
Bone Morphogenic Proteins and Their Antagonists in the Lower Airways of Stable COPD Patients

 ,
,  , , ,
, , ,  , and
, and
Abstract
Simple Summary
Abstract
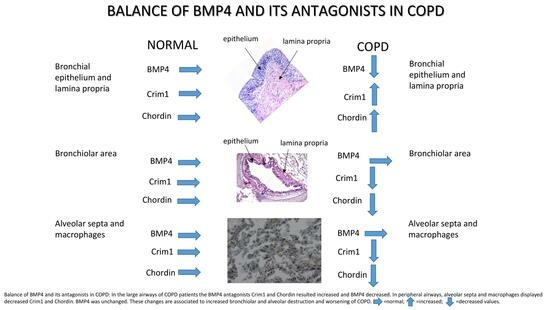
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Lung Function Tests and Volumes
2.3. Fiberoptic Bronchoscopy, Collection, and Processing of Bronchial Biopsies
2.4. Collection and Processing of the Peripheral Lung Tissue
2.5. Immunohistochemistry on OCT-Embedded Bronchial Biopsies
2.6. Immunohistochemistry in Human Peripheral Lung Tissue
2.7. Scoring System for Immunohistochemistry in the Bronchial Biopsies
2.8. Scoring System for Immunohistochemistry in the Peripheral Lung Tissue
2.9. RNA Extraction and Sequencing from Bronchial Rings and Lung Specimens
2.10. Analysis of RNA-seq Data
2.11. Cell Culture and Treatments
2.12. ELISA Tests in the Supernatants of Treated and Non-Treated 16HBE Cells
2.13. BrdU Test for Cell Proliferation
2.14. Statistical Analysis Applied to Functional and Morphological Data
3. Results
3.1. Clinical Characteristics of Subjects Providing Bronchial Biopsies
3.2. Immunohistochemistry for BMPs and BMP Antagonists in the Bronchial Epithelium of Bronchial Biopsies
3.3. Immunohistochemistry for BMPs and BMP Antagonists in the Bronchial Lamina Propria of Bronchial Biopsies
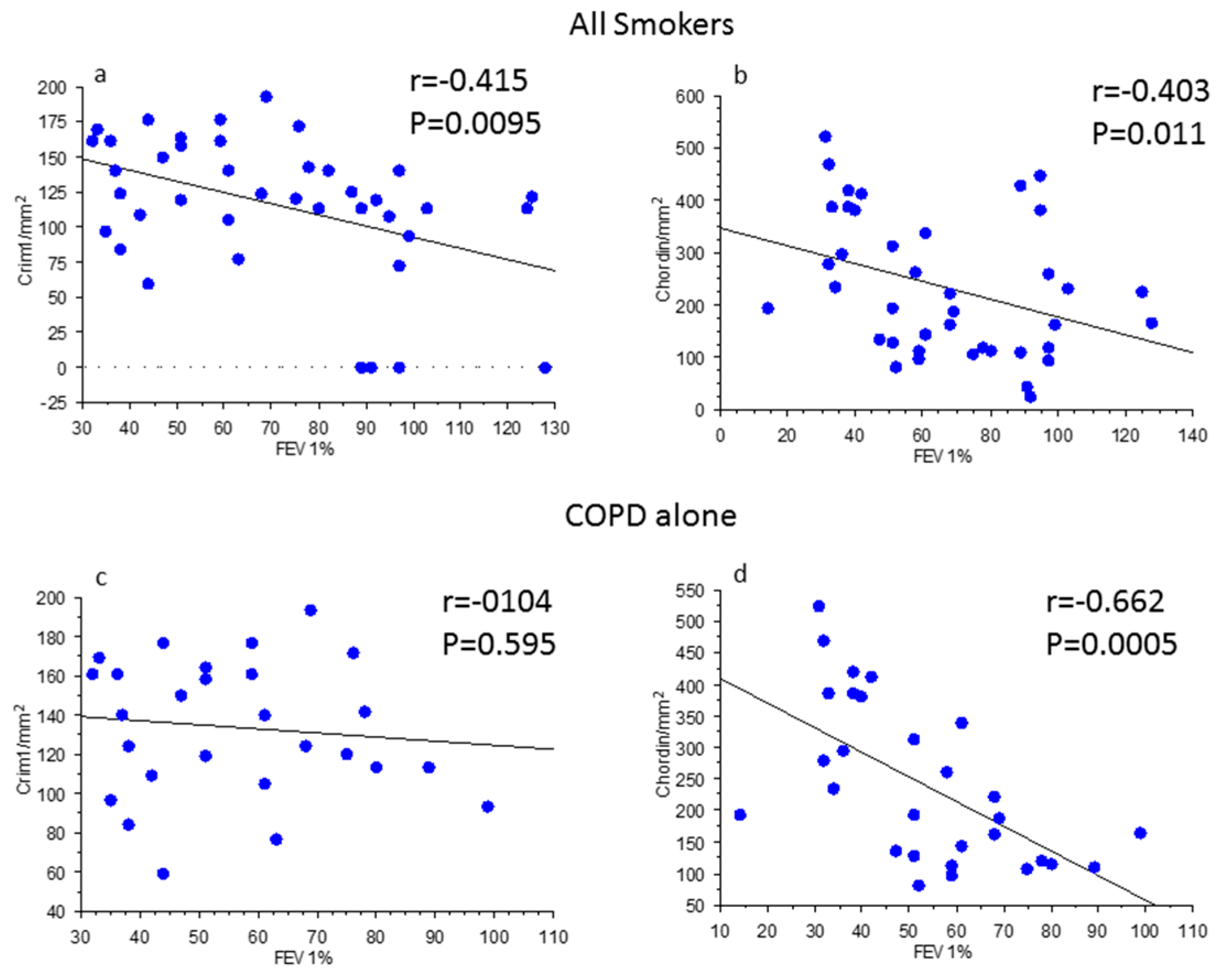
3.4. Correlations between Clinical Parameters, BMPs, and BMP Antagonists in Bronchial Biopsies
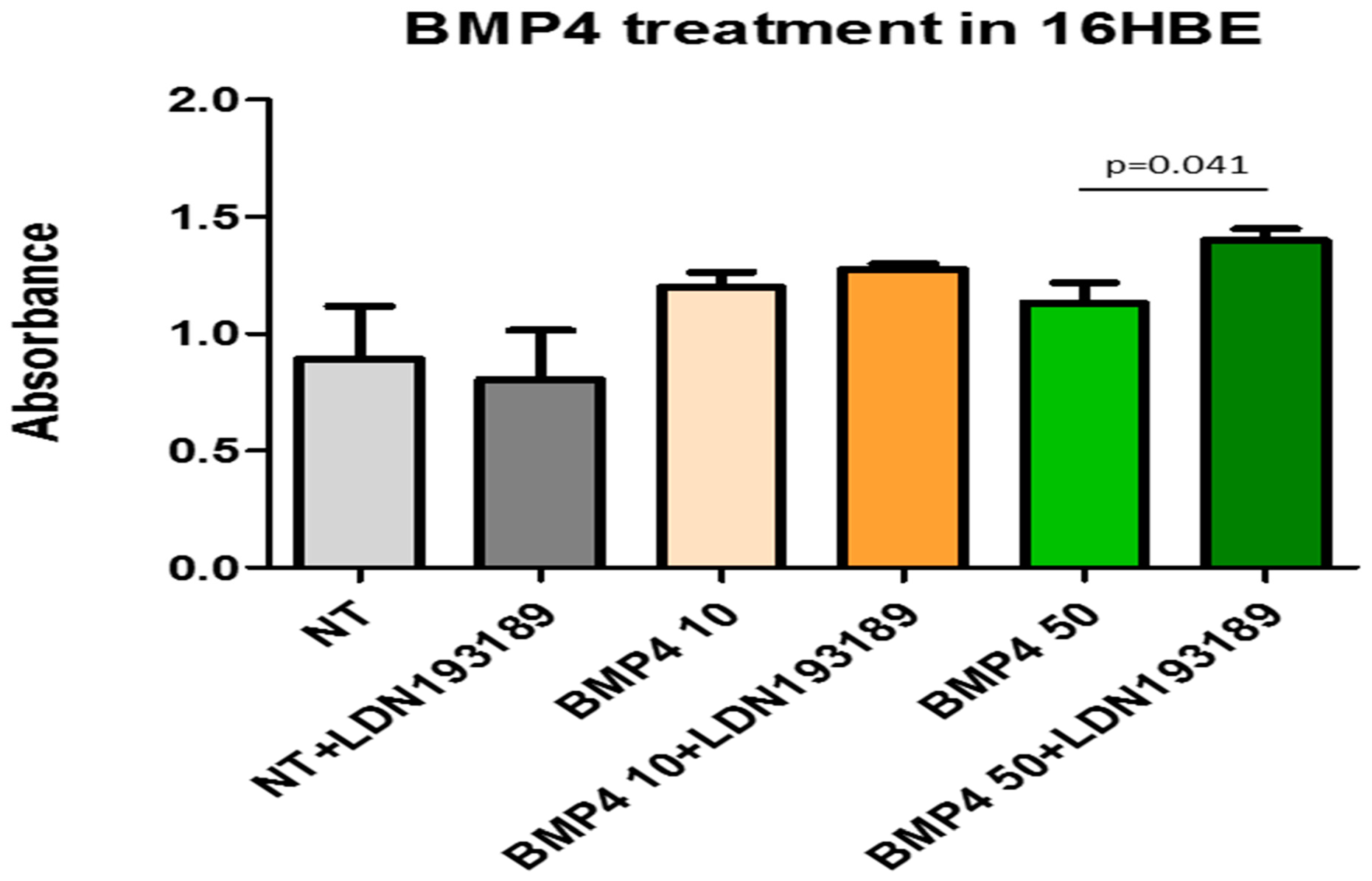
3.5. ELISA Tests for BMP4 and Chordin in the Supernatants of Pro-Inflammatory Protein Treated and Non-Treated 16HBE Cells
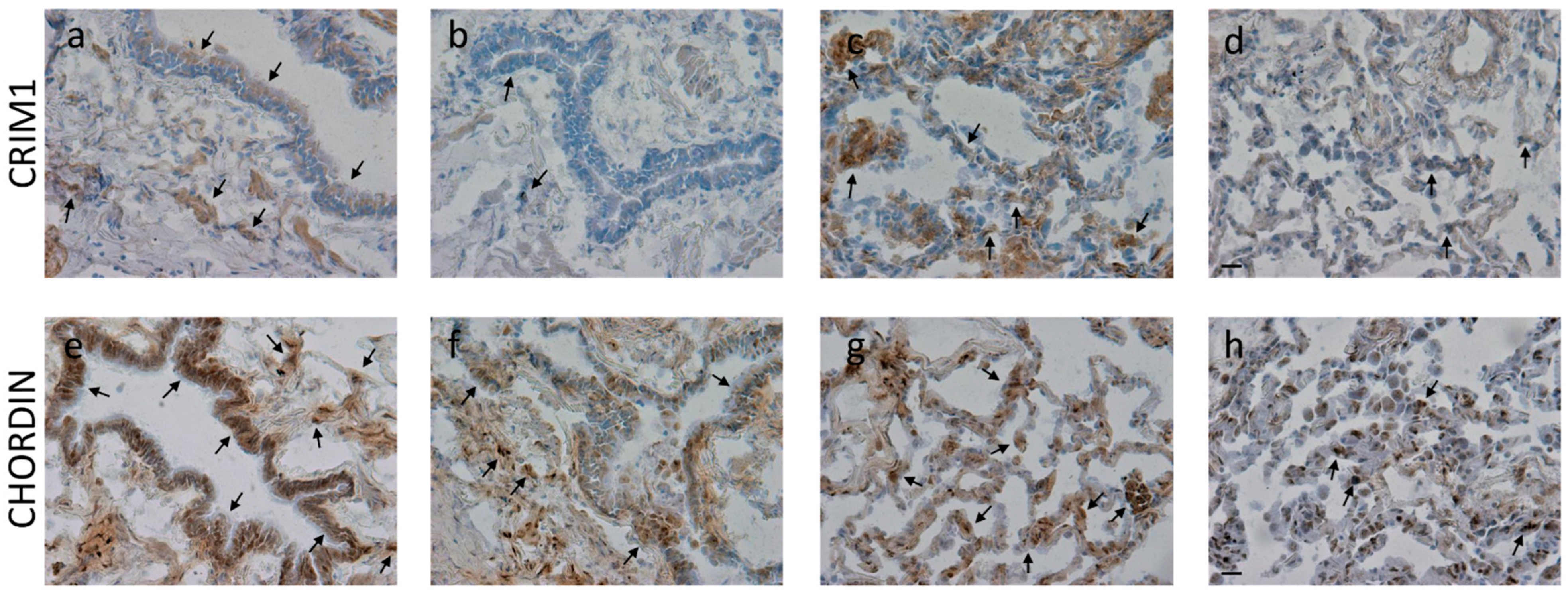
3.6. Immunohistochemistry for BMPs and BMP Antagonists in the Peripheral Airways and Lung Parenchyma
3.7. Gene Expression Level in Bronchial Rings and Lung Parenchyma
3.8. BrdU Test for Quantification of Bronchial Epithelial Cells Proliferation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rider, C.C.; Mulloy, B. Bone morphogenetic protein and growth differentiation factor cytokine families and their protein antagonists. Biochem. J. 2010, 429, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Brazil, D.P.; Church, R.H.; Surae, S.; Godson, C.; Martin, F. BMP signalling: Agony and antagony in the family. Trends Cell Biol. 2015, 25, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Tadokoro, T.; Gao, X.; Hong, C.C.; Hotten, D.; Hogan, B.L. BMP signaling and cellular dynamics during regeneration of airway epithelium from basal progenitors. Development 2016, 143, 764–773. [Google Scholar] [CrossRef]
