
Arbuscular Mycorrhizal Fungi Play More Important Roles in Saline–Sodic Soil than in Black Soil of the Paddy Field in Northeast China
 ,
, 
Abstract
1. Introduction
2. Experimental Materials and Methods
2.1. Experimental Materials
2.2. Soil Treatment
2.3. DNA Extraction, Amplification, and High-Throughput Genome Sequencing
2.4. Metagenomic Data Analysis of Rhizophagus intraradices Sequencing Data in the Rhizospheres of Rice Plants
2.5. Determination of Spore Density and Soil Physical and Chemical Properties Between Black Soil and Saline–Sodic Soil
2.6. Determination of Glomalin-Related Soil Protein Concentration in Black Soil and Saline–Sodic Soil
2.7. Determination of Aggregate Content of Black Soil and Saline–Sodic Soil
2.8. Data Processing
3. Results
3.1. Statistical Analysis of AMF Sequencing Data of Black Soil and Saline–Sodic Soil in Late Autumn Harvest and Heading Stage
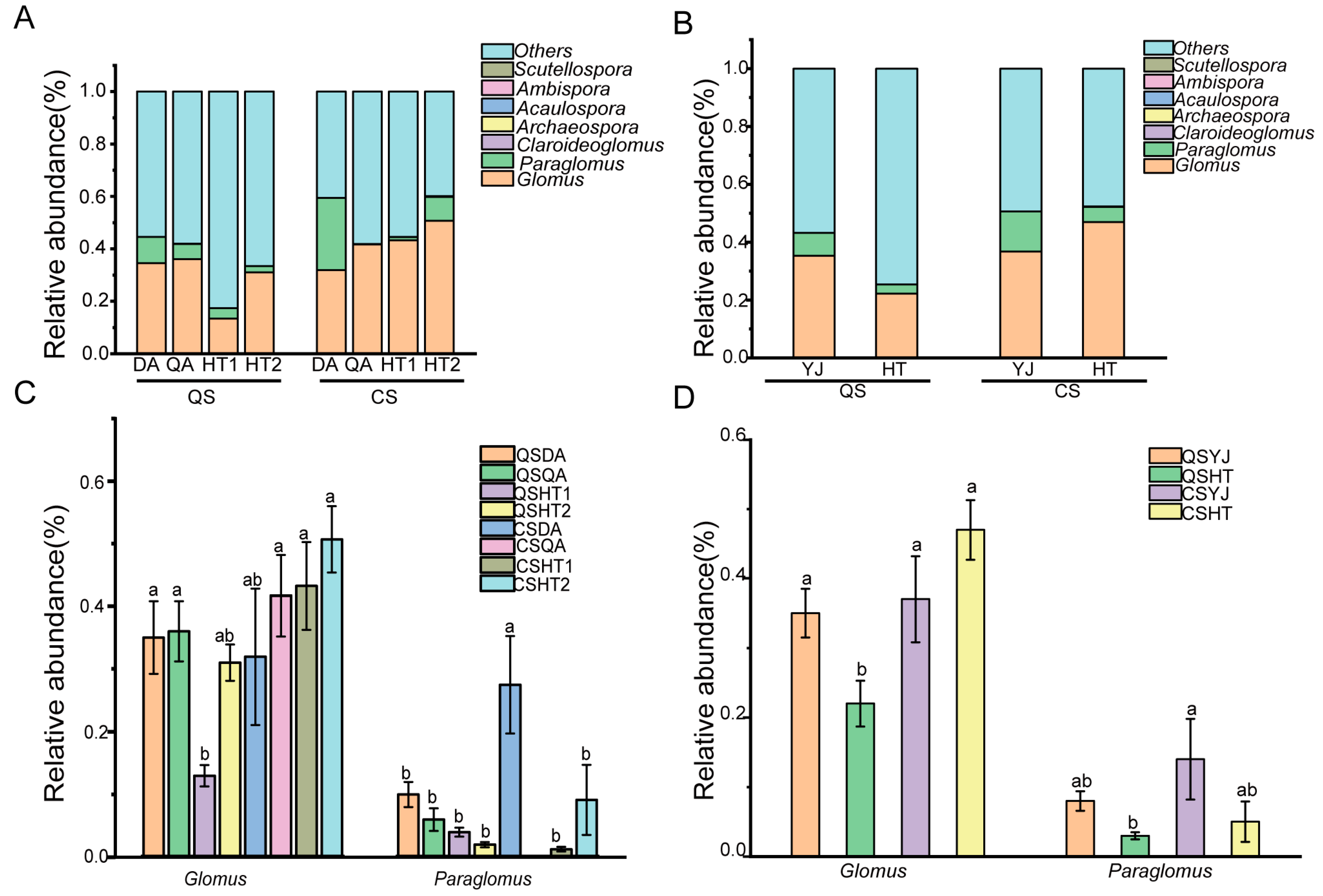
3.2. AMF Community Composition Analysis of Black Soil and Saline–Sodic Soil
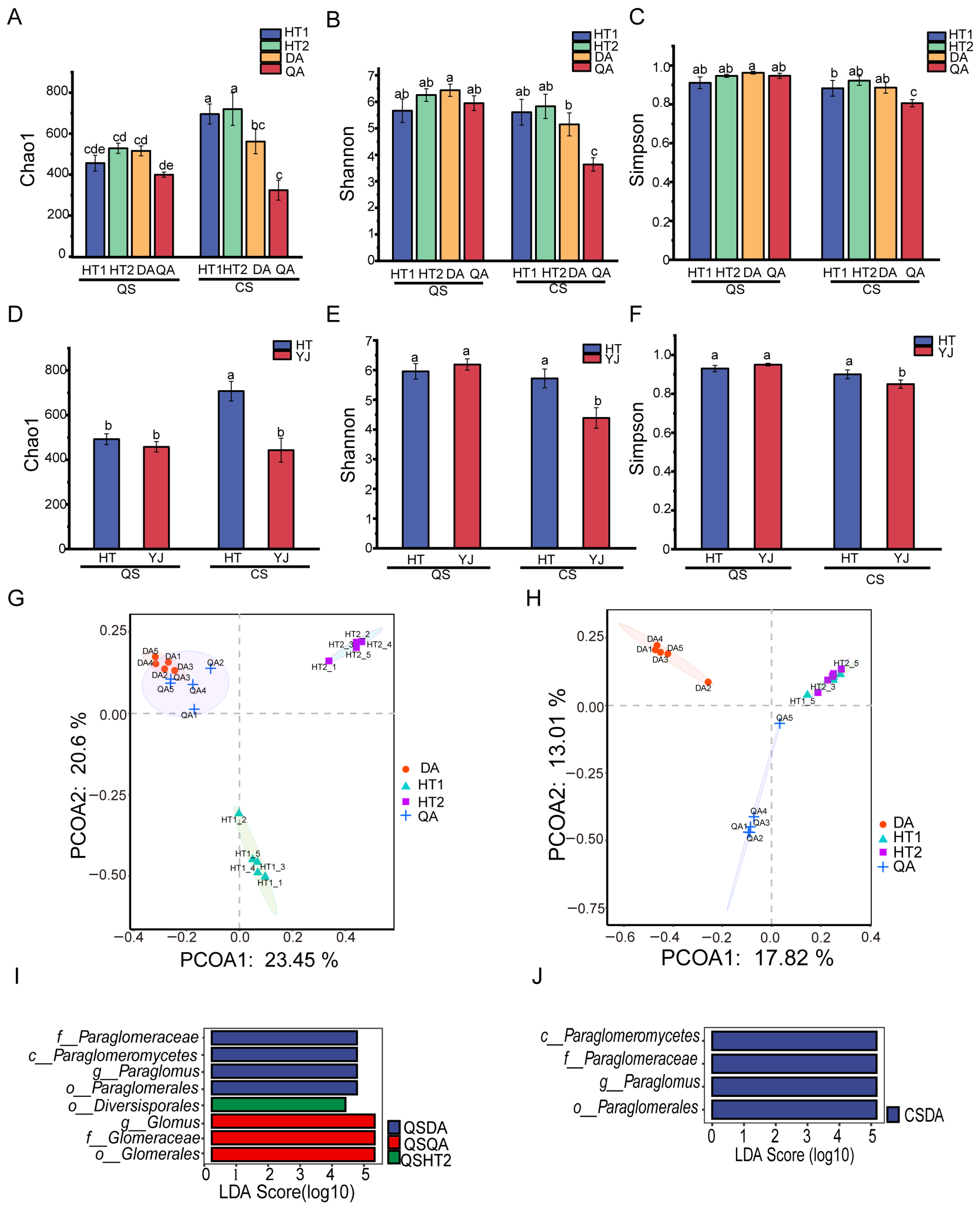
3.3. AMF Alpha Diversity of Black Soil and Saline–Sodic Soil
3.4. AMF Beta Diversity of Black Soil and Saline–Sodic Soil
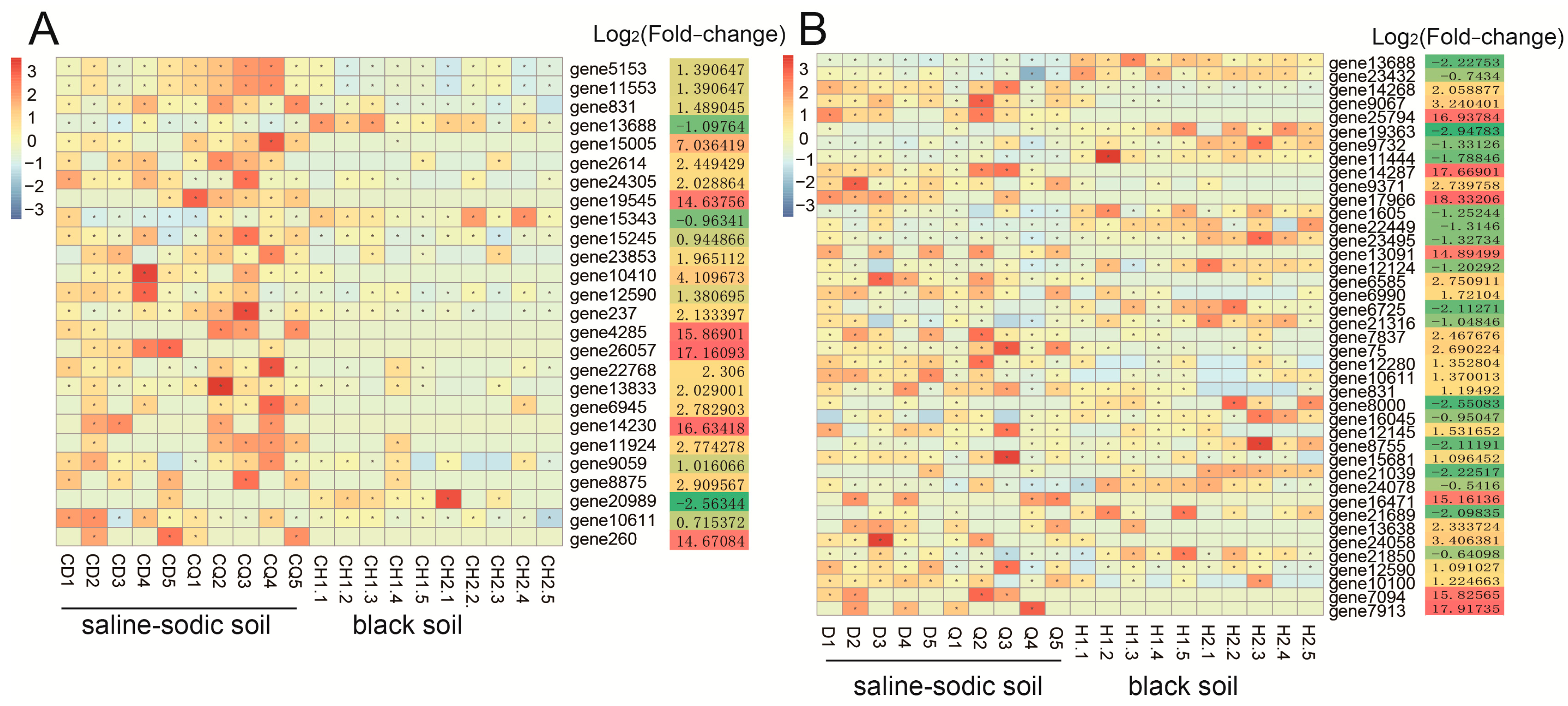
3.5. Metagenomic Data Analysis of R. intraradices-Related Genes in the Rhizospheres of Rice Plants
Metagenomic Data Analysis of R. intraradices in the Rhizospheres of Rice Plants
3.6. Physicochemical Properties and Glomalin-Related Soil Protein of Black Soil and Saline–Sodic Soil
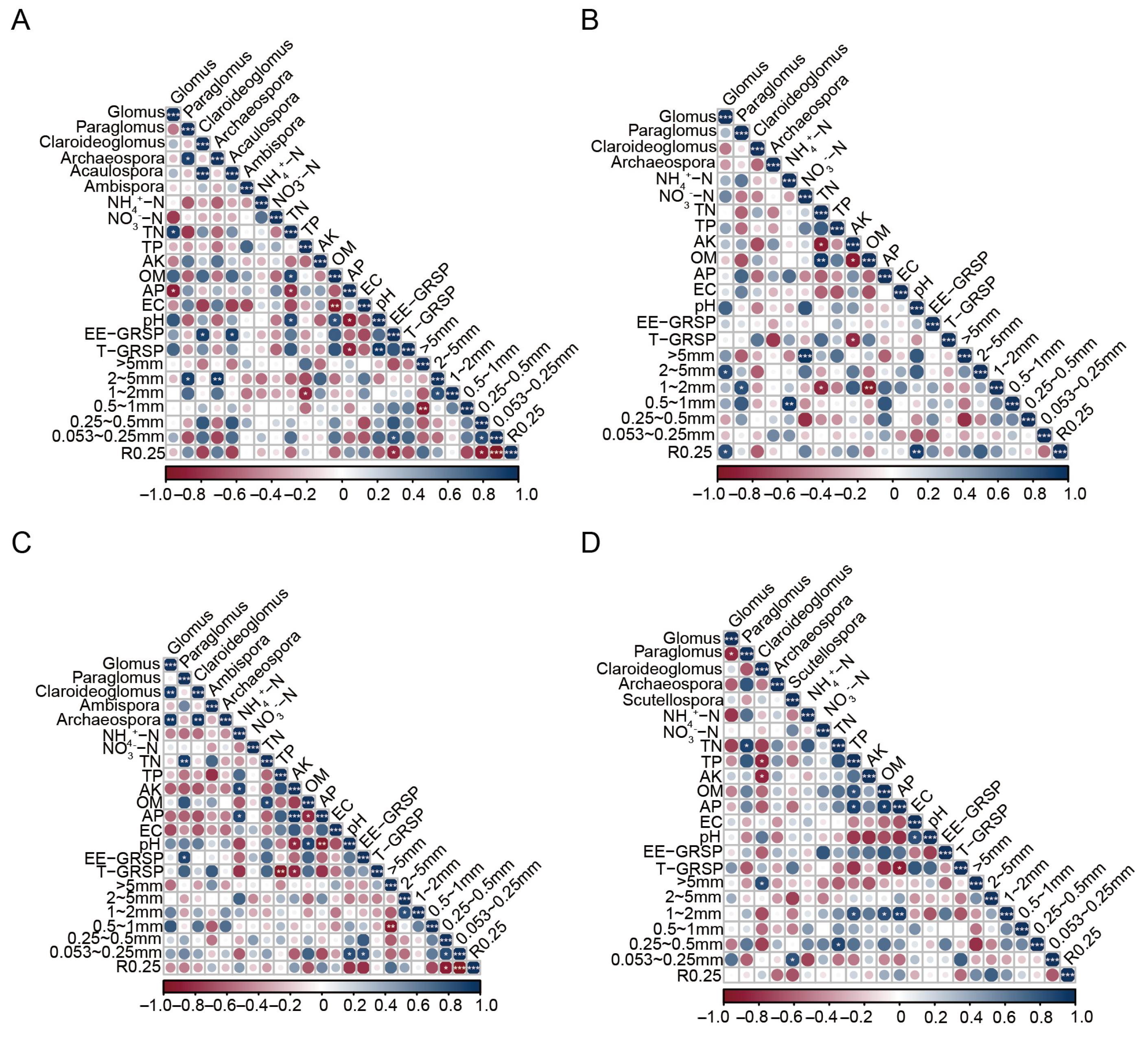
3.7. Correlation of the Genus Relative Abundance, Diversity, and Environmental Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bin Rahman, A.R.; Zhang, J. Trends in rice research: 2030 and beyond. Food Energy Secur. 2023, 12, e390. [Google Scholar] [CrossRef]
- Xinran, W. Analysis on the sustainable development of rice industry in Heilongjiang Province under the background of rural Revitalization. Shanxi Agric. Econ. 2025, 1, 195–197+220. (In Chinese) [Google Scholar] [CrossRef]
- Bing, H.; Chao, L.; Liang, Y.; Xiaoyang, W.; Dianyuan, C.; Guangbin, Y.; Zhenjiao, L. Analysis of Current Status and Development Strategies of Rice Seed Industry in Jilin Province. China Seed Ind. 2023, 12, 47–51+56. (In Chinese) [Google Scholar] [CrossRef]
- Fuyi, W. Analysis on Changes of Rice Planting in Liaoning Province. Agric. Sci. Technol. Equip. 2024, 04, 85–86. (In Chinese) [Google Scholar] [CrossRef]
