Ground Beetles (Carabidae) in the Short-Rotation Coppice Willow and Poplar Plants—Synergistic Benefits System



Abstract
1. Introduction
- -
- As a well-known group of epigeic insects, they can be treated as a monitoring group;
- -
- They play an important role, providing valuable ecosystem services for willow plantations and adjacent crops;
- -
- Compared to other insect groups for which the amount of data is negligible, there are more studies for Carabidae clusters, allowing for analysis.
2. Materials and Methods/Data Collection and Selection
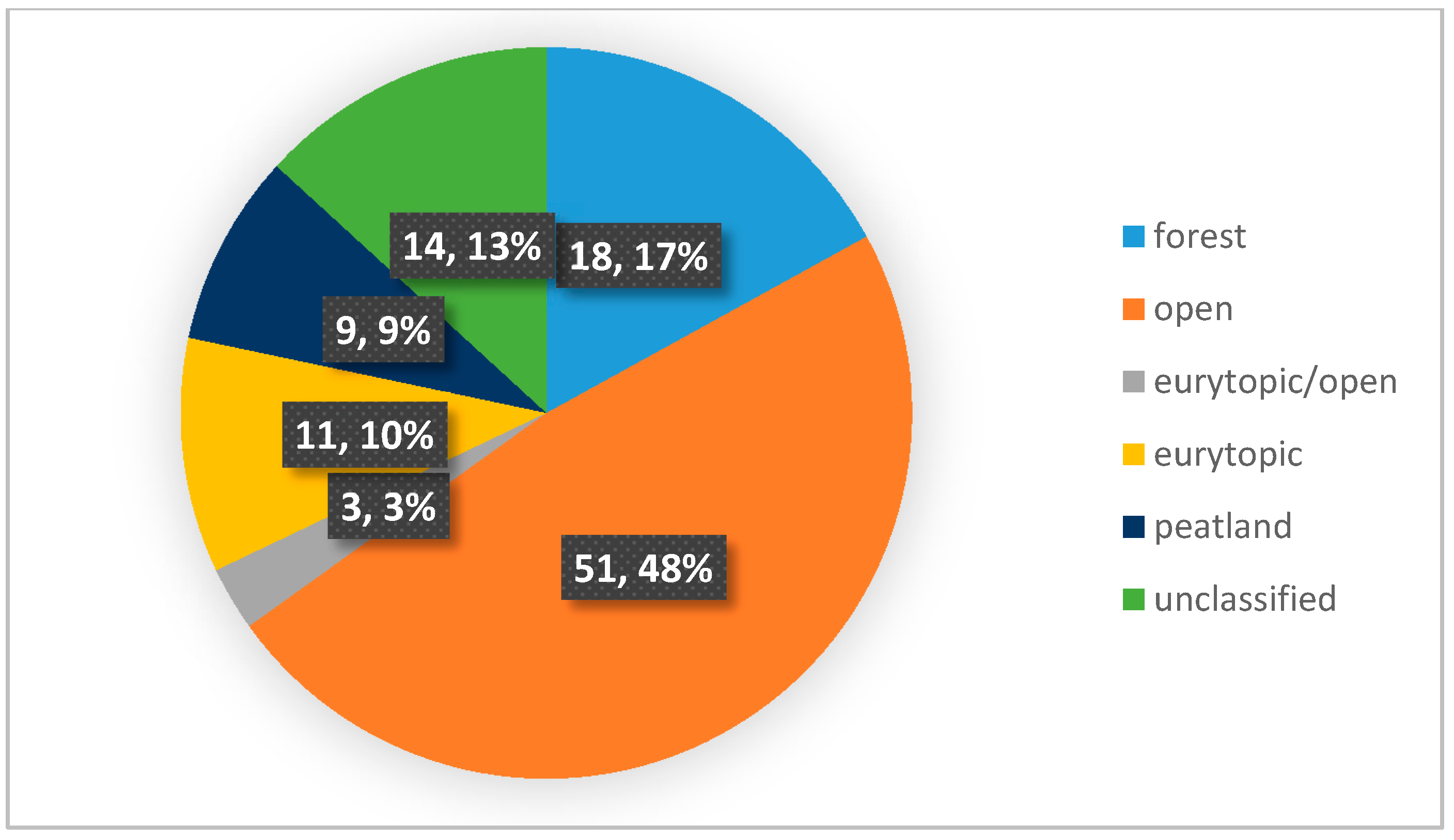
3. Results
4. Discussion
4.1. Factors Affecting Biodiversity
4.1.1. Rotation Length and Canopy Age vs. Plantation Age
4.1.2. Surrounding Area—Environmental Corridors and Islands
4.2. Carabidae as Bioindicators and Ecosystem Services Providers
4.3. Carabidae Assemblages Structure
5. Conclusions with Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lindegaard, K.N.; Adams, P.W.R.; Holley, M.; Lamley, A.; Henriksson, A.; Larsson, S.; von Engelbrechten, H.-G.; Esteban Lopez, G.; Pisarek, M. Short rotation plantations policy history in Europe: Lessons from the past and recommendations for the future. Food Energy Secur. 2016, 5, 125–152. [Google Scholar] [CrossRef] [PubMed]
- Volk, T.A.; Verwijst, T.; Tharakan, P.J.; Abrahamson, L.P.; White, E.H. Growing fuel: A sustainability assessment of willow biomass crops. Front. Ecol. Environ. 2004, 2, 411–418. [Google Scholar] [CrossRef]
- Groom, M.J.; Gray, E.M.; Townsend, P.A. Biofuels and Biodiversity: Principles for Creating Better Policies for Biofuel Production. Conserv. Biol. 2008, 22, 602–609. [Google Scholar] [CrossRef]
- Mohr, A.; Raman, S. Lessons from first-generation biofuels and implications for the sustainability appraisal of second-generation biofuels. Energy Policy 2013, 63, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Saladini, F.; Patrizi, N.; Pulselli, F.M.; Marchettini, N.; Bastianoni, S. Guidelines for emergy evaluation of first, second and third-generation biofuels. Renew. Sustain. Energy Rev. 2016, 66, 221–227. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, Y.; Zhang, F.; Yun, J.; Shen, Z. The impact of biofuel plantation on biodiversity: A review. Chin. Sci. Bull. 2014, 59, 4639–4651. [Google Scholar] [CrossRef]
- Stolarski, M.; Szczukowski, S.; Tworkowski, J.; Krzyżaniak, M. Extensive Willow Biomass Production on Marginal Land. Pol. J. Environ. Stud. 2019, 28, 4359–4367. [Google Scholar] [CrossRef]
- Dauber, J.; Jones, M.B.; Stout, J.C. The impact of biomass crop cultivation on temperate biodiversity: Biomass crops and biodiversity. GCB Bioenergy 2010, 2, 289–309. [Google Scholar] [CrossRef]
- Demirbas, M.F.; Balat, M.; Balat, H. Potential contribution of biomass to the sustainable energy development. Energy Convers. Manag. 2009, 50, 1746–1760. [Google Scholar] [CrossRef]
- Jensen, J.K.; Holm, P.E.; Nejrup, J.; Larsen, M.B.; Borggaard, O.K. The potential of willow for remediation of heavy metal polluted calcareous urban soils. Environ. Pollut. 2009, 157, 931–937. [Google Scholar] [CrossRef]
- Kacálková, L.; Tlustoš, P.; Száková, J. Phytoextraction of risk elements by willow and poplar trees. Int. J. Phytoremediation 2015, 17, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Pulford, I. Phytoremediation of heavy metal-contaminated land by trees—A review. Environ. Int. 2003, 29, 529–540. [Google Scholar] [CrossRef]
- Ruttens, A.; Boulet, J.; Weyens, N.; Smeets, K.; Adriaensen, K.; Meers, E.; van Slycken, S.; Tack, F.; Meiresonne, L.; Thewys, T.; et al. Short rotation coppice culture of willows and poplars as energy crops on metal contaminated agricultural soils. Int. J. Phytoremediation 2011, 13 (Suppl. 1), 194–207. [Google Scholar] [CrossRef]
- Riddell-Black, D. Heavy metal uptake by fast growing willow species. In A Biological Purification System: Conference and Workshop: Papers; Worldcat; Aronsson, P., Perttu, K., Eds.; Swedish University of Agricultural Sciences, Uppsala (Sweden) Department of Ecology and Environmental Research: Uppsala, Sweden, 1994; p. 231. ISSN 0282-6267. [Google Scholar]
- Punshon, T.; Lepp, N.W.; Dickinson, N.M. Resistance to copper toxicity in some British willows. J. Geochem. Explor. 1995, 52, 259–266. [Google Scholar] [CrossRef]
- Dickinson, N.M.; Punshon, T.; Hodkinson, R.B.; Lepp, N.W. Metal tolerance and accumulation in willows. In A Biological Purification System: Conference and Workshop: Papers; Worldcat; Aronsson, P., Perttu, K., Eds.; Swedish University of Agricultural Sciences, Uppsala (Sweden) Department of Ecology and Environmental Research: Uppsala, Sweden, 1994; p. 231. ISSN 0282-6267. [Google Scholar]
