
Vectors as Sentinels: Rising Temperatures Increase the Risk of Xylella fastidiosa Outbreaks

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
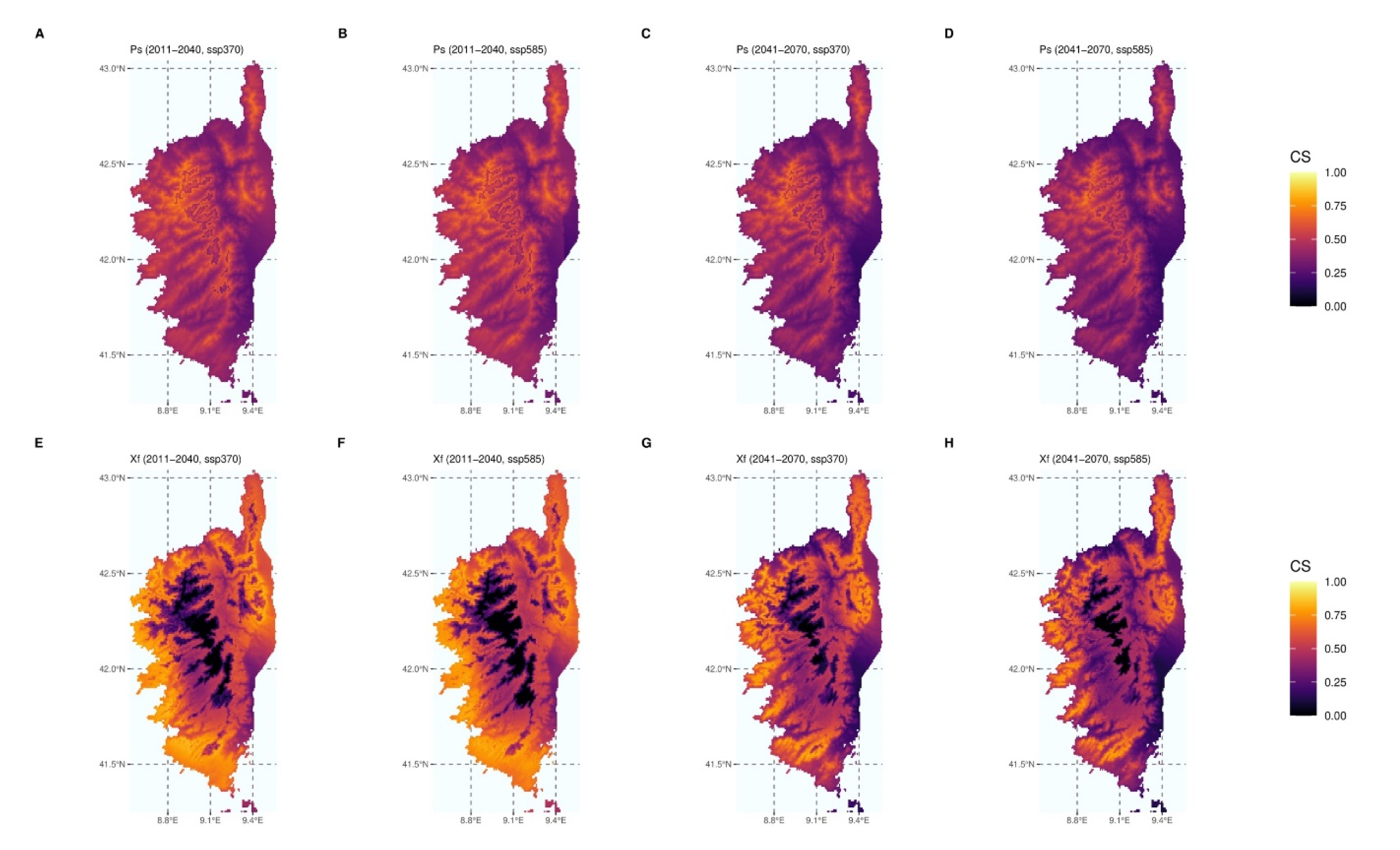
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Chaloner, T.M.; Gurr, S.J.; Bebber, D.P. Plant pathogen infection risk tracks global crop yields under climate change. Nat. Clim. Chang. 2021, 11, 710–715. [Google Scholar] [CrossRef]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef]
- Juroszek, P.; Racca, P.; Link, S.; Farhumand, J.; Kleinhenz, B. Overview on the review articles published during the past 30 years relating to the potential climate change effects on plant pathogens and crop disease risks. Plant Pathol. 2019, 69, 179–193. [Google Scholar] [CrossRef]
- IPPC Secretariat. Scientific Review of the Impact of Climate Change on Plant Pests—A Global Challenge to Prevent and Mitigate Plant Pest Risks in Agriculture, Forestry and Ecosystems; IPPC Secretariat: Rome, Italy, 2021. [Google Scholar]
- Jeger, M.J. The impact of climate change on disease in wild plant populations and communities. Plant Pathol. 2021, 71, 111–130. [Google Scholar] [CrossRef]
- Huang, W.; Reyes-Caldas, P.; Mann, M.; Seifbarghi, S.; Kahn, A.; Almeida, R.P.; Béven, L.; Heck, M.; Hogenhout, S.A.; Coaker, G. Bacterial Vector-Borne Plant Diseases: Unanswered Questions and Future Directions. Mol. Plant 2020, 13, 1379–1393. [Google Scholar] [CrossRef]
- Perilla-Henao, L.M.; Casteel, C.L. Vector-Borne Bacterial Plant Pathogens: Interactions with Hemipteran Insects and Plants. Front. Plant Sci. 2016, 7, 1163. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); Delbianco, A.; Gibin, D.; Pasinato, L.; Morelli, M. Update of the Xylella spp. host plant database—Systematic literature search up to 30 June 2021. EFSA J. 2022, 20, 7039. [Google Scholar]
- Chatterjee, S.; Almeida, R.P.P.; Lindow, S. Living in two Worlds: The Plant and Insect Lifestyles of Xylella fastidiosa. Annu. Rev. Phytopathol. 2008, 46, 243–271. [Google Scholar] [CrossRef]
- Cornara, D.; Morente, M.; Markheiser, A.; Bodino, N.; Tsai, C.-W.; Fereres, A.; Redak, R.A.; Perring, T.M.; Lopes, J.R.S. An overview on the worldwide vectors of Xylella fastidiosa. Èntomol. Gen. 2019, 39, 157–181. [Google Scholar] [CrossRef]
- Krugner, R.; Sisterson, M.S.; Backus, E.A.; Burbank, L.P.; Redak, R.A. Sharpshooters: A review of what moves Xylella fastidiosa. Austral Èntomol. 2019, 58, 248–267. [Google Scholar] [CrossRef]
