Genetic Landscape of Nonobstructive Azoospermia and New Perspectives for the Clinic

 and
and 
Abstract
1. Introduction
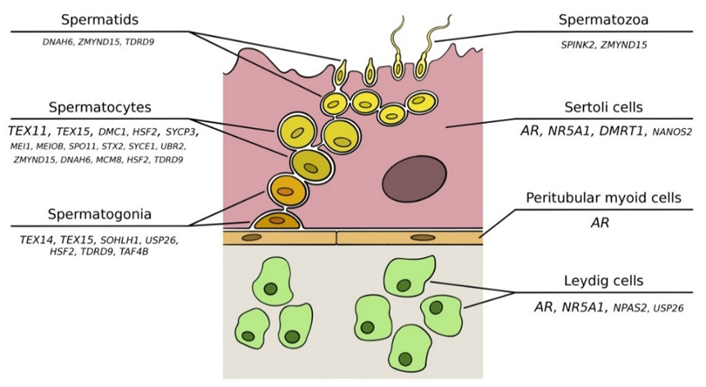
2. The Spermatogenic Process
3. Monogenic Causes of Nonobstructive Azoospermia
3.1. Karyotype Abnormalities
3.2. Y Chromosome Microdeletions
3.2.1. USP9Y
3.2.2. DDX3Y
3.2.3. EIF1AY
3.2.4. RPS4Y2
3.2.5. KDM5D
3.2.6. DAZ
3.3. Autosomal Monogenic Factors
3.3.1. AR
3.3.2. NR5A1
3.3.3. DMRT1
3.3.4. TEX11
3.3.5. TEX14 and TEX15
3.3.6. NPAS2
3.3.7. Other Putative Nonobstructive Azoospermia Genes
4. Common Variation Associated with Susceptibility to Nonobstructive Azoospermia
4.1. Candidate Gene Approach
4.2. Insights from Large-Scale Genetic Studies
4.2.1. Pilot GWAS in Europeans
4.2.2. First GWAS in East Asians
4.2.3. Second GWAS in East Asians
4.2.4. Fine-Mapping of the Major Histocompatibility Complex Region
4.2.5. GWAS in Hutterites
4.2.6. Other Genome-Wide Approaches
5. Clinical Relevance of the Genetic Studies of Nonobstructive Azoospermia
6. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Mascarenhas, M.N.; Flaxman, S.R.; Boerma, T.; Vanderpoel, S.; Stevens, G.A. National, regional, and global trends in infertility prevalence since 1990: A systematic analysis of 277 health surveys. PLoS Med. 2012, 9, e1001356. [Google Scholar] [CrossRef] [PubMed]
- Chandra, A.; Copen, C.E.; Stephen, E.H. Infertility and impaired fecundity in the United States, 1982–2010: Data from the National Survey of Family Growth. Natl. Health Stat. Rep. 2013, 67, 1–18. [Google Scholar]
- Jarow, J.P.; Espeland, M.A.; Lipshultz, L.I. Evaluation of the azoospermic patient. J. Urol. 1989, 142, 62–65. [Google Scholar] [CrossRef]
- Thonneau, P.; Marchand, S.; Tallec, A.; Ferial, M.L.; Ducot, B.; Lansac, J.; Lopes, P.; Tabaste, J.M.; Spira, A. Incidence and main causes of infertility in a resident population (1,850,000) of three French regions (1988–1989). Hum. Reprod. 1991, 6, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Mazzilli, F.; Rossi, T.; Delfino, M.; Sarandrea, N.; Dondero, F. Azoospermia: Incidence, and biochemical evaluation of seminal plasma by the differential pH method. Panminerva Med. 2000, 42, 27–31. [Google Scholar] [PubMed]
- Tuttelmann, F.; Werny, F.; Cooper, T.G.; Kliesch, S.; Simoni, M.; Nieschlag, E. Clinical experience with azoospermia: Aetiology and chances for spermatozoa detection upon biopsy. Int. J. Androl. 2011, 34, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef]
- Esteves, S.C. Clinical management of infertile men with nonobstructive azoospermia. Asian J. Androl. 2015, 17, 459–470. [Google Scholar] [CrossRef]
- Tournaye, H.; Krausz, C.; Oates, R.D. Novel concepts in the aetiology of male reproductive impairment. Lancet Diabetes Endocrinol. 2017, 5, 544–553. [Google Scholar] [CrossRef]
- Krausz, C.; Riera-Escamilla, A. Genetics of male infertility. Nat. Rev. Urol. 2018, 15, 369–384. [Google Scholar] [CrossRef]