- Greger, M.; Landberg, T. Use of Willow in Phytoextraction. Int. J. Phytoremediation 1999, 1, 115–123. [Google Scholar] [CrossRef]
- Pilipović, A.; Zalesny, R.S.; Rončević, S.; Nikolić, N.; Orlović, S.; Beljin, J.; Katanić, M. Growth, physiology, and phytoextraction potential of poplar and willow established in soils amended with heavy-metal contaminated, dredged river sediments. J. Environ. Manag. 2019, 239, 352–365. [Google Scholar] [CrossRef]
- Vysloužilová, M.; Tlustoš, P.; Száková, J.; Pavlíková, D. As, Cd, Pb and Zn uptake by Salix spp. clones grown in soils enriched by high loads of these elements. Plant Soil Environ. 2011, 49, 191–196. [Google Scholar] [CrossRef]
- Yang, W.; Zhao, F.; Ding, Z.; Wang, Y.; Zhang, X.; Zhu, Z.; Yang, X. Variation of tolerance and accumulation to excess iron in 24 willow clones: Implications for phytoextraction. Int. J. Phytoremediation 2018, 20, 1284–1291. [Google Scholar] [CrossRef]
- Dimitriou, I.; Mola-Yudego, B.; Aronsson, P.; Eriksson, J. Changes in Organic Carbon and Trace Elements in the Soil of Willow Short-Rotation Coppice Plantations. Bioenergy Res. 2012, 5, 563–572. [Google Scholar] [CrossRef]
- Dimitriou, I.; Baum, C.; Baum, S.; Busch, G.; Schulz, U.; Kohn, J.; Lamersdorf, N.; Leinweber, P.; Aronsson, P.; Weih, M.; et al. The impact of Short Rotation Coppice (SRC) cultivation on the environment. Landbauforsch. Volkenrode 2009, 59, 159–162. [Google Scholar]
- Klang-Westin, E.; Eriksson, J. Potential of Salix as phytoextractor for Cd on moderately contaminated soils. Plant Soil 2003, 249, 127–137. [Google Scholar] [CrossRef]
- Aronsson, P.; Perttu, K. Willow vegetation filters for wastewater treatment and soil remediation combined with biomass production. For. Chron. 2001, 77, 293–299. [Google Scholar] [CrossRef]
- Börjesson, P.; Berndes, G. The prospects for willow plantations for wastewater treatment in Sweden. Biomass Bioenergy 2006, 30, 428–438. [Google Scholar] [CrossRef]
- Dimitriou, I.; Mola-Yudego, B.; Aronsson, P. Impact of Willow Short Rotation Coppice on Water Quality. Bioenergy Res. 2012, 5, 537–545. [Google Scholar] [CrossRef]
- Urbaniak, M.; Wyrwicka, A.; Tołoczko, W.; Serwecińska, L.; Zieliński, M. The effect of sewage sludge application on soil properties and willow (Salix sp.) cultivation. Sci. Total Environ. 2017, 586, 66–75. [Google Scholar] [CrossRef]
- Dimitriou, I.; Eriksson, J.; Adler, A.; Aronsson, P.; Verwijst, T. Fate of heavy metals after application of sewage sludge and wood–ash mixtures to short-rotation willow coppice. Environ. Pollut. 2006, 142, 160–169. [Google Scholar] [CrossRef]
- Mao, R.; Zeng, D.-H.; Hu, Y.-L.; Li, L.-J.; Yang, D. Soil organic carbon and nitrogen stocks in an age-sequence of poplar stands planted on marginal agricultural land in Northeast China. Plant Soil 2010, 332, 277–287. [Google Scholar] [CrossRef]
- McIvor, I.R.; Douglas, G.B. Poplars and willows in hill country—Stabilising soils and storing carbon. Adv. Nutr. Manag. Gains Past Goals Future 2012, 25, 1–11. [Google Scholar]
- Coleman, M.D.; Isebrands, J.G.; Tolsted, D.N.; Tolbert, V.R. Comparing Soil Carbon of Short Rotation Poplar Plantations with Agricultural Crops and Woodlots in North Central United States. Environ. Manag. 2004, 33. [Google Scholar] [CrossRef]
- Mukherjee, A.; Coleman, M.D. Converting Conventional Agriculture to Poplar Bioenergy Crops: Soil Chemistry. Commun. Soil Sci. Plant Anal. 2020, 51, 364–379. [Google Scholar] [CrossRef]
- Lockwell, J.; Guidi, W.; Labrecque, M. Soil carbon sequestration potential of willows in short-rotation coppice established on abandoned farm lands. Plant. Soil 2012, 360, 299–318. [Google Scholar] [CrossRef]
- Langeveld, H.; Quist-Wessel, F.; Dimitriou, I.; Aronsson, P.; Baum, C.; Schulz, U.; Bolte, A.; Baum, S.; Köhn, J.; Weih, M.; et al. Assessing Environmental Impacts of Short Rotation Coppice (SRC) Expansion: Model Definition and Preliminary Results. Bioenergy Res. 2012, 5, 621–635. [Google Scholar] [CrossRef]
- Acharya, B.S.; Blanco-Canqui, H. Lignocellulosic-based bioenergy and water quality parameters: A review. GCB Bioenergy 2018, 10, 504–533. [Google Scholar] [CrossRef]
- Baum, C.; Leinweber, P.; Weih, M.; Lamersdorf, N.; Dimitriou, I. Effects of short rotation coppice with willows and poplar on soil ecology. Landbauforsch. vTI Agric. Res. 2009, 59, 183–196. [Google Scholar]
- Weissteiner, C.J.; García-Feced, C.; Paracchini, M.L. A new view on EU agricultural landscapes: Quantifying patchiness to assess farmland heterogeneity. Ecol. Indic. 2016, 61, 317–327. [Google Scholar] [CrossRef]
