
Effects on Steroid 5-Alpha Reductase Gene Expression of Thai Rice Bran Extracts and Molecular Dynamics Study on SRD5A2

 ,
,  ,
,  ,
,  ,
, 
 ,
,  ,
,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Plant Material and Extraction
2.3. Determination of Bioactive Compounds
2.3.1. γ-Oryzanol and Tocopherols
2.3.2. Total Phenolic Content
2.3.3. Total Flavonoid Content
2.4. Cell Culture
2.5. Cell Viability Assay
2.6. RNA Extraction and Semiquantitative RT-PCR Analysis
2.6.1. RNA Extraction
2.6.2. Semi-Quantitative RT-PCR
2.7. Computational Method Details
2.7.1. Protein and Ligand Preparation
2.7.2. Molecular Docking Study for 5-Alpha Reductase 2
2.7.3. Molecular Dynamics Simulation
2.7.4. Trajectory Analysis
2.8. Statistical Analysis
3. Results and Discussion
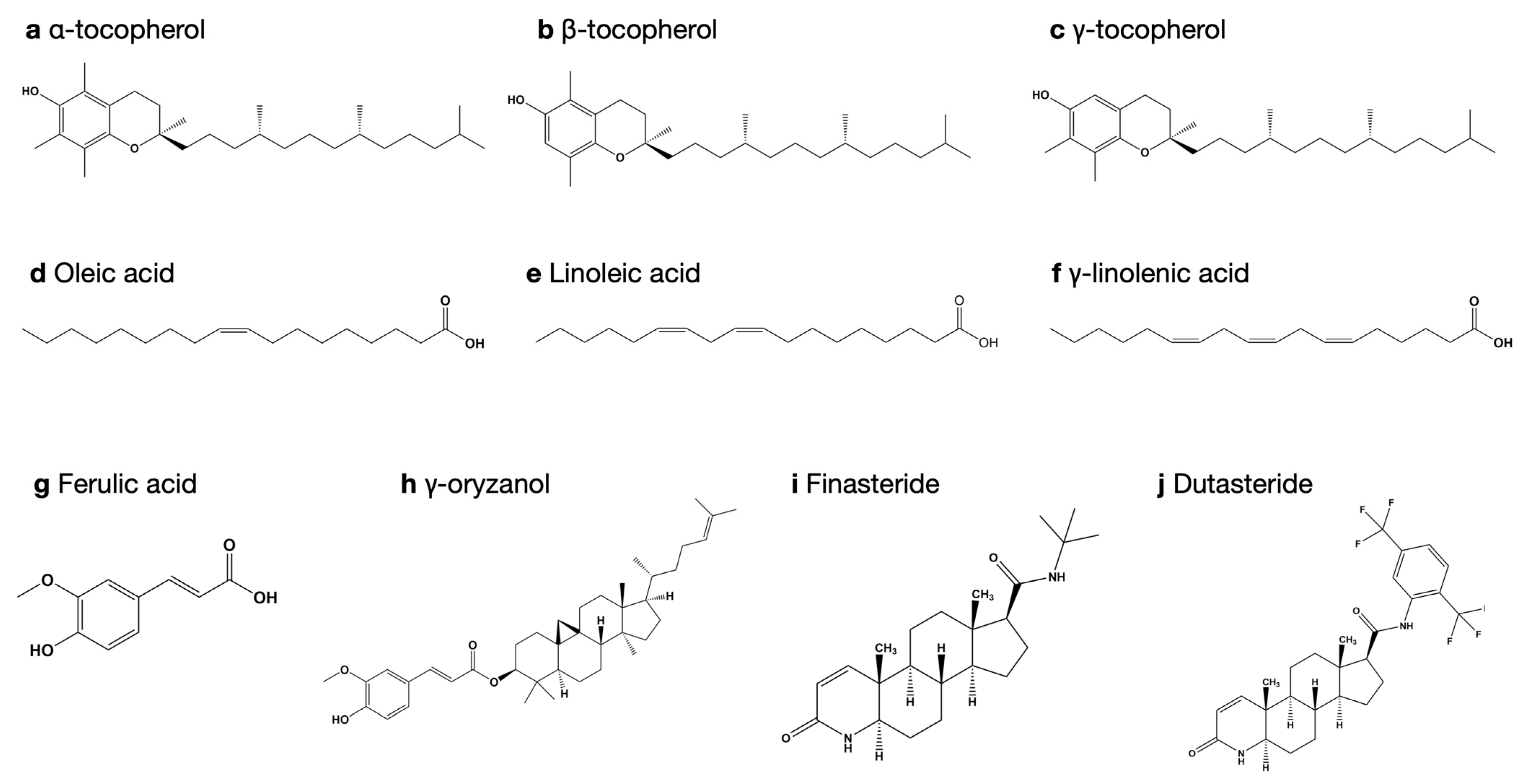
3.1. Extraction Yield and Bioactive Compounds
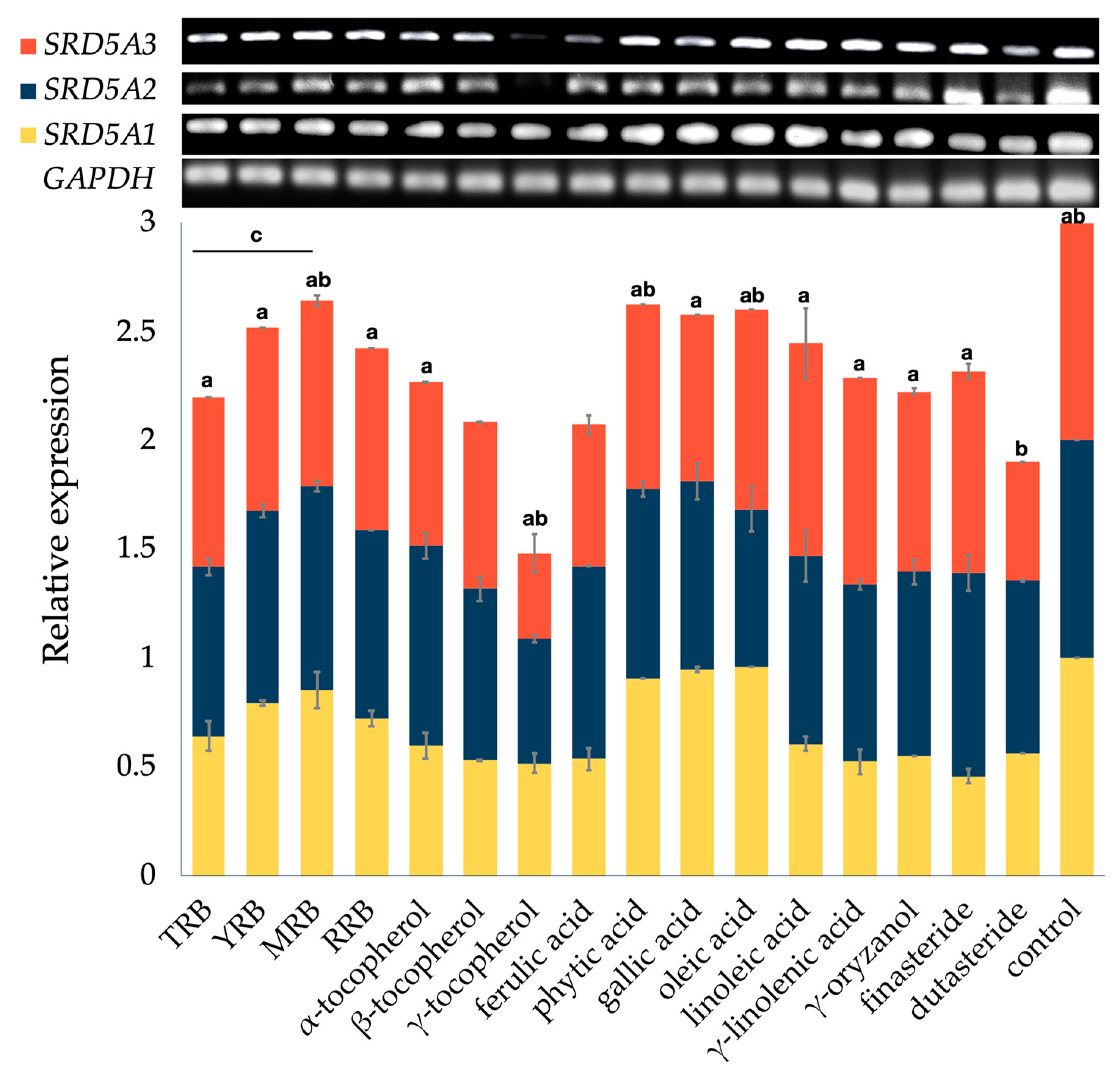
3.2. Effect on the Expression of 5-Alpha Reductase Isoenzymes
3.3. Correlation Analysis
3.3.1. Pearson’s Correlation
3.3.2. Correlation by Principal Component Analysis
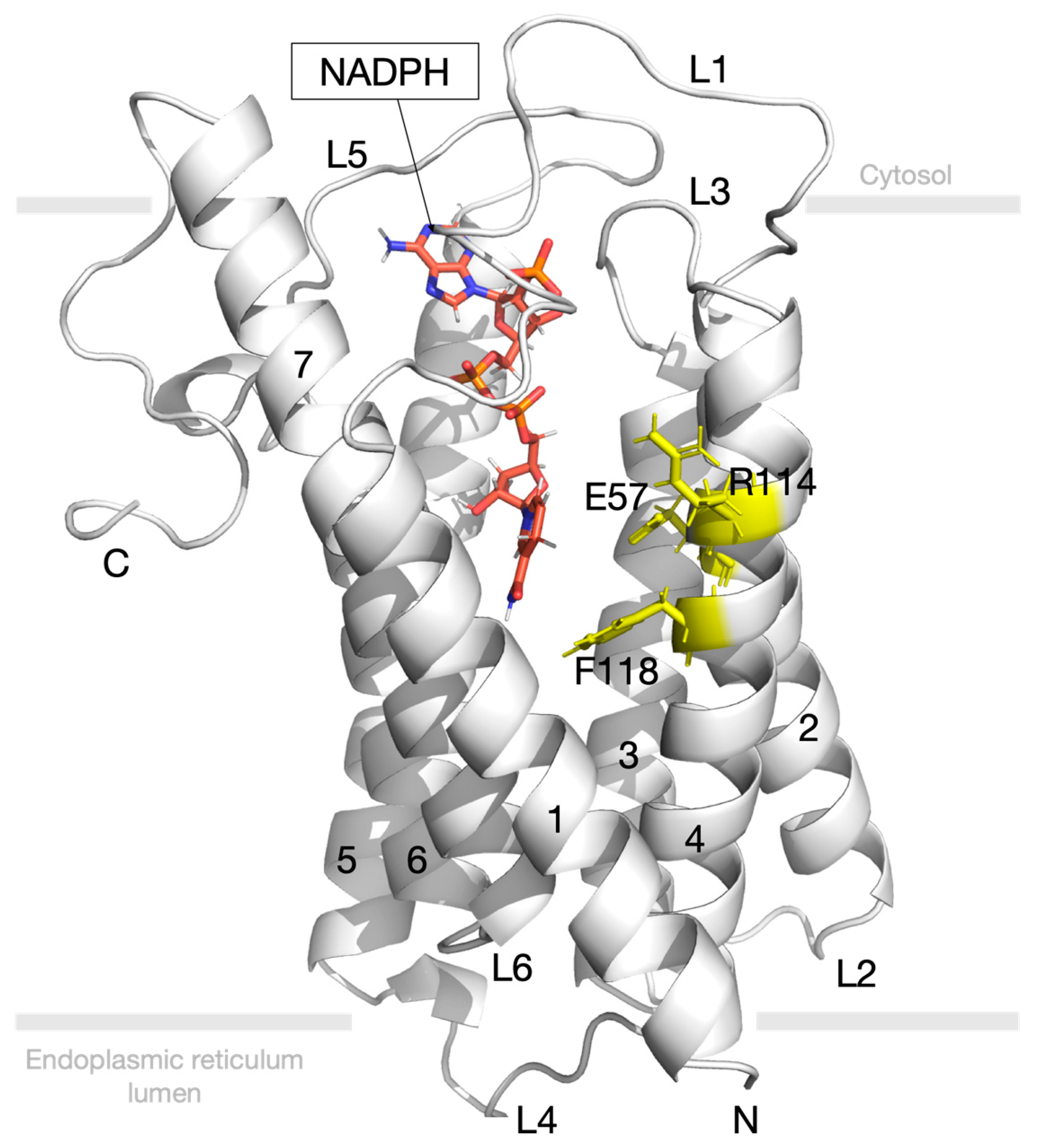
3.4. Molecular Docking Study for 5-Alpha Reductase 2
3.5. Molecular Dynamics Simulation
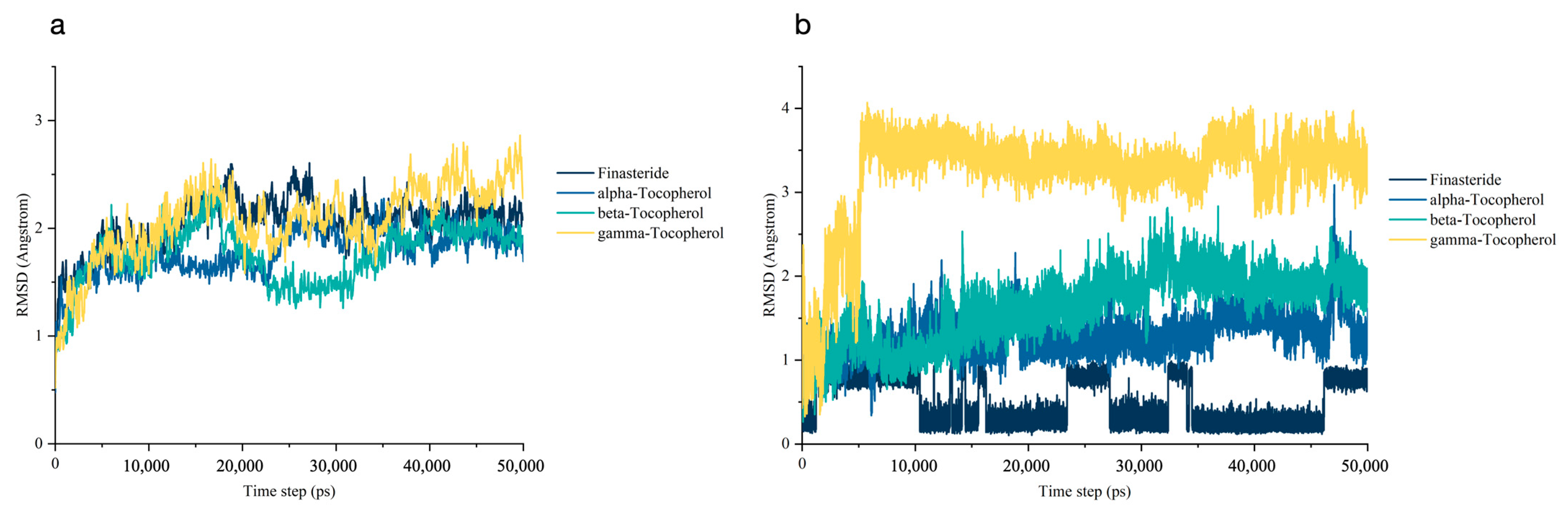
3.5.1. Stability of the Molecular Dynamics Trajectories
3.5.2. Binding Free Energy Analysis
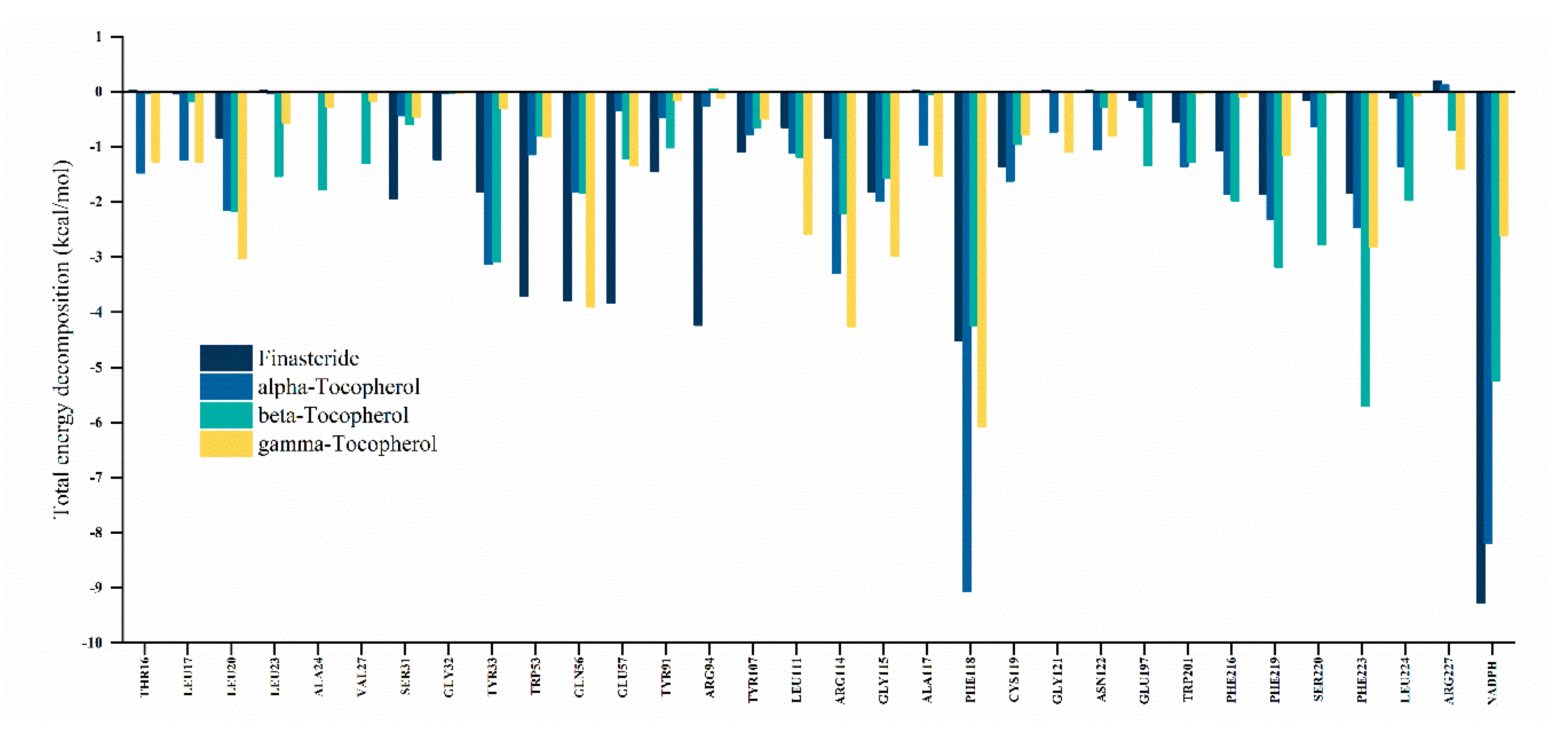
3.5.3. Decomposition of Binding Free Energy
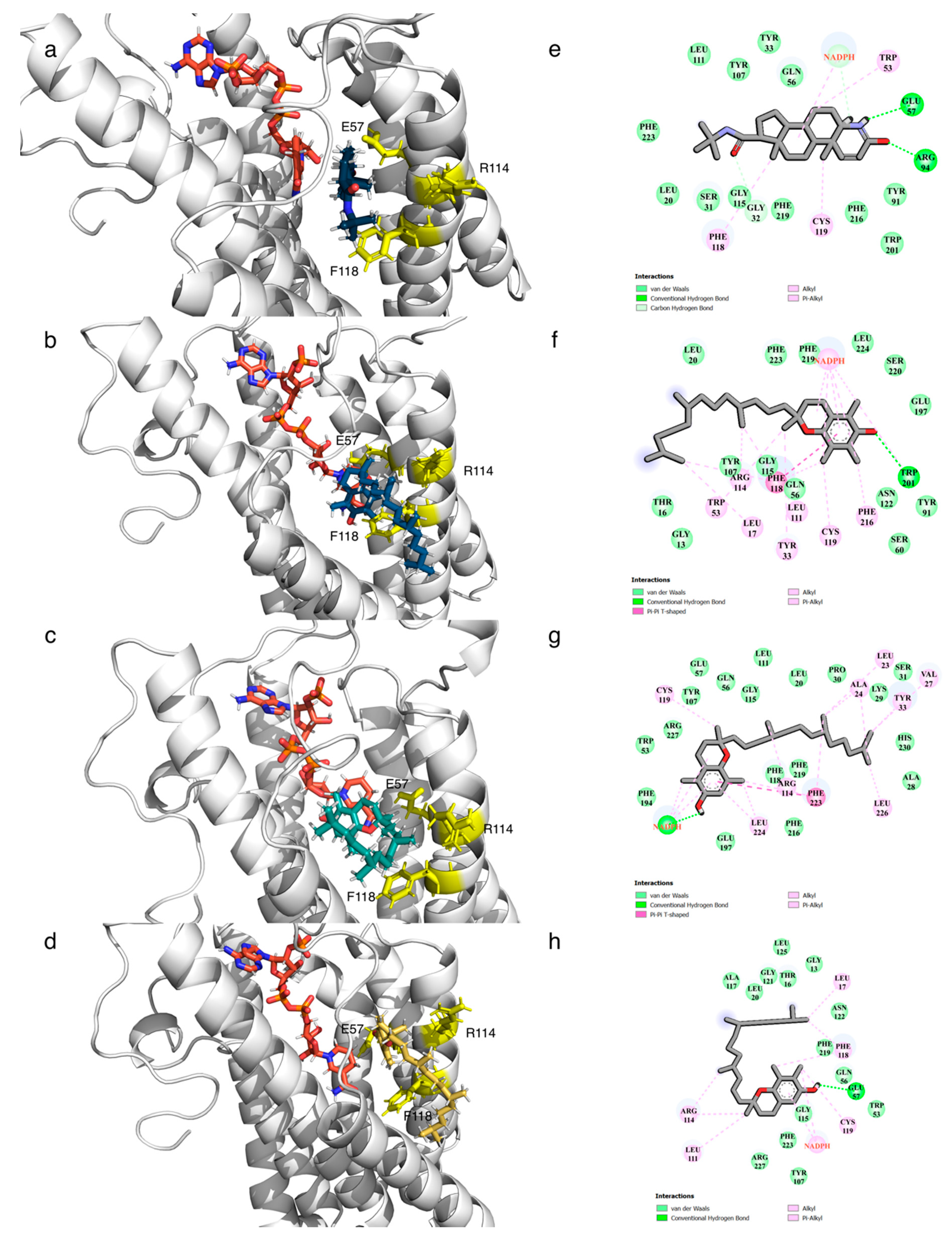
3.6. Post-Molecular Dynamics Simulation Binding Mode Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lolli, F.; Pallotti, F.; Rossi, A.; Fortuna, M.C.; Caro, G.; Lenzi, A.; Sansone, A.; Lombardo, F. Androgenetic alopecia: A review. Endocrine 2017, 57, 9–17. [Google Scholar] [CrossRef]
- Gupta, S.; Goyal, I.; Mahendra, A. Quality of life assessment in patients with androgenetic alopecia. Int. J. Trichol. 2019, 11, 147–152. [Google Scholar] [CrossRef]
- Gibson, D.A.; Saunders, P.T.K.; McEwan, I.J. Androgens and androgen receptor: Above and beyond. Mol. Cell. Endocrinol. 2018, 465, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Ceruti, J.M.; Leirós, G.J.; Balañá, M.E. Androgens and androgen receptor action in skin and hair follicles. Mol. Cell. Endocrinol. 2018, 465, 122–133. [Google Scholar] [CrossRef]
- Azzouni, F.; Godoy, A.; Li, Y.; Mohler, J. The 5 alpha-reductase isozyme family: A review of basic biology and their role in human diseases. Adv. Urol. 2011, 2012, 530121. [Google Scholar] [CrossRef]
- Robitaille, J.; Langlois, V.S. Consequences of steroid-5α-reductase deficiency and inhibition in vertebrates. Gen. Comp. Endocrinol. 2020, 290, 113400. [Google Scholar] [CrossRef] [PubMed]
- Katharopoulos, E.; Sauter, K.; Pandey, A.V.; Flück, C.E. In silico and functional studies reveal novel loss-of-function variants of SRD5A2, but no variants explaining excess 5α-reductase activity. J. Steroid Biochem. Mol. Biol. 2019, 190, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Chávez, B.; Ramos, L.; García-Becerra, R.; Vilchis, F. Hamster SRD5A3 lacks steroid 5α-reductase activity in vitro. Steroids 2015, 94, 41–50. [Google Scholar] [CrossRef]
