
The Microbial Functional Communities of Mollisol and Saline–Sodic Paddy Soils at Rice Heading and Harvest Stages in Northeast China

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Environmental Parameters
2.3. High-Throughput Sequencing of Soil
2.4. Metagenomic Data Analysis
2.5. Statistical Analysis
3. Result
3.1. Raw Sequencing Data
3.2. Alpha Diversity of Bacterial Communities
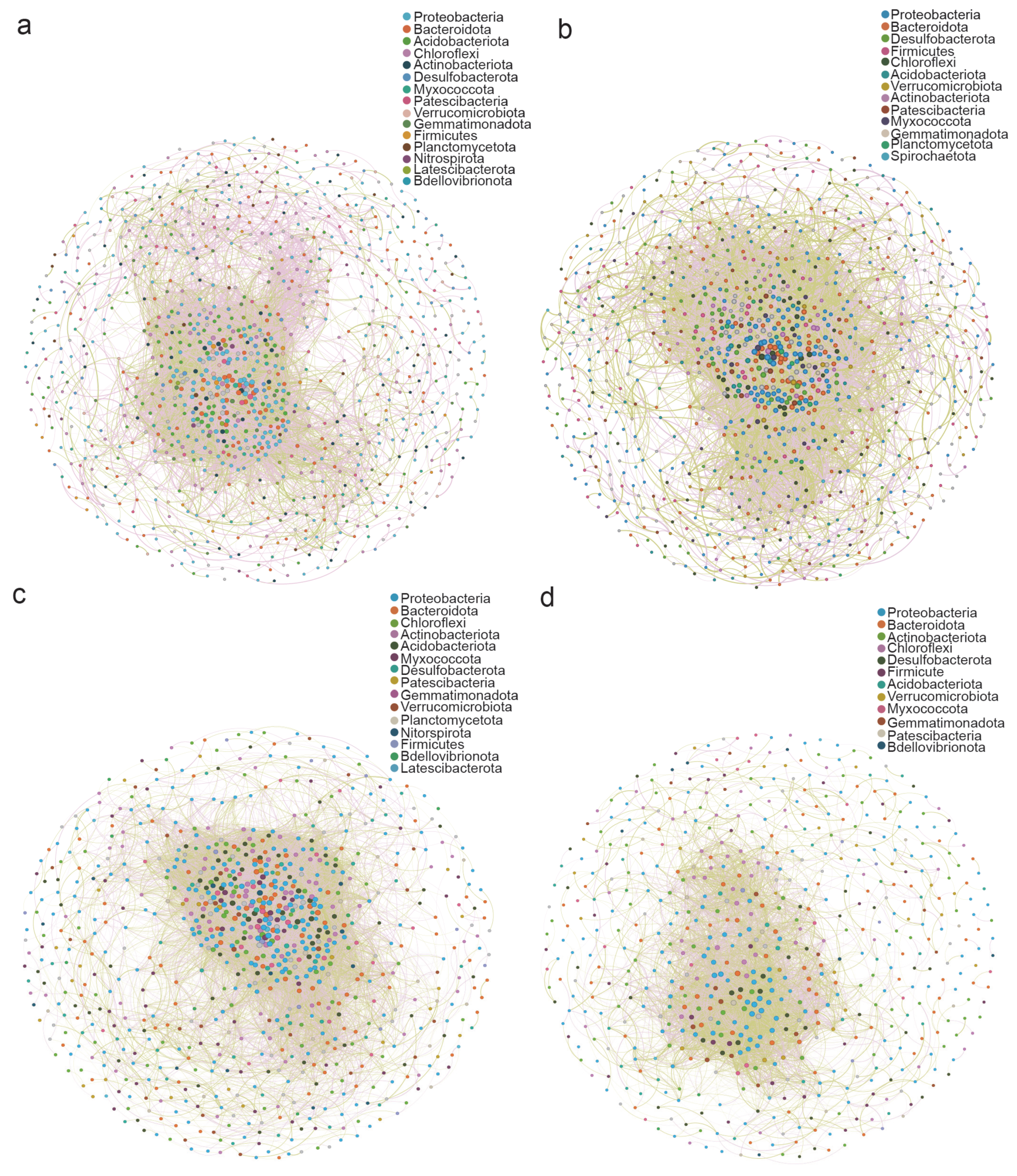
3.3. Beta Diversity of Bacterial Communities
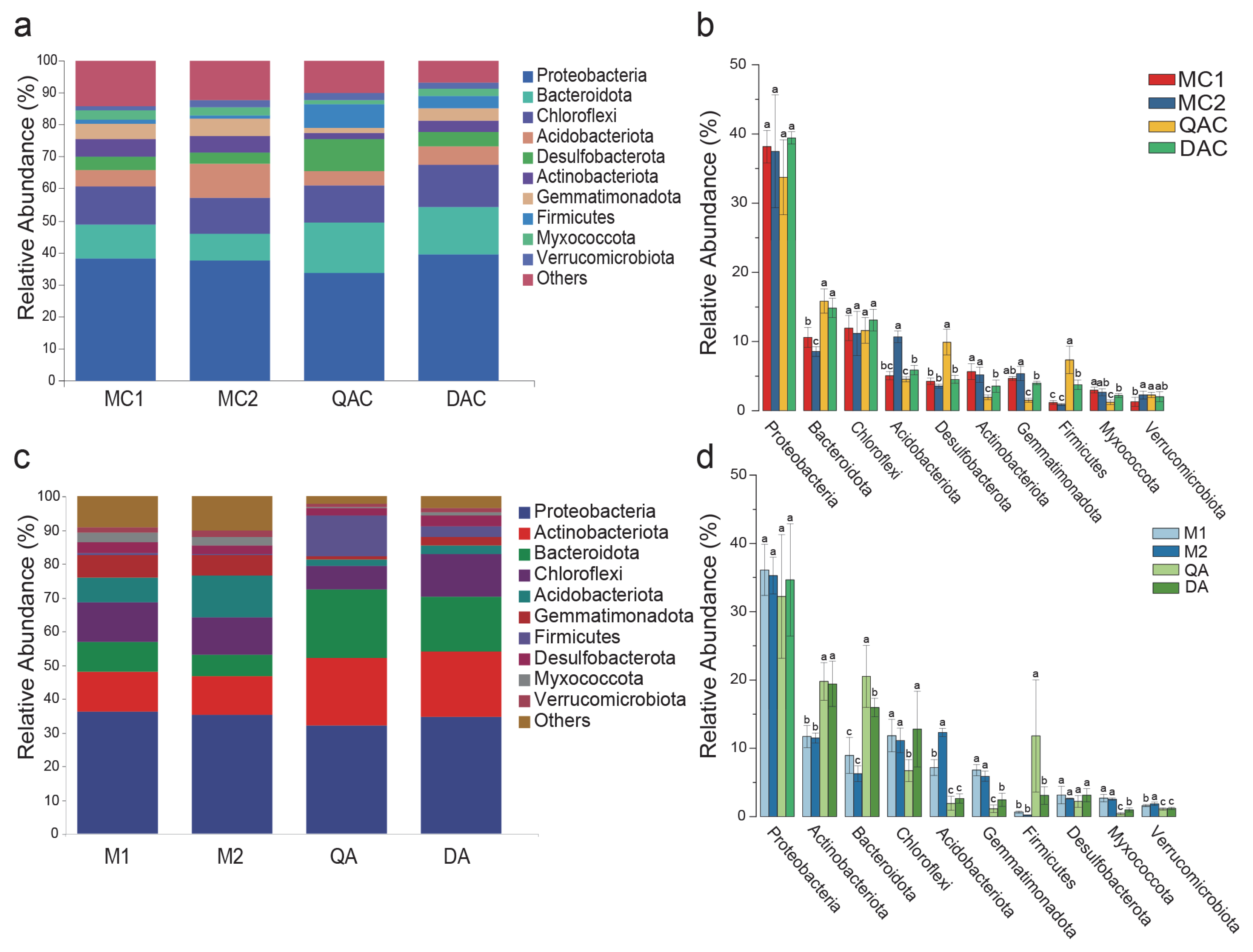
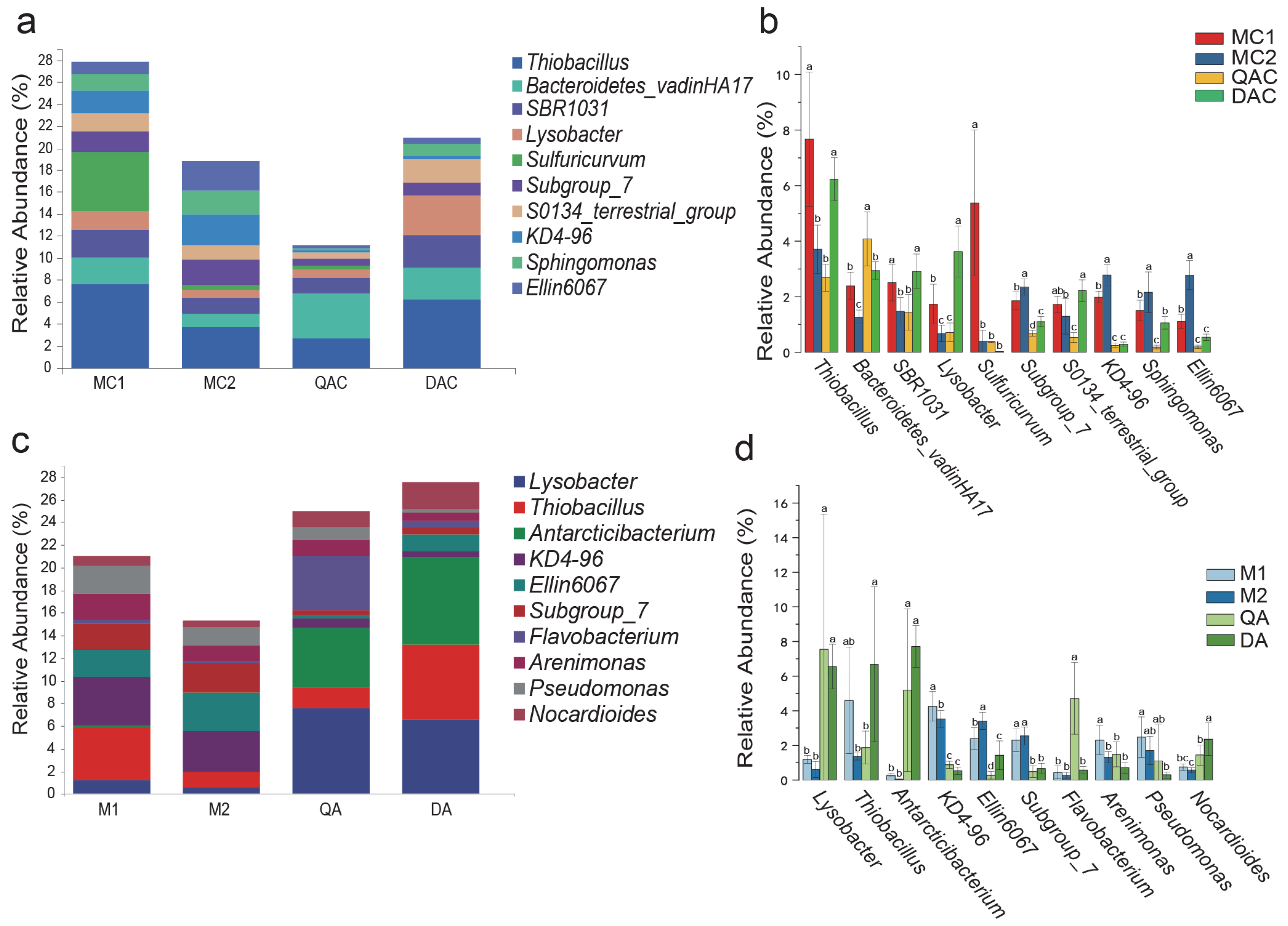
3.4. Soil Bacterial Community Composition and Structure
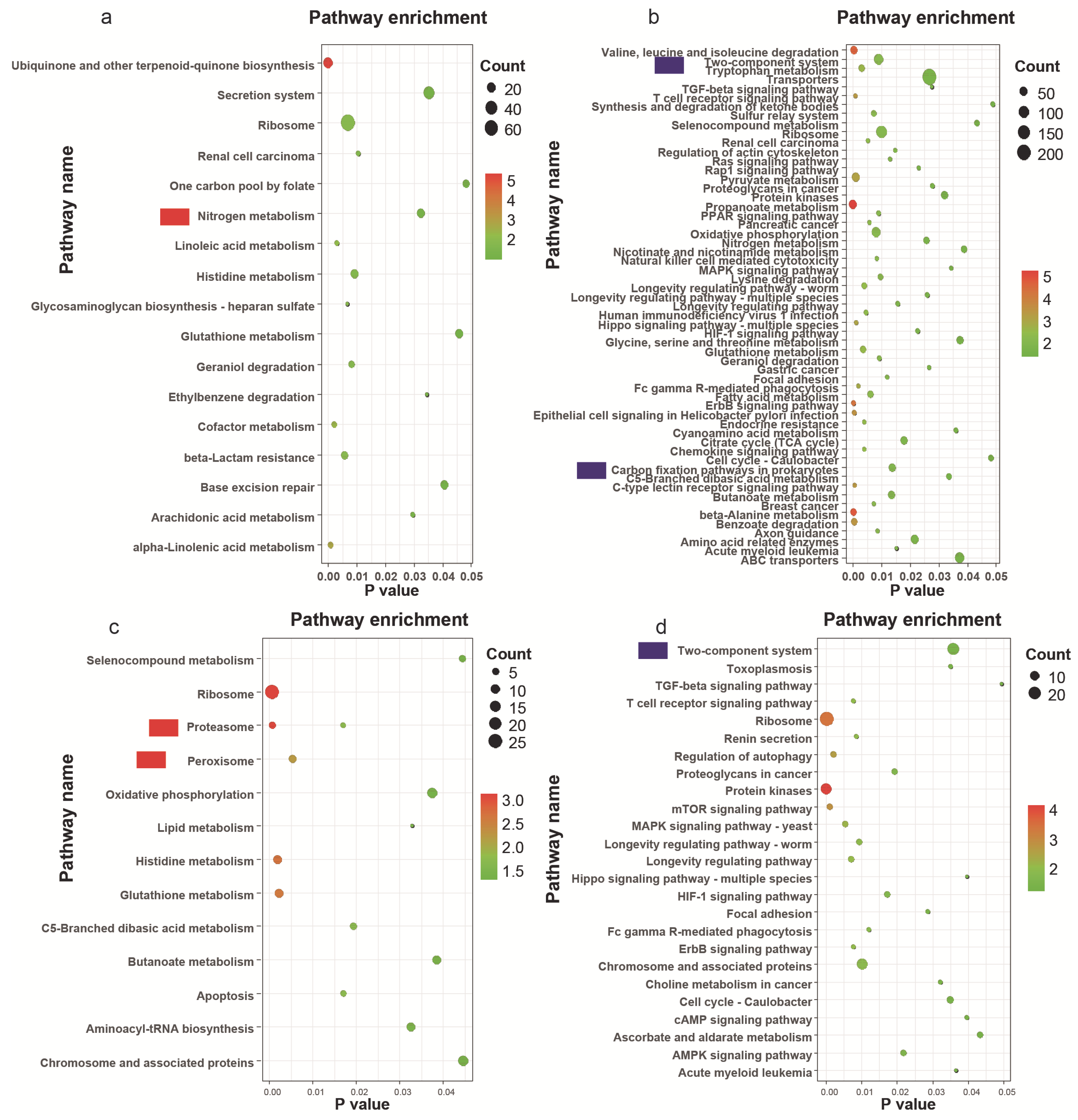
3.5. Metagenomic Data Analysis
3.6. RDA Analysis of the Influence of Soil Physicochemical Properties on Bacterial Community Diversity
4. Discussion
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Chang, J.; Shi, S.; Tian, L.; Leite, M.F.; Chang, C.; Ji, L.; Ma, L.; Tian, C.; Kuramae, E.E. Self-crossing leads to weak co-variation of the bacterial and fungal communities in the rice rhizosphere. Microorganisms 2021, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Xu, Z.X.; Wang, Z.Q. Influence of Soil Erosion to Soil Enzyme Activities in the Black Soil Area in Northeast China. Soil Water Conserv. China 2018, 9, 47–51+67. [Google Scholar] [CrossRef]
- Meng, F.J.; Yu, X.F.; Gao, J.L.; Zhao, Y.P. The Bottleneck and Breakthrough path of the Conservation Tillage Development in Black Soil of Northeast China. Issues Agric. Econ. 2020, 2, 135–142. [Google Scholar] [CrossRef]
- Li, Z.; Cao, Y.; Tang, J.; Wang, Y.; Duan, Y.; Jiang, Z.; Qu, Y. Relationships between temporal and spatial changes in lakes and climate change in the saline-alkali concentrated distribution area in the Southwest of Songnen Plain, Northeast China, from 1985 to 2015. Water 2020, 12, 3557. [Google Scholar] [CrossRef]
