1. Introduction
The interaction of engineered nanoparticles (ENPs) with cells is influenced by a corona of macromolecules that deposit on the ENP surface from the surrounding biological fluid. What macromolecules (often proteins) adhere to the ENP depends on the properties of the macromolecules and on the ENP surface structure, charge, hydrophobicity, and geometry [
1,
2,
3,
4]. Corona components may provide dominant features controlling the interaction of ENPs with specific cell surface binding sites, often followed by ENP internalization and a subsequent response by the cells. Understanding what corona components are present on an ENP and how they interface with cells is thus important to provide rational approaches for promoting positive responses, such as targeted drug delivery, or mitigating negative responses, such as toxicity. However, understanding ENP coronas is challenging because the potential corona components in complex biological environments are diverse and the properties of ENP surfaces vary widely. Single-walled carbon nanotubes (SWNTs) and multi-walled carbon nanotubes (MWNTs) are ENPs whose production is increasing due to a wide variety of commercial applications [
5,
6,
7,
8]. Nevertheless, there is ample evidence that carbon nanotubes can be toxic to organisms and the environment, but how their coronas contribute to toxicity is not well understood [
9,
10,
11].
We previously noted that carboxylated MWNTs (cMWNTs) coated with Pluronic
® F-108 (PF108) preferentially bind to and are accumulated by cells, whereas PF108-coated pristine MWNTs (pMWNTs) do not bind and are poorly accumulated [
12]. This suggested that surface receptors on macrophages selectively bind cMWNTs but not pMWNTs. Class A scavenger receptors (SR-As) are membrane glycoproteins that bind polyanionic compounds and modified proteins [
13,
14,
15], and several observations in the literature implicate SR-As as potential carbon nanotube receptors. For example, there is evidence that SWNTs coated with bovine serum albumin (BSA) are targeted to SR-As [
16]. There are also numerous reports where antagonists of class A-type 1 scavenger receptors (SR-A1s) affect cell responses to MWNTs: The accumulation of cMWNTs by RAW 264.7 macrophages correlated with the extent of carboxylation and was inhibited by the SR-A1 antagonist dextran sulfate [
17]; the rate of apoptosis induced by MWNTs could be reduced by treating the cells with poly I, another SR-A antagonist [
18]; and the accumulation of FITC-BSA-coated MWNTs by THP-1 macrophages was inhibited by the SR-A antagonist fucoidan [
19]. In addition, Hirano et al. found that MWNTs suspended in the surfactant Pluronic
® F-68 bind to MARCO (SR-A6) receptors on Chinese hamster ovary (CHO) cells overexpressing MARCO [
20]. We also observed that dextran sulfate reduced the binding of PF108-coated cMWNTs by macrophages [
12].
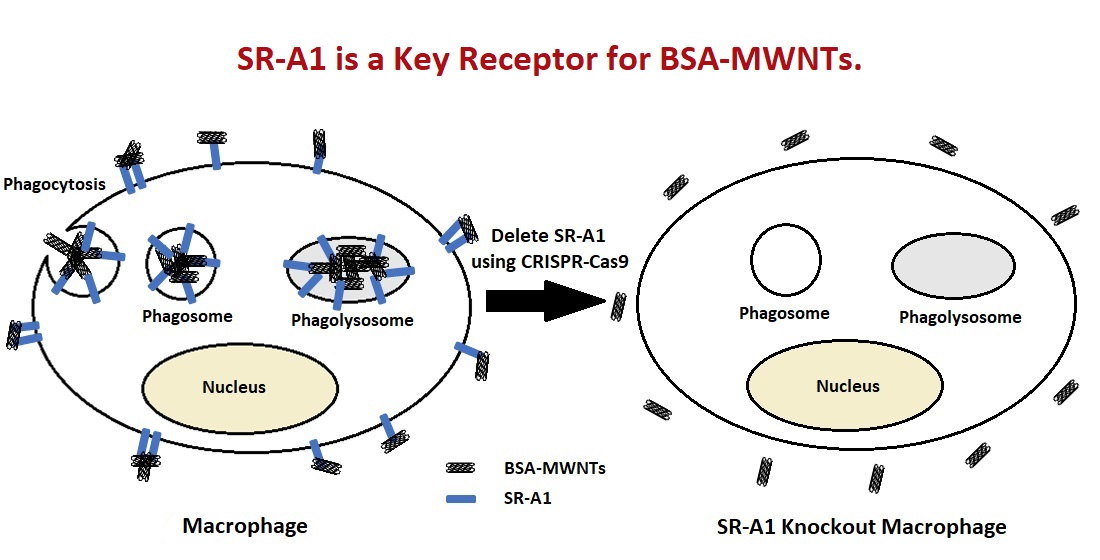
Recently, we reported that alveolar macrophages derived from SR-A1 knockout mice did not bind or accumulate PF108-cMWNTs whereas they were accumulated by CHO cells that ectopically expressed SR-A1 [
21]—strong evidence that SR-A1 is a receptor for PF108-coated cMWNTs. An interesting feature of PF108-coated cMWNTs is that they bind strongly to cells in the absence of serum or any exogenous protein, suggesting that a protein corona is not required for cMWNT binding to SR-A1 [
12]. Thus, some inherent structural feature of oxidized MWNTs, perhaps carboxyl groups, carbonyl groups, or hydroxyl groups, appear sufficient for interaction with SR-A1.
Herein, we replaced the PF108 coat with BSA and studied the interaction of cMWNTs and pMWNTs bearing a BSA corona with CHO cells that ectopically express SR-A1 and with RAW 264.7 cells that were deleted for SR-A1 using CRISPR-Cas9 technology. CHO cells expressing SR-A1, but not wild-type (WT) CHO cells, accumulated both BSA-coated cMWNTs and pMWNTs, but the amount of cMWNTs accumulated was 2–3 times more than pMWNTs. WT RAW 264.7 cells also accumulated approximately 2 times more BSA-coated cMWNTs than pMWNTs. Moreover, in binding studies with RAW 264.7 cells at 4 °C in the absence of serum, more BSA-cMWNTs than BSA-pMWNTs were bound. These data suggest that there are more binding sites on the RAW 264.7 cell surface for BSA-cMWNTs than BSA-pMWNTs. To assess what effect the absence of SR-A1 would have, the binding and accumulation of BSA-coated cMWNTs and pMWNTs to SR-A1 knockout RAW 264.7 cells at 4 °C in medium without serum and at 37 °C was measured. The amount of bound or accumulated BSA-MWNTs in the knockout SR-A1 cells was significantly decreased for both BSA-pMWNTs and BSA-cMWNTs compared to the WT RAW 264.7 cells. These observations suggest that pMWNTs coated with a BSA protein corona gain the capacity to bind SR-A1. Overall, BSA-cMWNTs have enhanced binding to SR-A1 above that observed with BSA-pMWNTs, emphasizing the differences between how BSA-coated cMWNTs and pMWNTs interact with receptors. Models to account for the differences are presented.
2. Materials and Methods
2.1. Nanomaterials
The pMWNT (product 1236-YJS, lot 2015-041709) and cMWNT (product 1256-YJF, lot 2015-070510) powders were purchased from Nanostructured & Amorphous Materials, Inc. (Houston, TX). pMWNTs and cMWNTs were synthesized using a Fe/Co/Ni-catalyzed chemical vapor deposition process. Caution should be taken, and a fine particulate respirator and other appropriate personal protective equipment should be worn when handling dry MWNT powders. Both MWNT products were reported by the manufacturer to be >95% in purity and to contain MWNTs with outer diameters of 10–20 nm, inner diameters of 5–10 nm, and lengths of 0.5–2 µm. The cMWNT powder was oxidized using sulfuric acid and potassium permanganate and comprised 1.9–2.1% by weight carboxylic acid groups. Elemental analyses of MWNTs were performed using a previously described combustion analysis technique [
22]. The combined carbon, hydrogen, nitrogen, sulfur, and oxygen elemental analyses of the pMWNTs and cMWNTs were 99.52% and 98.18%, respectively, indicative of MWNT powders that are essentially metal-free. An extensive physical and chemical characterization of the pMWNTs and cMWNTs powders appears elsewhere [
23]. The major similarities of the pMWNTs and cMWNTs were their outer diameters (18 ± 3 nm and 19 ± 5 nm, respectively) and inner diameters (5.6 ± 1.3 and 5.7 ± 1.7 nm, respectively), as determined using transmission electron microscopy. The key difference was the presence of a carbonyl vibrational stretching mode associated with carboxyl groups in the infrared spectra of cMWNTs that was not observed in the pMWNT spectra.
2.2. Chemicals and Solutions
Dulbecco’s modified Eagle medium (DMEM) and Ham’s F-12K complete medium were purchased from Gibco (Grand Island, NY, USA), fetal bovine serum (FBS) from Atlanta Biologicals (Flowery Branch, GA, USA), Geneticin® selective antibiotic G418 sulfate from Calbiochem (San Diego, CA, USA), and AccumaxTM from Innovative Cell Technologies (San Diego, CA, USA). SR-AI/MSR Alexa Fluor® 488-conjugated antibody and rat IgG2B Alexa Fluor® 488-conjugated Isotype Control were purchased from R&D Systems (Minneapolis, MN, USA). Bovine serum albumin (BSA), dextran sulfate (product # D6001), chondroitin sulfate (product # C9819), penicillin (10,000 U/mL), streptomycin (10 mg/mL), and all other chemicals were purchased from Millipore Sigma (Burlington, MA, USA). All chemicals were used as received. Deionized water (18.3 MΩ·cm) was obtained using a Milli-Q® Integral water purification system (Billerica, MA, USA). Phosphate buffered saline (PBS; 0.8 mM phosphate, 150 mM NaCl, pH 7.4) was sterilized by autoclaving at 121 °C for 45 min. Stock solutions of 100 mg/mL BSA were prepared by dissolving 10 g of BSA in 100 mL of deionized water and adjusting the pH to 7.4. Working solutions of 0.10 mg/mL BSA were prepared by diluting stock BSA solutions with aqueous 10 mM HEPES (pH 7.4) and filtering the solutions through a 0.22-μm pore membrane; stock and working solutions of BSA were stored at 4 °C in the dark.