- Shortall, O.K. “Marginal land” for energy crops: Exploring definitions and embedded assumptions. Energy Policy 2013, 62, 19–27. [Google Scholar] [CrossRef]
- Leather, S.R. “Ecological Armageddon”—More evidence for the drastic decline in insect numbers: Insect declines. Ann. Appl. Biol. 2018, 172, 1–3. [Google Scholar] [CrossRef]
- Simmons, B.I.; Balmford, A.; Bladon, A.J.; Christie, A.P.; De Palma, A.; Dicks, L.V.; Gallego-Zamorano, J.; Johnston, A.; Martin, P.A.; Purvis, A.; et al. Worldwide insect declines: An important message, but interpret with caution. Ecol. Evol. 2019, 9, 3678–3680. [Google Scholar] [CrossRef]
- Shortall, C.R.; Moore, A.; Smith, E.; Hall, M.J.; Woiwod, I.P.; Harrington, R. Long-term changes in the abundance of flying insects. Insect Conserv. Divers. 2009, 2, 251–260. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef]
- van Klink, R.; Bowler, D.E.; Gongalsky, K.B.; Swengel, A.B.; Gentile, A.; Chase, J.M. Meta-analysis reveals declines in terrestrial but increases in freshwater insect abundances. Science 2020, 368, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Bhattacharya, M.; Primack, R.B.; Gerwein, J. Are roads and railroads barriers to bumblebee movement in a temperate suburban conservation area? Biol. Conserv. 2003, 109, 37–45. [Google Scholar] [CrossRef]
- Keller, I.; Nentwig, W.; Largiadèr, C.R. Recent habitat fragmentation due to roads can lead to significant genetic differentiation in an abundant flightless ground beetle: Differentiation due to habitat fragmentation. Mol. Ecol. 2004, 13, 2983–2994. [Google Scholar] [CrossRef]
- Keller, I.; Excoffier, L.; Largiader, C.R. Estimation of effective population size and detection of a recent population decline coinciding with habitat fragmentation in a ground beetle. J. Evol. Biol. 2005, 18, 90–100. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botias, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- van Lexmond, M.B.; Bonmatin, J.-M.; Goulson, D.; Noome, D.A. Worldwide integrated assessment on systemic pesticides: Global collapse of the entomofauna: Exploring the role of systemic insecticides. Environ. Sci. Pollut. Res. 2015, 22, 1–4. [Google Scholar] [CrossRef]
- Stokstad, E. Field Research on Bees Raises Concern about Low-Dose Pesticides. Science 2012, 335, 1555. [Google Scholar] [CrossRef]
- Wallisdevries, M.F.; Van Swaay, C.A.M. Global warming and excess nitrogen may induce butterfly decline by microclimatic cooling. Glob. Chang. Biol. 2006, 12, 1620–1626. [Google Scholar] [CrossRef]
- Müller-Kroehling, S.; Jantsch, M.; Fischer, H.; Fischer, A. Modelling the effects of global warming on the ground beetle (Coleoptera: Carabidae) fauna of beech forests in Bavaria, Germany. Eur. J. Entomol. 2014, 111, 35–49. [Google Scholar] [CrossRef]
- Blanco, H.; Lal, R. Principles of Soil Conservation and Management; Springer: Dordrecht, The Netherlands, 2010; ISBN 978-1-4020-8709-7. [Google Scholar]
- Fry, D.A.; Slater, F.M. Early rotation short rotation willow coppice as a winter food resource for birds. Biomass Bioenergy 2011, 35, 2545–2553. [Google Scholar] [CrossRef]
- Sage, R.B.; Robertson, P.A. Factors affecting songbird communities using new short rotation coppice habitats in spring. Bird Study 1996, 43, 201–213. [Google Scholar] [CrossRef]
- Londo, M.; Dekker, J.; Terkeurs, W. Willow short-rotation coppice for energy and breeding birds: An exploration of potentials in relation to management. Biomass Bioenergy 2005, 28, 281–293. [Google Scholar] [CrossRef]
- Sage, R.; Cunningham, M.; Boatman, N. Birds in willow short-rotation coppice compared to other arable crops in central England and a review of bird census data from energy crops in the UK: Birds in short-rotation coppice. IBIS 2006, 148, 184–197. [Google Scholar] [CrossRef]
- Riffell, S.; Verschuyl, J.; Miller, D.; Wigley, T.B. A meta-analysis of bird and mammal response to short-rotation woody crops: Meta-analysis of bird and mammal response. GCB Bioenergy 2011, 3, 313–321. [Google Scholar] [CrossRef]
- Berg, Å. Breeding birds in short-rotation coppices on farmland in central Sweden—The importance of Salix height and adjacent habitats. Agric. Ecosyst. Environ. 2002, 90, 265–276. [Google Scholar] [CrossRef]
- Christian, D.P.; Hoffman, W.; Hanowski, J.M.; Niemi, G.J.; Beyea, J. Bird and mammal diversity on woody biomass plantations in North America. In Biomass and Bioenergy; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Campbell, S.P.; Frair, J.L.; Gibbs, J.P.; Volk, T.A. Use of short-rotation coppice willow crops by birds and small mammals in central New York. Biomass Bioenergy 2012, 47, 342–353. [Google Scholar] [CrossRef]
