scholar] [CrossRef]
- Demirci-Çekiç, S.; Özkan, G.; Avan, A.N.; Uzunboy, S.; Çapanoğlu, E.; Apak, R. Biomarkers of Oxidative Stress and Antioxidant Defense. J. Pharm. Biomed. Anal. 2022, 209, 114477. [Google Scholar] [CrossRef]
- Gulcin, İ.; Alwasel, S.H. DPPH Radical Scavenging Assay. Processes 2023, 11, 2248. [Google Scholar] [CrossRef]
- Brighente, I.M.C.; Dias, M.; Verdi, L.G.; Pizzolatti, M.G. Antioxidant Activity and Total Phenolic Content of Some Brazilian Species. Pharm. Biol. 2007, 45, 156–161. [Google Scholar] [CrossRef]
- Fetni, S.; Bertella, N.; Ouahab, A. LC–DAD/ESI–MS/MS Characterization of Phenolic Constituents in Rosa canina L. and Its Protective Effect in Cells. Biomed. Chromatogr. 2020, 34, e4961. [Google Scholar] [CrossRef]
- Fascella, G.; Mammano, M.M.; D’Angiolillo, F. Leaf Methanolic Extracts from Four Sicilian Rose Species: Bioactive Compounds Content and Antioxidant Activity. Acta Hortic. 2019, 1232, 81–88. [Google Scholar] [CrossRef]
- Mourabit, Y.; El Hajjaji, S.; Taha, D.; Badaoui, B.; El Yadini, M.; Rusu, M.E.; Lee, L.-H.; Bouyahya, A.; Bourais, I. HPLC-DAD-ESI/MS Phytochemical Investigation, Antioxidant, and Antidiabetic Activities of Moroccan Rosa canina L. Extracts. Biocatal. Agric. Biotechnol. 2023, 52, 102817. [Google Scholar] [CrossRef]
- Sytar, O.; Hemmerich, I.; Zivcak, M.; Rauh, C.; Brestic, M. Comparative Analysis of Bioactive Phenolic Compounds Composition from 26 Medicinal Plants. Saudi J. Biol. Sci. 2018, 25, 631–641. [Google Scholar] [CrossRef]
- Roman, I.; Stănilă, A.; Stănilă, S. Bioactive Compounds and Antioxidant Activity of Rosa canina L. Biotypes from Spontaneous Flora of Transylvania. Chem. Cent. J. 2013, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Alecu, A.; Albu, C.; Badea, G.-I.; Alionte, A.; Enache, A.-A.; Radu, G.-L.; Litescu, S.-C. Infrared Laser-Assisted Extraction of Bioactive Compounds from Rosa canina L. Int. J. Mol. Sci. 2025, 26, 992. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kaur, M.; Sogi, D.S.; Purewal, S.S. A Comparative Study of Phytochemicals, Antioxidant Potential and in-Vitro DNA Damage Protection Activity of Different Oat (Avena sativa) Cultivars from India. J. Food Meas. Charact. 2019, 13, 347–356. [Google Scholar] [CrossRef]
- Martemucci, G.; Costagliola, C.; Mariano, M.; D’andrea, L.; Napolitano, P.; D’Alessandro, A.G. Free Radical Properties, Source and Targets, Antioxidant Consumption and Health. Oxygen 2022, 2, 48–78. [Google Scholar] [CrossRef]
- Albishi, T.; John, J.A.; Al-Khalifa, A.S.; Shahidi, F. Antioxidant, Anti-Inflammatory and DNA Scission Inhibitory Activities of Phenolic Compounds in Selected Onion and Potato Varieties. J. Funct. Foods 2013, 5, 930–939. [Google Scholar] [CrossRef]
- Kerasioti, E.; Apostolou, A.; Kafantaris, I.; Chronis, K.; Kokka, E.; Dimitriadou, C.; Tzanetou, E.N.; Priftis, A.; Koulocheri, S.D.; Haroutounian, S.A.; et al. Polyphenolic Composition of Rosa canina, Rosa sempervivens and Pyrocantha coccinea Extracts and Assessment of Their Antioxidant Activity in Human Endothelial Cells. Antioxidants 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Crouch, S.P.M.; Kozlowski, R.; Slater, K.J.; Fletcher, J. The Use of ATP Bioluminescence as a Measure of Cell Proliferation and Cytotoxicity. J. Immunol. Methods 1993, 160, 81–88. [Google Scholar] [CrossRef]
- Deters, A.M.; Schröder, K.R.; Hensel, A. Kiwi Fruit (Actinidia chinensis L.) Polysaccharides Exert Stimulating Effects on Cell Proliferation via Enhanced Growth Factor Receptors, Energy Production, and Collagen Synthesis of Human Keratinocytes, Fibroblasts, and Skin Equivalents. J. Cell. Physiol. 2005, 202, 717–722. [Google Scholar] [CrossRef]
