Decoding Antioxidant and Antibacterial Potentials of Malaysian Green Seaweeds: Caulerpa racemosa and Caulerpa lentillifera



Abstract
1. Introduction
2. Results
2.1. Yield Percentage of Seaweed Extracts
2.2. Determination of Total Phenolic Content (TPC) and Total Flavonoid Content (TFC)
2.3. Determination of DPPH Radical Scavenging Activity
2.4. Antibacterial Assay
2.4.1. Screening of Antibacterial Effect at 250 μg/mL
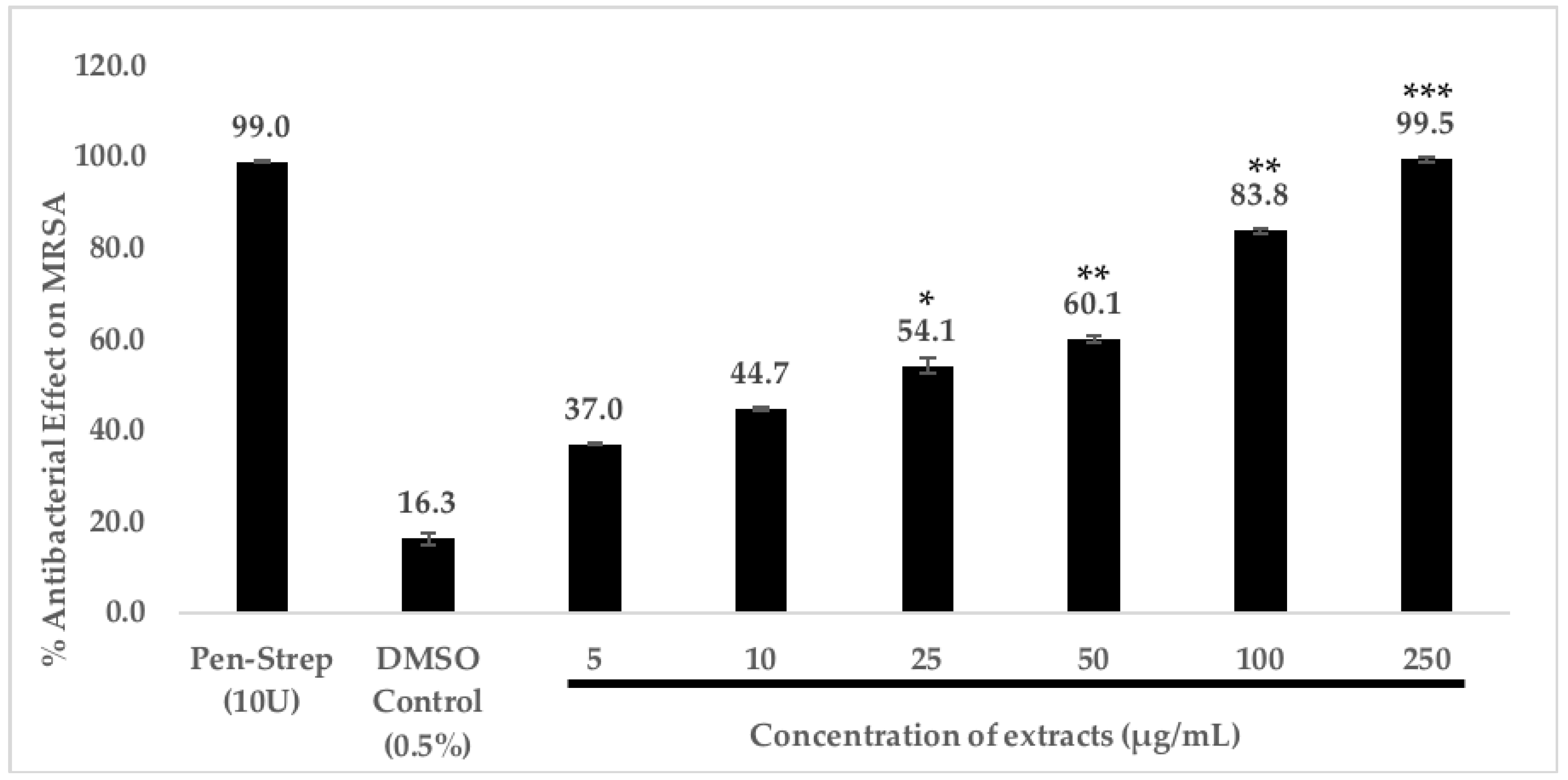
2.4.2. Dose-Dependent Antibacterial Effect of C. racemosa Chloroform Extract
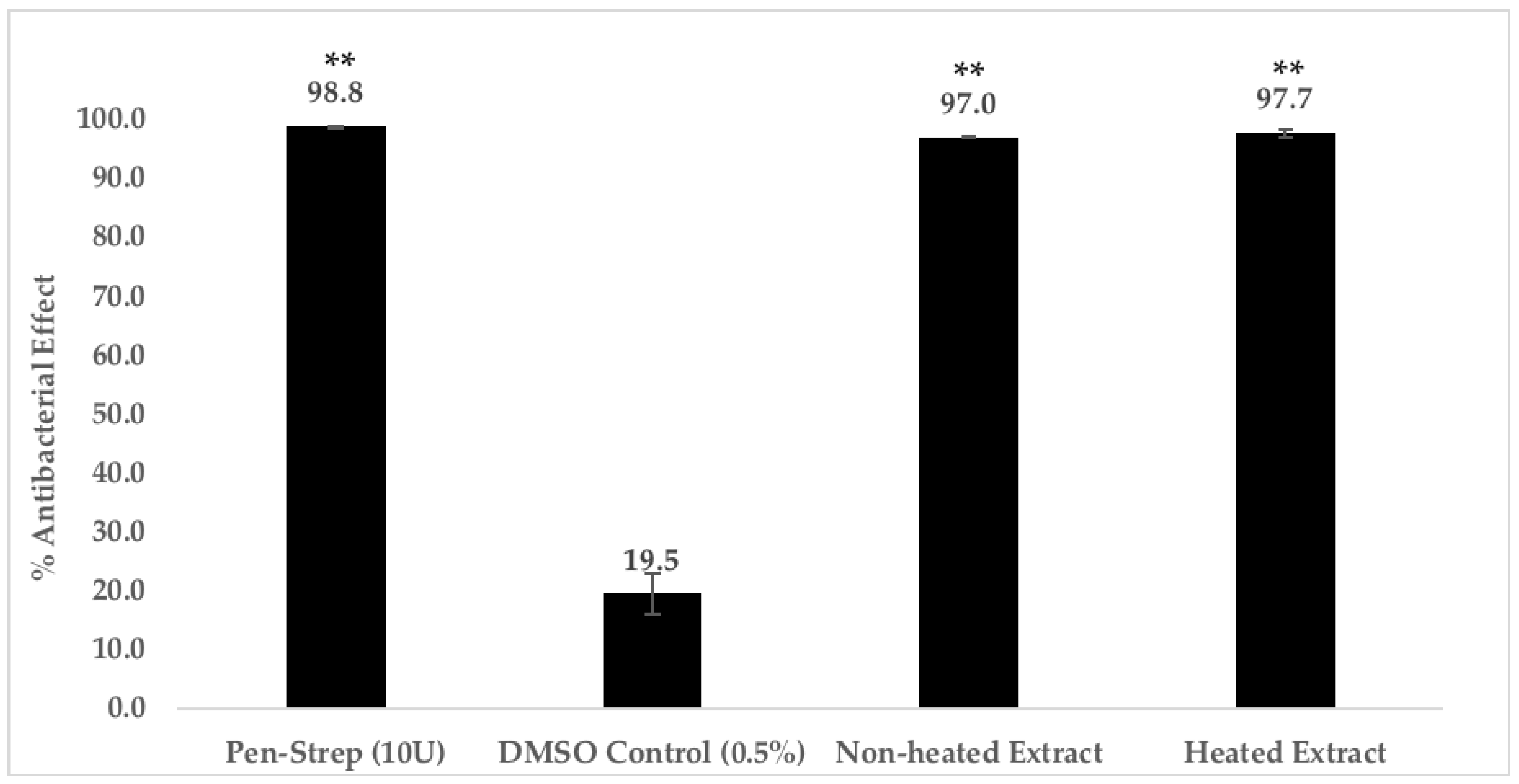
2.4.3. Heat Treatment of C. racemosa Chloroform Extract
2.5. Liquid Chromatography–Mass Spectrometry (LCMS) Analysis
3. Discussion
4. Materials and Methods
4.1. Seaweed Preparation
4.2. Crude Extract Preparations
4.3. Phytochemical Screening
4.3.1. Determination of Total Phenolic Content (TPC)
4.3.2. Determination of Total Flavonoid Content (TFC)
4.4. Determination of DPPH Radical Scavenging Activity
4.5. Antibacterial Assay
4.5.1. Screening of Antibacterial Effect at 250 μg/mL
4.5.2. Dose-Dependent Antibacterial Effect of C. racemosa Chloroform Extract
4.5.3. Heat Treatment of C. racemosa Chloroform Extract
4.6. Liquid Chromatography–Mass Spectrometry (LCMS) Analysis
4.6.1. Liquid Chromatography–Mass Spectrometry (LCMS): Separation and Analysis
4.6.2. Identification of Compounds Through Matching with Library
4.7. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, A.M.M.B.; de Morais, R.M.S.C. Emergent sources of prebiotics: Seaweeds and microalgae. Mar. Drugs 2016, 14, 27. [Google Scholar] [CrossRef]
- Yang, P.; Liu, D.Q.; Liang, T.J.; Li, J.; Zhang, H.Y.; Liu, A.H.; Guo, Y.W.; Mao, S.C. Bioactive constituents form the green alga Caulerpa racemosa. Bioorg. Med. Chem. 2015, 23, 38–45. [Google Scholar] [CrossRef]
- Kantachumpoo, A.M.; Chirapart, A. Components and antimicrobial activity of polysaccharides extracted from Thai brown seaweeds. Kasetsart J. (Nat. Sci.) 2010, 44, 220–233. [Google Scholar]
- Suganya, A.M.; Sanjivkumar, M.; Chandran, M.N.; Palavesam, A.; Immanuel, G. Pharmacological importance of sulphated polysaccharide carrageenan from red seaweed Kappaphycus alvarezii in comparison with commercial carrageenan. Biomed. Pharmacother. 2016, 84, 1300–1312. [Google Scholar] [CrossRef]
- Alshalmani, S.K.; Zobi, N.H.; Bozakouk, I.H. Antibacterial activity of Libyan seaweed extracts. Int. J. Pharm. Sci. Res. 2014, 5, 5425–5429. [Google Scholar] [CrossRef]
- Jeeva, S.; Antonisamy, J.M.; Domettila, C.; Anantham, B.; Mahesh, M. Preliminary phytochemical studies on some selected seaweeds from Gulf of Mannar, India. Asian Pac. J. Trop. Biomed. 2012, 2, 30–33. [Google Scholar] [CrossRef]