- Jain, R.; Monthakantirat, O.; Tengamnuay, P.; Deeknamkul, W. Identification of a new plant extract for androgenic alopecia treatment using a non-radioactive human hair dermal papilla cell-based assay. BMC Complement. Med. Ther. 2015, 16, 18. [Google Scholar] [CrossRef]
- Inui, S.; Itami, S. Androgen actions on the human hair follicle: Perspectives. Exp. Dermatol. 2013, 22, 168–171. [Google Scholar] [CrossRef]
- Scaglione, A.; Montemiglio, L.C.; Parisi, G.; Asteriti, I.A.; Bruni, R.; Cerutti, G.; Testi, C.; Savino, C.; Mancia, F.; Lavia, P.; et al. Subcellular localization of the five members of the human steroid 5α-reductase family. Biochim. Open 2017, 4, 99–106. [Google Scholar] [CrossRef]
- Leirós, G.J.; Ceruti, J.M.; Castellanos, M.L.; Kusinsky, A.G.; Balañá, M.E. Androgens modify Wnt agonists/antagonists expression balance in dermal papilla cells preventing hair follicle stem cell differentiation in androgenetic alopecia. Mol. Cell. Endocrinol. 2017, 439, 26–34. [Google Scholar] [CrossRef]
- Sawaya, M.E.; Price, V.H. Different levels of 5alpha-reductase type I and II, aromatase, and androgen receptor in hair follicles of women and men with androgenetic alopecia. J. Investig. Dermatol. 1997, 109, 296–300. [Google Scholar] [CrossRef]
- Cantagrel, V.; Lefeber, D.J.; Ng, B.G.; Guan, Z.; Silhavy, J.L.; Bielas, S.L.; Lehle, L.; Hombauer, H.; Adamowicz, M.; Swiezewska, E.; et al. SRD5A3 is required for converting polyprenol to dolichol and is mutated in a congenital glycosylation disorder. Cell 2010, 142, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Uemura, M.; Tamura, K.; Chung, S.; Honma, S.; Okuyama, A.; Nakamura, Y.; Nakagawa, H. Novel 5α-steroid reductase (SRD5A3, type-3) is overexpressed in hormone-refractory prostate cancer. Cancer Sci. 2008, 99, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Yamana, K.; Labrie, F.; Luu The, V. Human type 3 5α-reductase is expressed in peripheral tissues at higher levels than types 1 and 2 and its activity is potently inhibited by finasteride and dutasteride. Horm. Mol. Biol. Clin. Investig. 2010, 2, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Godoy, A.; Kawinski, E.; Li, Y.; Oka, D.; Alexiev, B.; Azzouni, F.; Titus, M.A.; Mohler, J.L. 5α-reductase type 3 expression in human benign and malignant tissues: A comparative analysis during prostate cancer progression. Prostate 2011, 71, 1033–1046. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.K.; Levine, P.H.; Cleary, S.D.; Hoffman, H.J.; Graubard, B.I.; Cook, M.B. Male pattern baldness in relation to prostate cancer-specific mortality: A prospective analysis in the nhanes i epidemiologic follow-up study. Am. J. Epidemiol. 2016, 183, 210–217. [Google Scholar] [CrossRef]
- Jin, T.; Wu, T.; Luo, Z.; Duan, X.; Deng, S.; Tang, Y. Association between male pattern baldness and prostate disease: A meta-analysis. Urol. Oncol. 2018, 36, 80.e7–80.e15. [Google Scholar] [CrossRef]
- Sánchez, P.; Serrano Falcón, C.; Torres, J.; Serrano, S.; Ortega, E. 5α-Reductase isozymes and aromatase mRNA levels in plucked hair from young women with female pattern hair loss. Arch. Dermatol. Res. 2018, 310, 77–83. [Google Scholar] [CrossRef]
