

Crystallographic Texture of the Mineral Matter in the Bivalve Shells of Gryphaea dilatata Sowerby, 1816


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
- Determination of the crystallographic texture sharpness of the G. dilatata mineral substance;
- Assessment of the influence of various habitat conditions and diagenetic processes on the texture;
- Reveal the crystallographic texture dependence on the valve shape;
- Reveal the texture similarity in two representatives of the Gryphaeidae family;
- A comparison of recent and fossil valves of O. edulis;
- Reveal the common features of the crystallographic texture in thick-walled shells.
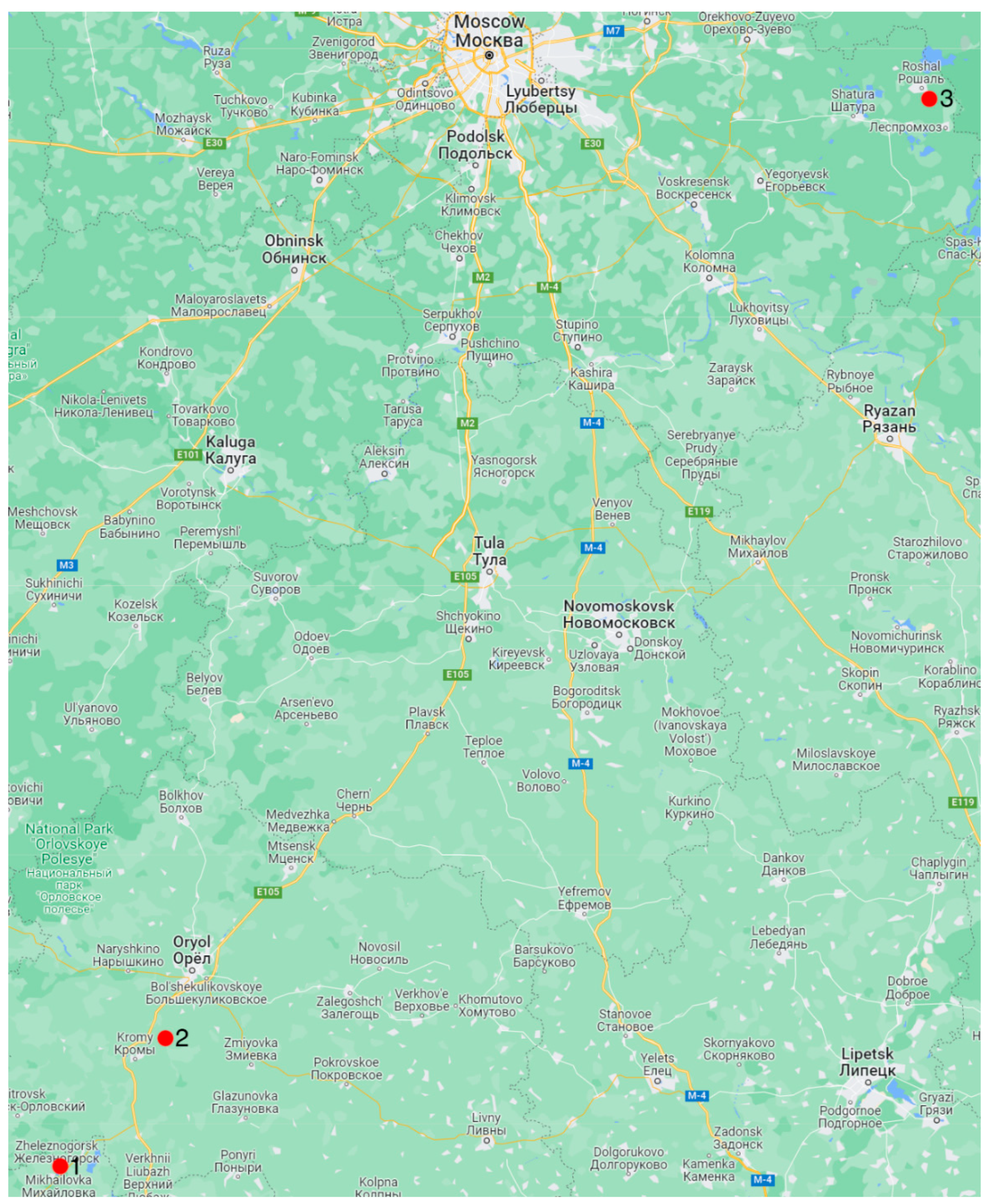
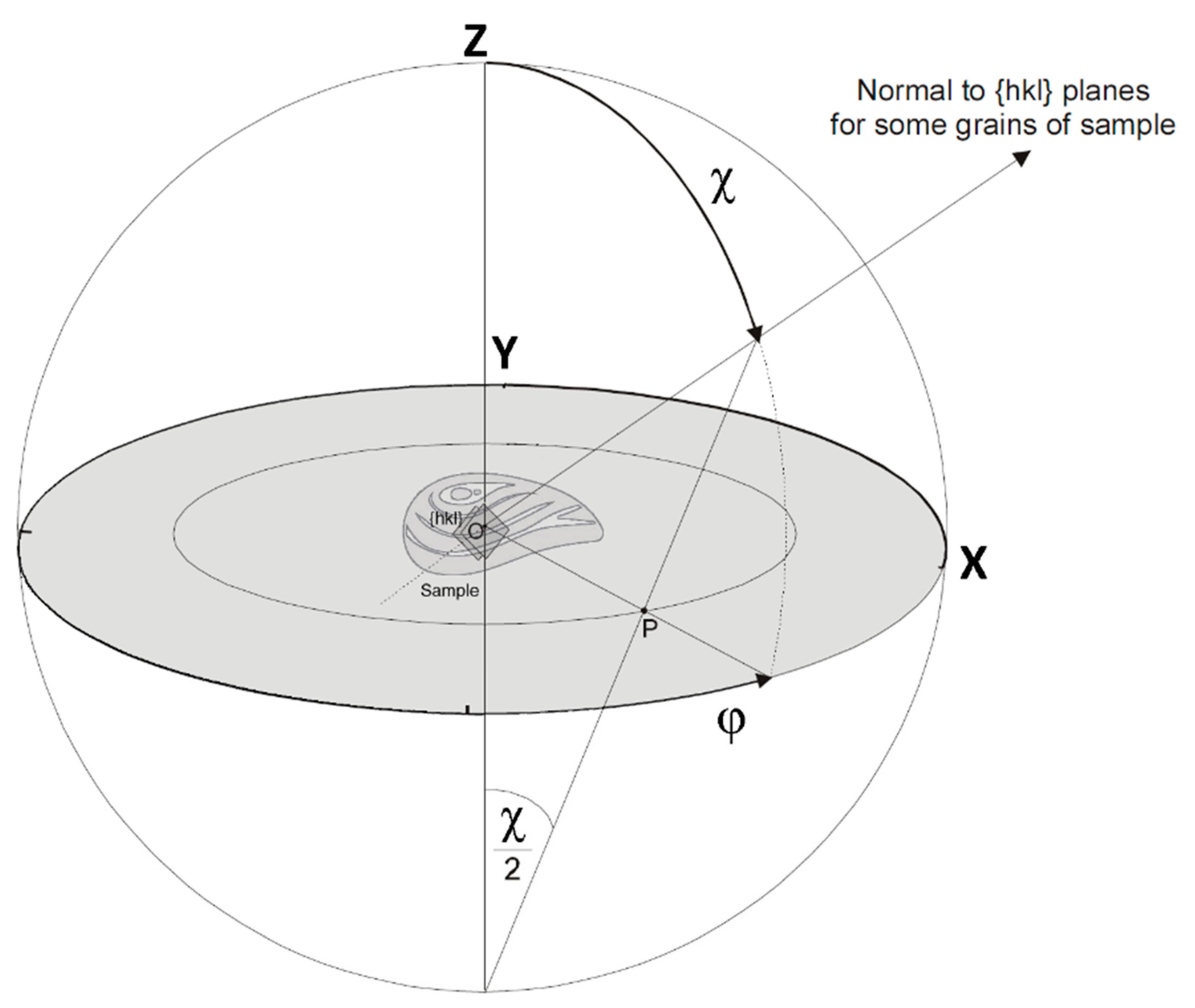
2. Materials and Methods
3. Results
3.1. Determination of the Shells’ Geologic Age
3.2. Analysis of the Diagenesis Features
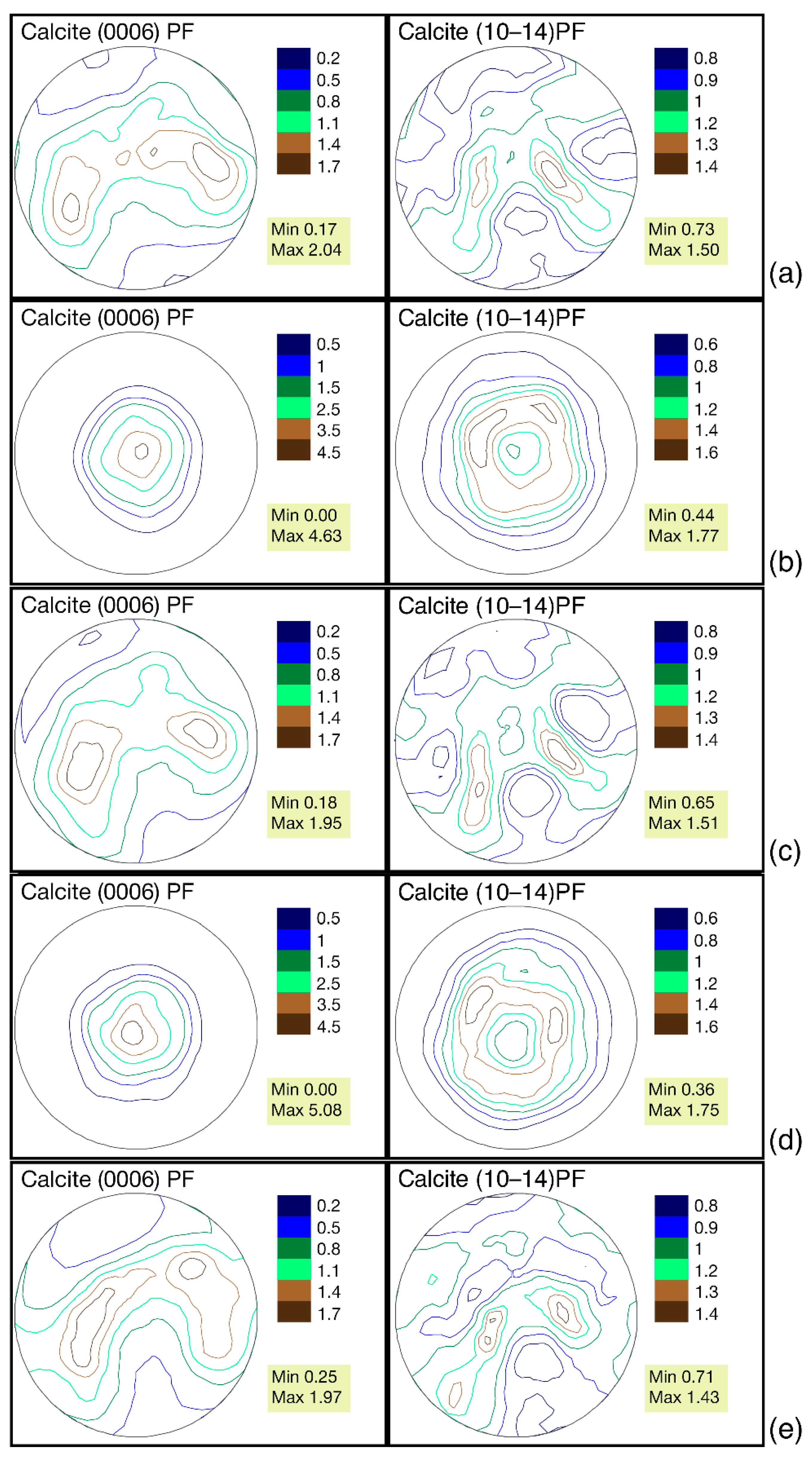
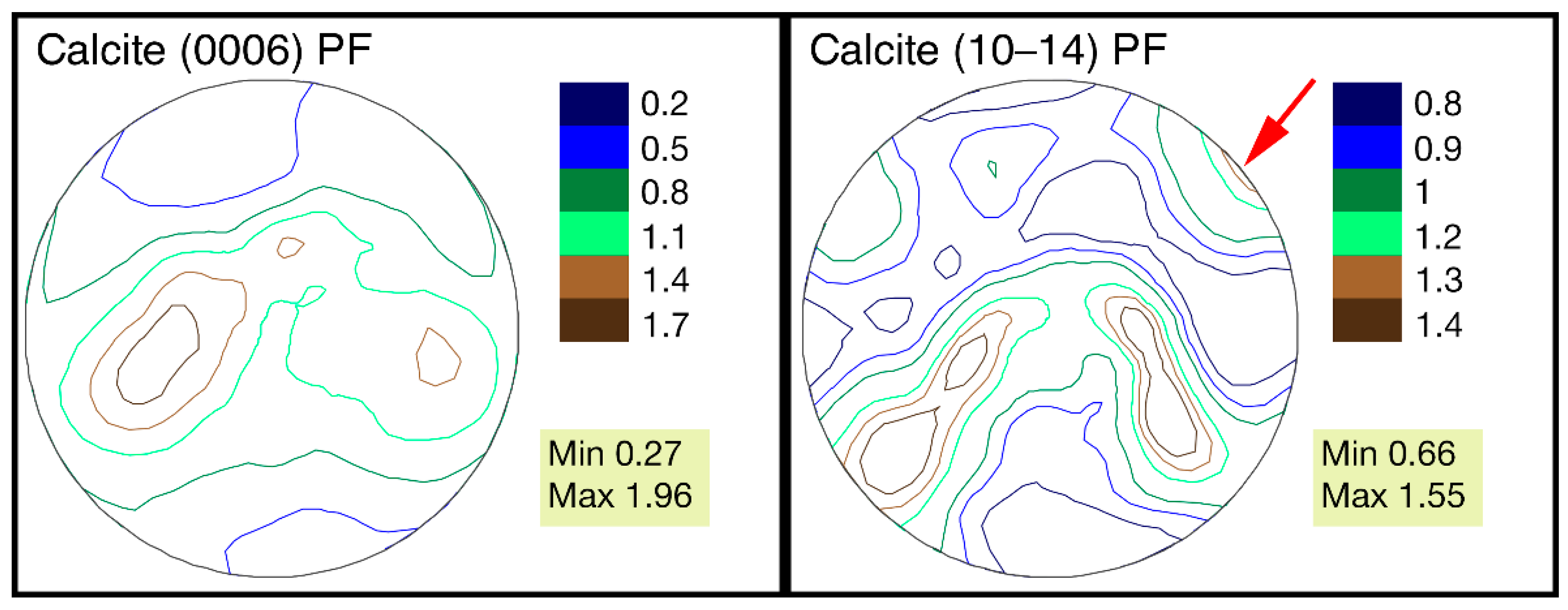
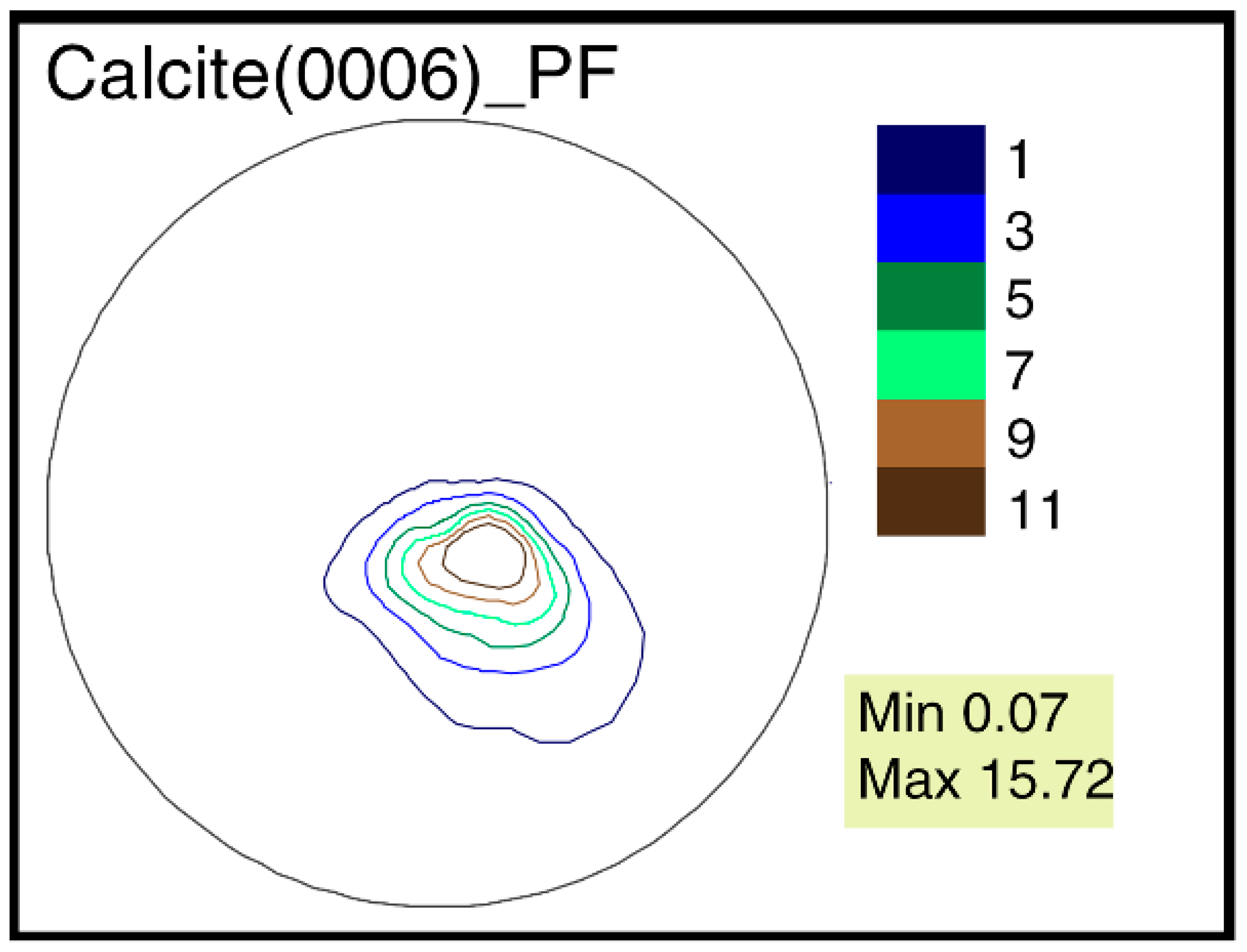
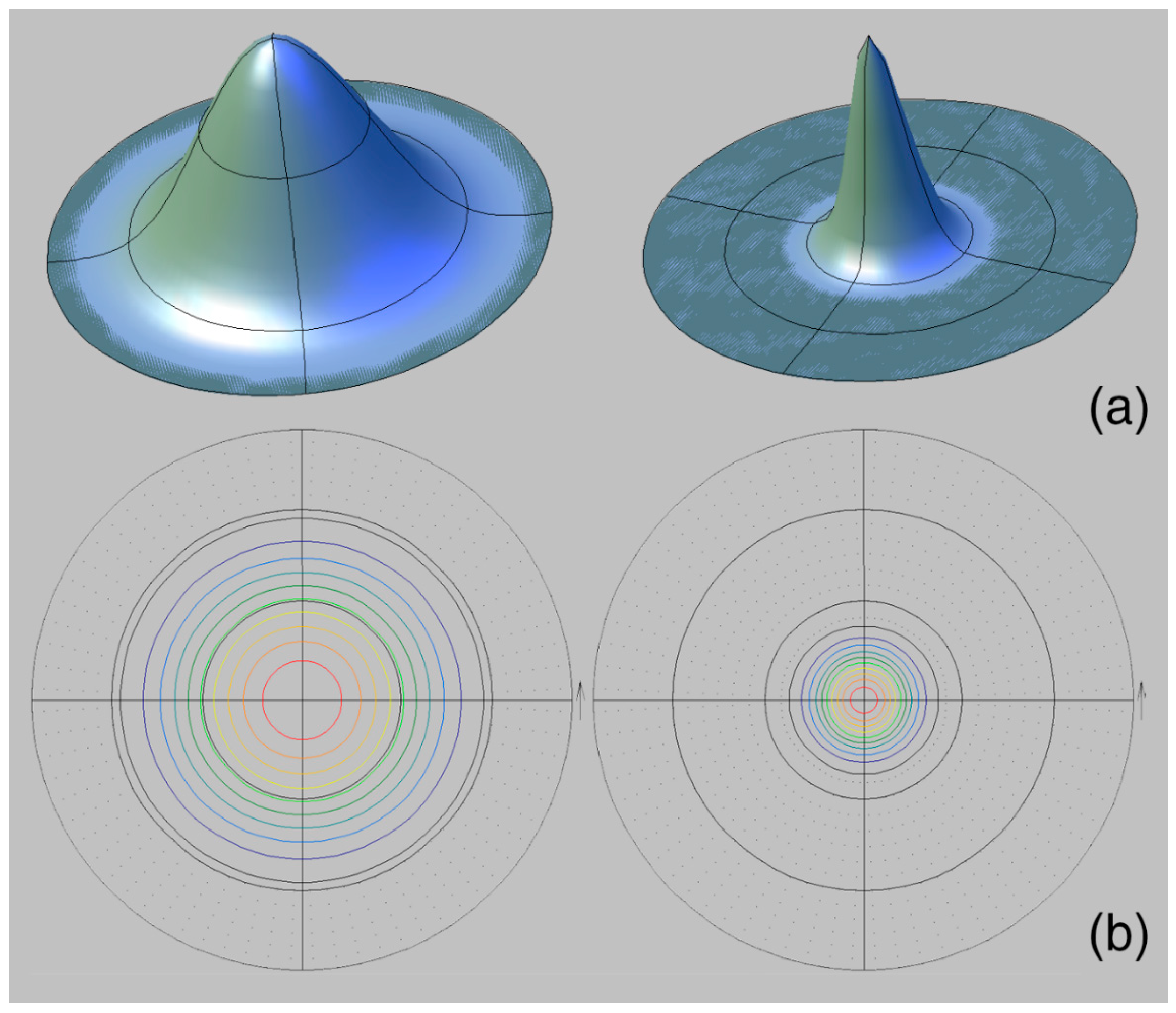
3.3. Features of Crystallographic Texture
4. Discussion