- Baxter, D.A.; Sage, R.B.; Hall, D.O. A methodology for assessing gamebird use of short rotation coppice. Biomass Bioenergy 1996, 10, 301–306. [Google Scholar] [CrossRef]
- Dhondt, A.A.; Wrege, P.H.; Sydenstricker, K.V.; Cerretani, J. Clone preference by nesting birds in short-rotation coppice plantations in central and western New York. Biomass Bioenergy 2004, 27, 429–435. [Google Scholar] [CrossRef]
- Dhondt, A.A.; Wrege, P.H.; Cerretani, J.; Sydenstricker, K.V. Avian species richness and reproduction in short-rotation coppice habitats in central and western New York. Bird Study 2007, 54, 12–22. [Google Scholar] [CrossRef]
- Göransson, G. Bird fauna of cultivated energy shrub forests at different heights. Biomass Bioenergy 1994, 6, 49–52. [Google Scholar] [CrossRef]
- Hanowski, J.M.; Niemi, G.J.; Christian, D.C. Influence of Within-Plantation Heterogeneity and Surrounding Landscape Composition on Avian Communities in Hybrid Poplar Plantations. Influencia de la Heterogeneidad Intra-plantacion y de la Composicion del Paisaje Circundante Sobre Comunidades de Aves en Plantaciones de Alamo Hibrido. Conserv. Biol. 1997, 11, 936–944. [Google Scholar] [CrossRef]
- Wilson, J. The breeding bird community of willow scrub at Leighton Moss, Lancashire. Bird Study 1978, 25, 239–244. [Google Scholar] [CrossRef]
- Bergström, R.; Guillet, C. Summer browsing by large herbivores in short-rotation willow plantations. Biomass Bioenergy 2002, 23, 27–32. [Google Scholar] [CrossRef]
- Christian, D.P. Wintertime use of hybrid poplar plantations by deer and medium-sized mammals in the midwestern U.S. Biomass Bioenergy 1997, 12, 35–40. [Google Scholar] [CrossRef]
- Christian, D.P.; Niemi, G.J.; Hanowski, J.M.; Collins, P. Perspectives on biomass energy tree plantations and changes in habitat for biological organisms. Biomass Bioenergy 1994, 6, 31–39. [Google Scholar] [CrossRef]
- Giordano, M.; Alberto, M. Use by small mammals of short-rotation plantations in relation to their structure and isolation. Hystrix Ital. J. Mammal. 2010, 20. [Google Scholar] [CrossRef]
- Moser, B.W.; Pipas, M.J.; Witmer, G.W.; Engeman, R.M. Small mammal use of hybrid poplar plantations relative to stand age. Northwest. Sci. 2002, 76, 158–165. [Google Scholar]
- Björkman, C.; Bommarco, R.; Eklund, K.; Höglund, S. Harvesting disrupts biological control of herbivores in a short-rotation coppice system. Ecol. Appl. 2004, 14, 1624–1633. [Google Scholar] [CrossRef]
- Langer, V. The potential of leys and short rotation coppice hedges as reservoirs for parasitoids of cereal aphids in organic agriculture. Agric. Ecosyst. Environ. 2001, 87, 81–92. [Google Scholar] [CrossRef]
- Minor, M.; Norton, R. Erratum: Effects of soil amendments on assemblages of soil mites (Acari: Oribatida, Mesostigmata) in short-rotation willow plantings in central New York. Can. J. For. Res. 2004, 34, 1417–1425. [Google Scholar] [CrossRef]
- Minor, M.A.; Volk, T.A.; Norton, R.A. Effects of site preparation techniques on communities of soil mites (Acari: Oribatida, Acari: Gamasida) under short-rotation forestry plantings in New York, USA. Appl. Soil Ecol. 2004, 25, 181–192. [Google Scholar] [CrossRef]
- Mueller, M.; Klein, A.-M.; Scherer-Lorenzen, M.; Nock, C.A.; Staab, M. Tree genetic diversity increases arthropod diversity in willow short rotation coppice. Biomass Bioenergy 2018, 108, 338–344. [Google Scholar] [CrossRef]
- Mueller, A.L.; Dauber, J. Hoverflies (Diptera: Syrphidae) benefit from a cultivation of the bioenergy crop Silphium perfoliatum L. (Asteraceae) depending on larval feeding type, landscape composition and crop management. Agric. For. Entomol. 2016, 18, 419–431. [Google Scholar] [CrossRef]
- Peacock, L.; Batley, J.; Dungait, J.; Barker, J.H.A.; Powers, S.; Karp, A. A comparative study of interspecies mating of Phratora vulgatissima and P. vitellinae using behavioural tests and molecular markers. Entomol. Exp. Appl. 2004, 110, 231–241. [Google Scholar] [CrossRef]
- Peacock, L.; Herrick, S.; Brain, P. Spatio-temporal dynamics of willow beetle (Phratora vulgatissima) in short-rotation coppice willows grown as monocultures or a genetically diverse mixture. Agric. For. Entomol. 1999, 1, 287–296. [Google Scholar] [CrossRef]
- Rowe, R.L.; Goulson, D.; Doncaster, C.P.; Clarke, D.J.; Taylor, G.; Hanley, M.E. Evaluating ecosystem processes in willow short rotation coppice bioenergy plantations. GCB Bioenergy 2013, 5, 257–266. [Google Scholar] [CrossRef]