- Cibois, M.; Luxardi, G.; Chevalier, B.; Thomé, V.; Mercey, O.; Zaragosi, L.E.; Barbry, P.; Pasini, A.; Marcet, B.; Kodjabachian, L. BMP signalling controls the construction of vertebrate mucociliary epithelia. Development 2015, 142, 2352–2363. [Google Scholar] [CrossRef] [PubMed]
- Zuo, W.L.; Yang, J.; Strulovici-Barel, Y.; Salit, J.; Rostami, M.; Mezey, J.G.; O’Beirne, S.L.; Kaner, R.J.; Crystal, R.G. Exaggerated BMP4 signalling alters human airway basal progenitor cell differentiation to cigarette smoking-related phenotypes. Eur. Respir. J. 2019, 53, 1702553. [Google Scholar] [CrossRef]
- Wang, D.; Prakash, J.; Nguyen, P.; Davis-Dusenbery, B.N.; Hill, N.S.; Layne, M.D.; Hata, A.; Lagna, G.J. Bone morphogenetic protein signaling in vascular disease: Anti-inflammatory action through myocardin-related transcription factor A. Biol. Chem. 2012, 287, 28067–28077. [Google Scholar] [CrossRef]
- Li, Z.; Wang, J.; Wang, Y.; Jiang, H.; Xu, X.; Zhang, C.; Li, D.; Xu, C.; Zhang, K.; Qi, Y.; et al. Bone morphogenetic protein 4 inhibits liposaccharide-induced inflammation in the airway. Eur. J. Immunol. 2014, 44, 3283–3294. [Google Scholar] [CrossRef]
- Jasuja, R.; Allen, B.L.; Pappano, W.N.; Rapraeger, A.C.; Greenspan, D.S. Cell-surface heparan sulfate proteoglycans potentiate chordin antagonism of bone morphogenetic protein signaling and are necessary for cellular uptake of chordin. J. Biol. Chem. 2004, 279, 51289–51297. [Google Scholar] [CrossRef]
- Nakayama, N.; Han, C.E.; Scully, S.; Nishinakamura, R.; He, C.; Zeni, L.; Yamane, H.; Chang, D.; Yu, D.; Yokota, T.; et al. A novel chordin-like protein inhibitor for bone morphogenetic proteins expressed preferentially in mesenchymal cell lineages. Dev. Biol. 2001, 232, 372–387. [Google Scholar] [CrossRef]
- Cyr-Depauw, C.; Northey, J.J.; Tabariès, S.; Annis, M.G.; Dong, Z.; Cory, S.; Hallett, M.; Rennhack, J.P.; Andrechek, E.R.; Siegel, P.M. Chordin-Like 1 Suppresses Bone Morphogenetic Protein 4-Induced Breast Cancer Cell Migration and Invasion. Mol. Cell. Biol. 2016, 36, 1509–1525. [Google Scholar] [CrossRef]
- Kane, R.; Godson, C.; O’Brien, C. Chordin-like 1, a bone morphogenetic protein-4 antagonist, is upregulated by hypoxia in human retinal pericytes and plays a role in regulating angiogenesis. Mol. Vis. 2008, 14, 1138–1148. [Google Scholar] [PubMed]