- Chen, W.; Qi, W.; Yuan, F.; Li, Z. Comparative analysis of rice yield prediction in Jilin Province Based on time series and cross-sectional data. China Agric. Inf. 2018, 30, 91–101. [Google Scholar]
- Wang, S.; Huang, Y. Research progress on saline-alkali land improvement in Songnen Plain. Soils Crop 2023, 12, 206–217. [Google Scholar]
- Qadir, M.; Tubeileh, A.; Akhtar, J.; Larbi, A.; Minhas, P.; Khan, M. Productivity enhancement of salt-affected environments through crop diversification. Land Degrad. Dev. 2008, 19, 429–453. [Google Scholar] [CrossRef]
- Chen, B.D.; Yu, M.; Hao, Z.P.; Xie, W.; Zhang, X. Research progress in arbuscular mycorrhizal technology. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2019, 30, 1035–1046. [Google Scholar]
- Vosátka, M.; Látr, A.; Gianinazzi, S.; Albrechtová, J. Development of arbuscular mycorrhizal biotechnology and industry: Current achievements and bottlenecks. Symbiosis 2012, 58, 29–37. [Google Scholar] [CrossRef]
- Yang, M.; Guo, H.; Duan, G.; Wang, Z.; Fan, G.; Li, J. Role and mechanism of arbuscular mycorrhizal fungi in enhancing plant stress resistance and soil Improvement: A review. China Powder Sci. Technol 2024, 30, 164–172. [Google Scholar]
- Mathur, S.; Sharma, M.P.; Jajoo, A. Improved photosynthetic efficacy of maize (Zea mays) plants with arbuscular mycorrhizal fungi (AMF) under high temperature stress. J. Photochem. Photobiol. B Biol. 2018, 180, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Cao, Q.; Sun, L.; Yang, X.; Yang, W.; Zhang, H. Stomatal conductance and morphology of arbuscular mycorrhizal wheat plants response to elevated CO2 and NaCl stress. Front. Plant Sci. 2018, 9, 410525. [Google Scholar] [CrossRef]
- Shao, Y.-D.; Zhang, D.-J.; Hu, X.-C.; Wu, Q.-S.; Jiang, C.-J.; Xia, T.-J.; Gao, X.-B.; Kuča, K. Mycorrhiza-induced changes in root growth and nutrient absorption of tea plants. Plant Soil Environ. 2018, 64, 283–289. [Google Scholar] [CrossRef]
- Huang, Z.; Zhao, F.; Wang, M.; Qi, K.; Wu, J.; Zhang, S. Soil chemical properties and geographical distance exerted effects on arbuscular mycorrhizal fungal community composition in pear orchards in Jiangsu Province, China. Appl. Soil Ecol. 2019, 142, 18–24. [Google Scholar] [CrossRef]
- Augé, R.M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 2001, 11, 3–42. [Google Scholar] [CrossRef]
- Hodge, A.; Storer, K. Arbuscular mycorrhiza and nitrogen: Implications for individual plants through to ecosystems. Plant Soil 2015, 386, 1–19. [Google Scholar] [CrossRef]