5. Conclusions
- For the first time, the complete pole figures of G. dilatata shells were measured using time-of-flight neutron diffraction.
- The studied G. dilatata lived at considerable distances from each other. The mollusks likely did not coincide in their time of habitation. This means that their habitats were different.
- G. dilatata from the three localities almost do not differ in their crystallographic texture maximum sharpness, i.e., the habitat conditions and fossilization did not affect the texture, and it can be considered as a very stable feature.
- The largest differences of the crystallographic texture sharpness have been noted for the calcite pole figure (0006). Therefore, it is possible to reveal the greatest variability by comparing these figures’ sharpness for different objects.
- The features of G. dilatata valve recrystallization, which affect the crystallographic texture, were revealed for the first time using time-of-flight neutron diffraction. They were reflected in the pole figure isolines, but do not appear in the values of maximum sharpness, since the entire valve was not recrystallized and the pole density was low in the recrystallized part. The maximum texture sharpness was determined by the non-recrystallized part of the valve.
- It has been established that for one mollusk, the crystallographic texture of the left and right valves of various shapes differs: to a greater extent by the pole figures’ isoline patterns, to a lesser extent by the values of maximum sharpness.
- The calcite pole figures and the values of the maximum sharpness of the G. dilatata and O. edulis left valves are similar. The pole figures isoline patterns and values of the pole density maximum of G. dilatata right valves are similar, and along the same parameters as the P. mirabilis left valve. All of these species are characterized by either an axial pole figure (or close to it) or a figure with an arcuately-curved sharpness maximum. Transitions between them are found for the recent O. edulis. It is possible that these pole figures with have isoline patterns that are characteristic of thick-walled shells.