- Sakai, H.; Horiguchi, M.; Ozawa, C.; Akita, T.; Hirota, K.; Shudo, K.; Terada, H.; Makino, K.; Kubo, H.; Yamashita, C. Pulmonary administration of Am80 regenerates collapsed alveoli. J. Control. Release 2014, 196, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, L.; Kolle, G.; Wen, D.; Piper, M.; Scott, J.; Little, M. CRIM1 regulates the rate of processing and delivery of bone morphogenetic proteins to the cell surface. J. Biol. Chem. 2003, 278, 34181–34188. [Google Scholar] [CrossRef]
- Iyer, S.; Chhabra, Y.; Harvey, T.J.; Wang, R.; Chiu, H.S.; Smith, A.G.; Thomas, W.G.; Pennisi, D.J.; Piper, M. CRIM1 is necessary for coronary vascular endothelial cell development and homeostasis. J. Mol. Histol. 2017, 48, 53–61. [Google Scholar] [CrossRef]
- Zeng, H.; Zhang, Y.; Yi, Q.; Wu, Y.; Wan, R.; Tang, L. CRIM1, a newfound cancer-related player, regulates the adhesion and migration of lung cancer cells. Growth Factors 2015, 33, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Prosdocimo, D.A.; Jain, M.K. BMPing up angiogenesis via BMPER. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2167–2168. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Heinke, J.; Juschkat, M.; Charlet, A.; Mnich, L.; Helbing, T.; Bode, C.; Patterson, C.; Moser, M. Antagonism and synergy between extracellular BMP modulators Tsg and BMPER balance blood vessel formation. J. Cell Sci. 2013, 126, 3082–3094. [Google Scholar] [CrossRef]
- Helbing, T.; Rothweiler, R.; Heinke, J.; Goetz, L.; Diehl, P.; Zirlik, A.; Patterson, C.; Bode, C.; Moser, M. BMPER is upregulated by statins and modulates endothelial inflammation by intercellular adhesion molecule-1. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 554–560. [Google Scholar] [CrossRef]
- Ankeny, R.F.; Thourani, V.H.; Weiss, D.; Vega, J.D.; Taylor, W.R.; Nerem, R.M.; Jo, H. Preferential activation of SMAD1/5/8 on the fibrosa endothelium in calcified human aortic valves--association with low BMP antagonists and SMAD6. PLoS ONE 2011, 6, e20969. [Google Scholar] [CrossRef]
- Helbing, T.; Wiltgen, G.; Hornstein, A.; Brauers, E.Z.; Arnold, L.; Bauer, A.; Esser, J.S.; Diehl, P.; Grundmann, S.; Fink, K.; et al. Bone Morphogenetic Protein-Modulator BMPER Regulates Endothelial Barrier Function. Inflammation 2017, 40, 442–453. [Google Scholar] [CrossRef]