- Bian, J.; Nie, S.; Wang, R.; Wan, H.; Liu, C. Hydrochemical characteristics and quality assessment of groundwater for irrigation use in central and eastern Songnen Plain, Northeast China. Environ. Monit. Assess. 2018, 190, 1–16. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, X.; Meng, Q.; Zeng, X.; Zhang, J.; Li, D.; Wang, J.; Du, W.; Ma, X. Additional application of aluminum sulfate with different fertilizers ameliorates saline-sodic soil of Songnen Plain in Northeast China. J. Soils Sediments 2019, 19, 3521–3533. [Google Scholar] [CrossRef]
- Andreo-Jimenez, B.; Schilder, M.T.; Nijhuis, E.H.; Te Beest, D.E.; Bloem, J.; Visser, J.H.; van Os, G.; Brolsma, K.; de Boer, W.; Postma, J. Chitin-and keratin-rich soil amendments suppress Rhizoctonia solani disease via changes to the soil microbial community. Appl. Environ. Microbiol. 2021, 87, e00318–e00321. [Google Scholar] [CrossRef] [PubMed]
- Andreo-Jimenez, B.; Te Beest, D.E.; Kruijer, W.; Vannier, N.; Kadam, N.N.; Melandri, G.; Jagadish, S.K.; Van Der Linden, G.; Ruyter-Spira, C.; Vandenkoornhuyse, P. Genetic mapping of the root mycobiota in rice and its role in drought tolerance. Rice 2023, 16, 26. [Google Scholar] [CrossRef]
- Song, B.; Wang, T.; Wan, C.; Cai, Y.; Mao, L.; Ge, Z.; Yang, N. Diversity Patterns and Drivers of Soil Bacterial and Fungal Communities in a Muddy Coastal Wetland of China. J. Fungi 2024, 10, 770. [Google Scholar] [CrossRef]
- Bossio, D.; Scow, K.; Gunapala, N.; Graham, K. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef]