2.3. Cell lines and Cell Culture
Abelson murine leukemia virus transformed RAW 264.7 macrophages were purchased from the American Type Culture Collection (ATCC® TIB-71™; Manassas, VA, USA). A scavenger receptor A1 (SR-A1) knockout RAW 264.7 cell pool was purchased from Synthego Corporation (Silicon Valley, CA, USA). The cell pool was generated using CRISPR-Cas9 technologies with the guide RNA sequence CAGCAUCCUCUCGUUCAUGA. Synthego validated, via genome sequencing, that 70% of the SR-A1 knockout pool of RAW 264.7 cells had insertion(s) or deletion(s) between base pairs 41 and 42 of the SR-A1 gene. Because the site of alteration is at the beginning of the gene, expression of SR-A1.1, which is a splice variant of SR-A1, would also be affected. A dilution scheme was used to clone cells that did not express SR-A1 receptors on their surface. Serial dilutions of the SR-A1 knockout RAW 264.7 cell pool were plated in 96-well plates and incubated for 7 days. Cells that had arisen from a single colony were grown for several passages before selecting clones that lacked surface SR-A1 expression using immunofluorescence microscopy and flow cytometry. All RAW 264.7 cells and SR-A1 knockout RAW 264.7 cells were grown in DMEM supplemented with 1.5 mg/mL sodium bicarbonate, 10 mM HEPES (pH 7.4), and 10% (v/v) FBS.
Chinese hamster ovary (CHO) cells stably transfected with mouse SR-A1 cDNA (CHO[mSR-AI] cells) were generously provided by Professor Monty Krieger (Massachusetts Institute of Technology) [
24]. The control WT CHO cell line for CHO[mSR-AI] cells were CHO-K1 cells (ATCC
® CCL-61™). All CHO cells were grown in Ham’s F-12K medium supplemented with 2.0 mg/mL sodium bicarbonate, 10 mM HEPES (pH 7.4), 10% (
v/v) FBS, 100 units/mL penicillin, and 100 μg/mL streptomycin; the mSR-AI cells were additionally maintained under 0.25 mg/mL G418. The standard incubation conditions for all cell lines were 37 °C in a 5% CO
2 and 95% air environment.
2.4. Preparation of BSA-MWNT Suspensions
The sonication and centrifugation protocol described in our previous works [
12,
25] was used with slight modifications to prepare purified BSA-coated MWNT suspensions, as summarized in
Scheme 1. MWNTs were coated with BSA to match the albumin in the FBS used in growth media. A total of 10.0 mg of pMWNT or cMWNT powder was weighed into a pre-cleaned 20-mL glass vial and baked at 200 °C for 2 h to inactivate potential endotoxin contaminants [
26]. Next, 10 mL of a 0.10 mg/mL BSA working solution was added to the vial and the mixture was sonicated. Specifically, a single vial was secured in a hanging rack and sonicated for 240 min using an ultrasonic bath sonicator (Elmasonic P30H; Elma Ultrasonic, Singen, Germany) that was operated at 120 W and 37 kHz in a 4 °C cold room. During sonication, the temperature of the bath water was maintained below 18 °C by using a refrigerated water bath circulator (Isotemp 1006S). After sonication, the solution was divided by transferring 1-mL aliquots into ten 1.5-mL centrifuge tubes. One of the 1-mL aliquots of each non-centrifuged BSA-pMWNT or BSA-cMWNT suspension was set aside as the standard suspension, and each standard solution was serially diluted with a 0.10 mg/mL-BSA working solution. The absorbance at 500 nm of the dilutions determined using a BioTek SynergyMx plate reader (Winooski, VT, USA) was used to construct pMWNT or cMWNT calibration curves. The remaining nine aliquots were centrifuged at 20,000 RCF for 5 min at 4 °C using an Eppendorf 5417R centrifuge to remove MWNT bundles and other impurities, as demonstrated in our previous work [
27]. The top 900 µL from each supernatant was collected without disturbing the pellet and combined in a sterile vial to afford ~9 mL of a purified BSA-pMWNT or BSA-cMWNT suspension. The concentration of MWNTs in each purified suspension was determined using the measured absorbance at 500 nm and the calibration curves described above. Purified BSA-MWNT suspensions were stored at 4 °C in the dark.
2.5. Characterization of MWNT Suspensions
The particle size distributions, in terms of hydrodynamic diameter, of BSA-MWNT suspensions were determined by dynamic light scattering (DLS). In brief, aliquots of purified pMWNT or cMWNT suspensions were diluted 1:10 in a 0.10 mg/mL BSA working solution and analyzed using a 633-nm laser and a backscatter measurement angle of 173° (Zetasizer Nano-ZS 3600, Malvern Instruments, Worcestershire, UK). The instrument was calibrated with Polybead® standards (Polysciences, Warrington, PA, USA) and ten consecutive 30-s runs were taken per measurement at 25 °C. The hydrodynamic diameter was calculated using a viscosity and refractive index of 0.8872 cP and 1.330, respectively, for deionized water, and an absorption and refractive index of 0.010 and 1.891, respectively, for MWNTs. Zeta potential values were also determined for purified BSA-coated MWNT suspensions that were diluted 1:10 with deionized water, medium with serum, or serum-free medium. In addition, DLS and zeta potential analyses were performed periodically on purified MWNT suspensions stored at 4 °C to detect any changes. Typically, MWNT suspensions were stable in storage for months, indicated by the lack of aggregates detected by DLS and constant zeta potential results.
2.6. Crystal Violet Cell Proliferation Assay
For the assays with RAW 264.7 cells, purified BSA-MWNT suspensions were first diluted with a freshly prepared 0.10 mg/mL-BSA working solution to a concentration twice the desired MWNT concentration to be tested. The diluted MWNT suspensions were then mixed 1:1 in equal volumes with 2X-concentrated medium that contained 3.0 mg/mL sodium bicarbonate, 20 mM HEPES (pH 7.4), 20% (
v/v) FBS, 200 units/mL penicillin, and 0.2 mg/mL streptomycin. The result is a test medium with the same concentration of 10 mM HEPES and 10% FBS as the control medium. A total of ~3.5 × 10
4 RAW 264.7 cells/well were seeded in 48-well plates and incubated at 37 °C overnight before the medium was replaced with freshly prepared control medium or test medium containing MWNTs and incubated for 24 h. At the end of the incubation, cells were washed 3 times with fresh medium, 2 times with PBS, air-dried, and fixed with 4% (
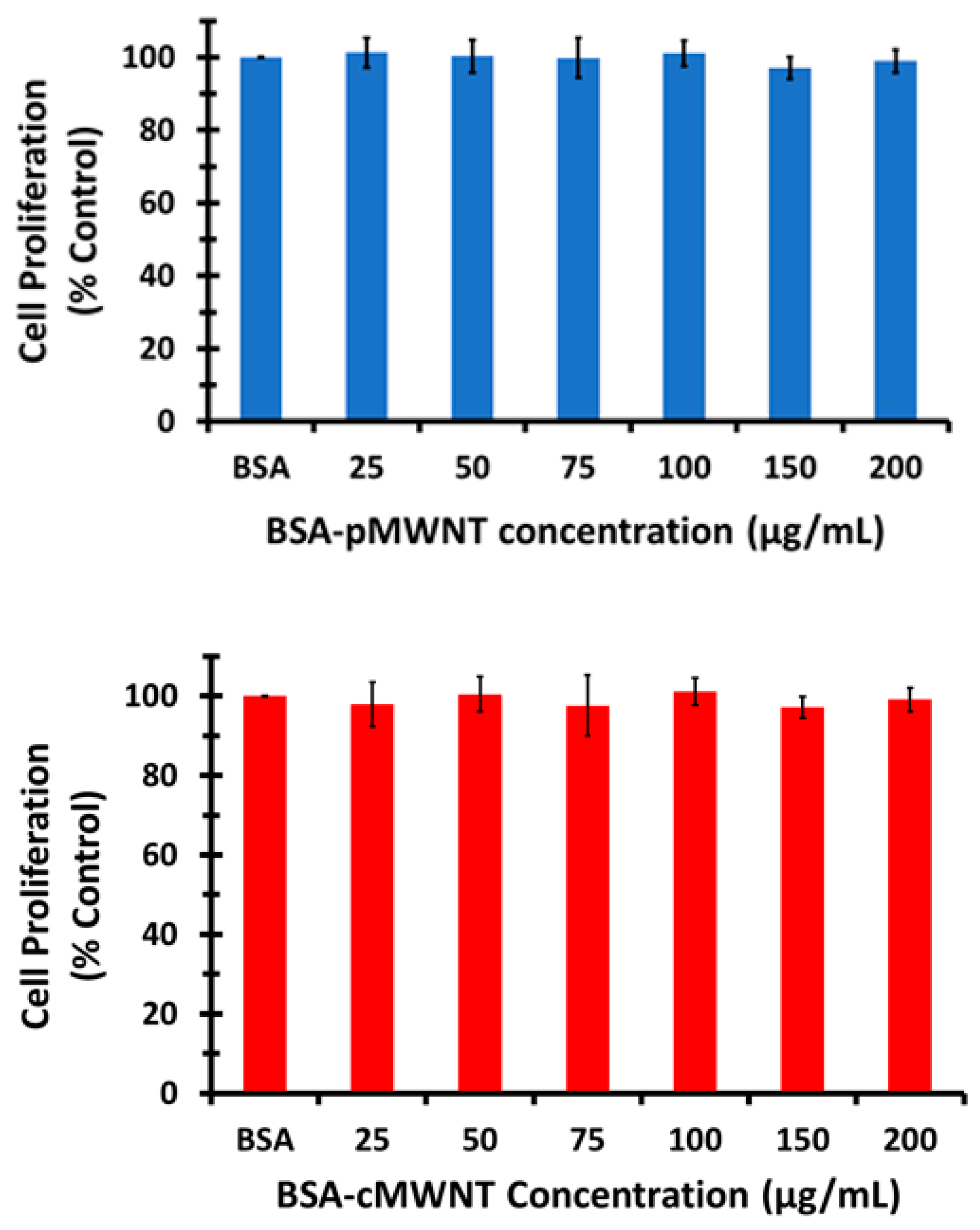
w/v) paraformaldehyde in PBS. Cell proliferation was determined using a standardized crystal violet assay, as described in our previous work where it was demonstrated that MWNTs do not interfere with the assay [
28].