- Purcell, A.H.; Finlay, A. Evidence of noncirculative transmission of Pierce’s Disease bacterium by sharpshooter leafhoppers. Phytopathology 1979, 69, 393–395. [Google Scholar] [CrossRef]
- Bové, J.M.; Ayres, A.J. Etiology of three recent diseases of citrus in São Paulo State: Sudden death, variegated chlorosis and huanglongbing. IUBMB Life 2007, 59, 346–354. [Google Scholar] [CrossRef]
- Tumber, K.P.; Alston, J.M.; Fuller, K.B. Pierce’s disease costs California $104 million per year. Calif. Agric. 2014, 68, 20–29. [Google Scholar] [CrossRef]
- Saponari, M.; Giampetruzzi, A.; Loconsole, G.; Boscia, D.; Saldarelli, P. Xylella fastidiosa in Olive in Apulia: Where We Stand. Phytopathology 2019, 109, 175–186. [Google Scholar] [CrossRef]
- Schneider, K.; van der Werf, W.; Cendoya, M.; Mourits, M.; Navas-Cortés, J.A.; Vicent, A.; Lansink, A.O. Impact of Xylella fastidiosa subspecies pauca in European olives. Proc. Natl. Acad. Sci. USA 2020, 117, 9250–9259. [Google Scholar] [CrossRef]
- Daugherty, M.P.; Cooper, M.; Smith, R.; Varela, L.; Almeida, R. Has Climate Contributed to a Pierce’s Disease Resurgence in North Coast Vineyards? PWV: San Rafael, CA, USA, 2019. [Google Scholar]
- Daugherty, M.P.; Zeilinger, A.R.; Almeida, R.P.P. Conflicting Effects of Climate and Vector Behavior on the Spread of a Plant Pathogen. Phytobiomes J. 2017, 1, 46–53. [Google Scholar] [CrossRef]
- Daugherty, M.; Bosco, D.; Almeida, R. Temperature mediates vector transmission efficiency: Inoculum supply and plant infection dynamics. Ann. Appl. Biol. 2009, 155, 361–369. [Google Scholar] [CrossRef]
- Gruber, B.R.; Daugherty, M.P. Predicting the effects of seasonality on the risk of pathogen spread in vineyards: Vector pressure, natural infectivity, and host recovery. Plant Pathol. 2013, 62, 194–204. [Google Scholar] [CrossRef]
- Sicard, A.; Zeilinger, A.R.; Vanhove, M.; Schartel, T.E.; Beal, D.J.; Daugherty, M.P.; Almeida, R.P. Xylella fastidiosa: Insights into an Emerging Plant Pathogen. Annu. Rev. Phytopathol. 2018, 56, 181–202. [Google Scholar] [CrossRef] [Green Version]
- Colegrave, N.; Ruxton, G.D. Using Biological Insight and Pragmatism When Thinking about Pseudoreplication. Trends Ecol. Evol. 2018, 33, 28–35. [Google Scholar] [CrossRef]
- Faraway, J.J. Extending the Linear Model with R: Generalized Linear, Mixed Effects and Non-Parametric Regression Models, 2nd ed.; Chapman & Hall: London, UK; CRC Taylor & Francis Group: Boca Raton, FL, USA, 2016. [Google Scholar]