- Xu, Y.; Li, J.; Qiao, C.; Yang, J.; Li, J.; Zheng, X.; Wang, C.; Cao, P.; Li, Y.; Chen, Q. Rhizosphere bacterial community is mainly determined by soil environmental factors, but the active bacterial diversity is mainly shaped by plant selection. BMC Microbiol. 2024, 24, 450. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Liu, Y.; Cheng, F. Impact of Tree Species Mixture on Microbial Diversity and Community Structure in Soil Aggregates of Castanopsis hystrix Plantations. Microorganisms 2025, 13, 578. [Google Scholar] [CrossRef]
- Zhang, Y.; Miao, S.; Song, Y.; Wang, X.; Jin, F. Biochar application reduces saline–alkali stress by improving soil functions and regulating the diversity and abundance of soil bacterial community in highly saline–alkali paddy field. Sustainability 2024, 16, 1001. [Google Scholar] [CrossRef]
- Hu, X.J.; Liu, J.J.; Yu, Z.H.; Wei, D.; Zhou, B.K.; Chen, X.L.; Wang, G.H. Response of nirS-type denitrifier community and network structures tolong-term application of chemical fertilizers in a black soil of northeast China. J. Plant Nutr. Fertil. 2020, 26, 1–9. [Google Scholar]
- Hao, Z.Y.; Shao, Y.X.; Liu, T.; Zhang, S.; Wang, W.D.; Zhou, N.Y.; Yan, L. Research advances of carbon-fixing microorganisms in the black soil of northeast China. Microbiol. China 2024, 51, 1873–1886. [Google Scholar] [CrossRef]
- Zhang, G.; Bai, J.; Zhai, Y.; Jia, J.; Zhao, Q.; Wang, W.; Hu, X. Microbial diversity and functions in saline soils: A review from a biogeochemical perspective. J. Adv. Res. 2024, 59, 129–140. [Google Scholar] [CrossRef]
- Shi, W.; Takano, T.; Liu, S. Isolation and characterization of novel bacterial taxa from extreme alkali-saline soil. World J. Microbiol. Biotechnol. 2012, 28, 2147–2157. [Google Scholar] [CrossRef]
- Yang, H.T.; Leng, X.Y.; Shi, C.F. Research Progress on Bacterial Communities in Saline-alkali Soil. Environ. Sci. Technol. 2021, 44, 230–236. [Google Scholar] [CrossRef]