- Giampieri, F.; Alvarez-Suarez, J.; Mazzoni, L.; Forbes-Hernandez, T.; Gasparrini, M.; Gonzàlez-Paramàs, A.; Santos-Buelga, C.; Quiles, J.; Bompadre, S.; Mezzetti, B.; et al. Polyphenol-Rich Strawberry Extract Protects Human Dermal Fibroblasts against Hydrogen Peroxide Oxidative Damage and Improves Mitochondrial Functionality. Molecules 2014, 19, 7798–7816. [Google Scholar] [CrossRef]
- Tsai, H.-Z.; Lin, R.-K.; Hsieh, T.-S. Drosophila Mitochondrial Topoisomerase III Alpha Affects the Aging Process via Maintenance of Mitochondrial Function and Genome Integrity. J. Biomed. Sci. 2016, 23, 38. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The Chemokine System in Diverse Forms of Macrophage Activation and Polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, Oxidative Stress and Autophagy in Skin Aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Zhang, W.; Wang, Y.; Qian, P.; Huang, H. Inflammation and Aging: Signaling Pathways and Intervention Therapies. Signal Transduct. Target. Ther. 2023, 8, 239. [Google Scholar] [CrossRef] [PubMed]
- Coltelli, M.-B.; Aliotta, L.; Vannozzi, A.; Morganti, P.; Panariello, L.; Danti, S.; Neri, S.; Fernandez-Avila, C.; Fusco, A.; Donnarumma, G.; et al. Properties and Skin Compatibility of Films Based on Poly(Lactic Acid) (PLA) Bionanocomposites Incorporating Chitin Nanofibrils (CN). J. Funct. Biomater. 2020, 11, 21. [Google Scholar] [CrossRef]
- Azimi, B.; Thomas, L.; Fusco, A.; Kalaoglu-Altan, O.I.; Basnett, P.; Cinelli, P.; De Clerck, K.; Roy, I.; Donnarumma, G.; Coltelli, M.-B.; et al. Electrosprayed Chitin Nanofibril/Electrospun Polyhydroxyalkanoate Fiber Mesh as Functional Nonwoven for Skin Application. J. Funct. Biomater. 2020, 11, 62. [Google Scholar] [CrossRef] [PubMed]
- Solá, P.; Mereu, E.; Bonjoch, J.; Casado-Peláez, M.; Prats, N.; Aguilera, M.; Reina, O.; Blanco, E.; Esteller, M.; Di Croce, L.; et al. Targeting Lymphoid-Derived IL-17 Signaling to Delay Skin Aging. Nat. Aging 2023, 3, 688–704. [Google Scholar] [CrossRef]
- Liang, J.; Dai, W.; Liu, C.; Wen, Y.; Chen, C.; Xu, Y.; Huang, S.; Hou, S.; Li, C.; Chen, Y.; et al. Gingerenone A Attenuates Ulcerative Colitis via Targeting IL-17RA to Inhibit Inflammation and Restore Intestinal Barrier Function. Adv. Sci. 2024, 11, 2400206. [Google Scholar] [CrossRef]
- Donnarumma, G.; Paoletti, I.; Fusco, A.; Perfetto, B.; Buommino, E.; de Gregorio, V.; Baroni, A. β-Defensins: Work in Progress. In Advances in Microbiology, Infectious Diseases and Public Health; Springer: Cham, Switzerland, 2015; pp. 59–76. [Google Scholar]
- Fusco, A.; Savio, V.; Cammarota, M.; Alfano, A.; Schiraldi, C.; Donnarumma, G. Beta-Defensin-2 and Beta-Defensin-3 Reduce Intestinal Damage Caused by Salmonella typhimurium Modulating the Expression of Cytokines and Enhancing the Probiotic Activity of Enterococcus faecium. J. Immunol. Res. 2017, 2017, 6976935. [Google Scholar] [CrossRef]
- Gęgotek, A.; Skrzydlewska, E. The Role of Transcription Factor Nrf2 in Skin Cells Metabolism. Arch. Dermatol. Res. 2015, 307, 385–396. [Google Scholar] [CrossRef]
- Gurjala, A.N.; Liu, W.R.; Mogford, J.E.; Procaccini, P.S.A.; Mustoe, T.A. Age-Dependent Response of Primary Human Dermal Fibroblasts to Oxidative Stress: Cell Survival, pro-Survival Kinases, and Entrance into Cellular Senescence. Wound Repair Regen. 2005, 13, 565–575. [Google Scholar] [CrossRef]