2.7. Quantitation of MWNTs Extracted from Cell Lysates by SDS-PAGE
The SDS-PAGE method with optical detection [
29], previously validated by a large-area Raman scan technique [
12], was used for quantifying MWNTs extracted from RAW 264.7 cells or CHO cells. In brief, aliquots of known amounts of pMWNT or cMWNT standard suspensions, lysates of control cells, and lysates of cells treated with MWNTs were mixed with 5% 2-mercaptoethanol, 10% glycerol, 62.5 mM Tris-HCl, pH 6.2, and 2X-concentrated SDS sample loading buffer to a final concentration of 2% SDS, and boiled for 3 min. Samples at various dilutions and volumes were subsequently loaded into the wells of an SDS-polyacrylamide gel composed of a 4% stacking gel on top of a 10% resolving gel. An electric current was applied at a constant 100 V for 2 h. MWNTs in standard suspensions and in the lysates bind SDS in the sample loading buffer to become negatively charged and migrate toward the anode upon electrophoresis. The large aspect ratio of MWNTs prevents them from sieving through the pores of a 4% polyacrylamide gel mesh; thus, the MWNTs accumulate at the bottom of the sample loading well during electrophoresis and form a sharp dark band. Following electrophoresis, optical images of the gels were obtained using a flatbed scanner (HP Scanjet G3110, Hewlett Packard Enterprise, Fort Collin, CO, USA), and the pixel intensity of each dark band was quantified using ImageJ software (NIH ImageJ system, Bethesda, MD, USA). The known amount of MWNTs in the standards and their corresponding pixel intensities form a linear calibration curve that was used to determine the unknown amount of MWNTs in cell lysates, based on the pixel intensities of lysate bands loaded in the same gel as the standards.
2.8. Accumulation of MWNTs by Cells at 37 °C
The following procedure was used to detect the accumulation of pMWNTs and cMWNTs by RAW 264.7 or CHO cells at 37 °C for 24 h. MWNT suspensions were first diluted in a freshly prepared 0.10 mg/mL BSA working solution to twice the desired final MWNT concentrations specified in the experiment. The diluted MWNT suspension samples were then mixed 1:1 with the appropriate 2X-concentrated medium. A total of ~3.5 × 105 cells/well were seeded in 6-well plates and incubated in medium at 37 °C overnight to allow the cells to adhere to the plates. The medium was removed the next day and 2 mL of the appropriate freshly prepared control medium that contained no MWNTs or test medium that contained an MWNT suspension at a specified concentration was added to each well. Cells were incubated in a control or test medium at 37 °C for 24 h, as described in each experiment. At the end of the incubation, the control and test media were removed by aspiration and the cells were washed 3 times with fresh medium followed by 2 washes with PBS. Cells were then lifted off the well using 0.5 mL AccumaxTM, transferred to a centrifuge tube, and the well was rinsed with 1.5 mL PBS that was subsequently added to the tube to make a final cell suspension of 2 mL/well/tube. Three aliquots of cell suspension, 100 µL each, were used to determine cell counts in each sample using a Beckman Coulter particle counter (Miami, FL, USA) and the cells in the remaining 1.7-mL cell suspension were collected by centrifugation at 1000× g for 5 min at 4 °C. The cells in the pellet were lysed in 200 µL of cell lysis buffer that contained 0.25 M Tris-HCl (pH 6.8), 8% (w/v) SDS, and 20% (v/v) 2-mercaptoethanol. To ensure complete lysis of the cells, the lysate samples were heated in a boiling water bath for 2 h and then stored at 4 °C. The amounts of MWNTs in the cell lysate samples were determined using the SDS-PAGE method, as described previously herein.
2.9. Surface Binding of MWNTs to Cells at 4 °C
To detect and compare the association of pMWNTs and cMWNTs to the surface of RAW 264.7 cells in the absence of endocytic or phagocytic activity, ~5.0 × 105 RAW 264.7 cells/well were first seeded in 6-well plates and incubated in the appropriate medium at 37 °C overnight. Then, the cells were incubated in the appropriate serum-free medium for 2 h at 37 °C to deplete the serum in the cells. In order to incubate cells at a low temperature outside of the 37 °C incubator, the medium was replaced with the respective serum-free medium that additionally did not contain sodium bicarbonate. The 6-well plates were then placed on a shallow ice-water bath and incubated in a 4 °C cold room for 30 min. The appropriate 2X-concentrated, serum- and sodium bicarbonate-free medium was pre-chilled to 4 °C before mixing 1:1 with a MWNT suspension, such that the final test medium contained MWNTs at the desired concentration specified in the experiment. After chilling down to 4 °C, the cells were incubated for 1 h at 4 °C with the appropriate pre-chilled serum- and sodium bicarbonate-free medium that did not contain MWNTs (control), or test serum- and sodium bicarbonate-free medium that contained a MWNT suspension at the specified final MWNT concentration. Because phagocytosis and endocytosis are blocked at low temperature, MWNTs in the test medium were free to interact with cell surface components without subsequently entering the vacuolar compartment of the cells. After incubation, the cells were washed, harvested, and the subsequent procedures for cell counting and lysate preparation were followed, as described in the previous sections. The amounts of cell-surface bound MWNTs in the cell lysate samples were determined using the SDS-PAGE method, as described previously herein.
2.10. Dissociation of Bound BSA-cMWNTs and BSA-pMWNTs from RAW 264.7 Cells at 4 °C
MWNTs suspended in a 0.10 mg/mL BSA working solution were mixed with an equal volume of 2X-concentrated, serum- and sodium bicarbonate-free medium to give a final MWNT concentration of 100 µg/mL. Equivalent number of RAW 264.7 cells were seeded in 6-well plates and incubated at 37 °C under standard cell culture conditions for 24 h prior to the experiment. Next, the cells were pre-incubated with serum-free medium (in the absence of MWNTs) for 2 h at 37 °C to deplete the serum in the cells. The cells were then pre-chilled to 4 °C and incubated at 4 °C for 1 h in serum- and sodium bicarbonate-free medium that contained either BSA-pMWNTs or BSA-cMWNTs. Finally, the cells were then incubated with serum- and sodium bicarbonate-free medium for an additional 20, 40, 60, 90, or 120 min, and then washed 3 times with serum- and sodium bicarbonate-free medium, then 2 times with PBS. After incubation, surface-bound MWNTs were extracted and quantified by the SDS-PAGE method, as described previously herein.