- Helbing, T.; Herold, E.M.; Hornstein, A.; Wintrich, S.; Heinke, J.; Grundmann, S.; Patterson, C.; Bode, C.; Moser, M. Inhibition of BMP activity protects epithelial barrier function in lung injury. J. Pathol. 2013, 231, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, L.B.; De Jesús-Escobar, J.M.; Harland, R.M. The Spemann organizer signal noggin binds and inactivates bone morphogenetic protein 4. Cell 1996, 86, 599–606. [Google Scholar] [CrossRef]
- Yang, K.; Lu, W.; Jia, J.; Zhang, J.; Zhao, M.; Wang, S.; Jiang, H.; Xu, L.; Wang, J. Noggin inhibits hypoxia-induced proliferation by targeting store-operated calcium entry and transient receptor potential cation channels. Am. J. Physiol. Cell Physiol. 2015, 308, C869–C878. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, A.; Caramori, G.; Barczyk, A.; Vicari, C.; Brun, P.; Zanini, A.; Cappello, F.; Garofano, E.; Padovani, A.; Contoli, M.; et al. Innate immunity but not NLRP3 inflammasome activation correlates with severity of stable COPD. Thorax 2014, 69, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, A.; Sangiorgi, C.; Gnemmi, I.; Casolari, P.; Brun, P.; Ricciardolo, F.L.M.; Contoli, M.; Papi, A.; Maniscalco, P.; Ruggeri, P.; et al. TGF-β Signaling Pathways in Different Compartments of the Lower Airways of Patients With Stable COPD. Chest 2018, 153, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Cozens, A.L.; Yezzi, M.J.; Kunzelmann, K.; Ohrui, T.; Chin, L.; Eng, K.; Finkbeiner, W.E.; Widdicombe, J.H.; Gruenert, D.C. CFTR expression and chloride secretion in polarized immortal human bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 1994, 10, 38–47. [Google Scholar] [CrossRef]
- McCormack, N.; Molloy, E.L.; O’Dea, S. Bone morphogenetic proteins enhance an epithelial-mesenchymal transition in normal airway epithelial cells during restitution of a disrupted epithelium. Respir. Res. 2013, 14, 36. [Google Scholar] [CrossRef]
- McCormack, N.; O’Dea, S. Regulation of epithelial to mesenchymal transition by bone morphogenetic proteins. Cell Signal. 2013, 25, 2856–2862. [Google Scholar] [CrossRef]
- Di Stefano, A.; Caramori, G.; Gnemmi, I.; Contoli, M.; Bristot, L.; Capelli, A.; Ricciardolo, F.L.; Magno, F.; D’Anna, S.E.; Zanini, A.; et al. Association of increased CCL5 and CXCL7 chemokine expression with neutrophil activation in severe stable COPD. Thorax 2009, 64, 968–975. [Google Scholar] [CrossRef]