- Zhou, Z.; Song, S.; Gao, Z.; Wu, J.; Ma, J.; Cui, Y. The efficacy and safety of dutasteride compared with finasteride in treating men with androgenetic alopecia: A systematic review and meta-analysis. Clin. Interv. Aging 2019, 14, 399–406. [Google Scholar] [CrossRef]
- Herman, A.; Herman, A.P. Mechanism of action of herbs and their active constituents used in hair loss treatment. Fitoterapia 2016, 114, 18–25. [Google Scholar] [CrossRef]
- Khatun, A.; Waters, D.L.E.; Liu, L. A review of rice starch digestibility: Effect of composition and heat-moisture processing. Starke 2019, 71, 1900090. [Google Scholar] [CrossRef]
- Liu, J.; Rahman, S.; Sriboonchitta, S.; Wiboonpongse, A. Enhancing productivity and resource conservation by eliminating inefficiency of Thai rice farmers: A zero inefficiency stochastic frontier approach. Sustainability 2017, 9, 770. [Google Scholar] [CrossRef]
- Sen, S.; Chakraborty, R.; Kalita, P. Rice—Not just a staple food: A comprehensive review on its phytochemicals and therapeutic potential. Trends Food Sci. Technol. 2020, 97, 265–285. [Google Scholar] [CrossRef]
- Verma, D.K.; Srivastav, P.P. Bioactive compounds of rice (Oryza sativa L.): Review on paradigm and its potential benefit in human health. Trends Food Sci. Technol. 2020, 97, 355–365. [Google Scholar] [CrossRef]
- Liang, T.; Liao, S. Inhibition of steroid 5 α-reductase by specific aliphatic unsaturated fatty acids. Biochem. J. 1992, 285, 557–562. [Google Scholar] [CrossRef]
- Ruksiriwanich, W.; Manosroi, J.; Abe, M.; Manosroi, W.; Manosroi, A. 5α-Reductase type 1 inhibition of Oryza sativa bran extract prepared by supercritical carbon dioxide fluid. J. Supercrit. Fluids 2011, 59, 61–71. [Google Scholar] [CrossRef]
- Camacho-Martinez, F.M. Hair loss in women. In Seminars in Cutaneous Medicine and Surgery; Elsevier: Amsterdam, The Netherlands, 2009; pp. 19–32. [Google Scholar]
- Pestana Bauer, V.R.; Zambiazi, R.C.; Mendonca, C.R.; Beneito Cambra, M.; Ramis Ramos, G. γ-Oryzanol and tocopherol contents in residues of rice bran oil refining. Food Chem. 2012, 134, 1479–1483. [Google Scholar] [CrossRef]
- Chen, C.; Wang, L.; Wang, R.; Luo, X.; Li, Y.; Li, J.; Li, Y.; Chen, Z. Phenolic contents, cellular antioxidant activity and antiproliferative capacity of different varieties of oats. Food Chem. 2018, 239, 260–267. [Google Scholar] [CrossRef]
- Zeng, J.; Shang, X.; Zhang, P.; Wang, H.; Gu, Y.; Tan, J.-N. Combined use of deep eutectic solvents, macroporous resins, and preparative liquid chromatography for the isolation and purification of flavonoids and 20-hydroxyecdysone from Chenopodium quinoa willd. Biomolecules 2019, 9, 776. [Google Scholar] [CrossRef]
- Papazisis, K.; Geromichalos, G.; Dimitriadis, K.; Kortsaris, A. Optimization of the sulforhodamine B colorimetric assay. J. Immunol. Methods 1997, 208, 151–158. [Google Scholar] [CrossRef]
- Xiao, Q.; Wang, L.; Supekar, S.; Shen, T.; Liu, H.; Ye, F.; Huang, J.; Fan, H.; Wei, Z.; Zhang, C. Structure of human steroid 5α-reductase 2 with the anti-androgen drug finasteride. Nat. Commun. 2020, 11, 5430. [Google Scholar] [CrossRef]
- Frisch, M.; Trucks, G.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G. Gaussian 09, Revision D. 01; Gaussian, Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef]