2.11. Additive Binding Test for BSA-cMWNTs and BSA-pMWNTs to RAW 264.7 Cells
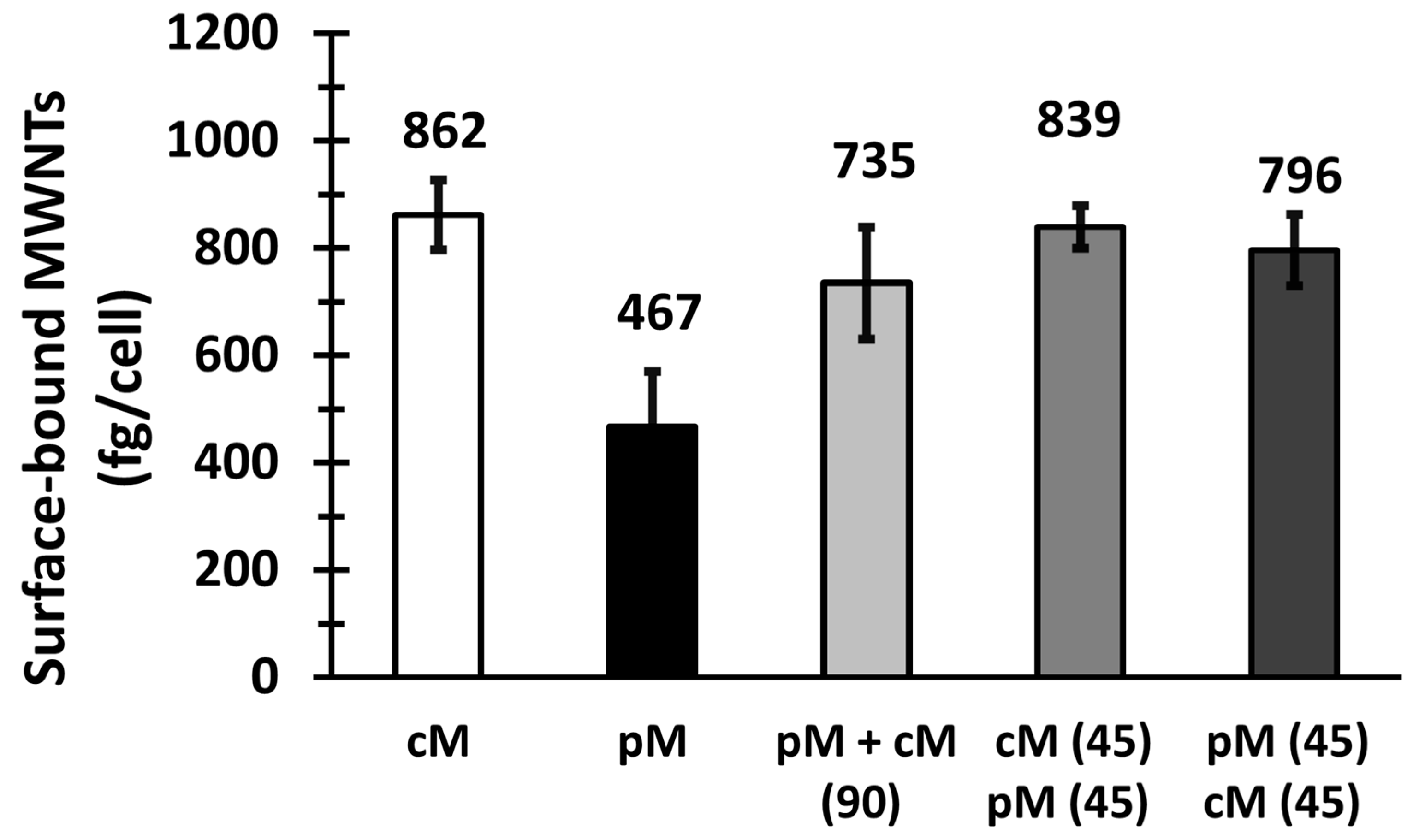
To determine whether BSA-cMWNTs and BSA-pMWNTs use independent surface binding sites, ~5.0 × 105 RAW 264.7 cells/well were first seeded in 6-well plates and incubated in medium at 37 °C overnight. Cells were then incubated in a serum-free medium for 2 h at 37 °C to deplete the serum in the cells. Next, this medium was replaced with a serum-free medium that did not contain sodium bicarbonate. The 6-well plates were placed on a shallow ice-water bath and incubated in a 4 °C cold room for 30 min. A 2X-concentrated, serum- and sodium bicarbonate-free medium was pre-chilled to 4 °C before mixing 1:1 with a MWNT suspension such that the final test serum- and sodium bicarbonate-free medium contained 100 μg/mL MWNTs. After chilling to 4 °C, the cells were incubated with either BSA-cMWNTs or BSA-pMWNTs separately at 4 °C for 90 min or simultaneously with both ligands at 4 °C for 90 min. In a slightly different experimental design, the ligands were added sequentially, first BSA-cMWNTs for 45 min at 4 °C followed by washing the cells and the addition of BSA-pMWNTs, for 45 min at 4 °C for a total incubation time of 90 minutes. The order of the ligand addition was then reversed with another set of cells. The amounts of cell-surface bound MWNTs in the cell lysate samples were determined using the SDS-PAGE method, as described previously herein.
2.12. Surface Binding of MWNTs to RAW 264.7 Cells in the Presence of Dextran Sulfate, an SR-A1 Antagonist
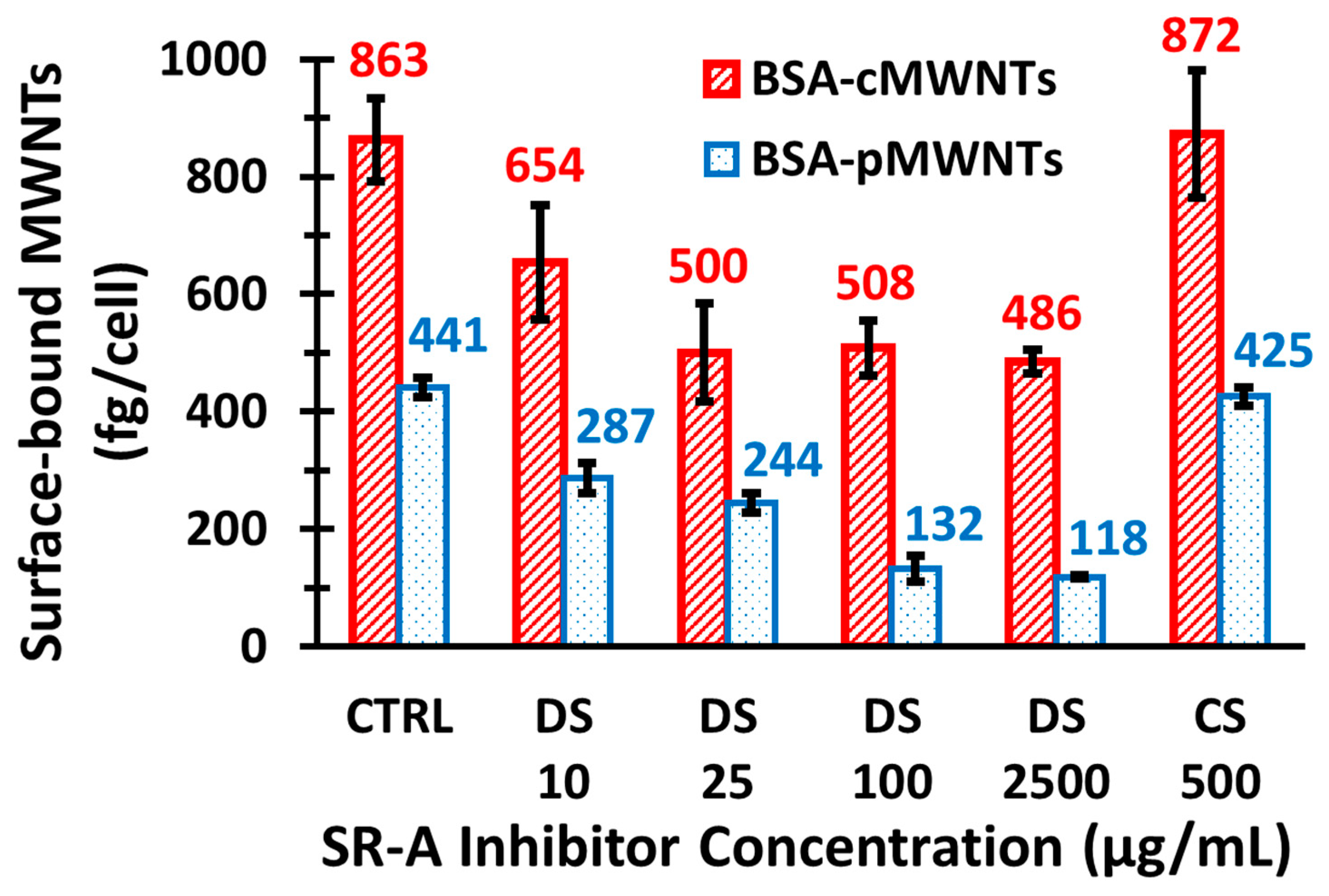
To determine the effects of dextran sulfate on the association of pMWNTs and cMWNTs to the surfaces of RAW 264.7 cells, ~5.0 × 105 RAW 264.7 cells/well were seeded in 6-well plates and incubated in medium at 37 °C overnight. Then, RAW 264.7 cells were incubated in serum-free medium for 2 h at 37 °C to deplete the serum in the cells. To incubate cells at low temperature outside of the 37 °C incubator, the serum-free medium was replaced with serum-free medium that did not contain sodium bicarbonate. The 6-well plates were then placed on a shallow ice-water bath and incubated in a 4 °C cold room for 30 min. A 2X-concentrated, serum- and sodium bicarbonate-free medium was pre-chilled to 4 °C before mixing 1:1 with a MWNT suspension followed by the addition of dextran sulfate (or chondroitin sulfate, a control that is not an SR-A1 antagonist) at various concentrations such that the final test serum- and sodium bicarbonate-free medium contained 100 μg/mL MWNTs. After chilling down to 4 °C, the cells were incubated for 1 h at 4 °C with test serum- and sodium bicarbonate-free medium that contained 100 μg/mL MWNTs, washed 3 times with serum- and sodium bicarbonate-free medium, and then washed 2 times with PBS. In all cases, the amounts of cell-surface bound MWNTs in the cell lysate samples were determined using the SDS-PAGE method, as described previously herein.