- Garg, N.; Chandel, S. Effect of mycorrhizal inoculation on growth, nitrogen fixation, and nutrient uptake in Cicer arietinum (L.) under salt stress. Turk. J. Agric. For. 2011, 35, 205–214. [Google Scholar] [CrossRef]
- Bao, X.; Ma, Y.; Zou, J.; Wu, L.; Yang, T.; Huang, Q.; Zhang, B.; Chu, H. Research progress of arbuscular mycorrhizal fungi in paddy fields. Microbiol. China 2023, 50, 392–412. [Google Scholar]
- Cozzolino, V.; Di Meo, V.; Piccolo, A. Impact of arbuscular mycorrhizal fungi applications on maize production and soil phosphorus availability. J. Geochem. Explor. 2013, 129, 40–44. [Google Scholar] [CrossRef]
- Qian, S.; Xu, Y.; Zhang, Y.; Wang, X.; Niu, X.; Wang, P. Effect of AMF Inoculation on Reducing Excessive Fertilizer Use. Microorganisms 2024, 12, 1550. [Google Scholar] [CrossRef]
- Pang, F.; Li, Q.; Solanki, M.K.; Wang, Z.; Xing, Y.-X.; Dong, D.-F. Soil phosphorus transformation and plant uptake driven by phosphate-solubilizing microorganisms. Front. Microbiol. 2024, 15, 1383813. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liu, P.; Xu, K.; Lu, Z. Ecological distribution of AM bacteria in saline-alkali soil in China. J. Appl. Ecol. 1999, 10, 721. [Google Scholar]
- Yang, W. Arbuscular Mycorrhizal Fungal Diversity and Its Response to Soil Organic Carbon in the Black Soil Region of Northeast China. Ph.D. Thesis, University of Chinese Academy of Sciences, Changchun, China, 2022. (In Chinese). [Google Scholar]
- Bergen, M.; Koske, R. Vesicular-arbuscular mycorrhizal fungi from sand dunes of Cape Cod, Massachusetts. Trans. Br. Mycol. Soc. 1984, 83, 157–158. [Google Scholar] [CrossRef]
- Bainard, L.D.; Klironomos, J.N.; Gordon, A.M. Arbuscular mycorrhizal fungi in tree-based intercropping systems: A review of their abundance and diversity. Pedobiologia 2011, 54, 57–61. [Google Scholar] [CrossRef]
- Tian, L.; Wang, J.; Chen, H.; Li, W.; Tran, L.-S.P.; Tian, C. Integrative multi-omics approaches reveal that Asian cultivated rice domestication influences its symbiotic relationship with arbuscular mycorrhizal fungi. Pedosphere 2024, 34, 315–327. [Google Scholar] [CrossRef]
- Liu, H.; Chen, M.; Huang, Y.; Ren, J.; Fan, D.; Zhao, J. Diversity of arbuscular mycorrhizal fungi in the rhizosphere of tea plant from Anhui tea area, China. Chin. J. Appl. Ecol. 2017, 28, 2897–2906. [Google Scholar]