- Bodino, N.; Cavalieri, V.; Pegoraro, M.; Altamura, G.; Canuto, F.; Zicca, S.; Fumarola, G.; Almeida, R.P.P.; Saponari, M.; Dongiovanni, C.; et al. Temporal dynamics of the transmission of Xylella fastidiosa subsp. pauca by Philaenus spumarius to olive plants. Èntomol. Gen. 2021, 41, 463–480. [Google Scholar] [CrossRef]
- Cruaud, A.; Gonzalez, A.-A.; Godefroid, M.; Nidelet, S.; Streito, J.-C.; Thuillier, J.-M.; Rossi, J.-P.; Santoni, S.; Rasplus, J.-Y. Using insects to detect, monitor and predict the distribution of Xylella fastidiosa: A case study in Corsica. Sci. Rep. 2018, 8, 15628. [Google Scholar] [CrossRef]
- Yuan, X.; Morano, L.; Bromley, R.; Spring-Pearson, S.; Stouthamer, R.; Nunney, L. Multilocus Sequence Typing ofXylella fastidiosaCausing Pierce’s Disease and Oleander Leaf Scorch in the United States. Phytopathology 2010, 100, 601–611. [Google Scholar] [CrossRef]
- Cruaud, P.; Rasplus, J.-Y.; Rodriguez, L.J.; Cruaud, A. High-throughput sequencing of multiple amplicons for barcoding and integrative taxonomy. Sci. Rep. 2017, 7, 41948. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm v2: Highly-scalable and high-resolution amplicon clustering. PeerJ 2015, 3, e1420. [Google Scholar] [CrossRef]
- Mehmood, T.; Liland, K.H.; Snipen, L.; Sæbø, S. A Review of Variable Selection Methods in Partial Least Squares Regression; Chemometr. Intell. Lab.: Amsterdam, The Netherlands, 2012; Volume 118, pp. 62–69. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Holland, R.M.; Christiano, R.S.C.; Gamliel-Atinsky, E.; Scherm, H. Distribution of Xylella fastidiosa in Blueberry Stem and Root Sections in Relation to Disease Severity in the Field. Plant Dis. 2014, 98, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Dupas, E.; Legendre, B.; Olivier, V.; Poliakoff, F.; Manceau, C.; Cunty, A. Comparison of real-time PCR and droplet digital PCR for the detection of Xylella fastidiosa in plants. J. Microbiol. Methods 2019, 162, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Costa, H.S.; Blua, M.S.; Bethke, J.A.; Redak, R.A. Transmission of Xylella fastidiosa to Oleander by the Glassywinged Sharp-shooter, Homalodisca coagulata. HortScience 2000, 35, 1265–1267. [Google Scholar] [CrossRef]
- Feil, H.; Purcell, A.H. Temperature-Dependent Growth and Survival of Xylella fastidiosa in Vitro and in Potted Grapevines. Plant Dis. 2001, 85, 1230–1234. [Google Scholar] [CrossRef] [PubMed]
- Purcell, A.H. Cold therapy of Pierce’s disease of grapevines. Plant Dis. Rep. 1977, 61, 514–518. [Google Scholar]