- Brennan, J.; Capel, B. One tissue, two fates: Molecular genetic events that underlie testis versus ovary development. Nat. Rev. Genet. 2004, 5, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Phillips, B.T.; Gassei, K.; Orwig, K.E. Spermatogonial stem cell regulation and spermatogenesis. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 1663–1678. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.J.; DeFalco, T. Role of the testis interstitial compartment in spermatogonial stem cell function. Reproduction 2017, 153, R151–R162. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yan, M.; Ge, R.; Cheng, C.Y. Crosstalk between Sertoli and Germ Cells in Male Fertility. Trends Mol. Med. 2019. [Google Scholar] [CrossRef]
- Fayomi, A.P.; Orwig, K.E. Spermatogonial stem cells and spermatogenesis in mice, monkeys and men. Stem Cell Res. 2018, 29, 207–214. [Google Scholar] [CrossRef]
- Larose, H.; Shami, A.N.; Abbott, H.; Manske, G.; Lei, L.; Hammoud, S.S. Gametogenesis: A journey from inception to conception. Curr. Top. Dev. Biol. 2019, 132, 257–310. [Google Scholar]
- Shiraishi, K. Hormonal therapy for non-obstructive azoospermia: Basic and clinical perspectives. Reprod. Med. Biol. 2015, 14, 65–72. [Google Scholar] [CrossRef]
- Younes, A.H.; Hamed, H.B.; Mohamed, E.M.; Makki, M.A.; Gaber, N.; Mohamed, H.M. Oestrogen receptors beta genotype in infertile Egyptian men with nonobstructive azoospermia. Andrologia 2016, 48, 1289–1293. [Google Scholar] [CrossRef]
- Schultz, N.; Hamra, F.K.; Garbers, D.L. A multitude of genes expressed solely in meiotic or postmeiotic spermatogenic cells offers a myriad of contraceptive targets. Proc. Natl. Acad. Sci. USA 2003, 100, 12201–12206. [Google Scholar] [CrossRef]
- Gunes, S.; Al-Sadaan, M.; Agarwal, A. Spermatogenesis, DNA damage and DNA repair mechanisms in male infertility. Reprod. Biomed. Online 2015, 31, 309–319. [Google Scholar] [CrossRef]
- Robles, V.; Herraez, P.; Labbe, C.; Cabrita, E.; Psenicka, M.; Valcarce, D.G.; Riesco, M.F. Molecular basis of spermatogenesis and sperm quality. Gen. Comp. Endocrinol. 2017, 245, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Jungwirth, A.; Giwercman, A.; Tournaye, H.; Diemer, T.; Kopa, Z.; Dohle, G.; Krausz, C.; European Association of Urology Working Group on Male Infertility. European Association of Urology guidelines on Male Infertility: The 2012 update. Eur. Urol. 2012, 62, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Berookhim, B.M.; Schlegel, P.N. Azoospermia due to spermatogenic failure. Urol. Clin. N. Am. 2014, 41, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Barratt, C.L.R.; Bjorndahl, L.; De Jonge, C.J.; Lamb, D.J.; Osorio Martini, F.; McLachlan, R.; Oates, R.D.; van der Poel, S.; St John, B.; Sigman, M.; et al. The diagnosis of male infertility: An analysis of the evidence to support the development of global WHO guidance-challenges and future research opportunities. Hum. Reprod. Update 2017, 23, 660–680. [Google Scholar] [CrossRef]
- Palermo, G.D.; Colombero, L.T.; Hariprashad, J.J.; Schlegel, P.N.; Rosenwaks, Z. Chromosome analysis of epididymal and testicular sperm in azoospermic patients undergoing ICSI. Hum. Reprod. 2002, 17, 570–575. [Google Scholar] [CrossRef]
- Chandley, A.C. Infertility and chromosome abnormality. Oxf. Rev. Reprod. Biol. 1984, 6, 1–46. [Google Scholar]