- Yang, Y.; Wu, F.; Chen, J.; Wu, Z.; Liu, L.; Zhang, D.; Ma, H.; Wu, J. Correlation between root rot of Camellia oleifera and arbuscular mycorrhizal fungi. Southwest China J. Agric. Sci. 2023, 36, 2426–2436. [Google Scholar]
- Bao, S. Soil Agrochemical Analysis; China Agriculture Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Wright, S.F.; Upadhyaya, A. A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi. Plant Soil 1998, 198, 97–107. [Google Scholar] [CrossRef]
- Janos, D.P.; Garamszegi, S.; Beltran, B. Glomalin extraction and measurement. Soil Biol. Biochem. 2008, 40, 728–739. [Google Scholar] [CrossRef]
- Cambardella, C.; Elliott, E. Carbon and nitrogen distribution in aggregates from cultivated and native grassland soils. Soil Sci. Soc. Am. J. 1993, 57, 1071–1076. [Google Scholar] [CrossRef]
- Leng, C.-X.; Zheng, F.-Y.; Zhao, B.-P.; Liu, H.-Y.; Wang, Y.-J. Advances on alkaline tolerance of rice. Biotechnol. Bull. 2020, 36, 103. [Google Scholar]
- Li, B.; Liu, Z.; Huang, F.; Yang, X.; Liu, Z.; Wan, W.; Wang, J.; Xu, Y.; Li, Z.; Ren, T. Ensuring national food security by strengthening high-productivity black soil granary in Northeast China. Bull. Chin. Acad. Sci. (Chin. Version) 2021, 36, 1184–1193. [Google Scholar]
- Zhang, S.-H.; Mi, C.-X.; Yu, Y.-J.; Liu, G.-Q.; Zhu, C.-Q.; Tian, W.-H.; Zhu, L.-F.; Cao, X.-C.; Zhang, J.-H.; Kong, Y.-L. Physiological characteristics of arbuscular mycorrhizal fungi in alleviating saline alkali stress in rice. China Rice 2023, 29, 56. [Google Scholar]
- Wang, Y.; Fan, J.; Shi, Z. Molecular diversity of arbuscular mycorrhizal fungal in China. Microbiol. China 2018, 45, 2399–2408. [Google Scholar]
- He, X.; Geng, X.; Zhao, L.; Niu, K. Seasonal variation of AM fungal diversity in the rhizosphere of Lonicera japonica. Ecol. Environ. Sci. 2013, 22, 90–94. [Google Scholar]
- Chen, K.; Tian, Q.; Liu, Z.; Wang, H.; Xiong, J.; Lei, Y.; Sun, Y. Diversity of arbuscular mycorrhizal fungi in cotton rhizosphere soil in Shihezi and surrounding areas, Xinjiang. Cotton Sci. 2022, 34, 69–78. [Google Scholar]
- Oehl, F.; Sieverding, E.; Ineichen, K.; Maeder, P.; Wiemken, A.; Boller, T. Distinct sporulation dynamics of arbuscular mycorrhizal fungal communities from different agroecosystems in long-term microcosms. Agric. Ecosyst. Environ. 2009, 134, 257–268. [Google Scholar] [CrossRef]