- Jiang, D.; Yan, Y.; Li, J.; Zhang, C.; Huangfu, S.; Sun, Y.; Sun, C.; Huang, L.; Tian, L. Arbuscular Mycorrhizal Fungi Play More Important Roles in Saline–Sodic Soil than in Black Soil of the Paddy Field in Northeast China. Agriculture 2025, 15, 951. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Chang, J.; Shi, S.; Ji, L.; Zhang, J.; Sun, Y.; Li, X.; Li, X.; Xie, H.; Cai, Y.; et al. Comparison of methane metabolism in the rhizomicrobiomes of wild and related cultivated rice accessions reveals a strong impact of crop domestication. Sci. Total Environ. 2022, 803, 150131. [Google Scholar] [CrossRef] [PubMed]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Xu, S.; Qiu, Q.; Yan, C.; Xiong, J. Structure, acquisition, assembly, and function of the root-associated microbiomes in Japonica rice and hybrid rice. Agric. Ecosyst. Environ. 2024, 373, 109122. [Google Scholar] [CrossRef]
- Cassán, F.; Diaz-Zorita, M. Azospirillum sp. in current agriculture: From the laboratory to the field. Soil Biol. Biochem. 2016, 103, 117–130. [Google Scholar] [CrossRef]
- Meng, X.Y.; Fan, S.X.; Dong, L.; Li, K.; Li, X.L. Response of Understory Plant Diversity to Soil Physical and Chemical Properties in Urban Forests in Beijing, China. Forests 2023, 14, 571. [Google Scholar] [CrossRef]
- Xu, M.P.; Wang, J.Y.; Zhu, Y.F.; Han, X.H.; Ren, C.J.; Yang, G.H. Plant Biomass and Soil Nutrients Mainly Explain the Variation of Soil Microbial Communities During Secondary Succession on the Loess Plateau. Microb. Ecol. 2022, 83, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Ehtaiwesh, A.F. The effect of salinity on nutrient availability and uptake in crop plants. Sci. J. Appl. Sci. Sabratha Univ. 2022, 9, 55–73. [Google Scholar]