- Nadvorny, D.; Soares Sobrinho, J.L.; de La Roca Soares, M.F.; Ribeiro, A.J.; Veiga, F.; Seabra, G.M. Molecular dynamics simulations reveal the influence of dextran sulfate in nanoparticle formation with calcium alginate to encapsulate insulin. J. Biomol. Struct. Dyn. 2018, 36, 1255–1260. [Google Scholar] [CrossRef]
- Bayly, C.I.; Cieplak, P.; Cornell, W.; Kollman, P.A. A well-behaved electrostatic potential based method using charge restraints for deriving atomic charges: The RESP model. J. Phys. Chem. 1993, 97, 10269–10280. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Beck, D.A.; Daggett, V. Methods for molecular dynamics simulations of protein folding/unfolding in solution. Methods 2004, 34, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Pastor, R.W.; Brooks, B.R.; Szabo, A. An analysis of the accuracy of Langevin and molecular dynamics algorithms. Mol. Phys. 1988, 65, 1409–1419. [Google Scholar] [CrossRef]
- Ryckaert, J.P.; Ciccotti, G.; Berendsen, H.J. Numerical integration of the cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- DeLano, W.L. PyMOL; Texas A&M University: College Station, TX, USA, 2002. [Google Scholar]
- Roe, D.R.; Cheatham, T.E., III. PTRAJ and CPPTRAJ: Software for processing and analysis of molecular dynamics trajectory data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.R., III; McGee, T.D., Jr.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA. py: An efficient program for end-state free energy calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef]
- Kollman, P.A.; Massova, I.; Reyes, C.; Kuhn, B.; Huo, S.; Chong, L.; Lee, M.; Lee, T.; Duan, Y.; Wang, W. Calculating structures and free energies of complex molecules: Combining molecular mechanics and continuum models. Acc. Chem. Res. 2000, 33, 889–897. [Google Scholar] [CrossRef]
- Cruz, J.N.; Costa, J.F.; Khayat, A.S.; Kuca, K.; Barros, C.A.; Neto, A. Molecular dynamics simulation and binding free energy studies of novel leads belonging to the benzofuran class inhibitors of Mycobacterium tuberculosis Polyketide Synthase 13. J. Biomol. Struct. Dyn. 2019, 37, 1616–1627. [Google Scholar] [CrossRef]
- Onufriev, A.; Bashford, D.; Case, D.A. Exploring protein native states and large-scale conformational changes with a modified generalized born model. Proteins 2004, 55, 383–394. [Google Scholar] [CrossRef]
- Weiser, J.; Shenkin, P.S.; Still, W.C. Approximate atomic surfaces from linear combinations of pairwise overlaps (LCPO). J. Comput. Chem. 1999, 20, 217–230. [Google Scholar] [CrossRef]
- Bhat, F.M.; Riar, C.S.; Seesuriyachan, P.; Sommano, S.R.; Chaiyaso, T.; Promuthai, C. Status of bioactive compounds from bran of pigmented traditional rice varieties and their scope in production of medicinal food with nutraceutical importance. Agronomy 2020, 10, 1817. [Google Scholar] [CrossRef]
- Goufo, P.; Trindade, H. Rice antioxidants: Phenolic acids, flavonoids, anthocyanins, proanthocyanidins, tocopherols, tocotrienols, γ-oryzanol, and phytic acid. Food Sci. Nutr. 2014, 2, 75–104. [Google Scholar] [CrossRef]