- Sage, R.B.; Tucker, K. The distribution of Phratora vulgatissima (Coleoptera: Chrysomelidae) on cultivated willows in Britain and Ireland. For. Pathol 1998, 28, 289–296. [Google Scholar] [CrossRef]
- Sage, R.B.; Fell, D.; Tucker, K.; Sotherton, N.W. Post hibernation dispersal of three leaf-eating beetles (Coleoptera: Chrysomelidae) colonising cultivated willows and poplars. Agric. For. Entomol. 1999, 1, 61–70. [Google Scholar] [CrossRef]
- Reddersen, J. SRC-willow (Salixviminalis) as a resource for flower-visiting insects. Biomass Bioenergy 2001, 20, 171–179. [Google Scholar] [CrossRef]
- Kosewska, A.; Nietupski, M.; Agnieszka, L.-D.; Dolores, C. Biegaczowate (Col. Carabidae) zasiedlające uprawy wierzby krzewiastej w okolicach Olsztyna. Prog. Plant. Prot. 2010, 50, 1504–1510. [Google Scholar]
- Walerys, G.; Kosewska, A.; Wojciech, S. Uprawa wierzby krzewiastej Salix spp. miejscem bytowania drapieżnych biegaczowatych Carabidae. Fragm. Agron. 2008, XXV, 158–169. [Google Scholar]
- Konieczna, K.; Melke, A.; Olbrycht, T. Unexploited willow’s plantation (Salix viminalis) as a reservoir of epigeic ground beetles (Col., Carabidae) and rove beetles (Col., Staphylinidae). Prog. Plant. Prot. 2013, 53, 319–326. [Google Scholar]
- Weger, J.; Vávrová, K.; Kašparová, L.; Bubeník, J.; Komárek, A. The influence of rotation length on the biomass production and diversity of ground beetles (Carabidae) in poplar short rotation coppice. Biomass Bioenergy 2013, 54, 284–292. [Google Scholar] [CrossRef]
- Müller-Kroehling, S.; Hohmann, G.; Helbig, C.; Liesebach, M.; Lübke-Al Hussein, M.; Al Hussein, I.A.; Burmeister, J.; Jantsch, M.C.; Zehlius-Eckert, W.; Müller, M. Biodiversity functions of short rotation coppice stands—Results of a meta study on ground beetles (Coleoptera: Carabidae). Biomass Bioenergy 2020, 132, 105416. [Google Scholar] [CrossRef]
- Burel, F. Landscape structure effects on carabid beetles spatial patterns in western France. Landsc. Ecol 1989, 2, 215–226. [Google Scholar] [CrossRef]
- Czerniakowski, Z.W.; Olbrycht, T. Ground beetles (Coleoptera, Carabidae) in the short-rotation biomass plantations. Zesz. Nauk. Południowo Wschod. Oddziału Pol. Tow. Inżynierii Ekol. Z Siedzibą W Rzesz. I Pol. Tow. Glebozn. Oddział W Rzesz. 2009, 11, 39–42. [Google Scholar]
- Allegro, G.; Sciaky, R. Assessing the potential role of ground beetles (Coleoptera, Carabidae) as bioindicators in poplar stands, with a newly proposed ecological index (FAI). For. Ecol. Manag. 2003, 175, 275–284. [Google Scholar] [CrossRef]
- Brauner, O.; Schulz, U. Ground beetle communities (Carabidae) on short rotation coppices and adjacent crop areas—Investigations in Saxony and Brandenburg. Entomol. Blaetter Fuer Biol. Und Syst. Der Kaefer 2011, 107, 31–64. [Google Scholar]
- Sage, R.; Cunningham, M.; Haughton, A.J.; Mallott, M.D.; Bohan, D.A.; Riche, A.; Karp, A. The environmental impacts of biomass crops: Use by birds of miscanthus in summer and winter in southwestern England: Birds in miscanthus. IBIS 2010, 152, 487–499. [Google Scholar] [CrossRef]
- Holland, J. Carabid Beetles: Their Ecology, Survival and Use in Agroecosystems. In The Agroecology of Carabid Beetles; Intercept: New York, NY, USA, 2002; pp. 1–40. ISBN 1-898298-76-9. [Google Scholar]
- Schulz, U.; Brauner, O.; Gruss, H. Animal diversity on short-rotation coppices—A review. Vti Agric. Res. 2009, 3, 171–182. [Google Scholar]
- Kromp, B. Carabid beetles in sustainable agriculture: A review on pest control efficacy, cultivation impacts and enhancement. Agric. Ecosyst. Environ. 1999, 74, 187–228. [Google Scholar] [CrossRef]
- Regulska, E. Carabidae in landscape research on the basis on literature, 2005–2008. Pol. J. Environ. Stud. 2011, 20, 733–741. [Google Scholar]
- Kotze, D.J.; Brandmayr, P.; Casale, A.; Dauffy-Richard, E.; Dekoninck, W.; Koivula, M.J.; Lövei, G.L.; Mossakowski, D.; Noordjik, J.; Paarmann, W.; et al. Forty years of carabid beetle research in Europe—From taxonomy, biology, ecology and population studies to bioindication, habitat assessment and conservation. ZooKeys 2011, 100, 55–148. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Booij, C.J.H.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. B 2006, 273, 1715–1727. [Google Scholar] [CrossRef]
- Thiele, H.-U. Carabid Beetles in Their Environments: A Study on Habitat Selection by Adaptations in Physiology and Behaviour; Springer: Berlin/Heidelberg, Germany, 1977; ISBN 978-3-642-81154-8. [Google Scholar]
- Szyszko, J. Próba Waloryzacji Środowisk Leśnych Przy Pomocy Biegaczowatych (Carabidea, Col.) Waloryzacja Ekosystemów Leśnych Metodami Bioidnykacyjnymi.; SGGW: Warszawa, Poland, 1997. [Google Scholar]