- Rivas, F.; Garcia-Esquivel, L.; Diaz, M.; Rivera, H.; Cantu, J.M. Cytogenetic evaluation of 163 azoospermics. J. Genet. Hum. 1987, 35, 291–298. [Google Scholar]
- Yatsenko, A.N.; Yatsenko, S.A.; Weedin, J.W.; Lawrence, A.E.; Patel, A.; Peacock, S.; Matzuk, M.M.; Lamb, D.J.; Cheung, S.W.; Lipshultz, L.I. Comprehensive 5-year study of cytogenetic aberrations in 668 infertile men. J. Urol. 2010, 183, 1636–1642. [Google Scholar] [CrossRef]
- Bojesen, A.; Gravholt, C.H. Klinefelter syndrome in clinical practice. Nat. Clin. Pract. Urol. 2007, 4, 192–204. [Google Scholar] [CrossRef]
- Denschlag, D.; Tempfer, C.; Kunze, M.; Wolff, G.; Keck, C. Assisted reproductive techniques in patients with Klinefelter syndrome: A critical review. Fertil. Steril. 2004, 82, 775–779. [Google Scholar] [CrossRef]
- Tiepolo, L.; Zuffardi, O. Localization of factors controlling spermatogenesis in the nonfluorescent portion of the human Y chromosome long arm. Hum. Genet. 1976, 34, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Vogt, P.H.; Edelmann, A.; Kirsch, S.; Henegariu, O.; Hirschmann, P.; Kiesewetter, F.; Köhn, F.M.; Schill, W.B.; Farah, S.; Ramos, C.; et al. Human Y chromosome azoospermia factors (AZF) mapped to different subregions in Yq11. Hum. Mol. Genet. 1996, 5, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Sharkey, A.; Kirsch, S.; Vogt, P.; Keil, R.; Hargreave, T.B.; McBeath, S.; Chandley, A.C. Towards the molecular localisation of the AZF locus: Mapping of microdeletions in azoospermic men within 14 subintervals of interval 6 of the human Y chromosome. Hum. Mol. Genet. 1992, 1, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Repping, S.; Skaletsky, H.; Lange, J.; Silber, S.; Van Der Veen, F.; Oates, R.D.; Page, D.C.; Rozen, S. Recombination between palindromes P5 and P1 on the human Y chromosome causes massive deletions and spermatogenic failure. Am. J. Hum. Genet. 2002, 71, 906–922. [Google Scholar] [CrossRef] [PubMed]
- Krausz, C.; Casamonti, E. Spermatogenic failure and the Y chromosome. Hum. Genet. 2017, 136, 637–655. [Google Scholar] [CrossRef] [PubMed]
- Kamp, C.; Hirschmann, P.; Voss, H.; Huellen, K.; Vogt, P.H. Two long homologous retroviral sequence blocks in proximal Yq11 cause AZFa microdeletions as a result of intrachromosomal recombination events. Hum. Mol. Genet. 2000, 9, 2563–2572. [Google Scholar] [CrossRef]
- Blanco, P.; Shlumukova, M.; Sargent, C.A.; Jobling, M.A.; Affara, N.; Hurles, M.E. Divergent outcomes of intrachromosomal recombination on the human Y chromosome: Male infertility and recurrent polymorphism. J. Med. Genet. 2000, 37, 752–758. [Google Scholar] [CrossRef]
- Sun, C.; Skaletsky, H.; Rozen, S.; Gromoll, J.; Nieschlag, E.; Oates, R.; Page, D.C. Deletion of azoospermia factor a (AZFa) region of human Y chromosome caused by recombination between HERV15 proviruses. Hum. Mol. Genet. 2000, 9, 2291–2296. [Google Scholar] [CrossRef]
- Kuroda-Kawaguchi, T.; Skaletsky, H.; Brown, L.G.; Minx, P.J.; Cordum, H.S.; Waterston, R.H.; Wilson, R.K.; Silber, S.; Oates, R.; Rozen, S.; et al. The AZFc region of the Y chromosome features massive palindromes and uniform recurrent deletions in infertile men. Nat. Genet. 2001, 29, 279–286. [Google Scholar] [CrossRef]