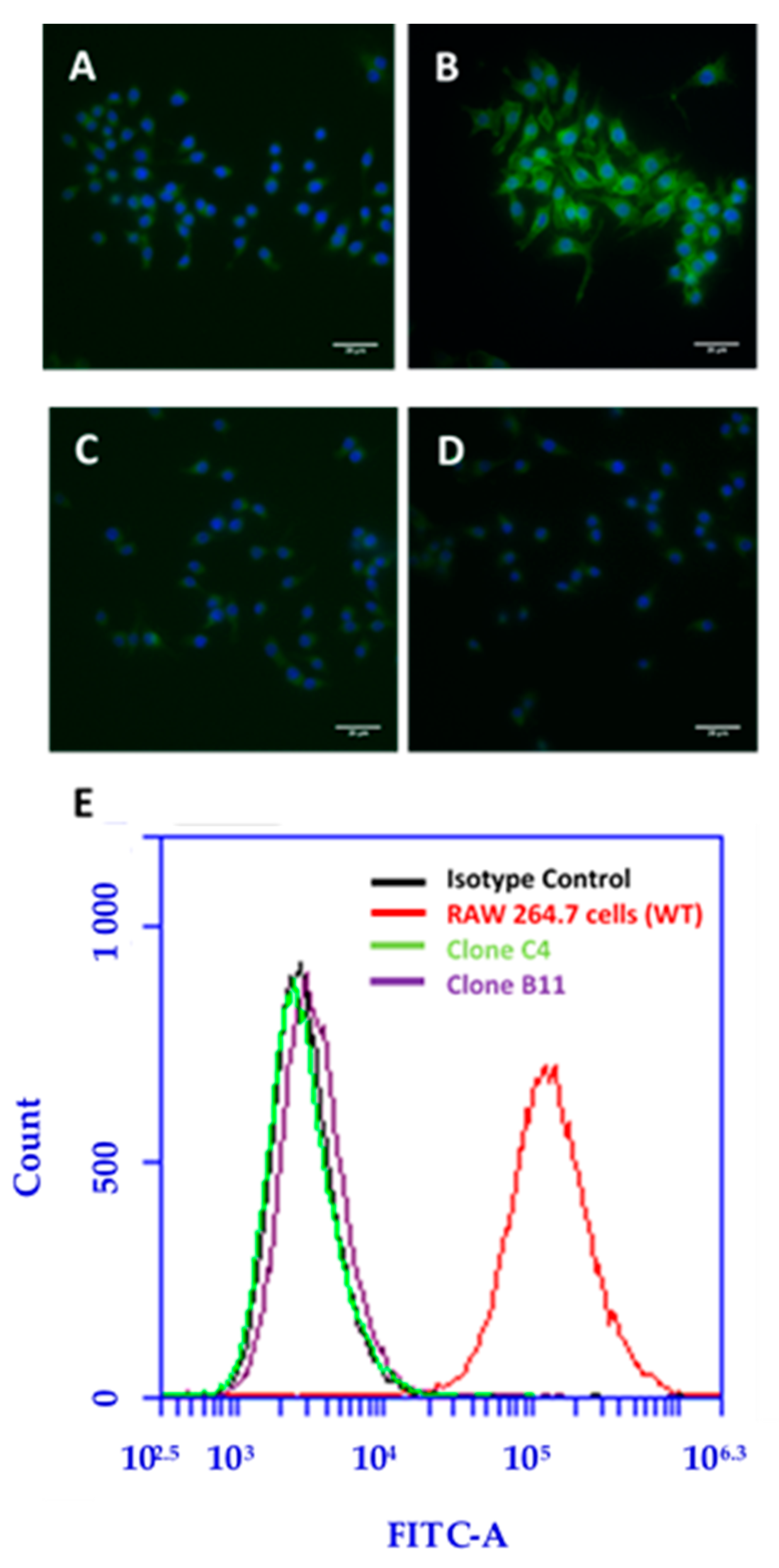
2.13. Immunofluorescence Microscopy of WT and SR-A1 Knockout RAW 264.7 Cells
A total of ~2 × 104 RAW 264.7 cells were seeded on coverslips in 4-well plates and incubated in medium at 37 °C for 48 h to allow the cells to adhere to the plates. RAW 264.7 cells were incubated in serum-free medium for 1 h at 37 °C to deplete the serum in the cells. The cells were washed three times with media and 2 times with PBS. Then the cells were fixed with 4% paraformaldehyde at room temperature for 20 min followed by washing with PBS. The cells were incubated in blocking buffer containing 4% fish gelatin in PBS at room temperature for 1 hour to block non-specific protein-protein interactions. The cells were incubated with mouse SR-AI/MSR Alexa Fluor® 488-conjugated antibody or a rat IgG2B Alexa Fluor® 488-conjugated monoclonal antibody as the isotype control at room temperature for 1 h in the dark; control cells were not treated with any antibody. After rinsing, cell nuclei were stained with Hoechst 33342 dye for 10 min at room temperature. Then the cells were washed two times with PBS to remove excess dye. The coverslips were mounted on the glass slide using Fluoromount-G™. Images were taken with a Nikon Eclipse TE-2000 fluorescence microscope using a 60× oil-immersion objective with a NA of 1.4; the images for Hoechst 33342 (Ex. 350 nm; Em. 435–485 nm) and Alexa Fluor® 488 (Ex. 488 nm; Em. 520–550 nm) were overlaid using ImageJ software.
2.14. Flow Cytometry for Surface Receptor(s) on WT and SR-A1 Knockout RAW 264.7 Cells
A total of ~2 × 106 RAW 264.7 cells were seeded in 10-mm plates and incubated in medium at 37 °C for 48 h to allow the cells to adhere to the plates. The cells were rinsed and harvested with warm FACS staining buffer (1% BSA in PBS) in 15 mL centrifuge tube followed by centrifugation (1000× g) for 5 min. The cells were suspended in 1 mL of FACS staining buffer, then three 100 µL-aliquots of the cell suspension were used to determine cell counts in each aliquot using a Beckman Coulter particle counter. A total of ~1 × 106 cells in 100 μL FACS staining buffer were aliquoted into 2 mL tubes. The cells were incubated in blocking buffer containing 5 μg IgG for 15 min at 4 °C to block non-specific protein interactions. The cells were stained with 5 μg mouse SR-AI/MSR Alexa Fluor® 488-conjugated antibody (R&D Systems cat. No. FAB1797G) or a rat IgG2B Alexa Fluor® 488-conjugated monoclonal antibody (R&D Systems cat. No. IC013G) as the isotype control for 30 min at 4 °C in the dark. Unbound antibody was removed by washing and re-suspending the cells in 1.5 mL FACS staining buffer thrice. The cells were re-suspended in 500 μL of FACS staining buffer for the final flow cytometric analysis. Flow cytometry analysis and data processing were performed using BD Accuri™ C6 Plus flow cytometer and CSampler™ Plus software (Becton and Dickinson Company, Franklin Lakes, NJ, USA) to determine the mean fluorescent index of each sample using a 518–548 nm emission filter.