- Feil, H.; Feil, W.S.; Purcell, A.H. Effects of Date of Inoculation on the Within-Plant Movement of Xylella fastidiosa and Persistence of Pierce’s Disease Within Field Grapevines. Phytopathology 2003, 93, 244–251. [Google Scholar] [CrossRef]
- Cornara, D.; Bosco, D.; Fereres, A. Philaenus spumarius: When an old acquaintance becomes a new threat to European agri-culture. J. Pest Sci. 2018, 91, 957–972. [Google Scholar] [CrossRef]
- Cornara, D.; Panzarino, O.; Santoiemma, G.; Bodino, N.; Loverre, P.; Mastronardi, M.G.; Mattia, C.; De Lillo, E.; Addante, R. Natural areas as reservoir of candidate vectors of Xylella fastidiosa. B. Insectol. 2021, 74, 173–180. [Google Scholar]
- Bodino, N.; Cavalieri, V.; Dongiovanni, C.; Plazio, E.; Saladini, M.A.; Volani, S.; Simonetto, A.; Fumarola, G.; Di Carolo, M.; Porcelli, F.; et al. Phenology, seasonal abundance and stage-structure of spittlebug (Hemiptera: Aphrophoridae) populations in olive groves in Italy. Sci. Rep. 2019, 9, 17725. [Google Scholar] [CrossRef] [PubMed]
- Albre, J.; Carrasco, J.M.G.; Gibernau, M. Ecology of the meadow spittlebug Philaenus spumarius in the Ajaccio region (Corsica)—I: Spring. Bull. Èntomol. Res. 2020, 111, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Roper, C.; Castro, C.; Ingel, B. Xylella fastidiosa: Bacterial parasitism with hallmarks of commensalism. Curr. Opin. Plant Biol. 2019, 50, 140–147. [Google Scholar] [CrossRef]
- Krugner, R.; Backus, E.A. Plant Water Stress Effects on Stylet Probing Behaviors of Homalodisca vitripennis (Hemiptera: Cicadellidae) Associated with Acquisition and Inoculation of the Bacterium Xylella fastidiosa. J. Econ. Èntomol. 2014, 107, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Daugherty, M.P.; Almeida, R.P.P. Understanding How an Invasive Vector Drives Pierce’s Disease Epidemics: Seasonality and Vine-to-Vine Spread. Phytopathology 2019, 109, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.P.; Blua, M.J.; Lopes, J.R.; Purcell, A.H. Vector Transmission of Xylella fastidiosa: Applying Fundamental Knowledge to Generate Disease Management Strategies. Ann. Èntomol. Soc. Am. 2005, 98, 775–786. [Google Scholar] [CrossRef]
- Mesmin, X.; Chartois, M.; Borgomano, S.; Rasplus, J.Y.; Rossi, J.P.; Cruaud, A. Interaction networks between spittlebugs and plants in and around olive and clementine groves of Corsica; implications for the management of Xylella fastidiosa. Agr. Ecosyst. Environ. 2022, 334, 107979. [Google Scholar] [CrossRef]