- Chatter, R.; Othman, R.B.; Rabhi, S.; Kladi, M.; Tarhouni, S.; Vagias, C.; Roussis, V.; Guizani-Tabbane, L.; Kharrat, R. In vivo and in vitro anti-inflammatory activity of neorogioltriol, a new diterpene extracted from the red algae Laurencia glandulifera. Mar. Drugs 2011, 9, 1293–1306. [Google Scholar] [CrossRef]
- Eccles, R.; Winther, B.; Johnston, S.L.; Robinson, P.; Trampisch, M.; Koelsch, S. Efficacy and safety of iota-carrageenan nasal spray versus placebo in early treatment of the common cold in adults: The ICICC trial. Respir. Res. 2015, 16, 121. [Google Scholar] [CrossRef]
- Kwiatek, M.A.; Roman, S.; Fareeduddin, A.; Pandolfino, J.E.; Kahrilas, P.J. An alginate-antacid formulation (Gaviscon Double Action Liquid) can eliminate or displace the postprandial ‘acid pocket’ in symptomatic GERD patients. Aliment. Pharmacol. Ther. 2011, 34, 59–66. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects–A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Park, S.; Han, S.S.; Park, C.H.; Hahm, E.R.; Lee, S.J.; Park, H.K.; Lee, S.H.; Kim, W.S.; Jung, C.W.; Park, K.; et al. L-ascorbic acid induces apoptosis in acute myeloid leukemia cells via hydrogen peroxide-mediated mechanisms. Int. J. Biochem. Cell Biol. 2004, 36, 2180–2195. [Google Scholar] [CrossRef]
- Park, S. The effects of high concentrations of vitamin C on cancer cells. Nutrients 2013, 5, 3496–3505. [Google Scholar] [CrossRef]
- Chauhan, V.; Chauhan, A. Oxidative stress in Alzheimer’s disease. Pathophysiology 2006, 13, 195–208. [Google Scholar] [CrossRef]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The role of oxidative stress in neurodegenerative diseases. Exp. Neurobiol. 2015, 24, 325–340. [Google Scholar] [CrossRef]
- Schumacker, P.T. Reactive oxygen species in cancer: A dance with the devil. Cancer Cell 2015, 27, 156–157. [Google Scholar] [CrossRef]
- Gao, P.; Zhang, H.; Dinavahi, R.; Li, F.; Xiang, Y.; Raman, V.; Bhujwalla, Z.M.; Felsher, D.W.; Cheng, L.; Pevsner, J.; et al. HIF-dependent anti-tumorigenic effect of anti-oxidants in vivo. Cancer Cell 2007, 12, 230–238. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Gladyshev, V.N. Role of reactive oxygen species-mediated signaling in aging. Antioxid. Redox Signal. 2013, 19, 1362–1372. [Google Scholar] [CrossRef]
- Sampath-Wiley, P.; Neefus, C.D.; Jahnke, L.S. Seasonal effects of sun exposure and emersion on intertidal seaweed physiology: Fluctuations in antioxidant contents, photosynthetic pigments and photosynthetic efficiency in the red alga Porphyra umbilicalis Kützing (Rhodophyta, Bangiales). J. Exp. Mar. Biol. Ecol. 2008, 361, 83–91. [Google Scholar] [CrossRef]
- Fisal, A.; Sulaiman, M.R.; Saimon, W.; Chye, F.Y.; Matanjun, P. Proximate compositions and total phenolic contents of selected edible seaweed from Semporna, Sabah, Malaysia. Borneo Sci. 2012, 31, 85–96. [Google Scholar]