- Ling, B.; Lyng, J.G.; Wang, S. Effects of hot air-assisted radio frequency heating on enzyme inactivation, lipid stability and product quality of rice bran. LWT 2018, 91, 453–459. [Google Scholar] [CrossRef]
- Perretti, G.; Miniati, E.; Montanari, L.; Fantozzi, P. Improving the value of rice by-products by SFE. J. Supercrit. Fluids 2003, 26, 63–71. [Google Scholar] [CrossRef]
- Tsuzuki, W.; Komba, S.; Kotakec Nara, E. Diversity in γ-oryzanol profiles of Japanese black-purple rice varieties. J. Food Sci. Technol. 2019, 56, 2778–2786. [Google Scholar] [CrossRef]
- Surin, S.; Surayot, U.; Seesuriyachan, P.; You, S.; Phimolsiripol, Y. Antioxidant and immunomodulatory activities of sulphated polysaccharides from purple glutinous rice bran (Oryza sativa L.). Int. J. Food Sci. Technol. 2018, 53, 994–1004. [Google Scholar] [CrossRef]
- Muntana, N.; Prasong, S. Study on total phenolic contents and their antioxidant activities of Thai white, red and black rice bran extracts. Pak. J. Biol. Sci. 2010, 13, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Mbanjo, E.G.N.; Kretzschmar, T.; Jones, H.; Ereful, N.; Blanchard, C.; Boyd, L.A.; Sreenivasulu, N. The genetic basis and nutritional benefits of pigmented rice grain. Front. Genet. 2020, 11, 229. [Google Scholar] [CrossRef]
- Surin, S.; You, S.; Seesuriyachan, P.; Muangrat, R.; Wangtueai, S.; Jambrak, A.R.; Phongthai, S.; Jantanasakulwong, K.; Chaiyaso, T.; Phimolsiripol, Y. Optimization of ultrasonic-assisted extraction of polysaccharides from purple glutinous rice bran (Oryza sativa L.) and their antioxidant activities. Sci. Rep. 2020, 10, 10410. [Google Scholar] [CrossRef] [PubMed]
- Min, B.; Gu, L.; McClung, A.M.; Bergman, C.J.; Chen, M.H. Free and bound total phenolic concentrations, antioxidant capacities, and profiles of proanthocyanidins and anthocyanins in whole grain rice (Oryza sativa L.) of different bran colours. Food Chem. 2012, 133, 715–722. [Google Scholar] [CrossRef]
- Ehterami, A.; Salehi, M.; Farzamfar, S.; Samadian, H.; Vaez, A.; Ghorbani, S.; Ai, J.; Sahrapeyma, H. Chitosan/alginate hydrogels containing alpha-tocopherol for wound healing in rat model. J. Drug Deliv. Sci. Technol. 2019, 51, 204–213. [Google Scholar] [CrossRef]
- Harun, M.S.; Wong, T.W.; Fong, C.W. Advancing skin delivery of α-tocopherol and γ-tocotrienol for dermatitis treatment via nanotechnology and microwave technology. Int. J. Pharm. 2021, 593, 120099. [Google Scholar] [CrossRef]
- Nachbar, F.; Korting, H. The role of vitamin E in normal and damaged skin. J. Mol. Med. 1995, 73, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, M.; Sasaki, N. Effect of topical application of vitamin E on the hair growth of rabbits. J. Vitaminol. 1965, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Nohno, T.; Yoshida, H.; Yokoya, H. Trans-4-hydroxy-3-methoxycinnamic acid (ferulic acid) inhibits the effect of androgens on the rat prostate. Experientia 1979, 35, 696–699. [Google Scholar] [CrossRef] [PubMed]
- Abu El Saad, A.S.; Mahmoud, H.M. Phytic acid exposure alters aflatoxinb1-induced reproductive and oxidative toxicity in albino rats (Rattus norvegicus). Evid. Based Complement. Alternat. Med. 2009, 6, 107398. [Google Scholar] [CrossRef]