- Godefroid, M.; Morente, M.; Schartel, T.; Cornara, D.; Purcell, A.; Gallego, D.; Moreno, A.; Pereira, J.A.; Fereres, A. Climate tolerances of Philaenus spumarius should be considered in risk assessment of disease outbreaks related to Xylella fastidiosa. J. Pest. Sci. 2021, 95, 855–868. [Google Scholar] [CrossRef]
- Godefroid, M.; Cruaud, A.; Streito, J.C.; Rasplus, J.Y.; Rossi, J.P. Climate change and the potential distribution of Xylella fas-tidiosa in Europe. bioRxiv 2018. [Google Scholar] [CrossRef]
- Hällfors, M.H.; Liao, J.; Dzurisin, J.; Grundel, R.; Hyvärinen, M.; Towle, K.; Wu, G.C.; Hellmann, J.J. Addressing potential local adaptation in species distribution models: Implications for conservation under climate change. Ecol. Appl. 2016, 26, 1154–1169. [Google Scholar] [CrossRef]
- Vanhove, M.; Retchless, A.C.; Sicard, A.; Rieux, A.; Coletta-Filho, H.; De La Fuente, L.; Stenger, D.C.; Almeida, R.P.P. Genomic Diversity and Recombination among Xylella fastidiosa Subspecies. Appl. Environ. Microbiol. 2019, 85, e02972-18. [Google Scholar] [CrossRef] [PubMed]
- EPPO PM 7/24 (4) Xylella Fastidiosa. EPPO Bull. 2019, 49, 175–227. [CrossRef]
- Marshall, O.J. PerlPrimer: Cross-Platform, Graphical Primer Design for Standard, Bisulphite and Real-Time PCR. Bioinformatics 2004, 20, 2471–2472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcelletti, S.; Scortichini, M. Genome-Wide Comparison and Taxonomic Relatedness of Multiple Xylella Fastidiosa Strains Reveal the Occurrence of Three Subspecies and a New Xylella Species. Arch. Microbiol. 2016, 198, 803–812. [Google Scholar] [CrossRef]
- Martin, J.-F. Creating Error-Proof Indexes for High Throughput Sequencing 2019. Methods Mol. Biol. 2019, 840, 197–228. [Google Scholar]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Wistrom, C.; Purcell, A.H. The Fate of Xylella Fastidiosa in Vineyard Weeds and Other Alternate Hosts in California. Plant Dis. 2005, 89, 994–999. [Google Scholar] [CrossRef]
- Lieth, J.H.; Meyer, M.M.; Yeo, K.-H.; Kirkpatrick, B.C. Modeling Cold Curing of Pierce’s Disease in Vitis Vinifera ‘Pinot Noir’ and ‘Cabernet Sauvignon’ Grapevines in California. PhytopathologyTM 2011, 101, 1492–1500. [Google Scholar] [CrossRef]
- Purcell, A. Paradigms: Examples from the Bacterium Xylella Fastidiosa. Annu. Rev. Phytopathol. 2013, 51, 339–356. [Google Scholar] [CrossRef]
- Hewitt, W.B.; Frazier, N.W.; Freitag, J.H.; Winkler, A.J. Pierce’s Disease Investigations. Hilgardia 1949, 19, 207–264. [Google Scholar]
- R Core Team. R Version 3.5.1 (Feather Spray): A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: Https://Www.R-Project.Org/ (accessed on 12 July 2022).