- Matanjun, P.; Suhaila, M.; Noordin, M.M.; Kharidah, M.; Cheng, H.M. Antioxidant activities and phenolics content of eight species of seaweeds from north Borneo. J. Appl. Phycol. 2008, 20, 367–373. [Google Scholar] [CrossRef]
- Pereira, D.M.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Phenolics: From chemistry to biology. Molecules 2009, 14, 2202–2211. [Google Scholar] [CrossRef]
- Devi, K.P.; Suganthy, N.; Kesika, P.; Pandian, S.K. Bioprotective properties of seaweeds: In vitro evaluation of antioxidant activity and antimicrobial activity against food borne bacteria in relation to polyphenolic content. BMC Complement. Altern. Med. 2008, 8, 38. [Google Scholar] [CrossRef]
- Chakraborthy, K.; Joseph, D.; Praveen, N.K. Antioxidant activities and phenolic contents of three red seaweeds (Division: Rhodophyta) harvested from the Gulf of Mannar of Peninsular India. J. Food Sci. Technol. 2013, 52, 1924–1935. [Google Scholar] [CrossRef]
- Wang, B.G.; Zhang, W.W.; Duan, X.J.; Li, X.M. In vitro antioxidative activities of extract and semi-purified fractions of the marine red alga, Rhodomela confervoides (Rhodomelaceae). Food Chem. 2009, 113, 1101–1105. [Google Scholar] [CrossRef]
- Farasat, M.; Khavari-Nejad, R.-A.; Nabavi, S.M.B.; Namjooyan, F. Antioxidant properties of two edible green seaweeds from northern coasts of the Persian Gulf. Jundishapur J. Nat. Pharm. Prod. 2013, 8, 47–52. [Google Scholar] [CrossRef]
- Farasat, M.; Khavari-Nejad, R.-A.; Nabavi, S.M.B.; Namjooyan, F. Antioxidant activity, total phenolics and flavonoid contents of some edible green seaweeds from northern coasts of the Persian Gulf. Iran J. Pharm. Res. 2014, 13, 163–170. [Google Scholar]
- Kai, W.; Peisheng, Y.; Rui, M.; Wenwen, J.; Zongze, S. Diversity of culturable bacteria in deep-sea water from the South Atlantic Ocean. Bioengineered 2017, 8, 572–584. [Google Scholar] [CrossRef]
- Saha, M.; Wahl, M. Seasonal variation in the antifouling defence of the temperate brown alga Fucus vesiculosus. Biofouling 2013, 29, 661–668. [Google Scholar] [CrossRef]
- Nylund, G.M.; Cervin, G.; Hermansson, M.; Pavia, H. Chemical inhibition of bacteria colonization by the red alga Bonnemaisonia hamifera. Mar. Ecol. Prog. Ser. 2005, 302, 27–36. [Google Scholar] [CrossRef][Green Version]
- Mayer, A.M.S.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Fusetani, N. Marine pharmacology in 2009–2011: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar] [CrossRef]
- Rönnerstrand, B.; Sundell, K.A. Trust, reciprocity and collective action to fight antibiotic resistance. An experimental approach. Soc. Sci. Med. 2015, 142, 249–255. [Google Scholar] [CrossRef]
- Foon, T.S.; Ai, L.A.; Kuppusamy, P.; Yusoff, M.M.; Govindan, N. Studies on in vitro antioxidant activity of marine edible seaweed from east coastal region, Peninsular Malaysia using different extraction method. Res. J. Appl. Sci. 2014, 9, 141–146. [Google Scholar]