- Marinho, F.; Oehl, F.; da Silva, I.R.; Coyne, D.; da Nobrega Veras, J.S.; Maia, L.C. High diversity of arbuscular mycorrhizal fungi in natural and anthropized sites of a Brazilian tropical dry forest (Caatinga). Fungal Ecol. 2019, 40, 82–91. [Google Scholar] [CrossRef]
- Wang, F.; Liu, R. Preliminary investigation of AM fungi in saline soil in the Yellow River Delta. Biodiversity 2001, 9, 389. [Google Scholar]
- Hazard, C.; Gosling, P.; Van Der Gast, C.J.; Mitchell, D.T.; Doohan, F.M.; Bending, G.D. The role of local environment and geographical distance in determining community composition of arbuscular mycorrhizal fungi at the landscape scale. ISME J. 2013, 7, 498–508. [Google Scholar] [CrossRef]
- Oehl, F.; Laczko, E.; Oberholzer, H.-R.; Jansa, J.; Egli, S. Diversity and biogeography of arbuscular mycorrhizal fungi in agricultural soils. Biol. Fertil. Soils 2017, 53, 777–797. [Google Scholar] [CrossRef]
- Miller, S.P.; Sharitz, R. Manipulation of flooding and arbuscular mycorrhiza formation influences growth and nutrition of two semiaquatic grass species. Funct. Ecol. 2000, 14, 738–748. [Google Scholar] [CrossRef]
- Zhou, D.; Sun, H.; Zhu, C.; Zhao, Z.; Zhou, S.; Wu, S. Effects of nitrogen level on rice yield, AMF infection status and soil traits in rhizosphere. Shanghai J. Agric. 2021, 37, 35–41. [Google Scholar] [CrossRef]
- Karasawa, T.; Ariharal, J.; Kasahara, Y. Effects of previous crops on arbuscular mycorrhizal formation and growth of maize under various soil moisture conditions. Soil Sci. Plant Nutr. 2000, 46, 53–60. [Google Scholar] [CrossRef]
- Oehl, F.; Laczko, E.; Bogenrieder, A.; Stahr, K.; Bösch, R.; van der Heijden, M.; Sieverding, E. Soil type and land use intensity determine the composition of arbuscular mycorrhizal fungal communities. Soil Biol. Biochem. 2010, 42, 724–738. [Google Scholar] [CrossRef]
- Wright, S.; Upadhyaya, A.; Buyer, J. Comparison of N-linked oligosaccharides of glomalin from arbuscular mycorrhizal fungi and soils by capillary electrophoresis. Soil Biol. Biochem. 1998, 30, 1853–1857. [Google Scholar] [CrossRef]
- Agnihotri, R.; Sharma, M.P.; Prakash, A.; Ramesh, A.; Bhattacharjya, S.; Patra, A.K.; Manna, M.C.; Kurganova, I.; Kuzyakov, Y. Glycoproteins of arbuscular mycorrhiza for soil carbon sequestration: Review of mechanisms and controls. Sci. Total Environ. 2022, 806, 150571. [Google Scholar] [CrossRef]
- Li, S.; Chen, P.; Liu, H.; Hao, J.; Zhou, W.; Shi, L. Mechanism and Ecological Effects of Arbuscular Mycorrhizal Fungi on Improving Salt Tolerance of Plants in Coastal Saline-alkaline Land. Ecol. Environ. Sci. 2019, 28, 411–418. [Google Scholar]