- Kulkarmi, S.; Dosdall, L.; Willenborg, C. The Role of Ground Beetles (Coleoptera: Carabidae) in Weed Seed Consumption: A Review. Weed Sci. 2015, 63, 150304133636004. [Google Scholar] [CrossRef]
- Frei, B.; Guenay, Y.; Bohan, D.A.; Traugott, M.; Wallinger, C. Molecular analysis indicates high levels of carabid weed seed consumption in cereal fields across Central Europe. J. Pest. Sci 2019, 92, 935–942. [Google Scholar] [CrossRef]
- Huruk, S. Comparison of structure of carabid (Coleoptera: Carabidae) communities of hay meadows and adjacent cultivated fields. Wiadomości Entomol. 2006, 25, 9–32. [Google Scholar]
- Nakamura, M.; Kagata, H.; Ohgushi, T. Trunk cutting initiates bottom-up cascades in a tri-trophic system: Sprouting increases biodiversity of herbivorous and predaceous arthropods on willows. Oikos 2006, 113, 259–268. [Google Scholar] [CrossRef]
- Baum, S.; Weih, M.; Busch, G.; Kroiher, F.; Bolte, A. The impact of Short Rotation Coppice plantations on phytodiversity. Landbauforsch Volkenrode 2009, 59, 163–170. [Google Scholar]
- Briones, M.J.I.; Elias, D.M.O.; Grant, H.K.; McNamara, N.P. Plant identity control on soil food web structure and C transfers under perennial bioenergy plantations. Soil Biol. Biochem. 2019, 138, 107603. [Google Scholar] [CrossRef]
- Kahle, P.; Janssen, M. Impact of short-rotation coppice with poplar and willow on soil physical properties. J. Plant Nutr. Soil Sci. 2020, 183, 119–128. [Google Scholar] [CrossRef]
- Boháč, J.; Celjak, I.; Moudrý, J.; Kohout, P.; Wotavová, K. Communities of beetles in plantations of fast growing plant species for energetic purposes. Entomol.Rom. 2007, 12, 213–221. [Google Scholar]
- Sotherton, N.W. The distribution and abundance of predatory Coleoptera overwintering in field boundaries. Ann. Appl. Biol. 1985, 106, 17–21. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Szczukowski, S.; Tworkowski, J.; Klasa, A. Yield, energy parameters and chemical composition of short-rotation willow biomass. Ind. Crop. Prod. 2013, 46, 60–65. [Google Scholar] [CrossRef]
- Graham, J.B.; Nassauer, J.I. Wild bee abundance in temperate agroforestry landscapes: Assessing effects of alley crop composition, landscape configuration, and agroforestry area. Agroforest Syst 2019, 93, 837–850. [Google Scholar] [CrossRef]
- Vanbeveren, S.; Ceulemans, R. Biodiversity in short-rotation coppice. Renew. Sustain. Energy Rev. 2019, 111, 34–43. [Google Scholar] [CrossRef]
- Damschen, E.I. Landscape Corridors. In Encyclopedia of Biodiversity; Elsevier: Amsterdam, The Netherlands, 2013; pp. 467–475. ISBN 978-0-12-384720-1. [Google Scholar]
- Rogan, J.E.; Lacher, T.E. Impacts of Habitat Loss and Fragmentation on Terrestrial Biodiversity. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2018; p. B9780124095489110000. ISBN 978-0-12-409548-9. [Google Scholar]
- Fahrig, L. Why do several small patches hold more species than few large patches? Glob. Ecol. Biogeogr. 2020, 29, 615–628. [Google Scholar] [CrossRef]
- Rybicki, J.; Abrego, N.; Ovaskainen, O. Habitat fragmentation and species diversity in competitive communities. Ecol. Lett. 2020, 23, 506–517. [Google Scholar] [CrossRef]
- Tjørve, E. How to resolve the SLOSS debate: Lessons from species-diversity models. J. Theor. Biol. 2010, 264, 604–612. [Google Scholar] [CrossRef]
- Cartwright, J. Ecological islands: Conserving biodiversity hotspots in a changing climate. Front. Ecol. Env. 2019, fee.2058. [Google Scholar] [CrossRef]
- Losos, J.B.; Ricklefs, R.E.; MacArthur, R.H. The Theory of Island Biogeography Revisited; Princeton University Press: Princeton, NJ, USA, 2010; ISBN 978-1-4008-3192-0. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 2001; ISBN 978-0-691-08836-5. [Google Scholar]
- Banaszak, J. Wyspy Środowiskowe: Bioróżnorodność i Próby Typologii; Akademia Bydgoska im. Kazimierza Wielkiego: Bydgoszcz, Poland, 2002; ISBN 978-83-7096-459-7. [Google Scholar]
- Warren, B.H.; Simberloff, D.; Ricklefs, R.E.; Aguilée, R.; Condamine, F.L.; Gravel, D.; Morlon, H.; Mouquet, N.; Rosindell, J.; Casquet, J.; et al. Islands as model systems in ecology and evolution: Prospects fifty years after MacArthur-Wilson. Ecol. Lett. 2015, 18, 200–217. [Google Scholar] [CrossRef] [PubMed]
- Helmus, M.R.; Behm, J.E. Island Biogeography Revisited. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 978-0-12-409548-9. [Google Scholar]
- Corbet, S.A. Insects, plants and succession: Advantages of long-term set-aside. Agric. Ecosyst. Environ. 1995, 53, 201–217. [Google Scholar] [CrossRef]