- Manosroi, A.; Ruksiriwanich, W.; Abe, M.; Manosroi, W.; Manosroi, J. Transfollicular enhancement of gel containing cationic niosomes loaded with unsaturated fatty acids in rice (Oryza sativa) bran semi-purified fraction. Eur. J. Pharm. Biopharm. 2012, 81, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Manosroi, A.; Ruksiriwanich, W.; Abe, M.; Sakai, H.; Aburai, K.; Manosroi, W.; Manosroi, J. Physico-chemical properties of cationic niosomes loaded with fraction of rice (Oryza sativa) bran extract. J. Nanosci. Nanotechnol. 2012, 12, 7339–7345. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.T.; Zheng, Q.C.; Wu, Y.J.; Zhang, J.L.; Liang, C.Y.; Chen, L.; Xue, Q.; Zhang, H.X. Molecular dynamics (MD) simulations and binding free energy calculation studies between inhibitors and type II dehydroquinase (DHQ2). Mol. Simul. 2013, 39, 137–144. [Google Scholar] [CrossRef]
- Takayasu, S.; Adachi, K. The conversion of testosterone to 17β-hydroxy-5α-androstan-3-one (dihydrotestosterone) by human hair follicles. J. Clin. Endocrinol. Metab. 1972, 34, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Trüeb, R.M. Molecular mechanisms of androgenetic alopecia. Exp. Gerontol. 2002, 37, 981–990. [Google Scholar] [CrossRef]
- Smith, C.M.; Ballard, S.A.; Worman, N.; Buettner, R.; Masters, J.R. 5 alpha-reductase expression by prostate cancer cell lines and benign prostatic hyperplasia in vitro. J. Clin. Endocrinol. Metab. 1996, 81, 1361–1366. [Google Scholar]
- Lourith, N.; Kanlayavattanakul, M.; Chaikul, P. Para rubber seed oil: The safe and efficient bio-material for hair loss treatment. J. Cosmet. Dermatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Fertig, R.; Shapiro, J.; Bergfeld, W.; Tosti, A. Investigation of the plausibility of 5-alpha-reductase inhibitor syndrome. Skin Appendage Disord. 2016, 2, 120–129. [Google Scholar] [CrossRef]
- Ganzer, C.A.; Jacobs, A.R.; Iqbal, F. Persistent sexual, emotional, and cognitive impairment post-finasteride: A survey of men reporting symptoms. Am. J. Men’s Health 2015, 9, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Coskuner, E.R.; Ozkan, B.; Culha, M.G. Sexual problems of men with androgenic alopecia treated with 5-alpha reductase inhibitors. Sex. Med. Rev. 2019, 7, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Manosroi, A.; Chaikul, P.; Chankhampan, C.; Ruksiriwanich, W.; Manosroi, W.; Manosroi, J. 5α-reductase inhibition and melanogenesis induction of the selected Thai plant extracts. Chiang Mai J. Sci. 2018, 45, 220–236. [Google Scholar]
- Akoglu, H. User’s guide to correlation coefficients. Turk. J. Emerg. Med. 2018, 18, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Clavel, C.; Rendl, M.; Watt, F.M. Hair follicle dermal papilla cells at a glance. J. Cell Sci. 2011, 124, 1179–1182. [Google Scholar] [CrossRef]
- Imperato Mcginley, J.; Peterson, R.E.; Gautier, T.; Cooper, G.; Danner, R.; Arthur, A.; Morris, P.L.; Sweeney, W.J.; Shackleton, C. Hormonal evaluation of a large kindred with complete androgen insensitivity: Evidence for secondary 5α-reductase deficiency. J. Clin. Endocrinol. Metab. 1982, 54, 931–941. [Google Scholar] [CrossRef]