- Groppe, J.; Greenwald, J.; Wiater, E.; Rodriguez-Leon, J.; Economides, A.N.; Kwiatkowski, W.; Affolter, M.; Vale, W.W.; Izpisua Belmonte, J.C.; Choe, S. Structural basis of BMP signalling inhibition by the cystine knot protein Noggin. Nature 2002, 420, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Kelley, R.; Ren, R.; Pi, X.; Wu, Y.; Moreno, I.; Willis, M.; Moser, M.; Ross, M.; Podkowa, M.; Attisano, L.; et al. A concentration-dependent endocytic trap and sink mechanism converts Bmper from an activator to an inhibitor of Bmp signaling. J. Cell Biol. 2009, 184, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Heinke, J.; Wehofsits, L.; Zhou, Q.; Zoeller, C.; Baar, K.M.; Helbing, T.; Laib, A.; Augustin, H.; Bode, C.; Patterson, C.; et al. BMPER is an endothelial cell regulator and controls bone morphogenetic protein-4-dependent angiogenesis. Circ. Res. 2008, 103, 804–812. [Google Scholar] [CrossRef] [PubMed]
- Glienke, J.; Sturz, A.; Menrad, A.; Thierauch, K.H. CRIM1 is involved in endothelial cell capillary formation in vitro and is expressed in blood vessels in vivo. Mech. Dev. 2002, 119, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Ruozi, G.; Bortolotti, F.; Mura, A.; Tomczyk, M.; Falcione, A.; Martinelli, V.; Vodret, S.; Braga, L.; Dal Ferro, M.; Cannatà, A.; et al. Cardioprotective factors against myocardial infarction selected in vivo from an AAV secretome library. Sci. Transl. Med. 2022, 14, eabo0699. [Google Scholar] [CrossRef]
- Staudt, M.R.; Buro-Auriemma, L.J.; Walters, M.S.; Salit, J.; Vincent, T.; Shaykhiev, R.; Mezey, J.G.; Tilley, A.E.; Kaner, R.J.; Ho, M.W.; et al. Airway Basal stem/progenitor cells have diminished capacity to regenerate airway epithelium in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2014, 190, 955–958. [Google Scholar] [CrossRef]
- Galbán, C.J.; Han, M.K.; Boes, J.L.; Chughtai, K.A.; Meyer, C.R.; Johnson, T.D.; Galbán, S.; Rehemtulla, A.; Kazerooni, E.A.; Martinez, F.J.; et al. Computed tomography-based biomarker provides unique signature for diagnosis of COPD phenotypes and disease progression. Nat. Med. 2012, 18, 1711–1715. [Google Scholar] [CrossRef]
- Hogg, J.C.; McDonough, J.E.; Suzuki, M. Small airway obstruction in COPD: New insights based on micro-CT imaging and MRI imaging. Chest 2013, 143, 1436–1443. [Google Scholar] [CrossRef]
- Hogg, J.C.; McDonough, J.E.; Sanchez, P.G.; Cooper, J.D.; Coxson, H.O.; Elliott, W.M.; Naiman, D.; Pochettino, M.; Horng, D.; Gefter, W.B.; et al. Micro-computed tomography measurements of peripheral lung pathology in chronic obstructive pulmonary disease. Proc. Am. Thorac. Soc. 2009, 6, 546–549. [Google Scholar] [CrossRef]