- Lo Giacco, D.; Chianese, C.; Sanchez-Curbelo, J.; Bassas, L.; Ruiz, P.; Rajmil, O.; Sarquella, J.; Vives, A.; Ruiz-Castañé, E.; Oliva, R.; et al. Clinical relevance of Y-linked CNV screening in male infertility: New insights based on the 8-year experience of a diagnostic genetic laboratory. Eur. J. Hum. Genet. 2014, 22, 754–761. [Google Scholar] [CrossRef]
- Ginalski, K.; Rychlewski, L.; Baker, D.; Grishin, N.V. Protein structure prediction for the male-specific region of the human Y chromosome. Proc. Natl. Acad. Sci. USA 2004, 101, 2305–2310. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, A.; Pellman, D. Deubiquitinating enzymes: A new class of biological regulators. Crit. Rev. Biochem. Mol. Biol. 1998, 33, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Vogt, P.H.; Falcao, C.L.; Hanstein, R.; Zimmer, J. The AZF proteins. Int. J. Androl. 2008, 31, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, S.J.; Ross, A.R.; Ma, K.; Cooke, H.J.; Intyre, M.A.; Chandley, A.C.; Hargreave, T.B. Polymerase chain reaction screening for Y chromosome microdeletions: A first step towards the diagnosis of genetically-determined spermatogenic failure in men. Mol. Hum. Reprod. 1996, 2, 775–779. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.M.; Furlong, R.A.; Sargent, C.A.; Erickson, R.P.; Longepied, G.; Mitchell, M.; Jones, M.H.; Hargreave, T.B.; Cooke, H.J.; Affara, N.A. Characterisation of the coding sequence and fine mapping of the human DFFRY gene and comparative expression analysis and mapping to the Sxrb interval of the mouse Y chromosome of the Dffry gene. Hum. Mol. Genet. 1998, 7, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Krausz, C.; Degl’Innocenti, S.; Nuti, F.; Morelli, A.; Felici, F.; Sansone, M.; Varriale, G.; Forti, G. Natural transmission of USP9Y gene mutations: A new perspective on the role of AZFa genes in male fertility. Hum. Mol. Genet. 2006, 15, 2673–2681. [Google Scholar] [CrossRef]
- Nuti, F.; Krausz, C. Gene polymorphisms/mutations relevant to abnormal spermatogenesis. Reprod. Biomed. Online 2008, 16, 504–513. [Google Scholar] [CrossRef]
- Tyler-Smith, C.; Krausz, C. The will-o’-the-wisp of genetics--hunting for the azoospermia factor gene. N. Engl. J. Med. 2009, 360, 925–927. [Google Scholar] [CrossRef]
- Vogt, P.H. Azoospermia factor (AZF) in Yq11: Towards a molecular understanding of its function for human male fertility and spermatogenesis. Reprod. Biomed. Online 2005, 10, 81–93. [Google Scholar] [CrossRef]
- Mohr, S.; Stryker, J.M.; Lambowitz, A.M. A DEAD-box protein functions as an ATP-dependent RNA chaperone in group I intron splicing. Cell 2002, 109, 769–779. [Google Scholar] [CrossRef]
- Ditton, H.J.; Zimmer, J.; Kamp, C.; Rajpert-De Meyts, E.; Vogt, P.H. The AZFa gene DBY (DDX3Y) is widely transcribed but the protein is limited to the male germ cells by translation control. Hum. Mol. Genet. 2004, 13, 2333–2341. [Google Scholar] [CrossRef]