- Sharifi, N.; Auchus, R.J. Steroid biosynthesis and prostate cancer. Steroids 2012, 77, 719–726. [Google Scholar] [CrossRef]
- Han, Y.; Zhuang, Q.; Sun, B.; Lv, W.; Wang, S.; Xiao, Q.; Pang, B.; Zhou, Y.; Wang, F.; Chi, P.; et al. Crystal structure of steroid reductase SRD5A reveals conserved steroid reduction mechanism. Nat. Commun. 2021, 1, 449. [Google Scholar] [CrossRef] [PubMed]
- Suchonwanit, P.; Iamsumang, W.; Leerunyakul, K. Topical finasteride for the treatment of male androgenetic alopecia and female pattern hair loss: A review of the current literature. J. Dermatol. Treat. 2020, 31, 1–6. [Google Scholar] [CrossRef]
- Raynaud, J.P.; Cousse, H.; Martin, P.M. Inhibition of type 1 and type 2 5alpha-reductase activity by free fatty acids, active ingredients of Permixon. J. Steroid Biochem. Mol. Biol. 2002, 82, 233–239. [Google Scholar] [CrossRef]
- Park, W.-S.; Lee, C.-H.; Lee, B.-G.; Chang, I.-S. The extract of Thujae occidentalis semen inhibited 5α-reductase and androchronogenetic alopecia of B6CBAF1/j hybrid mouse. J. Dermatol. Sci. 2003, 31, 91–98. [Google Scholar] [CrossRef]
- Kumar, T.; Chaiyasut, C.; Suttajit, M. Screening of steroid 5-reductase inhibitory activity and total phenolic content of Thai plants. J. Med. Plant Res. 2011, 5, 1265–1271. [Google Scholar]
- Ganeshpurkar, A.; Singh, R.; Gore, P.G.; Kumar, D.; Gutti, G.; Kumar, A.; Singh, S.K. Structure-based screening and molecular dynamics simulation studies for the identification of potential acetylcholinesterase inhibitors. Mol. Simul. 2020, 46, 169–185. [Google Scholar] [CrossRef]
- Sk, M.F.; Roy, R.; Jonniya, N.A.; Poddar, S.; Kar, P. Elucidating biophysical basis of binding of inhibitors to SARS-CoV-2 main protease by using molecular dynamics simulations and free energy calculations. J. Biomol. Struct. Dyn. 2020, 38, 1–13. [Google Scholar] [CrossRef]
- Shamsara, J. Homology modeling of 5-alpha-reductase 2 using available experimental data. Interdiscip. Sci. 2019, 11, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Makridakis, N.; Akalu, A.; Reichardt, J.K. Identification and characterization of somatic steroid 5 α-reductase (SRD5A2) mutations in human prostate cancer tissue. Oncogene 2004, 23, 7399–7405. [Google Scholar] [CrossRef]
- Azzi, A. Tocopherols, tocotrienols and tocomonoenols: Many similar molecules but only one vitamin E. Redox Biol. 2019, 26, 101259. [Google Scholar] [CrossRef] [PubMed]
- Yui, Y.; Miyazaki, S.; Ma, Y.; Ohira, M.; Fiehn, O.; Ikegami, T.; McCalley, D.V.; Tanaka, N. Distinction of synthetic dl-α-tocopherol from natural vitamin E (d-α-tocopherol) by reversed-phase liquid chromatography enhanced selectivity of a polymeric C18 stationary phase at low temperature and/or at high pressure. J. Chromatogr. A 2016, 1450, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Goossens, K.; De Winter, H. Molecular dynamics simulations of membrane proteins: An overview. J. Chem. Inf. Model. 2018, 58, 2193–2202. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | NCBI Reference Sequence | Forward Primer | Reverse Primer |
|---|---|---|---|
| SRD5A1 | 001047.4 | AGCCATTGTGCAGTGTATGC | AGCCTCCCCTTGGTATTTTG |