- Jaswir, I.; Tope, A.-H.T.; Raus, R.A.; Monsur, H.A.; RamLi, N. Study on anti-bacterial properties of some Malaysian brown seaweeds. Food Hydrocoll. 2014, 42, 275–279. [Google Scholar] [CrossRef]
- Natrah, F.M.I.; Harah, Z.M.; Sidik, B.J.; Izzatul, N.M.S.; Syahidah, A. Antibacterial activities of selected seaweed and seagrass from Port Dickson coastal water against different aquaculture pathogens. Sains Malays. 2015, 44, 1269–1273. [Google Scholar] [CrossRef]
- Chan, Y.S.; Ong, C.W.; Chuah, B.L.; Khoo, K.S.; Chye, F.Y.; Sit, N.W. Antimicrobial, antiviral and cytotoxic activities of selected marine organisms collected from the coastal areas of Malaysia. J. Mar. Sci. Technol. 2018, 26, 128–136. [Google Scholar] [CrossRef]
- Phang, S.M. Potential products from tropical algae and seaweeds, especially with reference to Malaysia. Malays. J. Sci. 2010, 29, 160–166. [Google Scholar] [CrossRef]
- Nagappan, T.; Vairappan, C.S. Nutritional and bioactive properties of three edible species of green algae, genus Caulerpa (Caulerpaceae). J. Appl. Phycol. 2014, 26, 1019–1027. [Google Scholar] [CrossRef]
- de Gaillande, C.; Payri, C.; Remoissenet, G.; Zubia, M. Caulerpa consumption, nutritional value and farming in the Indo-Pacific region. J. Appl. Phycol. 2017, 29, 2249–2266. [Google Scholar] [CrossRef]
- Sarini, A.W.; Nor’ Aishah, H.; Zaini, N. Determination of antioxidant activity for seven types of macroalgae. In Proceedings of the 5th International Conference on Food Engineering and Biotechnology, Penang, Malaysia, 12–14 March 2014; IACSIT Press: Singapore, 2014. [Google Scholar]
- Chia, Y.Y.; Kanthimathi, M.S.; Khoo, K.S.; Rajarajeswaran, J.; Cheng, H.M.; Yap, W.S. Antioxidant and cytotoxic activities of three species of tropical seaweeds. BMC Complement. Altern. Med. 2015, 15, 339. [Google Scholar] [CrossRef]
- Khandhasamy, M.; Arunachalam, K.D. Evaluation of in vitro antibacterial property of seaweeds of southeast coast of India. Afr. J. Biotechnol. 2008, 7, 1958–1961. [Google Scholar]
- Radhika, D.; Veerabahu, C.; Priya, R. Antibacterial activity of some selected seaweeds from the Gulf of Mannar Coast, South India. Asian J. Pharm. Clin. Res. 2012, 5, 89–90. [Google Scholar]
- Nagaraj, S.R.; Osborne, J.W. Bioactive compounds from Caulerpa racemosa as a potent larvicidal and antibacterial agent. Front. Biol. 2014, 9, 300–305. [Google Scholar] [CrossRef]
- Salem, W.M.; Galal, H.; Nasr El-deen, F. Screening for antibacterial activities insome marine algae from the red sea (Hurghada, Egypt). Afr. J. Microbiol. Res. 2011, 5, 2160–2167. [Google Scholar] [CrossRef]
- Paul, N.A.; Neveux, N.; Magnusson, M.; de Nys, R. Comparative production and nutritional value of “sea grapes”–The tropical green seaweeds Caulerpa lentillifera and C. racemosa. J. Appl. Phycol. 2014, 26, 1833–1844. [Google Scholar] [CrossRef]