- Vasilescu, D.M.; Martinez, F.J.; Marchetti, N.; Galbán, C.J.; Hatt, C.; Meldrum, C.A.; Dass, C.; Tanabe, N.; Reddy, R.M.; Lagstein, A.; et al. Noninvasive Imaging Biomarker Identifies Small Airway Damage in Severe Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2019, 200, 575–581. [Google Scholar] [CrossRef]
- Vasilescu, D.M.; Hackett, T.L.; Martinez, F.J.; Curtis, J.L.; Hogg, J.C.; Han, M.K. Reply to Janssen and Wouters: Loss of Alveolar Attachments as a Pathomechanistic Link between Small Airway Disease and Emphysema. Am. J. Respir. Crit. Care Med. 2020, 201, 879–880. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | N | Age (Years) | M/F | Pack Years | Ex/Current Smokers | FEV1 (% Pred) Pre-β2 | FEV1 (% Pred) Post-β2 | FEV1/FVC (%) |
|---|---|---|---|---|---|---|---|---|
| Control non-smokers | 11 | 64 ± 11 | 10/1 | 0 | 0 | 111 ± 16 | ND | 84 ± 9 |
| Control smokers with normal lung function | 13 | 60 ± 8 | 9/4 | 35 ± 25 | 2/11 | 101 ± 14 | ND | 81 ± 5 |
| COPD grades I and II (mild/moderate) | 18 | 68 ± 9 | 15/3 | 46 ± 22 | 6/12 | 66 ± 13 # | 70 ± 12 | 58 ± 9 # |
| COPD grades III and IV (severe/very severe) | 16 | 66 ± 12 | 11/5 | 58 ± 39 | 13/3 | 36 ± 7 #& | 41 ± 6 | 44 ± 9 #& |
| Groups | N | Age (Year) | M/F | Ex/Current Smokers | Pack Years | FEV1 (% Pred) Pre-β2 | FEV1 (% Pred) Post-β2 | FEV1/FVC (%) |
|---|---|---|---|---|---|---|---|---|
| Control non-smokers | 9 | 71 ± 3 | 5/4 | --- | --- | 115 ± 4.4 | ND | 81 ± 1.5 |
| Control smokers | 11 | 69 ± 1.9 | 7/4 | 7/4 | 39 ± 16 | 98 ± 2.3 | ND | 76 ± 1.6 |
| Patients with COPD | 9 | 70 ± 1.3 | 8/1 | 8/1 | 48 ± 26 | 69 ± 4.9 # | 77 ± 4.8 | 61 ± 2.9 # |
| Target | Supplier | Cat. #a | Source | Dilution | Positive Control |
|---|---|---|---|---|---|
| BMP1 | R&D | AF1927 | Goat | 1:100 (2 µg/mL) | Nasal polyp |
| BMP2 | Peprotech | 500-p195 | Rabbit | 1:300 (0.3 µg/mL) | Nasal polyp |
| BMP4 | Thermo Fisher | PA137175 | Rabbit | 1:150 (1.3 µg/mL) | Nasal polyp |
| BMP7 | R&D | MAB3541 | Mouse | 1:300 (13 µg/mL) | Nasal polyp |
| BMP9 | Abcam | Ab71809 | Rabbit | 1:80 (5 µg/mL) | Nasal polyp |
| BMP10 | Bio Antibodies | Bs-9447R | Rabbit | 1:150 (6.5 µg/mL) | Nasal polyp |
| BMPER | Abcam | Ab-73900 | Rabbit | 1:400 (1.2 µg/mL) | Nasal polyp |
| CRIM1 | Sigma | HPA000556 | Rabbit | 1:25 (4 µg/mL) | Nasal polyp |