| SRD5A2 | 000348.4 | TGAATACCCTGATGGGTGG | CAAGCCACCTTGTGGAATC |
| SRD5A3 | 024592.5 | TCCTTCTTTGCCCAAACATC | CTGATGCTCTCCCTTTACGC |
| GAPDH | 001289745.3 | GGAAGGTGAAGGTCGGAGTC | CTCAGCCTTGACGGTGCCATG |
| Sample | α-Tocopherol | (β+γ)-Tocopherol | γ-Oryzanol | TPC | TFC |
|---|---|---|---|---|---|
| mg/kg Extract | mg/kg Extract | mg/kg Extract | mg GAE/100 g | mg EGCGE/100 g | |
| TRB | 20.76 ± 0.13 | 23.32 ± 0.01 | 8600.45 ± 0.13 | 180.44 ± 6.42 | 569.01 ± 90.42 |
| YRB | 12.52 ± 0.01 | 50.98 ± 0.02 | 3773.17 ± 0.01 | 254.97 ± 5.20 | 880.16 ± 22.86 |
| MRB | 7.61 ± 0.01 | 3.51 ± 0.01 | ND | 125.67 ± 0.49 | 545.30 ± 48.33 |
| RRB | 11.95 ± 0.04 | 16.97 ± 0.01 | 9174.01 ± 0.09 | 56.47 ± 2.82 | 441.49 ± 12.39 |
| Compounds | AutoDock Binding Free Energy, ∆G (kcal/mol) |
|---|---|
| Native finasteride | −10.13 |
| β-Tocopherol | −9.83 |
| γ-Tocopherol | −9.53 |
| α-Tocopherol | −9.47 |
| Dutasteride | −8.75 |
| γ-Linolenic acid | −7.03 |
| Linoleic acid | −6.62 |
| Oleic acid | −6.49 |
| Ferulic acid | −5.22 |
| γ-Oryzanol | −4.26 |
| Phytic acid | −2.02 |
| Compounds | Binding Free Energy (kcal/mol) | ||||||
|---|---|---|---|---|---|---|---|
| VDW | EEL | EGB | ESURF | ∆Ggas | ∆Gsolv | ∆GTotal | |
| Finasteride | −53.93 ± 3.23 | −18.81 ± 3.41 | 32.36 ± 3.03 | −6.15 ± 0.24 | −72.74 ± 4.33 | 26.20 ± 2.95 | −46.54 ± 3.12 |
| α-Tocopherol | −64.97 ± 3.21 | −2.26 ± 1.68 | 20.10 ± 1.80 | −7.42 ± 0.39 | −67.23 ± 3.68 | 12.68 ± 1.77 | −54.55 ± 3.67 |
| β-Tocopherol | −60.01 ± 4.30 | −8.13 ± 2.90 | 25.88 ± 2.35 | −8.15 ± 0.55 | −68.14 ± 5.02 | 17.73 ± 2.10 | −50.40 ± 4.33 |
| γ-Tocopherol | −51.24 ± 4.41 | −5.71 ± 4.69 | 23.02 ± 3.76 | −6.77 ± 0.47 | −56.96 ± 5.54 | 16.25 ± 3.65 | −40.71 ± 3.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khantham, C.; Yooin, W.; Sringarm, K.; Sommano, S.R.; Jiranusornkul, S.; Carmona, F.D.; Nimlamool, W.; Jantrawut, P.; Rachtanapun, P.; Ruksiriwanich, W. Effects on Steroid 5-Alpha Reductase Gene Expression of Thai Rice Bran Extracts and Molecular Dynamics Study on SRD5A2. Biology 2021, 10, 319. https://doi.org/10.3390/biology10040319
Khantham C, Yooin W, Sringarm K, Sommano SR, Jiranusornkul S, Carmona FD, Nimlamool W, Jantrawut P, Rachtanapun P, Ruksiriwanich W. Effects on Steroid 5-Alpha Reductase Gene Expression of Thai Rice Bran Extracts and Molecular Dynamics Study on SRD5A2. Biology. 2021; 10(4):319. https://doi.org/10.3390/biology10040319
Chicago/Turabian StyleKhantham, Chiranan, Wipawadee Yooin, Korawan Sringarm, Sarana Rose Sommano, Supat Jiranusornkul, Francisco David Carmona, Wutigri Nimlamool, Pensak Jantrawut, Pornchai Rachtanapun, and Warintorn Ruksiriwanich. 2021. "Effects on Steroid 5-Alpha Reductase Gene Expression of Thai Rice Bran Extracts and Molecular Dynamics Study on SRD5A2" Biology 10, no. 4: 319. https://doi.org/10.3390/biology10040319
APA StyleKhantham, C., Yooin, W., Sringarm, K., Sommano, S. R., Jiranusornkul, S., Carmona, F. D., Nimlamool, W., Jantrawut, P., Rachtanapun, P., & Ruksiriwanich, W. (2021). Effects on Steroid 5-Alpha Reductase Gene Expression of Thai Rice Bran Extracts and Molecular Dynamics Study on SRD5A2. Biology, 10(4), 319. https://doi.org/10.3390/biology10040319