- Wang, J.; Zhou, Z.; Ling, W. Distribution and environmental function of glomalin-related soil protein: A review. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2016, 27, 634–642. [Google Scholar]
- Tang, H.-L.; Liu, L.; Wang, L.; Ba, C.-J. Effect of land use type on profile distribution of glomalin. Chin. J. Eco-Agric. 2009, 17, 1137–1142. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Wright, S.F.; Clark, D.A.; Ruess, R.W. Soil stocks of glomalin produced by arbuscular mycorrhizal fungi across a tropical rain forest landscape. J. Ecol. 2004, 92, 278–287. [Google Scholar] [CrossRef]
- Rillig, M.C.; Wright, S.F.; Eviner, V.T. The role of arbuscular mycorrhizal fungi and glomalin in soil aggregation: Comparing effects of five plant species. Plant Soil 2002, 238, 325–333. [Google Scholar] [CrossRef]
- QF, W.; Xie, G.; Wang, J. The stability of organic carbon and aggregates of three limestone soils in the Western Zhejiang. Chin. J. Soil Sci. 2018, 49, 567–574. [Google Scholar]
- Schindler, F.V.; Mercer, E.J.; Rice, J.A. Chemical characteristics of glomalin-related soil protein (GRSP) extracted from soils of varying organic matter content. Soil Biol. Biochem. 2007, 39, 320–329. [Google Scholar] [CrossRef]
- Whiffen, L.K.; Midgley, D.J.; McGee, P.A. Polyphenolic compounds interfere with quantification of protein in soil extracts using the Bradford method. Soil Biol. Biochem. 2007, 39, 691–694. [Google Scholar] [CrossRef]
- Singh, A.K.; Rai, A.; Pandey, V.; Singh, N. Contribution of glomalin to dissolve organic carbon under different land uses and seasonality in dry tropics. J. Environ. Manag. 2017, 192, 142–149. [Google Scholar] [CrossRef]
- Gałązka, A.; Niedźwiecki, J.; Grządziel, J.; Gawryjołek, K. Evaluation of changes in Glomalin-Related Soil Proteins (GRSP) content, microbial diversity and physical properties depending on the type of soil as the important biotic determinants of soil quality. Agronomy 2020, 10, 1279. [Google Scholar] [CrossRef]
- Wilson, G.W.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef]
- Hood, L.; Swaine, M.D.; Mason, P. The influence of spatial patterns of damping-off disease and arbuscular mycorrhizal colonization on tree seedling establishment in Ghanaian tropical forest soil. J. Ecol. 2004, 92, 816–823. [Google Scholar] [CrossRef]