| CHORDIN | Cloud Clone | PAC126Hu01 | Rabbit | 1:80 (2.5 µg/mL) | Nasal polyp |
| NOGGIN | Santa Cruz | Sc-293439 | Mouse | 1:50 (2 µg/mL) | Nasal polyp |
| Control Non-Smokers | Control Smokers | Mild/Moderate COPD | Severe/Very Severe COPD | Kruskal–Wallis p-Value | |
|---|---|---|---|---|---|
| Epithelium (Score 0–3) | |||||
| BMP1 | 0 (0–1) | 0 (0–0.12) | 0 (0–1.25) | 0 (0–0.5) | 0.957 |
| BMP2 | 0 (0–0) | 0 (0–0) | 0 (0–0.12) | 0 (0–0.12) | 0.987 |
| BMP4 | 0.75 (0.25–1.25) | 0.5 (0.25–1.0) * | 0.5 (0.25–0.75) * | 0.37 (0.25–0.75) * | 0.010 |
| BMP7 | 0.25 (0.25–0.75) | 0.25 (0–1.0) | 0.5 (0.25–0.75) | 0.5 (0.25–1.25) | 0.264 |
| BMP9 | 0.5 (0–1) | 0.5 (0–1.25) | 0.25 (0–1) | 0.75 (0.25–1) | 0.561 |
| BMP10 | 0.5 (0–1.5) | 0.75 (0–1) | 0.62 (0.25–1.5) | 0.75 (0.25–1.25) | 0.745 |
| BMPER | 0 (0–0.25) | 0.25 (0–0.75) * | 0.5 (0–0.75) * | 0.5 (0–1) * | 0.0009 |
| CRIM1 | 0 (0–1) | 0.5 (0–1) | 0.5 (0.25–1.5) *& | 0.5 (0.25–1) *$ | 0.0025 |
| CHORDIN | 1.25 (0.5–2.0) | 1.87 (0.5–2.75) | 1.62 (1.25–2.5) | 2 (1.5–3.0) *$ | 0.0110 |
| NOGGIN | 0 (0–0) | 0 (0–0) | 0 (0–0) | 0 (0–0) | n.d. |
| Lamina Propria (cells/mm2) | |||||
| BMP1 | 6 (0–48) | 4.5 (0–55) | 12 (0–122) | 9 (0–125) | 0.765 |
| BMP2 | 0 (0–0) | 0 (0–0) | 0 (0–12) | 0 (0–64) | 0.824 |
| BMP4 | 76 (6–138) | 48 (21–104) | 69 (51–148) | 84 (52–136) | 0.054 |
| BMP7 | 43 (5–142) | 29 (9–116) | 55 (24–123) | 60 (20–97) | 0.160 |
| BMP9 | 48 (0–156) | 27 (0–149) | 14 (0–125) | 55 (24–102) | 0.109 |
| BMP10 | 26 (0–122) | 37 (6–90) | 22 (0–106) | 20 (0–52) | 0.524 |
| BMPER | 89 (16–177) | 150 (11–203) | 150 (64–212) | 135 (56–226) | 0.062 |
| CRIM1 | 5 (0–156) | 113 (0–140) | 124 (77–193) *& | 140 (59–177) *& | 0.0003 |
| CHORDIN | 175 (54–355) | 226 (24–447) | 145 (81–339) | 384 (135–523) *&$ | 0.0028 |
| NOGGIN | 0 (0–11) | 0 (0–8) | 0 (0–6) | 0 (0–6) | 0.805 |
| Localization | Control Non-Smokers | Control Smokers | COPD Patients | Kruskal–Wallis (p-Value) |
|---|---|---|---|---|
| Bronchiolar Epithelium (Score 0–3) | ||||
| BMP4 | 1.70 (0.5–2.5) | 1.75 (1.4–2.5) | 2.06 (0.5–2.5) | 0.811 |
| Chordin | 1.75 (0.75–2.5) | 1.50 (0.5–2.5) | 1.75 (0.87–2.0) | 0.495 |
| CRIM1 | 0.62 (0.12–1.5) * | 0.25 (0.12–1.0) | 0.50 (0.12–1.5) | 0.069 |
| BMPER | 0.25 (0–0.50) | 0.25 (0.12–0.50) | 0.50 (0.12–0.75) | 0.593 |
| Bronchiolar Lamina propria (Score 0–3) | ||||
| BMP4 | 0.50 (0.25–0.50) | 0.50 (0.25–0.50) | 0.50 (0–0.50) | 0.931 |
| Chordin | 1.12 (0.25–1.5) * | 0.50 (0.25–1.5) | 0.75 (0.5–1.5) | 0.054 |