4. Discussion
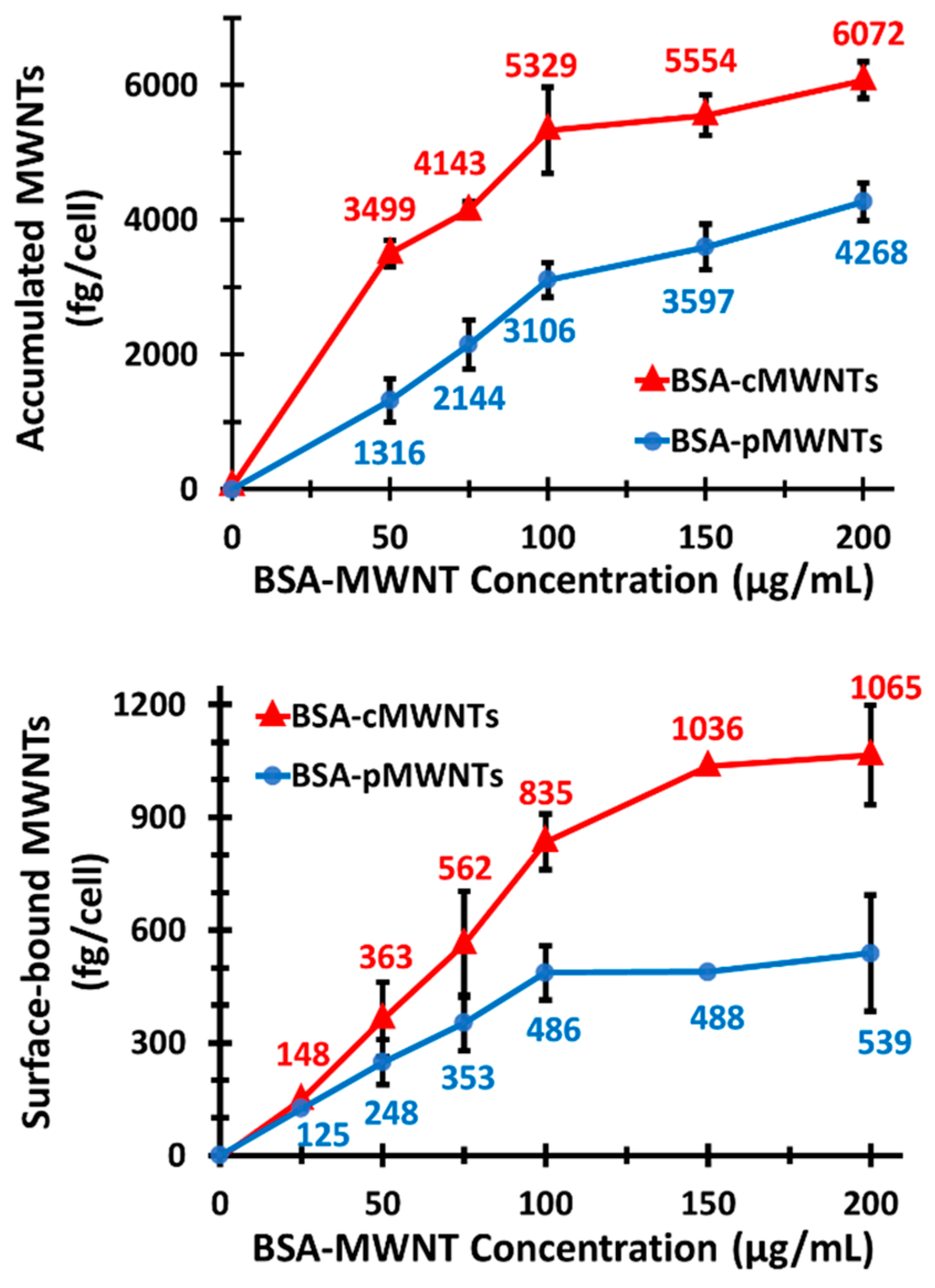
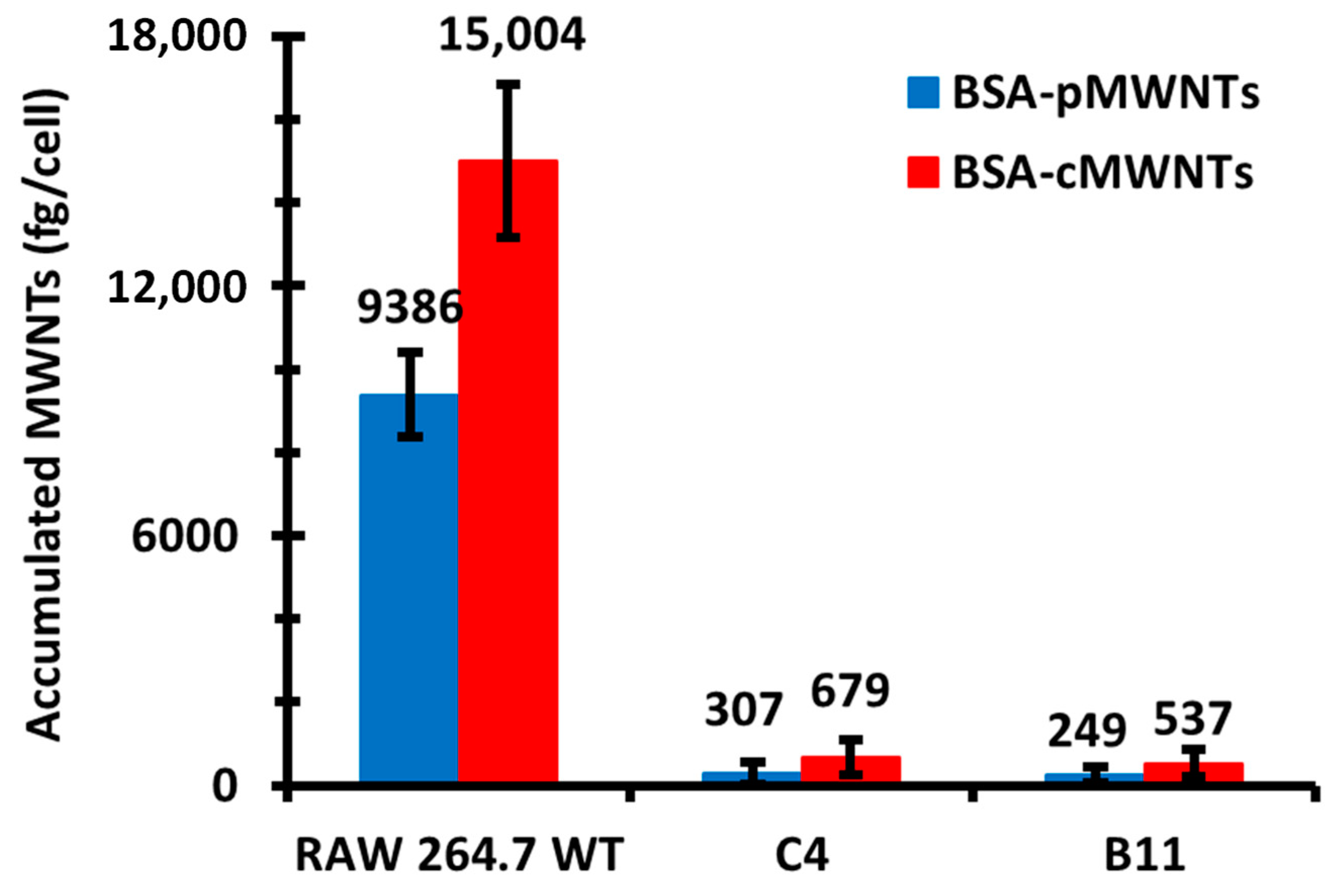
WT RAW 264.7 cells accumulated both BSA-cMWNTs and BSA-pMWNTs as a function of concentration after a 24 h exposure at 37 °C, although BSA-coated cMWNTs were accumulated to almost twice the amount of pMWNTs at each concentration tested. Uptake for both was near linear up to 100 µg/mL, after which the rate of accumulation was reduced. The break in the uptake curve at 100 µg/mL suggests a saturable receptor could be involved in the uptake process; however, accumulation depends not only on uptake, but also on potential loss of the MWNTs from cells by either recycling or degradation, or a loss of surface receptors that are internalized from the cell surface but not replaced. To focus on the initial interaction of MWNTs with cells, binding experiments were performed at 4 °C where internalization is inhibited. Moreover, serum proteins other than BSA that might confound the interpretation of the results were absent from the binding medium. Under these conditions, the binding of BSA-coated cMWNTs or pMWNTs to RAW 264.7 cells was near linear up to 100 µg/mL and then began to plateau, suggesting a saturable receptor-mediated binding event. There were two notable observations in comparing the binding of BSA-coated MWNTs to that we previously described for PF108-coated MWNTs. First, BSA-pMWNTs bound to cells, whereas previous studies showed that PF108-coated pMWNTs did not [
12,
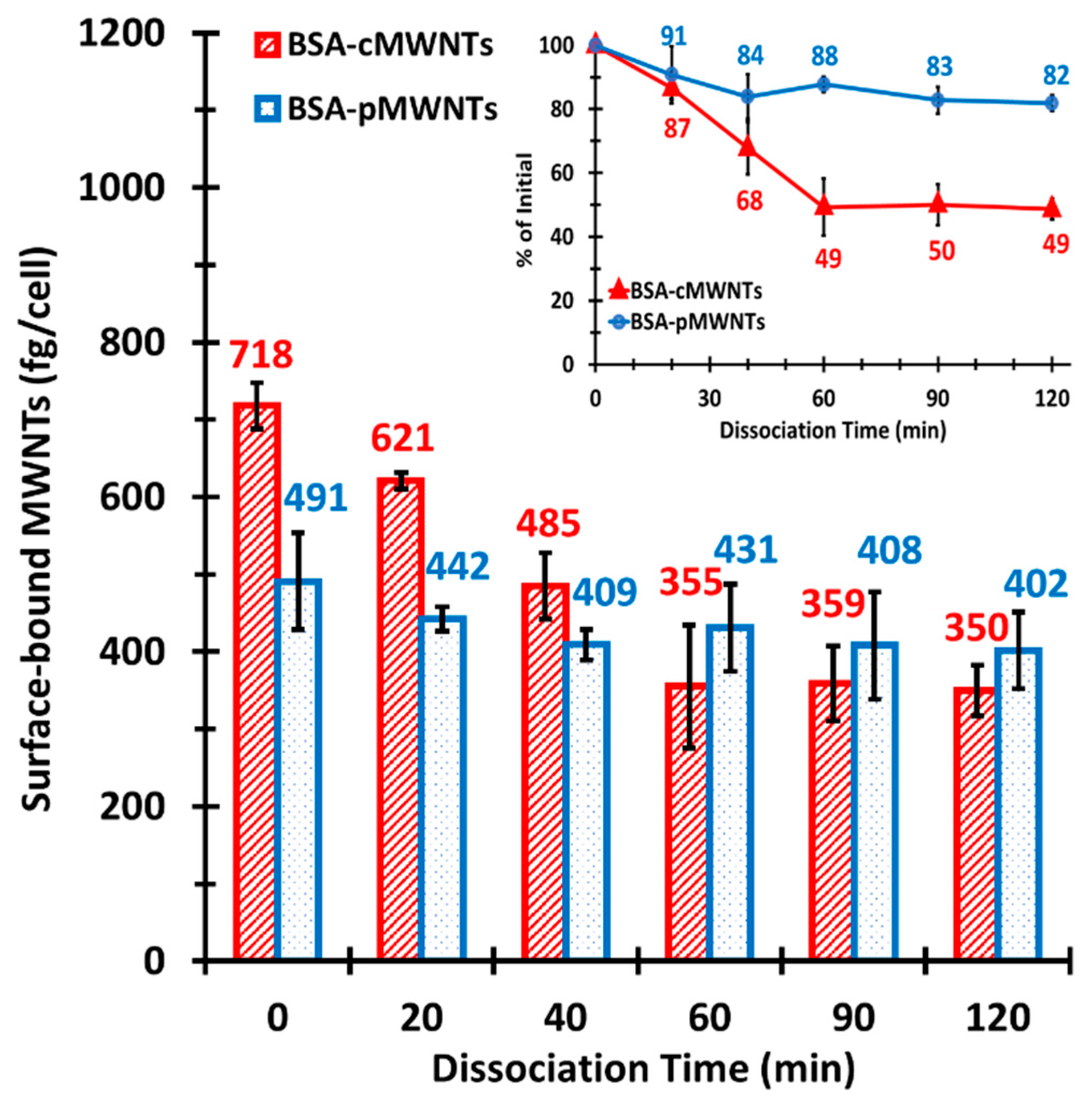
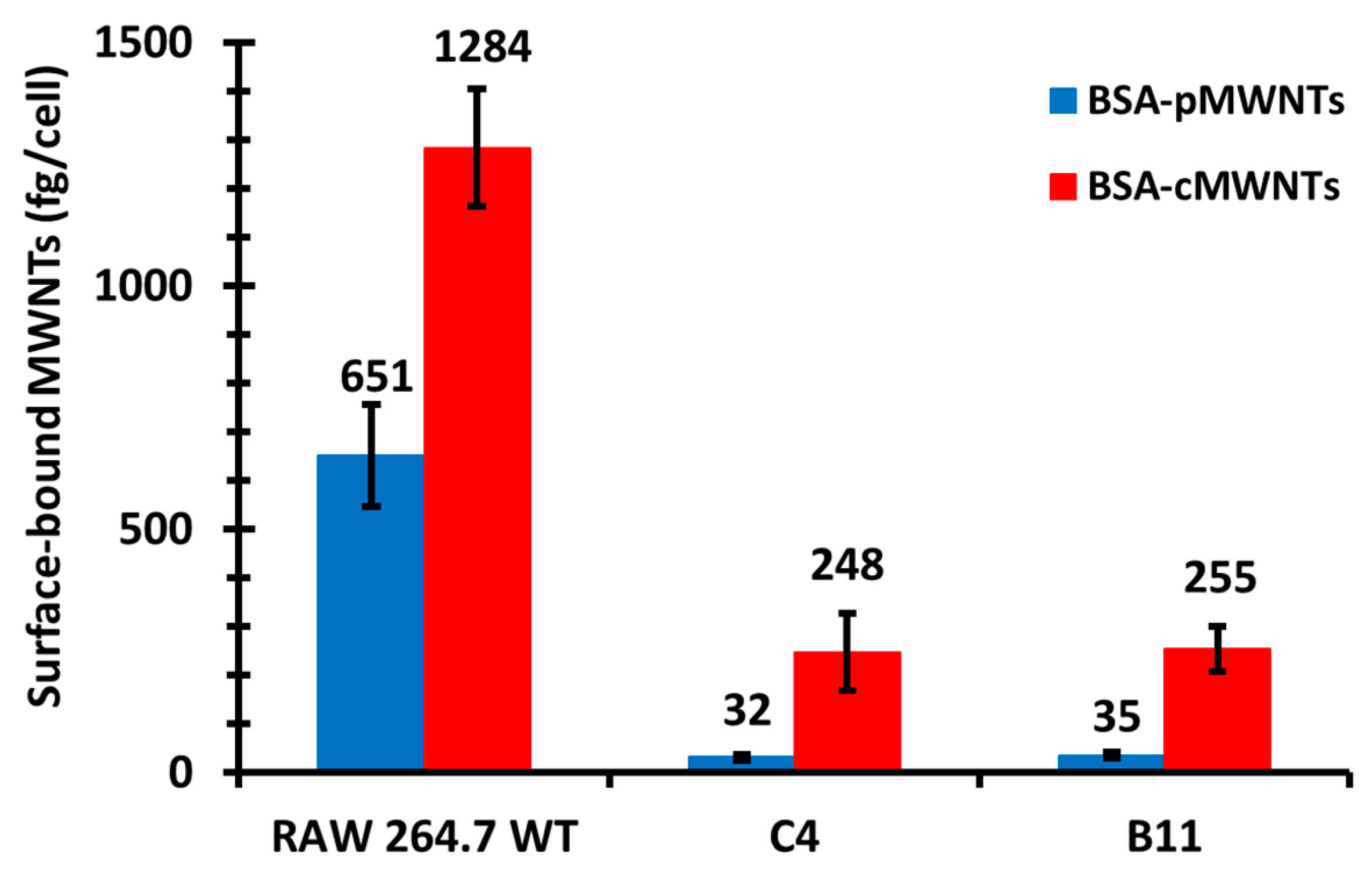
21]. This indicates that the BSA corona confers the ability of pMWNTs to bind cells. Second, the cells bound more BSA-cMWNTs than BSA-pMWNTs, evidence that there remains a difference in binding capacity between the two MWNT types. Differences between BSA-cMWNTs and BSA-pMWNTs were also seen in their kinetics of dissociation from cells: BSA-pMWNTs dissociated very slowly, whereas BSA-cMWNTs had a faster dissociating component followed by a slowly dissociating component.
One model to explain the difference in the binding of BSA-cMWNTs and BSA-pMWNTs to cells is that there are two independent receptors—one for each type of MWNT. If there are two receptors interacting independently with two ligands, then exposing cells simultaneously to both ligands should result in an amount bound that is the sum of both when added separately. However, this was not observed. The amount bound after simultaneous exposure to both BSA-cMWNTs and BSA-pMWNTs never exceed the amount bound to cells when BSA-cMWNTs were added alone, which is not a simple additive result. To further explore this issue, sequential binding experiments were undertaken. The level of cell-associated MWNTs when BSA-cMWNTs were added first, followed by BSA-pMWNTs, was equal to the amount of MWNTs