- Mathur, M.; Mathur, P. Restoration of Saline and Sodic Soil Through Using Halophytes as Agroforestry Components. In Sustainable Management and Conservation of Environmental Resources in India; Apple Academic Press: Palm Bay, FL, USA, 2024; pp. 311–354. [Google Scholar]
- Wang, H.; Chun, L.; Ji, L.; Na, R.; Wei, Z.; Han, W. Investigating the diversity and influencing factors of the rhizosphere bacterial community associated with Salicornia europaea L. populations in semi-arid grassland. Agriculture 2024, 14, 1018. [Google Scholar] [CrossRef]
- Awagul, T.; Zhang, X.Q.; Yuan, F.Z.; Zhu, Y.B.; Zhang, L.; Du, W.J.; Ma, Z.H. Analysis of soil microbial diversity and influencing factors in Hemaquan New District in Urumqi. China Environ. Sci. 2023, 43, 277–287. [Google Scholar] [CrossRef]
- Bi, W.; Sun, Y.; Yao, Z.; Zhao, Z.; Niu, Y. Bacillus halophilus BH-8 Combined with Coal Gangue as a Composite Microbial Agent for the Rehabilitation of Saline-Alkali Land. Microorganisms 2025, 13, 532. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Huang, J.; Yuan, L. A new function of the biocontrol bacterium Lysobacter enzymogenes LE16 in the mineralization of soil organic phosphorus. Plant Soil 2019, 442, 299–309. [Google Scholar] [CrossRef]
- Wen, S.F.; Jiang, R.H.; Zhu, C.Q.; Zhang, M.; Wang, C.X.; Hou, X.L. Effects of Phosphate-solubilizing Bacteria on Maize Growth and Rhizosphere Soil Microbial Community Structure. Ecol. Rural. Environ. 2025, 40, 1–18. [Google Scholar] [CrossRef]
- Xing, Y.; Gu, Y.T.; Liu, L.; Li, G.H.; Guo, H.C.; Li, J. Growth promotion effects of Bacillus amyloliquefaciens on potato under water deficit by combining microbiome and metabolome. South. Agric. 2023, 54, 1977–1987. [Google Scholar]
- Huang, H.; Lin, J.; Wang, W.; Li, S. Biopolymers Produced by Sphingomonas Strains and Their Potential Applications in Petroleum Production. Polymers 2022, 14, 1920. [Google Scholar] [CrossRef]