- Quintana-Seguí, P.; Le Moigne, P.; Durand, Y.; Martin, E.; Habets, F.; Baillon, M.; Canellas, C.; Franchisteguy, L.; Morel, S. Analysis of Near-Surface Atmospheric Variables: Validation of the SAFRAN Analysis over France. J. Appl. Meteorol. Climatol. 2008, 47, 92–107. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. GlmmTMB Balances Speed and Flexibility among Packages for Zero-Inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Chessel, D.; Dufour, A.-B.; Thioulouse, J. The Ade4 Package—I: One-Table Methods. R News 2004, 4, 5–10. [Google Scholar]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-Regression: A Basic Tool of Chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Faraway, J.J. Extending the Linear Model with R: Generalized Linear, Mixed Effects and Nonparametric Regression Models; Chapman and Hall/CRC: Boca Raton, FL, 2006. [Google Scholar]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized Linear Mixed Models: A Practical Guide for Ecology and Evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. 2020. Available online: https://cran.r-project.org/web/packages/DHARMa/vignettes/DHARMa.html (accessed on 12 July 2022).
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Social Sciences Mcmaster: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. 2021. Available online: https://cran.r-project.org/web/packages/emmeans/emmeans.pdf (accessed on 12 July 2022).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at High Resolution for the Earth’s Land Surface Areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef]
- Xu, T.; Hutchinson, M. ANUCLIM Version 6.1 User Guide; Australia National University: Canberra, Australia, 2011. [Google Scholar]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Dismo: Species Distribution Modeling. Available online: https://cran.r-project.org/web/packages/dismo/dismo.pdf (accessed on 12 July 2022).
- Karger, D.N.; Schmatz, D.R.; Dettling, G.; Zimmermann, N.E. High-Resolution Monthly Precipitation and Temperature Time Series from 2006 to 2100. Sci. Data 2020, 7, 248. [Google Scholar] [CrossRef]
- Buisson, L.; Thuiller, W.; Casajus, N.; Lek, S.; Grenouillet, G. Uncertainty in Ensemble Forecasting of Species Distribution. Glob. Change Biol. 2010, 16, 1145–1157. [Google Scholar] [CrossRef]
- O’Neill, B.C.; Kriegler, E.; Riahi, K.; Ebi, K.L.; Hallegatte, S.; Carter, T.R.; Mathur, R.; van Vuuren, D.P. A New Scenario Framework for Climate Change Research: The Concept of Shared Socioeconomic Pathways. Clim. Change 2014, 122, 387–400. [Google Scholar] [CrossRef]
- Godefroid, M.; Cruaud, A.; Streito, J.-C.; Rasplus, J.-Y.; Rossi, J.-P. Xylella Fastidiosa: Climate Suitability of European Continent. Sci. Rep. 2019, 9, 8844. [Google Scholar] [CrossRef] [PubMed]
- Falsini, S.; Tani, C.; Sambuco, G.; Papini, A.; Faraoni, P.; Campigli, S.; Ghelardini, L.; Bleve, G.; Rizzo, D.; Ricciolini, M.; et al. Anatomical and Biochemical Studies of Spartium Junceum Infected by Xylella fastidiosa Subsp. Multiplex ST 87. Protoplasma 2021, 259, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial Filtering to Reduce Sampling Bias Can Improve the Performance of Ecological Niche Models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. SpThin: An R Package for Spatial Thinning of Species Occurrence Records for Use in Ecological Niche Models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample Selection Bias and Presence-Only Distribution Models: Implications for Background and Pseudo-Absence Data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Naimi, B.; Hamm, N.A.S.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where Is Positional Uncertainty a Problem for Species Distribution Modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Varela, S.; Anderson, R.P.; García-Valdés, R.; Fernández-González, F. Environmental Filters Reduce the Effects of Sampling Bias and Improve Predictions of Ecological Niche Models. Ecography 2014, 2014, 1084–1091. [Google Scholar] [CrossRef]