- Kiers, E.T.; Lovelock, C.E.; Krueger, E.L.; Herre, E.A. Differential effects of tropical arbuscular mycorrhizal fungal inocula on root colonization and tree seedling growth: Implications for tropical forest diversity. Ecol. Lett. 2000, 3, 106–113. [Google Scholar] [CrossRef]
- Xu, X.; Chen, C.; Zhang, Z.; Sun, Z.; Chen, Y.; Jiang, J.; Shen, Z. The influence of environmental factors on communities of arbuscular mycorrhizal fungi associated with Chenopodium ambrosioides revealed by MiSeq sequencing investigation. Sci. Rep. 2017, 7, 45134. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wang, Z.; He, Y.; Li, G.; Lv, X.; Zhuang, L. High-throughput sequencing analysis of the rhizosphere arbuscular mycorrhizal fungi (AMF) community composition associated with Ferula sinkiangensis. BMC Microbiol. 2020, 20, 335. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Late Autumn Harvest | Heading Period | ||||
|---|---|---|---|---|---|---|
| Number of Original Sequences | Number of Effective Sequences | OTU | Number of Original Sequences | Number of Effective Sequences | OTU | |
| DA | 76,977 | 59,845 | 1512 | 95,340 | 77,812 | 1871 |
| QA | 77,481 | 63,450 | 1266 | 89,725 | 77,790 | 1292 |
| HT1 | 65,568 | 51,749 | 1305 | 93,178 | 72,114 | 2310 |
| HT2 | 59,876 | 45,986 | 1512 | 99,929 | 78,431 | 2510 |
| Total | 279,902 | 221,030 | 5595 | 378,172 | 306,147 | 7983 |
| Unit | HT1 | HT2 | DA | QA | |
|---|---|---|---|---|---|
| NH4+-N | mg/kg | 7 ± 2.12 a | 7.58 ± 1.16 a | 36.57 ± 7.88 a | 30.56 ± 22.51 a |
| NO3−-N | mg/kg | 65.44 ± 28.68 a | 26.23 ± 2.08 a | 24.67 ± 4.14 a | 50.3 ± 31.97 a |
| TN | mg/kg | 358.76 ± 40.22 b | 695.98 ± 35.42 a | 160.5 ± 10.16 c | 292.69 ± 12.93 b |
| TP | mg/kg | 637.66 ± 56.36 a | 597.86 ± 18.38 a | 265.52 ± 16.02 b | 315.75 ± 16.67 b |
| AK | mg/kg | 226.46 ± 21.90 a | 199.73 ± 15.49 ab | 196.06 ± 1.92 ab | 150.18 ± 14.68 b |
| OM | % | 1.81 ± 0.33 b | 2.88 ± 0.28 a | 0.92 ± 0.03 c | 1.25 ± 0.045 bc |
| AP | mg/kg | 65.53 ± 1.56 a | 36.63 ± 4.28 b | 28.37 ± 1.03 b | 15.94 ± 5.46 c |
| EC | us/cm | 815 ± 104.84 ab | 646 ± 95.63 b | 1187.67 ± 200.11 a | 880.67 ± 145.48 ab |
| pH | 6.69 ± 0.07 c | 7.11 ± 0.07 b | 7.51 ± 0.04 a | 7.51 ± 0.1 a |
| Unit | HT1 | HT2 | DA | QA | |
|---|---|---|---|---|---|
| NH4+-N | mg/kg | 2.83 ± 0.35 a | 1.83 ± 0.07 b | 2.34 ± 0.25 ab | 2.4 ± 0.16 ab |
| NO3−-N | mg/kg | 7.64 ± 5.42 a | 6.94 ± 3.24 a | 5.19 ± 1.10 a | 3.5 ± 1.10 a |