- Geisen, S.; Hu, S.; dela Cruz, T.E.E.; Veen, G. Protists as catalyzers of microbial litter breakdown and carbon cycling at different temperature regimes. ISME J. 2021, 15, 618–621. [Google Scholar] [CrossRef]
- Yang, Y.; Gerrity, S.; Collins, G.; Chen, T.; Li, R.; Xie, S.; Zhan, X. Enrichment and characterization of autotrophic Thiobacillus denitrifiers from anaerobic sludge for nitrate removal. Process Biochem. 2018, 68, 165–170. [Google Scholar] [CrossRef]
- Bhatti, A.A.; Haq, S.; Bhat, R.A. Actinomycetes benefaction role in soil and plant health. Microb. Pathog. 2017, 111, 458–467. [Google Scholar] [CrossRef]
- Zhang, L.; Zeng, Q.; Liu, X.; Chen, P.; Guo, X.; Ma, L.Z.; Dong, H.; Huang, Y. Iron reduction by diverse actinobacteria under oxic and pH-neutral conditions and the formation of secondary minerals. Chem. Geol. 2019, 525, 390–399. [Google Scholar] [CrossRef]
- An, X.; Wang, Z.; Teng, X.; Zhou, R.; Wang, X.; Xu, M.; Lian, B. Rhizosphere bacterial diversity and environmental function prediction of wild salt-tolerant plants in coastal silt soil. Ecol. Indic. 2022, 134, 108503. [Google Scholar] [CrossRef]
- Gao, L.; Huang, Y.; Liu, Y.; Mohamed, O.A.A.; Fan, X.; Wang, L.; Li, L.; Ma, J. Bacterial community structure and potential microbial coexistence mechanism associated with three halophytes adapting to the extremely hypersaline environment. Microorganisms 2022, 10, 1124. [Google Scholar] [CrossRef]
- Yue, H.; Zhao, L.; Yang, D.; Zhang, M.; Wu, J.; Zhao, Z.; Xing, X.; Zhang, L.; Qin, Y.; Guo, F. Comparative analysis of the endophytic bacterial diversity of Populus euphratica oliv. in environments of different salinity intensities. Microbiol. Spectr. 2022, 10, e00500–e00522. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Jin, T.; Zhang, H.; Peng, J.; Zuo, N.; Huang, Y.; Han, Y.; Tian, C.; Yang, Y.; Peng, K.; et al. Deciphering the diversity and functions of plastisphere bacterial communities in plastic-mulching croplands of subtropical China. J. Hazard. Mater. 2022, 422, 126865. [Google Scholar] [CrossRef]
- Ding, J.; Yu, S. Impacts of Land Use on Soil Nitrogen-Cycling Microbial Communities: Insights from Community Structure, Functional Gene Abundance, and Network Complexity. Life 2025, 15, 466. [Google Scholar] [CrossRef]
- Yuan, A.; Kumar, S.D.; Wang, H.; Wang, S.; Impa, S.; Wang, H.; Guo, J.; Wang, Y.; Yang, Q.; Liu, X.J.A.; et al. Dynamic interplay among soil nutrients, rhizosphere metabolites, and microbes shape drought and heat stress responses in summer maize. Soil Biol. Biochem. 2024, 191, 109357. [Google Scholar] [CrossRef]
- Nan, L.; Guo, E.Q.; Tan, J.H.; Kang, F.Y. Influence of rotation fallow mode on bacterial community in soil. Agric. Res. Arid. Areas 2020, 38, 128–134. [Google Scholar]
- Begmatov, S.; Beletsky, A.V.; Dedysh, S.N.; Mardanov, A.V.; Ravin, N.V. Genome analysis of the candidate phylum MBNT15 bacterium from a boreal peatland predicted its respiratory versatility and dissimilatory iron metabolism. Front. Microbiol. 2022, 13, 951761. [Google Scholar] [CrossRef]
- Guo, J.; Zhou, Y.; Guo, H.; Min, W. Saline and alkaline stresses alter soil properties and composition and structure of gene-based nitrifier and denitrifier communities in a calcareous desert soil. BMC Microbiol. 2021, 21, 246. [Google Scholar] [CrossRef]
- Tian, Y.H.; Li, H.B.; Li, Z.J.; Quan, W.C.; Kan, S.M.; Luo, P.; Zhou, D.P.; Zheng, L. Effects of PAM water retention improvement on soil properties and microbial characteristics in dry-hot valley loamy soils. Environ. Ecol. 2025, 7, 115–120. [Google Scholar]