- VanDerWal, J.; Shoo, L.P.; Graham, C.; Williams, S.E. Selecting Pseudo-Absence Data for Presence-Only Distribution Modeling: How Far Should You Stray from What You Know? Ecol. Model. 2009, 220, 589–594. [Google Scholar] [CrossRef]
- Seabra, S.G.; Rodrigues, A.S.B.; Silva, S.E.; Neto, A.C.; Pina-Martins, F.; Marabuto, E.; Thompson, V.; Wilson, M.R.; Yurtsever, S.; Halkka, A.; et al. Population Structure, Adaptation and Divergence of the Meadow Spittlebug, Philaenus Spumarius (Hemiptera, Aphrophoridae), Revealed by Genomic and Morphological Data. PeerJ 2021, 9, e11425. [Google Scholar] [CrossRef]
- Vollering, J.; Halvorsen, R.; Mazzoni, S. The MIAmaxent R Package: Variable Transformation and Model Selection for Species Distribution Models. Ecol. Evol. 2019, 9, 12051–12068. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, S.; Halvorsen, R.; Bakkestuen, V. MIAT: Modular R-Wrappers for Flexible Implementation of MaxEnt Distribution Modelling. Ecol. Inform. 2015, 30, 215–221. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A Review of Methods for the Assessment of Prediction Errors in Conservation Presence/Absence Models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Le Lay, G.; Helfer, V.; Randin, C.; Guisan, A. Evaluating the Ability of Habitat Suitability Models to Predict Species Presences. Ecol. Model. 2006, 199, 142–152. [Google Scholar] [CrossRef]
- Yackulic, C.B.; Chandler, R.; Zipkin, E.F.; Royle, J.A.; Nichols, J.D.; Campbell Grant, E.H.; Veran, S. Presence-Only Modelling Using MAXENT: When Can We Trust the Inferences? Methods Ecol. Evol. 2013, 4, 236–243. [Google Scholar] [CrossRef]
- Streito, J.-C.; Chartois, M.; Pierre, É.; Dusoulier, F.; Armand, J.-M.; Gaudin, J.; Rossi, J.-P. Citizen Science and Niche Modeling to Track and Forecast the Expansion of the Brown Marmorated Stinkbug Halyomorpha Halys (Stål, 1855). Sci. Rep. 2021, 11, 11421. [Google Scholar] [CrossRef]
- Broennimann, O.; Cola, V.D.; Guisan, A. Ecospat: Spatial Ecology Miscellaneous Methods; 2020. Available online: https://cran.r-project.org/web/packages/ecospat/index.html (accessed on 12 July 2022).
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An Improved Dual-Indexing Approach for Multiplexed 16S RRNA Gene Sequencing on the Illumina MiSeq Platform. Microbiome 2014, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. 2017. Available online: https://cran.r-project.org/web/packages/factoextra/readme/README.html (accessed on 12 July 2022).
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix. 2017. Available online: https://scirp.org/reference/referencespapers.aspx?referenceid=3067218 (accessed on 12 July 2022).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farigoule, P.; Chartois, M.; Mesmin, X.; Lambert, M.; Rossi, J.-P.; Rasplus, J.-Y.; Cruaud, A. Vectors as Sentinels: Rising Temperatures Increase the Risk of Xylella fastidiosa Outbreaks. Biology 2022, 11, 1299. https://doi.org/10.3390/biology11091299
Farigoule P, Chartois M, Mesmin X, Lambert M, Rossi J-P, Rasplus J-Y, Cruaud A. Vectors as Sentinels: Rising Temperatures Increase the Risk of Xylella fastidiosa Outbreaks. Biology. 2022; 11(9):1299. https://doi.org/10.3390/biology11091299
Chicago/Turabian StyleFarigoule, Pauline, Marguerite Chartois, Xavier Mesmin, Maxime Lambert, Jean-Pierre Rossi, Jean-Yves Rasplus, and Astrid Cruaud. 2022. "Vectors as Sentinels: Rising Temperatures Increase the Risk of Xylella fastidiosa Outbreaks" Biology 11, no. 9: 1299. https://doi.org/10.3390/biology11091299
APA StyleFarigoule, P., Chartois, M., Mesmin, X., Lambert, M., Rossi, J.-P., Rasplus, J.-Y., & Cruaud, A. (2022). Vectors as Sentinels: Rising Temperatures Increase the Risk of Xylella fastidiosa Outbreaks. Biology, 11(9), 1299. https://doi.org/10.3390/biology11091299