- Schroeder, P.; Pohl, C.; Calles, C.; Marks, C.; Wild, S.; Krutmann, J. Cellular Response to Infrared Radiation Involves Retrograde Mitochondrial Signaling. Free Radic. Biol. Med. 2007, 43, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.; Jubri, Z.; Rajab, N.; Rahim, K.; Yusof, Y.; Makpol, S. Gelam Honey Protects against Gamma-Irradiation Damage to Antioxidant Enzymes in Human Diploid Fibroblasts. Molecules 2013, 18, 2200–2211. [Google Scholar] [CrossRef]
- Hseu, Y.-C.; Chang, C.-T.; Gowrisankar, Y.V.; Chen, X.-Z.; Lin, H.-C.; Yen, H.-R.; Yang, H.-L. Zerumbone Exhibits Antiphotoaging and Dermatoprotective Properties in Ultraviolet A-Irradiated Human Skin Fibroblast Cells via the Activation of Nrf2/ARE Defensive Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 4098674. [Google Scholar] [CrossRef]
- Kim, Y.U.; Gi, H.; Jeong, E.K.; Han, S.; Seo, W.-Y.; Kim, Y.J.; Lee, S.B.; Kim, K. Development of a Highly Effective Recombinant Protein from Human Collagen Type III Alpha 1 (COL3A1) to Enhance Human Skin Cell Functionality. BMB Rep. 2024, 57, 424–429. [Google Scholar] [CrossRef]
- Neumann, U.; Louis, S.; Gille, A.; Derwenskus, F.; Schmid-Staiger, U.; Briviba, K.; Bischoff, S.C. Anti-Inflammatory Effects of Phaeodactylum tricornutum Extracts on Human Blood Mononuclear Cells and Murine Macrophages. J. Appl. Phycol. 2018, 30, 2837–2846. [Google Scholar] [CrossRef]
- Gasparrini, M.; Forbes-Hernandez, T.Y.; Giampieri, F.; Afrin, S.; Alvarez-Suarez, J.M.; Mazzoni, L.; Mezzetti, B.; Quiles, J.L.; Battino, M. Anti-Inflammatory Effect of Strawberry Extract against LPS-Induced Stress in RAW 264.7 Macrophages. Food Chem. Toxicol. 2017, 102, 1–10. [Google Scholar] [CrossRef]
- Zheng, Q.; Yang, S.-J.; Yi, E.-J.; Park, S.-J.; Jin, X.; Nguyen, T.T.M.; Yi, G.-S.; Jeon, Y.-J.; Yi, T.-H. Enzyme-Assisted Rosa Davurica Mitigates UV-Induced Skin Photodamage by Modulating Apoptosis through Nrf2/ARE and MAPK/NF-ΚB Pathways. J. Photochem. Photobiol. B 2025, 263, 113098. [Google Scholar] [CrossRef]
- Abdelkader, M.-A.E.; Mediatrice, H.; Lin, D.; Lin, Z.; Aggag, S.A. Mitigating Oxidative Stress and Promoting Cellular Longevity with Mushroom Extracts. Foods 2024, 13, 4028. [Google Scholar] [CrossRef]
- Park, B.; Hwang, E.; Seo, S.A.; Zhang, M.; Park, S.-Y.; Yi, T.-H. Dietary Rosa Damascena Protects against UVB-Induced Skin Aging by Improving Collagen Synthesis via MMPs Reduction through Alterations of c-Jun and c-Fos and TGF-Β1 Stimulation Mediated Smad2/3 and Smad7. J. Funct. Foods 2017, 36, 480–489. [Google Scholar] [CrossRef]
- Faur, C.-A.; Zăhan, M.; Bunea, C.I.; Hârșan, E.; Bora, F.-D.; Bunea, A. Antiproliferative and Biochemical Evaluation of Rose Extracts: Impact on Tumor and Normal Skin Cells. Front. Plant Sci. 2024, 15, 1477243. [Google Scholar] [CrossRef]
- Shin, E.J.; Han, A.-R.; Lee, M.-H.; Song, Y.-R.; Lee, K.M.; Nam, T.-G.; Lee, P.; Lee, S.-Y.; Lim, T.-G. Extraction Conditions for Rosa Gallica Petal Extracts with Anti-Skin Aging Activities. Food Sci. Biotechnol. 2019, 28, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Son, S.-R.; Choi, Y.J.; Kim, Y.; Ahn, S.-Y.; Jang, D.S.; Lee, S. Rosarugosides A and D from Rosa rugosa Flower Buds: Their Potential Anti-Skin-Aging Effects in TNF-α-Induced Human Dermal Fibroblasts. Plants 2024, 13, 1266. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.; Hao, Y.; Chen, Y.; Yu, S.; Qu, L. Chemical Constituents and Bioactivities of Fermented Rose (from Yunnan) Extract. Nat. Prod. Res. 2024, 1–8. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).