- Zhou, X.; Leite, M.; Zhang, Z.; Tian, L.; Chang, J.; Ma, L.; Li, X.; van Veen, J.; Tian, C.; Kuramae, E. Facilitation in the soil microbiome does not necessarily lead to niche expansion. Environ. Microbiome 2021, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sangwongchai, W.; Tananuwong, K.; Krusong, K.; Thitisaksakul, M. Yield, Grain Quality, and Starch Physicochemical Properties of 2 Elite Thai Rice Cultivars Grown under Varying Production Systems and Soil Characteristics. Foods 2021, 10, 2601. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Yan, Y.; Jiang, D.; Huangfu, S.; Ma, H.; Tian, L.; Huang, L. The Microbial Functional Communities of Mollisol and Saline–Sodic Paddy Soils at Rice Heading and Harvest Stages in Northeast China. Agriculture 2025, 15, 1261. https://doi.org/10.3390/agriculture15121261
Zhang J, Yan Y, Jiang D, Huangfu S, Ma H, Tian L, Huang L. The Microbial Functional Communities of Mollisol and Saline–Sodic Paddy Soils at Rice Heading and Harvest Stages in Northeast China. Agriculture. 2025; 15(12):1261. https://doi.org/10.3390/agriculture15121261
Chicago/Turabian StyleZhang, Jianfeng, Yuxin Yan, Dongxue Jiang, Shaoqi Huangfu, Hongyuan Ma, Lei Tian, and Lihua Huang. 2025. "The Microbial Functional Communities of Mollisol and Saline–Sodic Paddy Soils at Rice Heading and Harvest Stages in Northeast China" Agriculture 15, no. 12: 1261. https://doi.org/10.3390/agriculture15121261
APA StyleZhang, J., Yan, Y., Jiang, D., Huangfu, S., Ma, H., Tian, L., & Huang, L. (2025). The Microbial Functional Communities of Mollisol and Saline–Sodic Paddy Soils at Rice Heading and Harvest Stages in Northeast China. Agriculture, 15(12), 1261. https://doi.org/10.3390/agriculture15121261