- Devasagayam, T.P.; Tilak, J.C.; Boloor, K.K.; Sane, K.S.; Ghaskadbi, S.S.; Lele, R.D. Free radicals and antioxidants in human health: Current status and future prospects. J. Assoc. Phys. India 2004, 52, 794–804. [Google Scholar]
- Jiang, J.; Xiong, Y.L. Natural antioxidants as food and feed additives to promote health benefits and quality of meat products: A review. Meat. Sci. 2016, 120, 107–117. [Google Scholar] [CrossRef]
- Wang, T.; Jónsdóttir, R.; Liu, H.; Gu, L.; Kristinsson, H.G.; Raghavan, S.; Ólafsdóttir, G. Antioxidant capacities of phlorotannins extracted from the brown algae Fucus vesiculosus. J. Agric. Food Chem. 2012, 60, 5874–5883. [Google Scholar] [CrossRef]
- Li, Y.X.; Wijesekara, I.; Li, Y.; Kim, S.K. Phlorotannins as bioactive agents from brown algae. Process. Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- Chew, Y.L.; Lim, Y.Y.; Omar, M.; Khoo, K.S. Antioxidant activity of three edible seaweeds from two areas in South East Asia. LWT–Food Sci. Technol. 2008, 41, 1067–1072. [Google Scholar] [CrossRef]
- Li, Z.; Wang, B.; Zhang, Q.; Qu, Y.; Xu, H.; Li, G. Preparation and antioxidant property of extract and semipurified fractions of Caulerpa racemosa. J. Appl. Phycol. 2012, 24, 1527–1536. [Google Scholar] [CrossRef]
- Salmi, N.A.; Shamsul, M.; Ibrahim, C.O.; Hasmah, A. Radical scavenging activity and total phenolic content of Gracilaria manilaensis. In Proceedings of the Technology, Science, Social Sciences and Humanities International Conference 2012, Langkawi, Kedah, Malaysia, 14–15 November 2012. [Google Scholar]
- Cho, M.; Lee, H.S.; Kang, I.J.; Won, M.H.; You, S.G. Antioxidant properties of extract and fractions from Enteromorpha prolifera, a type of green seaweed. Food Chem. 2011, 127, 999–1006. [Google Scholar] [CrossRef]
- Zakaria, N.A.; Ibrahim, D.; Sulaiman, S.F.; Supardy, N.A. Assessment of antioxidant activity, total phenolic content and in vitro toxicity of Malaysian red seaweed, Acanthophora spicifera. J. Chem. Pharm. Res. 2011, 3, 182–191. [Google Scholar]
- Chibi, F.; Rchid, H.; Arsalane, W.; Nmila, R. Antioxidant activity and total phenolic content of the red alga Halopitys incurvus harvested from El Jadida Coast (Morocco). Int. J. Pharm. Phytochem. Res. 2018, 10, 176–181. [Google Scholar]
- Osuna-Ruiz, I.; López-Saiz, C.-M.; Burgos-Hernández, A.; Velázquez, C.; Nieves-Soto, M.; Hurtado-Oliva, M.A. Antioxidant, antimutagenic and antiproliferative activities in selected seaweed species from Sinaloa, Mexico. Pharm. Biol. 2016, 54, 2196–2210. [Google Scholar] [CrossRef]
- Global Action Plan on Antimicrobial Resistance. Available online: https://www.who.int/antimicrobial-resistance/global-action-plan/en/ (accessed on 11 June 2019).