| CRIM1 | 0.62 (0.12–1.5) * | 0.25 (0–0.75) | 0.37 (0–1.5) | 0.030 |
| BMPER | 0.50 (0.12–1.0) | 0.50 (0.12–0.75) | 0.75 (0.12–1.0) | 0.371 |
| Alveolar Macrophages (Score 0–3) | ||||
| BMP4 | 1.25 (1–2.5) | 2.0 (1–2.5) | 1.50 (1–2.0) | 0.172 |
| Chordin | 2.0 (1–2.5) * | 1.50 (1–2.0) | 1.12 (0.5–2.5) & | 0.032 |
| CRIM1 | 0.87 (0.75–2.5) | 0.75 (0.5–1.5) | 0.87 (0.25–1.5) | 0.720 |
| BMPER | 0.75 (0.25–1.5) | 0.75 (0.25–1.25) | 0.5 (0.25–1.5) | 0.128 |
| Alveolar Septa (Score 0–3) | ||||
| BMP4 | 0.25 (0.25–0.5) | 0.37 (0.25–0.5) | 0.5 (0.25–1.0) | 0.128 |
| Chordin | 1.50 (0.75–2.25) | 1.0 (0.5–2.0) | 0.87 (0.5–1.5) & | 0.087 |
| CRIM1 | 1.37 (0.25–2.0) * | 0.50 (0–1.0) | 0.50 (0–2.0) | 0.010 |
| BMPER | 0.75 (0.25–1.0) | 0.50 (0.12–0.75) | 0.25 (0.12–1.0) | 0.287 |
| Lung vessels (Score 0–3) | ||||
| BMP4 | 0.62 (0.25–1.5) | 0.87 (0.25–1.0) | 0.75 (0–1.5) | 0.978 |
| Chordin | 1.37 (0.5–2.0) | 0.75 (0–2.0) | 0.50 (0–1.5) | 0.318 |
| CRIM1 | 1.50 (0.75–2.5) | 1.5 (0.25–1.5) | 1.25 (0–2.5) | 0.426 |
| BMPER | 0.75 (0.25–1.0) | 0.37 (0–1.0) | 0.5 (0.25–1.0) | 0.561 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Stefano, A.; Rosani, U.; Levra, S.; Gnemmi, I.; Brun, P.; Maniscalco, M.; D’Anna, S.E.; Carriero, V.; Bertolini, F.; Ricciardolo, F.L.M. Bone Morphogenic Proteins and Their Antagonists in the Lower Airways of Stable COPD Patients. Biology 2023, 12, 1304. https://doi.org/10.3390/biology12101304
Di Stefano A, Rosani U, Levra S, Gnemmi I, Brun P, Maniscalco M, D’Anna SE, Carriero V, Bertolini F, Ricciardolo FLM. Bone Morphogenic Proteins and Their Antagonists in the Lower Airways of Stable COPD Patients. Biology. 2023; 12(10):1304. https://doi.org/10.3390/biology12101304
Chicago/Turabian StyleDi Stefano, Antonino, Umberto Rosani, Stefano Levra, Isabella Gnemmi, Paola Brun, Mauro Maniscalco, Silvestro Ennio D’Anna, Vitina Carriero, Francesca Bertolini, and Fabio L. M. Ricciardolo. 2023. "Bone Morphogenic Proteins and Their Antagonists in the Lower Airways of Stable COPD Patients" Biology 12, no. 10: 1304. https://doi.org/10.3390/biology12101304
APA StyleDi Stefano, A., Rosani, U., Levra, S., Gnemmi, I., Brun, P., Maniscalco, M., D’Anna, S. E., Carriero, V., Bertolini, F., & Ricciardolo, F. L. M. (2023). Bone Morphogenic Proteins and Their Antagonists in the Lower Airways of Stable COPD Patients. Biology, 12(10), 1304. https://doi.org/10.3390/biology12101304