- Staley, J.T.; Sparks, T.H.; Croxton, P.J.; Baldock, K.C.R.; Heard, M.S.; Hulmes, S.; Hulmes, L.; Peyton, J.; Amy, S.R.; Pywell, R.F. Long-term effects of hedgerow management policies on resource provision for wildlife. Biol. Conserv. 2012, 145, 24–29. [Google Scholar] [CrossRef]
- Wehling, S.; Diekmann, M. Importance of hedgerows as habitat corridors for forest plants in agricultural landscapes. Biol. Conserv. 2009, 142, 2522–2530. [Google Scholar] [CrossRef]
- Londo, M.; Roose, M.; Dekker, J.; de Graaf, H. Willow short-rotation coppice in multiple land-use systems: Evaluation of four combination options in the Dutch context. Biomass Bioenergy 2004, 27, 205–221. [Google Scholar] [CrossRef]
- Homburg, K.; Drees, C.; Boutaud, E.; Nolte, D.; Schuett, W.; Zumstein, P.; Ruschkowski, E.; Assmann, T. Where have all the beetles gone? Long-term study reveals carabid species decline in a nature reserve in Northern Germany. Insect Conserv. Divers. 2019, icad.12348. [Google Scholar] [CrossRef]
- Maes, D.; Titeux, N.; Hortal, J.; Anselin, A.; Decleer, K.; De Knijf, G.; Fichefet, V.; Luoto, M. Predicted insect diversity declines under climate change in an already impoverished region. J. Insect Conserv. 2010, 14, 485–498. [Google Scholar] [CrossRef]
- Wilson, R.J.; Maclean, I.M.D. Recent evidence for the climate change threat to Lepidoptera and other insects. J. Insect Conserv. 2011, 15, 259–268. [Google Scholar] [CrossRef]
- Wagner, D.L. Insect Declines in the Anthropocene. Annu. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef]
- Didham, R.K.; Basset, Y.; Collins, C.M.; Leather, S.R.; Littlewood, N.A.; Menz, M.H.M.; Müller, J.; Packer, L.; Saunders, M.E.; Schönrogge, K.; et al. Interpreting insect declines: Seven challenges and a way forward. Insect Conserv Divers. 2020, 13, 103–114. [Google Scholar] [CrossRef]
- Forister, M.L.; Pelton, E.M.; Black, S.H. Declines in insect abundance and diversity: We know enough to act now. Conserv. Sci Pr. 2019, 1. [Google Scholar] [CrossRef]
- Eyre, M.D.; McMillan, S.D.; Critchley, C.N.R. Ground beetles (Coleoptera, Carabidae) as indicators of change and pattern in the agroecosystem: Longer surveys improve understanding. Ecol. Indic. 2016, 68, 82–88. [Google Scholar] [CrossRef]
- Jowett, K.; Milne, A.E.; Metcalfe, H.; Hassall, K.L.; Potts, S.G.; Senapathi, D.; Storkey, J. Species matter when considering landscape effects on carabid distributions. Agric. Ecosyst. Environ. 2019, 285, 106631. [Google Scholar] [CrossRef]
- Zaller, J.G.; Moser, D.; Drapela, T.; Frank, T. Ground-dwelling predators can affect within-field pest insect emergence in winter oilseed rape fields. BioControl 2009, 54, 247–253. [Google Scholar] [CrossRef]
- Suenaga, H.; Hamamura, T. Occurrence of carabid beetles (Coleoptera: Carabidae) in cabbage fields and their possible impact on lepidopteran pests. Appl. Entomol. Zool. 2001, 36, 151–160. [Google Scholar] [CrossRef]
- Vichitbandha, P.; Wise, D.H. A field experiment on the effectiveness of spiders and carabid beetles as biocontrol agents in soybean. Agric. For. Ent 2002, 4, 31–38. [Google Scholar] [CrossRef]
- Warner, D.J.; Allen-Williams, L.J.; Warrington, S.; Ferguson, A.W.; Williams, I.H. Implications for conservation biocontrol of spatio-temporal relationships between carabid beetles and coleopterous pests in winter oilseed rape. Agric. For. Entomol. 2008, 10, 375–387. [Google Scholar] [CrossRef]
- Schlein, O.; Buechs, W. Approaches to assess the importance of carnivorous beetles as predators of oilseed rape pests. Iobc Wprs Bull. 2004, 27, 289–292. [Google Scholar]
- Schmidt, M.H.; Lauer, A.; Purtauf, T.; Thies, C.; Schaefer, M.; Tscharntke, T. Relative importance of predators and parasitoids for cereal aphid control. Proc. R. Soc. Lond. B 2003, 270, 1905–1909. [Google Scholar] [CrossRef]
- Woodcock, B.A.; Harrower, C.; Redhead, J.; Edwards, M.; Vanbergen, A.J.; Heard, M.S.; Roy, D.B.; Pywell, R.F. National patterns of functional diversity and redundancy in predatory ground beetles and bees associated with key UK arable crops. J. Appl. Ecol. 2014, 51, 142–151. [Google Scholar] [CrossRef]
- Honek, A.; Martinkova, Z.; Jarosik, V. Ground beetles (Carabidae) as seed predators. Eur. J. Entomol. 2003, 100, 531–544. [Google Scholar] [CrossRef]
- Menalled, F.D.; Smith, R.G.; Dauer, J.T.; Fox, T.B. Impact of agricultural management on carabid communities and weed seed predation. Agric. Ecosyst. Environ. 2007, 118, 49–54. [Google Scholar] [CrossRef]