- Peng, B.; Wang, C.; Li, H.; Su, Y.B.; Ye, J.Z.; Yang, M.J.; Jiang, M.; Peng, X.-X. Outer membrane proteins form specific patterns in antibiotic-resistant Edwardsiella tarda. Front. Microbiol. 2017, 8, 69. [Google Scholar] [CrossRef]
- Lin, X.M.; Yang, M.J.; Li, H.; Wang, C.; Peng, X.X. Decreased expression of LamB and Odp1 complex is crucial for antibiotic resistance in Escherichia coli. J. Proteom. 2014, 98, 244–253. [Google Scholar] [CrossRef]
- Al-Saif, S.S.A.; Abdel-Raouf, N.; El-Wazanani, H.A.; Aref, I.A. Antibacterial substances from marine algae isolated from Jedah coast of Red sea, Saudi Arabia. Saudi J. Biol. Sci. 2014, 21, 57–64. [Google Scholar] [CrossRef]
- Gao, X.; Liu, Y.; Deeb, D.; Arbab, A.S.; Gautam, S.C. Anticancer activity of pristimerin in ovarian carcinoma cells is mediated through the inhibition of prosurvival Akt/NF-κB/mTOR signaling. J. Exp. Ther. Oncol. 2014, 10, 275–283. [Google Scholar]
- Sheng, Q.; Fang, X.; Zhu, Z.; Xiao, W.; Wang, Z.; Ding, G.; Zhao, L.; Li, Y.; Yu, P.; Ding, Z.; et al. Seasonal variation of pheophorbide a and flavonoid in different organs of two Carpinus species and its correlation with immunosuppressive activity. In Vitro Cell Dev. Biol. Anim. 2016, 52, 654–661. [Google Scholar] [CrossRef]
- Stermitz, F.R.; Tawara-Matsuda, J.; Lorenz, P.; Mueller, P.; Zenewicz, L.; Lewis, K. 5′-Methoxyhydnocarpin-D and Pheophorbide A: Berberis species components that potentiate berberine growth inhibition of resistant Staphylococcus aureus. J. Nat. Prod. 2000, 63, 1146–1149. [Google Scholar] [CrossRef]
- Guerriero, A.; Meinesz, A.; D’Ambrosio, M.; Pietra, F. Isolation of toxic and potentially toxic sesqui-and monoterpenes from the tropical green seaweed Caulerpa taxifolia which has invaded the region of Cap Martin and Monaco. Helvetica 1992, 75, 689–695. [Google Scholar] [CrossRef]
- Smyrniotopoulos, V.; Abatis, D.; Tziveleka, L.-A.; Tsitsimpikou, C.; Roussis, V.; Loukis, A.; Vagias, C. Acetylene sesquiterpenoid esters from the green alga Caulerpa racemosa. J. Nat. Prod. 2003, 66, 21–24. [Google Scholar] [CrossRef]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef]
- Huang, H.L.; Wang, B.G. Antioxidant capacity and lipophilic content of seaweeds collected from the Qingdao coastline. J. Agric. Food Chem. 2004, 52, 4993–4997. [Google Scholar] [CrossRef]
- Ismail, A.; Ktari, L.; Romdhane, Y.B.R.; Aoun, B.; Sadok, S.; Boudabous, A.; El Bour, M. Antimicrobial fatty acids from green alga Ulva rigida (Chlorophyta). BioMed Res. Int. 2018, 2018, 3069595. [Google Scholar] [CrossRef]
- Desbois, A.P.; Lawlor, K.C. Antibacterial activity of long-chain polyunsaturated fatty acids against Propionibacterium acnes and Staphylococcus aureus. Mar. Drugs 2013, 11, 4544–4557. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.J. Antibacterial free fatty acids and monoglycerides: Biological activities, experimental testing, and therapeutic applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef]
- Das, U.N. Arachidonic acid and other unsaturated fatty acids and some of their metabolites function as endogenous antimicrobial molecules: A review. J. Adv. Res. 2018, 11, 57–66. [Google Scholar] [CrossRef]
- Hamzah, A.; Kipli, S.H.; Ismail, S.R. Microbiological study in coastal water of Port Dickson, Malaysia. Sains Malays. 2011, 40, 93–99. [Google Scholar]
- Plaza, A.; Keffer, J.L.; Bifulco, G.; Llyod, J.R.; Bewley, C.A. Chrysophaentins A-H, antibacterial bisdiarylbutene macrocycles that inhibit the bacterial cell division protein FtsZ. J. Am. Chem. Soc. 2010, 132, 9069–9077. [Google Scholar] [CrossRef]
- Lane, A.L.; Stout, E.P.; Lin, A.S.; Prudhomme, J.; Roch, K.L.; Fairchild, C.R.; Franzblau, S.G.; Hay, M.E.; Aalbersberg, W.; Kubanek, J. Antimalarial bromophycolides J-Q from the Fijian red alga Callophycus serratus. J. Org. Chem. 2009, 74, 2736–2742. [Google Scholar] [CrossRef]