bound when BSA-cMWNTs were added alone, which is not additive. However, when the order was reversed and BSA-pMWNTs were added first followed by BSA-cMWNTs, there was more binding than observed when BSA-pMWNTs were added alone, and the amount was again equal to the increased binding seen with BSA-cMWNTs alone, an additive result. Altogether, the results of the binding experiments suggest a semi-additive model: BSA-cMWNTs can occupy all the binding sites available to BSA-pMWNTs, plus additional sites not available to BSA-pMWNTs. Thus, when BSA-cMWNTs are added first, no binding of BSA-pMWNTs occurs because the sites are occupied by BSA-cMWNTs. However, when BSA-pMWNTs are added first, there remain sites available for BSA-cMWNTs to which BSA-pMWNTs cannot bind.
The semi-additive data are compatible with a two-receptor model and also with a model where a single receptor has two binding sites. In the two-receptor model, one receptor would bind both cMWNTs and pMWNTs, and the other receptor would bind only cMWNTs. To help address the question of whether one or two receptors were involved in binding cMWNTs and pMWNTs, the accumulation and binding of BSA-coated MWNTs was studied with RAW 264.7 cells in which the SR-A1 gene had been knocked out. Two clones isolated from the knockout pool, which were shown to lack immunologically detectable SR-A1 on their surfaces, failed to accumulate either BSA-coated cMWNTs or pMWNTs at 37 ℃. In binding studies at 4 ℃, the binding of BSA-pMWNTs was negligible and the binding of BSA-cMWNTs was reduced by 80%. It is not clear what is responsible for the 20% of BSA-cMWNT binding in the knockout cells, but perhaps one or more minor receptors for BSA-cMWNTs are present at low levels, and their contributions are seen in SR-A1 knockout cells. Nevertheless, it appears that knocking out SR-A1 severely affects the accumulation and binding of both BSA-cMWNTs and BSA-pMWNTs.
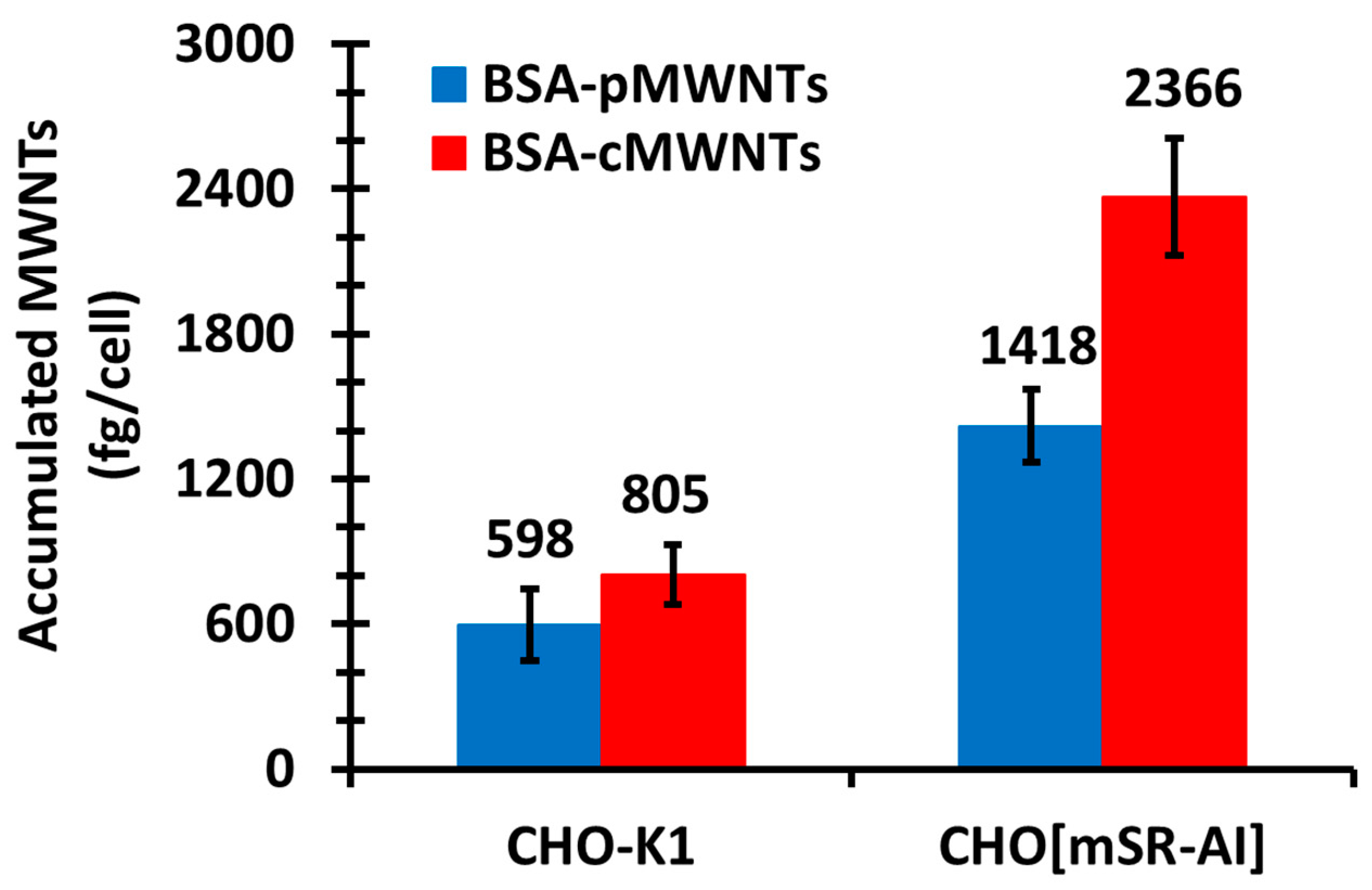
The simplest explanation for the knockout results is that SR-A1 is a receptor for both BSA-cMWNTs and BSA-pMWNTs. However, an alternative explanation is that knocking out SR-A1 suppresses the expression of one or more other cell surface proteins that could be major receptors for BSA-coated MWNTs. Two lines of evidence argue against this possibility. One is that dextran sulfate, a known antagonist of ligand binding to SR-A1, at least partially inhibited the binding of both BSA-coated pMWNTs and cMWNTs to cells, supporting the idea that SR-A1 is a receptor for these ligands. Second, CHO-K1 cells that ectopically express SR-A1 accumulated significantly more BSA-coated cMWNTs and pMWNTs than normal CHO-K1 cells. It seems unlikely that a covert receptor is activated in CHO cells, a cell type very different than RAW 264.7 macrophages, upon expression of SR-A1. Altogether, the simplest interpretation of the evidence argues that SR-A1 binds both BSA-cMWNTs and BSA-pMWNTs.