- Jaroszynski, L.; Zimmer, J.; Fietz, D.; Bergmann, M.; Kliesch, S.; Vogt, P.H. Translational control of the AZFa gene DDX3Y by 5’UTR exon-T extension. Int. J. Androl. 2011, 34, 313–326. [Google Scholar] [CrossRef]
- Sekiguchi, T.; Iida, H.; Fukumura, J.; Nishimoto, T. Human DDX3Y, the Y-encoded isoform of RNA helicase DDX3, rescues a hamster temperature-sensitive ET24 mutant cell line with a DDX3X mutation. Exp. Cell Res. 2004, 300, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Rauschendorf, M.A.; Zimmer, J.; Ohnmacht, C.; Vogt, P.H. DDX3X, the X homologue of AZFa gene DDX3Y, expresses a complex pattern of transcript variants only in the male germ line. Mol. Hum. Reprod. 2014, 20, 1208–1222. [Google Scholar] [CrossRef]
- Lahn, B.T.; Page, D.C. Functional coherence of the human Y chromosome. Science 1997, 278, 675–680. [Google Scholar] [CrossRef]
- Mitchell, S.F.; Lorsch, J.R. Should I stay or should I go? Eukaryotic translation initiation factors 1 and 1A control start codon recognition. J. Biol. Chem. 2008, 283, 27345–27349. [Google Scholar] [CrossRef]
- Kleiman, S.E.; Yogev, L.; Hauser, R.; Botchan, A.; Maymon, B.B.; Paz, G.; Yavetz, H. Expression profile of AZF genes in testicular biopsies of azoospermic men. Hum. Reprod. 2007, 22, 151–158. [Google Scholar] [CrossRef][Green Version]
- Nygard, O.; Nika, H. Identification by RNA-protein cross-linking of ribosomal proteins located at the interface between the small and the large subunits of mammalian ribosomes. EMBO J. 1982, 1, 357–362. [Google Scholar] [CrossRef]
- Andres, O.; Kellermann, T.; Lopez-Giraldez, F.; Rozas, J.; Domingo-Roura, X.; Bosch, M. RPS4Y gene family evolution in primates. BMC Evol. Biol. 2008, 8, 142. [Google Scholar] [CrossRef]
- Skaletsky, H.; Kuroda-Kawaguchi, T.; Minx, P.J.; Cordum, H.S.; Hillier, L.; Brown, L.G.; Repping, S.; Pyntikova, T.; Ali, J.; Bieri, T.; et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 2003, 423, 825–837. [Google Scholar] [CrossRef]
- Lopes, A.M.; Miguel, R.N.; Sargent, C.A.; Ellis, P.J.; Amorim, A.; Affara, N.A. The human RPS4 paralogue on Yq11.223 encodes a structurally conserved ribosomal protein and is preferentially expressed during spermatogenesis. BMC Mol. Biol. 2010, 11, 33. [Google Scholar] [CrossRef]
- Bernstein, B.E.; Meissner, A.; Lander, E.S. The mammalian epigenome. Cell 2007, 128, 669–681. [Google Scholar] [CrossRef]
- Liu, X.; Secombe, J. The Histone Demethylase KDM5 Activates Gene Expression by Recognizing Chromatin Context through Its PHD Reader Motif. Cell Rep. 2015, 13, 2219–2231. [Google Scholar] [CrossRef]
- Mizukami, H.; Kim, J.D.; Tabara, S.; Lu, W.; Kwon, C.; Nakashima, M.; Fukamizu, A. KDM5D-mediated H3K4 demethylation is required for sexually dimorphic gene expression in mouse embryonic fibroblasts. J. Biochem. 2019, 165, 335–342. [Google Scholar] [CrossRef]
- Blair, L.P.; Cao, J.; Zou, M.R.; Sayegh, J.; Yan, Q. Epigenetic Regulation by Lysine Demethylase 5 (KDM5) Enzymes in Cancer. Cancers 2011, 3, 1383–1404. [Google Scholar] [CrossRef]
- Akimoto, C.; Kitagawa, H.; Matsumoto, T.; Kato, S. Spermatogenesis-specific association of SMCY and MSH5. Genes Cells Devoted Mol. Cell. Mech. 2008, 13, 623–633. [Google Scholar] [CrossRef]