- Klevens, R.M.; Morrison, M.A.; Nadle, J.; Petit, S.; Gershman, K.; Ray, S.; Harrison, L.H.; Lynfield, R.; Dumyati, G.; Townes, J.M.; et al. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 2007, 298, 1763–1771. [Google Scholar] [CrossRef]
- Mera, R.M.; Suaya, J.A.; Amrine-Madsen, H.; Hogea, C.S.; Miller, L.A.; Lu, E.P.; Sahm, D.F.; O’Hara, P.; Acosta, C.J. Increasing role of Staphylococcus aureus and community-acquired methicillin-resistant Staphylococcus aureus infections in the United States: A 10-year trend of replacement and expansion. Microb. Drug Resist. 2011, 17, 321–328. [Google Scholar] [CrossRef]
- Lee, C.H.; Wang, M.C.; Huang, I.W.; Chen, F.J.; Lauderdale, T.L. Development of daptomycin nonsusceptibility with heterogeneous vancomycin-intermediate resistance and oxacillin susceptibility in methicillin-resistant Staphylococcus aureus during high-dose daptomycin treatment. Antimicrob. Agents Chemother. 2010, 54, 4038–4040. [Google Scholar] [CrossRef]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar]
- Szymanek-Majchrzak, K.; MLynarczyk, A.; MLynarczyk, G. Characteristics of glycopeptide-resistant Staphylococcus aureus strains isolated from inpatients of three teaching hospitals in Warsaw, Poland. Antimicrob. Resist. Infect. Control 2018, 7, 105. [Google Scholar] [CrossRef]
- Adeyankinnu, F.A.; Motayo, B.O.; Akinduti, A.; Akinbo, J.; Ogiogwa, J.I.; Aboderin, B.W.; Agunlejika, R.A. A multicenter study of beta-lactamase resistant Escherichia coli and Klebsiella pneumoniae reveals high level chromosome mediated extended spectrum β lactamase resistance in Ogun State, Nigeria. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 819896. [Google Scholar] [CrossRef]
- Liang, W.J.; Liu, H.Y.; Duan, G.C.; Zhao, Y.X.; Chen, S.Y.; Yang, H.Y.; Xi, Y.L. Emergence and mechanism of carbapenem-resistant Escherichia coli in Henan, China, 2014. J. Infect. Public Health 2018, 11, 347–351. [Google Scholar] [CrossRef]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar]
- Moubayed, N.M.S.; Al Houri, H.J.; Al Khulaifi, M.M.; Farraj, D.A. Antimicrobial, antioxidant properties and chemical composition of seaweeds collected from Saudi Arabia (Red Sea and Arabian Gulf). Saudi J. Biol. Sci. 2016, 24, 162–169. [Google Scholar] [CrossRef]
- Airanthi, M.K.W.A.; Hosokawa, M.; Miyashita, K. Comparative antioxidant activity of edible Japanese brown seaweeds. J. Food Sci. 2011, 76, 104–111. [Google Scholar] [CrossRef]
- Chakraborty, K.; Raola, V.K. In vitro bioactive analysis and antioxidant activity of two species of seaweeds from the Gulf of Mannar. Nat. Prod Res. 2018, 32, 2729–2734. [Google Scholar] [CrossRef]
- Egan, S.; Harder, T.; Burke, C.; Steinberg, P.; Kjelleberg, S.; Thomas, T. The seaweed holobiont: Understanding seaweed-bacteria interactions. FEMS Microbiol. Rev. 2013, 37, 462–476. [Google Scholar] [CrossRef]
- Ding, L.; Ma, Y.; Huang, B.; Chen, S. Effects of seawater salinity and temperature on growth and pigment contents in Hypnea cervicornis J. Agardh (Gigartinales, Rhodophyta). BioMed Res. Int. 2013, 2018, 594308. [Google Scholar]
- Singh, R.P.; Reddy, C.R. Seaweed-microbial interactions: Key functions of seaweed-associated bacteria. FEMS Microbiol. Ecol. 2014, 88, 213–230. [Google Scholar] [CrossRef]
- Ismail, A.; Ktari, L.; Ahmed, M.; Bolhuis, H.; Boudabbous, A.; Stal, L.J.; Cretoiu, M.S.; El Bour, M. Antimicrobial activities of bacteria associated with the brown alga Padina pavonica. Front. Microbiol. 2016, 7, 1072. [Google Scholar] [CrossRef]
- Sujuliyani; Thaib, E.A.; Indriati, N.; Liananda, M. Antibacterial activity of the symbiotic bacteria of green algae Caulerpa racemosa from Pulau Lima Indonesia. In Proceedings of the 3rd EMBRIO International Workshop on Marine Biodiversity: Understanding, Utilization, Conversation, Bogor, Indonesia, 9–10 October 2018; IOP Publishing Ltd.: Bristol, UK, 2019. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Packer, L., Ed.; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Pękal, A.; Pyrzynska, K. Evaluation of aluminium complexation reaction for flavonoid content assay. Food Anal. Methods 2014, 7, 1776–1782. [Google Scholar] [CrossRef]