- The thick-walled valves of G. dilatata, O. edulis, and P. mirabilis have small values of maximum texture sharpness. They are very close for these mollusks. Perhaps, it is a characteristic feature of all thick-walled valves and reflects an adaptation to shell construction.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nevesskaya, L.A.; Popov, S.V.; Goncharova, I.A.; Guzhov, A.V.; Yanin, B.T.; Polubotko, I.V.; Byakov, A.S.; Gavrilova, V.A. Bivalves of Russia and Neighboring Countries in the Phanerozoic; Scientific World: Moscow, Russia, 2013; 524p. [Google Scholar]
- Kennedy, W.J.; Taylor, J.D.; Hall, A. Environmental and Biological Controls on Bivalve Shell Mineralogy. Biol. Rev. 1969, 44, 499–530. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.G.; Clark, G.R. Classification and phylogenetic significance of molluscan shell microstructure. Ser. Geol. Notes Short Cour. 1985, 13, 50–71. [Google Scholar] [CrossRef]
- Zakhera, M.; Kassab, A.; Chinzei, K. Hyotissocameleo, a new Cretaceous oyster subgenus and its shell microstructure, from Wadi Tarfa, Eastern Desert of Egypt. Paleontol. Res. 2001, 5, 77–86. [Google Scholar]
- Nikolayev, D.I.; Lychagina, T.A.; Pakhnevich, A.V. Experimental neutron pole figures of minerals composing bivalve mollusk shells. Nat. Appl. Sci. 2019, 1, 344. [Google Scholar] [CrossRef]
- Chateigner, D.; Hedegaard, C.; Wenk, H.-R. Mollusc shell microstructures and crystallographic textures. J. Struct. Geol. 2000, 22, 1723–1735. [Google Scholar] [CrossRef]
- Nekhoroshkov, P.; Zinicovscaia, I.; Nikolayev, D.; Lychagina, T.; Pakhnevich, A.; Yushin, N.; Bezuidenhout, J. Effect of the elemental content of shells of the bivalve mollusks (Mytilus galloprovincialis) from Saldanha Bay (South Africa) on their crystallographic texture. Biology 2021, 10, 1093. [Google Scholar] [CrossRef]
- Bunge, H.J. Texture Analysis in Material Science, Mathematical Methods; Butterworths Publ.: London, UK, 1982; 595p. [Google Scholar]
- Ouhenia, S.; Chateigner, D.; Belkhir, M.A.; Guilmeau, E. Microstructure and crystallographic texture of Charonia lampas lampas shell. J. Struct. Biol. 2008, 163, 175–184. [Google Scholar] [CrossRef]
- Chateigner, D.; Kaptein, R.; Dupont-Nivet, M. X-ray quantitative texture analysis on Helix aspersa aspera (Pulmonata) shells selected or not for increased weight. Amer. Malac. Bull. 2009, 27, 157–165. [Google Scholar] [CrossRef]
- Frýda, J.; Klicnarová, K.; Frýdová, B.; Mergl, M. Variability in the crystallographic texture of bivalve nacre. Bull. Geosci. 2010, 85, 645–662. [Google Scholar] [CrossRef][Green Version]
- Hahn, S.; Rodolfo-Metalpa, R.; Griesshaber, E.; Schmahl, W.W.; Buhl, D.; Hall-Spencer, J.M.; Baggini, C.; Fehr, K.T.; Immenhauser, A. Marine bivalve shell geochemistry and ultrastructure from recent low pH environments: Environmental effect versus experimental bias. Biogeosciences 2012, 9, 1897–1914. [Google Scholar] [CrossRef]
- Fitzer, S.C.; Phoenix, V.R.; Cusack, M.; Kamenos, N.A. Ocean acidification impacts mussel control on biomineralization. Sci. Rep. 2014, 4, 6218. [Google Scholar] [CrossRef] [PubMed]
- Almagro, I.; Drzymała, P.; Berent, K.; Saínz-Díaz, C.I.; Willinger, M.G.; Bonarsky, J.; Checa, A.G. New crystallo-graphic relationships in biogenic aragonite: The crossed-lamellar microstructures of mollusks. Cryst. Growth Des. 2016, 16, 2083–2093. [Google Scholar] [CrossRef]