Understanding what features of BSA-coated MWNTs interact with SR-A1 is an interesting challenge. Previous work established that PF108-coated cMWNTs bound to and were accumulated by macrophages that expressed SR-A1 in the absence of serum or serum proteins [
12], whereas alveolar macrophages derived from mice knocked out for SR-A1 failed to accumulate the MWNTs [
21]. PF108-coated pMWNTs were not bound or accumulated by either SR-A1 positive or negative macrophages [
12,
21]. Thus, no protein corona was necessary for SR-A1 to interact with cMWNTs. This suggested that one or more oxidized functionalities intrinsic to cMWNTs (carboxyl, hydroxyl, phenolic, etc.) are structural features potentially recognized by SR-A1. SR-A1 access to cMWNT surface features might occur at nanotube ends where the high curvature may not support coat binding and where oxidized functionalities are often located due to ring strain [
37,
38,
39,
40]. In addition, the residence time of BSA on MWNTs appears to be short and not all the surface is covered with protein at one time [
41]. Thus, it is likely that SR-A1 would have access to oxidized groups intrinsic to the MWNT surface of BSA-coated cMWNTs.
It is understood now that while native BSA does not interact with SR-A1, conformational changes in BSA upon binding several types of nanoparticles uncover latent sites that do bind SR-A1 [
34,
35,
36]. Moreover, BSA undergoes significant conformation changes upon binding to cMWNTs [
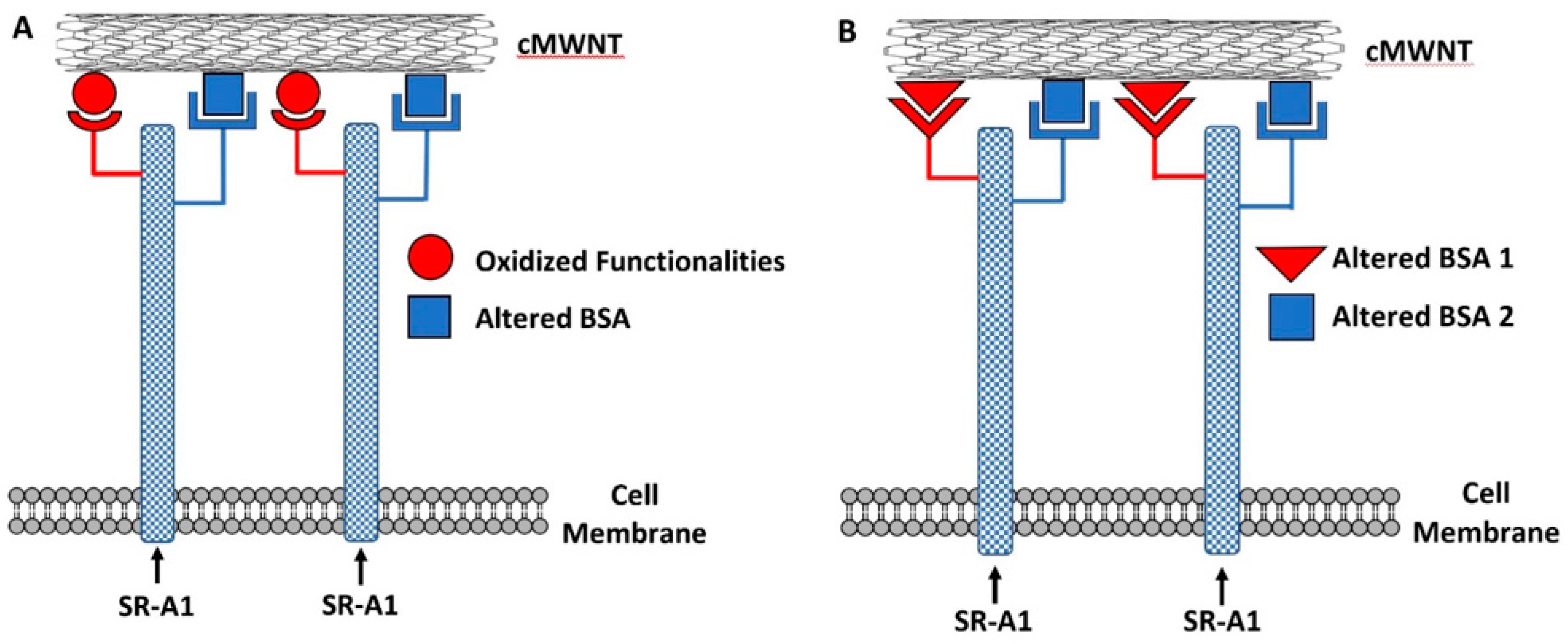
42]. This leads to Binding Hypothesis 1 in
Figure 10A, where BSA-coated cMWNTs present two sites that can interact with SR-A1—one for oxidized groups inherent to the nanotube and another for the coat of conformationally altered BSA protein. This model may explain why more BSA-cMWNTs bind cells than BSA-pMWNTs, and also is consistent with the semi-additive binding data: all binding sites are occupied by BSA-cMWNTs, whereas only the BSA binding sites are occupied by BSA-pMWNTs. The model is also consistent with the differences in dissociation of the two MWNT types from cells assuming BSA-cMWNTs and BSA-pMWNTs bound to SR-A1 at BSA binding sites dissociate slowly and that BSA-cMWNTs bound to oxidized functionalities dissociates more rapidly.
An alternative model is one in which all the oxidized binding sites on cMWNTs are unavailable because they are covered by BSA, and that binding of BSA to cMWNTs exposes additional latent SR-A1 binding sites that are not exposed when BSA binds to pMWNTs; hence, cells bind more BSA-cMWNTs than BSA-pMWNTs. A model of this type shown in
Figure 10B cannot be ruled out with the available data.
SR-A1 is a homotrimer and each monomer comprises an N-terminal cytoplasmic tail, a transmembrane domain, a spacer region, an α-helical coiled coil domain, a collagenous domain, and a C-terminal scavenger receptor cysteine rich (SRCR) domain [
15,
30,
43]. Depending on the ligand, either the collagenous domain, the SRCR, or both, may be involved in ligand binding of various scavenger receptors, but the details are not well understood and appear to depend on the structural context within each receptor type. For example, there is evidence from mutational studies with SR-A1 that positively charged residues in the collagenous domain are important for binding oxidized LDL [
31,
44]. Further, SR-A1.1, an alternatively spliced variant of SR-A1 lacking the SRCR domain, still binds oxidized LDL, suggesting that the collagenous domain is the major binding site for this ligand, although this does not rule out that the SRCR domain of SR-A1 may also interact with oxidized LDL or other protein ligands. Indeed, recent work suggests that the SR-A1 SRCR domain binds spectrin [
45] and ferritin [
46]. The SRCR domain is involved in the ligand binding by MARCO, a member of the class A scavenger receptors that shares the highly conserved SRCR domain with SR-A1 [
47,
48,
49]. The functional unit of many scavenger receptor family members is a trimer, including SR-A1, and the potential for three ligand binding sites per trimer is believed to enhance binding avidity to larger ligands, such as intact bacteria, and which would presumably include large ENPs such as MWNTs [
33]. This feature is not explicitly shown in the models of
Figure 10, but could be accommodated. Nevertheless, given the intricacies of how different domains in scavenger receptors interact with ligands, it is difficult to parse which SR-A1 domains interact with what features of BSA-coated MWNTs.
Additional complexities in scavenger receptor interaction with ligands arise from evidence that scavenger receptors, including SR-A1, may form complexes with other pattern recognition receptors, termed co-receptors, that also interact with the same ligand. The resulting complexes can then recruit components to form “Signalosomes” that contain two or more receptors bound to the same ligand plus associated signaling components that may activate cell signaling pathways [
32,
33,
50]. For example, there is evidence from computational work [
51] and from molecular docking studies that SWNTs may bind toll-like receptor 4 (TLR4) [
52]. It would be interesting to know whether the ~20% of cMWNT binding to RAW 264.7 cells lacking SR-A1 seen in
Figure 9 is due to TLR4. Thus, the simple models in
Figure 10 may not capture the range of possibilities for how MWNTs interact with SR-A1 and other cell components via co-receptors. Nevertheless, SR-A1 is a key player evidenced by the major loss of binding in SR-A1 knockout cells and the gain of binding in CHO cells that ectopically express SR-A1.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}