- Pang, J.R.; Goh, V.M.J.; Tan, C.Y.; Phang, S.M.; Wong, K.H.; Yow, Y.Y. Neuritogenic and in vitro antioxidant activities of Malaysian Gracilaria manilaensis Yamamoto & Trono. J. Appl. Phycol. 2018, 30, 3253–3260. [Google Scholar]
- Lee, S.; Duce, I.; Atkins, H.; Khan, N.A. Cockroaches and locusts: Physicians’ answer to infectious diseases. Int. J. Antimicrob. Agents 2010, 37, 270–280. [Google Scholar] [CrossRef]
- Khan, N.A.; Osman, K.; Goldsworthy, G.J. Lysates of Locusta migratoria brain exhibit potent broad-spectrum antibacterial activity. J. Antimicrob. Chemother. 2008, 62, 634–635. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Seaweed Species | Extract | Mass of Extract Obtained (g) | Yield (%) |
|---|---|---|---|
| Chloroform | 0.053 ± 0.001 | 5.212 ± 0.106 a | |
| C. racemosa | Methanol | 0.126 ± 0.004 | 12.554 ± 0.434 b |
| Water | 0.133 ± 0.002 | 12.920 ± 0.289 b | |
| Chloroform | 0.052 ± 0.005 | 5.081 ± 0.462 a | |
| C. lentillifera | Methanol | 0.138 ± 0.015 | 13.450 ± 1.429 b |
| Water | 0.319 ± 0.020 | 30.884 ± 2.006 c |
| Seaweed Species | Extract | TPC (mg GAE/g) | TFC (mg QE/g) |
|---|---|---|---|
| Chloroform | 13.41 ± 0.86 a | 5.46 ± 0.41 a,c,e | |
| C. racemosa | Methanol | 10.33 ± 0.02 b | 24.52 ± 2.17 b |
| Water | 1.74 ± 0.09 c | 2.50 ± 0.10 a,c | |
| Chloroform | 5.47 ± 0.75 d | 0.28 ± 0.03 d | |
| C. lentillifera | Methanol | 4.52 ± 0.42 d | 4.93 ± 0.27 a,e |
| Water | 2.04 ± 0.36 c | 1.17 ± 0.03 f |
| Standard and Seaweed Species | Control and Extract | EC50 (mg/mL) |
|---|---|---|
| Standard | Ascorbic Acid | 0.01 ± 0.0005 a |
| Chloroform | 0.65 ± 0.03 d | |
| C. racemosa | Methanol | 2.51 ± 0.09 c,g |
| Water | 7.46 ± 0.20 b,f | |
| Chloroform | 2.20 ± 0.10 c,g | |
| C. lentillifera | Methanol | 9.74 ± 0.59 b,f |
| Water | 81.55 ± 4.22 e |
| Seaweed Extracts and Controls | Antibacterial Effect (%) | |
|---|---|---|
| MRSA | E. coli K1 | |
| C. racemosa Extract | ||
| Chloroform | 97.70 ± 0.30 * | 19.90 ± 4.05 |
| Methanol | 61.54 ± 2.19 *** | 42.91 ± 7.75 |
| Water | −237.79 ± 18.62 *** | −37.00 ± 2.86 ** |
| C. lentillifera Extract | ||
| Chloroform | 62.17 ± 6.60 * | 12.42 ± 3.83 |
| Methanol | 44.99 ± 2.42 ** | 42.26 ± 12.67 * |
| Water | −34.58 ± 2.71 | −43.05 ± 0.99 *** |
| Controls | ||
| Pen-strep (10 U) | 98.10 ± 0.10 | 100.00 ± 0.0 |
| DMSO (0.5%) | 15.90 ± 1.70 | 10.90 ± 3.70 |
| Methanol (0.5%) | 8.70 ± 1.56 | 11.80 ± 3.00 |
| Water (0.5%) | −28.30 ± 5.68 | −9.72 ± 0.92 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yap, W.-F.; Tay, V.; Tan, S.-H.; Yow, Y.-Y.; Chew, J. Decoding Antioxidant and Antibacterial Potentials of Malaysian Green Seaweeds: Caulerpa racemosa and Caulerpa lentillifera. Antibiotics 2019, 8, 152. https://doi.org/10.3390/antibiotics8030152
Yap W-F, Tay V, Tan S-H, Yow Y-Y, Chew J. Decoding Antioxidant and Antibacterial Potentials of Malaysian Green Seaweeds: Caulerpa racemosa and Caulerpa lentillifera. Antibiotics. 2019; 8(3):152. https://doi.org/10.3390/antibiotics8030152
Chicago/Turabian StyleYap, Wing-Fai, Vangene Tay, Sie-Hui Tan, Yoon-Yen Yow, and Jactty Chew. 2019. "Decoding Antioxidant and Antibacterial Potentials of Malaysian Green Seaweeds: Caulerpa racemosa and Caulerpa lentillifera" Antibiotics 8, no. 3: 152. https://doi.org/10.3390/antibiotics8030152
APA StyleYap, W.-F., Tay, V., Tan, S.-H., Yow, Y.-Y., & Chew, J. (2019). Decoding Antioxidant and Antibacterial Potentials of Malaysian Green Seaweeds: Caulerpa racemosa and Caulerpa lentillifera. Antibiotics, 8(3), 152. https://doi.org/10.3390/antibiotics8030152