- Kučeráková, M.; Rohlicek, J.; Vratislav, S.; Jarosova, M.; Kalvoda, L.; Lychagina, T.; Nikolayev, D.; Douda, K. Texture and element concentration of the freshwater shells from the Unionidae family collected in the Czech Republic by X-ray, neutron and electron diffraction. Crystals 2021, 11, 1483. [Google Scholar] [CrossRef]
- Pakhnevich, A.; Nikolayev, D.; Lychagina, T.; Balasoiu, M.; Ibram, O. Global Crystallographic Texture of Freshwater Bivalve Mollusks of the Unionidae Family from Eastern Europe Studied by Neutron Diffraction. Life 2022, 12, 730. [Google Scholar] [CrossRef] [PubMed]
- Brokmeier, H.-G. Neutron Diffraction Texture Analysis of Multi-Phase Systems. Textures Microstruct. 1989, 10, 325–346. [Google Scholar] [CrossRef]
- Pakhnevich, A.V.; Nikolayev, D.I.; Lychagina, T.A. Comparison of the Crystallographic Texture of the Recent, Fossil and Subfossil Shells of Bivalves. Paleontol. J. 2021, 55, 589–599. [Google Scholar] [CrossRef]
- Nikolayev, D.I.; Lychagina, T.A.; Nikishin, A.V.; Yudin, V.V. Study of error distribution in measured pole figures. Solid State Phenom. 2005, 105, 77–82. [Google Scholar] [CrossRef]
- Nikolayev, D.I.; Lychagina, T.A.; Nikishin, A.V.; Yudin, V.V. Investigation of measured pole figures errors. Mater. Sci. Forum 2005, 495–497, 307–312. [Google Scholar] [CrossRef]
- Nikolayev, D.I.; Ullemeyer, K. A note on preprocessing of diffraction pole density data. J. Appl. Cryst. 1994, 27, 517–520. [Google Scholar] [CrossRef]
- Gerasimov, P.A. Guide Fossils of the Mesozoic of the European Central Regions of the USSR. Part 1. Lamellar-Gill, Gastropods, Boat-Footed Mollusks and Brachiopods of the Jurassic Deposits; Gosgeoltekhizdat: Moscow, Russia, 1955; 379p. (In Russian) [Google Scholar]
- Gerasimov, P.A.; Mitta, V.V.; Kochanova, M.D.; Tesakova, E.M. The Callovian Fossils of Central Russia; VNIGNI–MosGorSUN: Moscow, Russia, 1996; 127p. (In Russian) [Google Scholar]
- Checa, A.G.; Esteban-Delgado, F.J.; Rodríguez-Navarro, A.B. Crystallographic structure of the foliated calcite of bivalves. J. Structur. Biol. 2007, 157, 393–402. [Google Scholar] [CrossRef]
- Checa, A.G.; Harper, E.M.; González-Segura, A. Structure and crystallography of foliated and chalk shell microstructures of the oyster Magallana: The same materials grown under different conditions. Sci. Rep. 2018, 8, 7507. [Google Scholar] [PubMed]
- Nevesskaya, L.A. Stages of Benthos Development in the Phanerozoic Seas, Mesozoic, Cenozoic; Transactions of PIN RAS; Rozanov, A.Y., Ed.; “Nauka”: Moscow, Russia, 1999; Volume 274, 438p. (In Russian) [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pakhnevich, A.; Nikolayev, D.; Lychagina, T. Crystallographic Texture of the Mineral Matter in the Bivalve Shells of Gryphaea dilatata Sowerby, 1816. Biology 2022, 11, 1300. https://doi.org/10.3390/biology11091300
Pakhnevich A, Nikolayev D, Lychagina T. Crystallographic Texture of the Mineral Matter in the Bivalve Shells of Gryphaea dilatata Sowerby, 1816. Biology. 2022; 11(9):1300. https://doi.org/10.3390/biology11091300
Chicago/Turabian StylePakhnevich, Alexey, Dmitry Nikolayev, and Tatiana Lychagina. 2022. "Crystallographic Texture of the Mineral Matter in the Bivalve Shells of Gryphaea dilatata Sowerby, 1816" Biology 11, no. 9: 1300. https://doi.org/10.3390/biology11091300
APA StylePakhnevich, A., Nikolayev, D., & Lychagina, T. (2022). Crystallographic Texture of the Mineral Matter in the Bivalve Shells of Gryphaea dilatata Sowerby, 1816. Biology, 11(9), 1300. https://doi.org/10.3390/biology11091300