| TN | mg/kg | 1153 ± 43.05 b | 1536.25 ± 137.07 a | 823.83 ± 47.38 c | 740.74 ± 28.99 c |
| TP | mg/kg | 640.83 ± 22.64 a | 561.5 ± 30.08 b | 317.68 ± 5.38 c | 264.3 ± 4.09 c |
| AK | mg/kg | 216.67 ± 9.73 a | 155.6 ± 3.98 b | 172.16 ± 6.91 b | 152.67 ± 3.33 b |
| OM | % | 2.71 ± 0.14 b | 3.43 ± 0.13 a | 1.64 ± 0.06 c | 1.41 ± 0.04 c |
| AP | mg/kg | 112.69 ± 2.05 a | 92 ± 1.42 b | 87.12 ± 1.21 b | 73.56 ± 039 c |
| EC | us/cm | 927.5 ± 143.35 ab | 358.75 ± 78.78 c | 760 ± 71.74 bc | 1205.6 ± 149.96 a |
| pH | 6.34± 0.27 c | 7.76 ± 0.02 b | 7.51 ± 0.14 b | 8.39 ± 0.13 a |
| Different Stage | Index | HT1 | HT2 | DA | QA |
|---|---|---|---|---|---|
| Heading stage | EE-GRSP (mg/g) | 0.25 ± 0.11 b | 0.53 ± 0.60 ab | 0.8 ± 0.03 a | 0.66 ± 0.22 ab |
| T-GRSP (mg/g) | 1.24 ± 0.69 b | 2.3 ± 0.68 a | 1.2 ± 0.34 c | 2.3 ± 0.55 c | |
| SD (spores/g) | 5.67 ± 0.33 c | 8.67 ± 2.89 bc | 17.67 ± 3.18 a | 10.33 ± 1.33 bc | |
| Harvest stage | EE-GRSP (mg/g) | 1.33 ± 0.13 b | 4.16 ± 1.72 a | 5.3 ± 0.89 a | 5.11 ± 0.02 a |
| T-GRSP (mg/g) | 9.27 ± 0.77 b | 17.41 ± 2.38 a | 7.7 ± 0.97 b | 9.85 ± 1.06 b | |
| SD (spores/g) | 5.67 ± 0.33 c | 8.33 ± 1.2 bc | 13 ± 2.08 ab | 9.33 ± 3.18 bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, D.; Yan, Y.; Li, J.; Zhang, C.; Huangfu, S.; Sun, Y.; Sun, C.; Huang, L.; Tian, L. Arbuscular Mycorrhizal Fungi Play More Important Roles in Saline–Sodic Soil than in Black Soil of the Paddy Field in Northeast China. Agriculture 2025, 15, 951. https://doi.org/10.3390/agriculture15090951
Jiang D, Yan Y, Li J, Zhang C, Huangfu S, Sun Y, Sun C, Huang L, Tian L. Arbuscular Mycorrhizal Fungi Play More Important Roles in Saline–Sodic Soil than in Black Soil of the Paddy Field in Northeast China. Agriculture. 2025; 15(9):951. https://doi.org/10.3390/agriculture15090951
Chicago/Turabian StyleJiang, Dongxue, Yuxin Yan, Jiaqi Li, Chenyu Zhang, Shaoqi Huangfu, Yang Sun, Chunyu Sun, Lihua Huang, and Lei Tian. 2025. "Arbuscular Mycorrhizal Fungi Play More Important Roles in Saline–Sodic Soil than in Black Soil of the Paddy Field in Northeast China" Agriculture 15, no. 9: 951. https://doi.org/10.3390/agriculture15090951
APA StyleJiang, D., Yan, Y., Li, J., Zhang, C., Huangfu, S., Sun, Y., Sun, C., Huang, L., & Tian, L. (2025). Arbuscular Mycorrhizal Fungi Play More Important Roles in Saline–Sodic Soil than in Black Soil of the Paddy Field in Northeast China. Agriculture, 15(9), 951. https://doi.org/10.3390/agriculture15090951