- Sasakawa, K. Field Observations of Climbing Behavior and Seed Predation by Adult Ground Beetles (Coleoptera: Carabidae) in a Lowland Area of the Temperate Zone: Table 1. Environ. Entomol. 2010, 39, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Pannwitt, H.; Westerman, P.R.; de Mol, F.; Selig, C.; Gerowitt, B. Biological control of weed patches by seed predators; responses to seed density and exposure time. Biol. Control. 2017, 108, 1–8. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| References | Investigated Issues | Local. | Plant. Type | Plant. Age | Canopy Age | Adjacent Habitat | Years Date |
|---|---|---|---|---|---|---|---|
| [86] | 1. Species richness and diversity, dominance structure. 2. Community structure: trophic, habitat preference, humidity preference, development type. | Northeast Poland | willow | 8–9 years | Not given | none | 2004–2005 |
| [85] | 1. Species richness and diversity, dominance structure. 2. Community structure: trophic, habitat preference, humidity preference, development type. | Northeast Poland | willow | 1 to 3 years | 1 to 3 years | none | 2005–2006 |
| [87] | 1. Species composition and abundance of ground beetles inhabiting unexploited willows plantation. 2. Population ecological characteristic and dominance structure. 3. Margalef’s index, Shannon’ diversity, Evenness H/log(N) Pielou. | Southeast Poland | willow | not given | 8 and 9 years | none | 2011–2012 |
| [92] | 1. Species richness and diversity, dominance structure. 2. The similarity to natural woodlands. | Po Valley, Italy | poplar | 1 to 10 years | not given | natural woods, crops: maize, tobacco | 1989–1999 |
| [102] | 1. Influence of the plantation vicinity and anthropogenic factors on Carabidae assemblages. 2. The structure of beetle communities. | South Bohemia (Czech Republic) | willow | 2, 4, 6, 8 years | not given | Field with a stream; pond, field, alder trees and meadow; pasture, field, cultural forest; | 2007 |
| [81] | 1. Comparison of how predation processes by ground arthropods varied between short rotation coppice (SRC) willow bioenergy plantations and alternative land-uses: arable and set-aside. 2. Predation pressure investigations: prey removal assay coupled with pitfall traps and direct searches | North Nottinghamshire, England | willow | 1 to 10 years | 1 to 9 years | set-aside, arable | 2008 |
| [88] | 1. Simpson biodiversity index, evenness, level of anthropogenic influence 2. The influence of the length of rotation on biodiversity parameters; | Peklov, Czech Republic | poplar | 9 years | 1, 3, 6 years rotations | none | 2003–2008 |
| [89] | 1. Taxonomy and identification. 2. Species traits and categorization. 3. Habitat preferences. 4. Endangered species. 5. Dispersal of forest species; corridor function. 6. Species traits concerning age of the SRC versus age of the SRC standing crop. 7. Factors influencing SRC biodiversity functions. | Germany (different sites) and the Czech Republic | willow poplar | 1 to 23 years | 1 to 9 years | different | Meta-study |
| Investigated Plots and Year of Investigation | Plantation Age (Years) | Canopy Age (Years) | Number of Forest Species (including ED and D) |
|---|---|---|---|
| [86] 2004 | 8 | Not given | 5 (1 ED) |
| [86] 2005 | 9 | Not given | 4 (2 ED) |
| [85] 2005 W1 | 2 | 2 | 2 (2 ED) |
| [85] 2006 W1 | 3 | 3 | 2 |
| [85] 2005 W2 | 1 | 1 | 2 (2 ED) |
| [85] 2006 W2 | 2 | 2 | 2 (1 ED) |
| [87] 2011 | Not given | 8 | 7 (1 ED) |
| [87] 2012 | Not given | 9 | 5 (1 ED) |
| [92] 1989 P1 | 2 | Not given | 5 |
| [92] 1999 P2 | 6 | Not given | 3 (1 ED, 1 D) |
| [92] 1999 P3 | 6 | Not given | 2 (1 D) |
| [92] 1991 P4 | 10 | Not given | 4 (2 ED) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piotrowska, N.S.; Czachorowski, S.Z.; Stolarski, M.J. Ground Beetles (Carabidae) in the Short-Rotation Coppice Willow and Poplar Plants—Synergistic Benefits System. Agriculture 2020, 10, 648. https://doi.org/10.3390/agriculture10120648
Piotrowska NS, Czachorowski SZ, Stolarski MJ. Ground Beetles (Carabidae) in the Short-Rotation Coppice Willow and Poplar Plants—Synergistic Benefits System. Agriculture. 2020; 10(12):648. https://doi.org/10.3390/agriculture10120648
Chicago/Turabian StylePiotrowska, Natalia Stefania, Stanisław Zbigniew Czachorowski, and Mariusz Jerzy Stolarski. 2020. "Ground Beetles (Carabidae) in the Short-Rotation Coppice Willow and Poplar Plants—Synergistic Benefits System" Agriculture 10, no. 12: 648. https://doi.org/10.3390/agriculture10120648
APA StylePiotrowska, N. S., Czachorowski, S. Z., & Stolarski, M. J. (2020). Ground Beetles (Carabidae) in the Short-Rotation Coppice Willow and Poplar Plants—Synergistic Benefits System. Agriculture, 10(12), 648. https://doi.org/10.3390/agriculture10120648