- Navarro-Costa, P.; Plancha, C.E.; Goncalves, J. Genetic dissection of the AZF regions of the human Y chromosome: Thriller or filler for male (in)fertility? J. Biomed. Biotechnol. 2010, 2010, 936569. [Google Scholar] [CrossRef]
- Bhowmick, B.K.; Satta, Y.; Takahata, N. The origin and evolution of human ampliconic gene families and ampliconic structure. Genome Res. 2007, 17, 441–450. [Google Scholar] [CrossRef]
- Repping, S.; van Daalen, S.K.; Brown, L.G.; Korver, C.M.; Lange, J.; Marszalek, J.D.; Pyntikova, T.; van der Veen, F.; Skaletsky, H.; Page, D.C.; et al. High mutation rates have driven extensive structural polymorphism among human Y chromosomes. Nat. Genet. 2006, 38, 463–467. [Google Scholar] [CrossRef]
- Page, D.C. 2003 Curt Stern Award address. On low expectation exceeded; or, the genomic salvation of the Y chromosome. Am. J. Hum. Genet. 2004, 74, 399–402. [Google Scholar] [CrossRef]
- Seboun, E.; Barbaux, S.; Bourgeron, T.; Nishi, S.; Agulnik, A.; Egashira, M.; Nikkawa, N.; Bishop, C.; Fellous, M.; McElreavey, K.; et al. Gene sequence, localization, and evolutionary conservation of DAZLA, a candidate male sterility gene. Genomics 1997, 41, 227–235. [Google Scholar] [CrossRef]
- Saxena, R.; de Vries, J.W.; Repping, S.; Alagappan, R.K.; Skaletsky, H.; Brown, L.G.; Ma, P.; Chen, E.; Hoovers, J.M.; Page, D.C. Four DAZ genes in two clusters found in the AZFc region of the human Y chromosome. Genomics 2000, 67, 256–267. [Google Scholar] [CrossRef]
- Kato, Y.; Katsuki, T.; Kokubo, H.; Masuda, A.; Saga, Y. Dazl is a target RNA suppressed by mammalian NANOS2 in sexually differentiating male germ cells. Nat. Commun. 2016, 7, 11272. [Google Scholar] [CrossRef]
- Ruggiu, M.; Speed, R.; Taggart, M.; McKay, S.J.; Kilanowski, F.; Saunders, P.; Dorin, J.; Cooke, H.J. The mouse Dazla gene encodes a cytoplasmic protein essential for gametogenesis. Nature 1997, 389, 73–77. [Google Scholar] [CrossRef]
- Oud, M.S.; Volozonoka, L.; Smits, R.M.; Vissers, L.; Ramos, L.; Veltman, J.A. A systematic review and standardized clinical validity assessment of male infertility genes. Hum. Reprod. 2019, 34, 932–941. [Google Scholar] [CrossRef]
- Tuttelmann, F.; Ruckert, C.; Ropke, A. Disorders of spermatogenesis: Perspectives for novel genetic diagnostics after 20 years of unchanged routine. Med. Genet. 2018, 30, 12–20. [Google Scholar] [CrossRef]
- Callewaert, L.; Christiaens, V.; Haelens, A.; Verrijdt, G.; Verhoeven, G.; Claessens, F. Implications of a polyglutamine tract in the function of the human androgen receptor. Biochem. Biophys. Res. Commun. 2003, 306, 46–52. [Google Scholar] [CrossRef]
- Roy, A.K.; Lavrovsky, Y.; Song, C.S.; Chen, S.; Jung, M.H.; Velu, N.K.; Bi, B.Y.; Chatterjee, B. Regulation of androgen action. Vitam. Horm. 1999, 55, 309–352. [Google Scholar]
- McLachlan, R.I.; Wreford, N.G.; O’Donnell, L.; de Kretser, D.M.; Robertson, D.M. The endocrine regulation of spermatogenesis: Independent roles for testosterone and FSH. J. Endocrinol. 1996, 148, 1–9. [Google Scholar] [CrossRef]
- Gottlieb, B.; Beitel, L.K.; Nadarajah, A.; Paliouras, M.; Trifiro, M. The androgen receptor gene mutations database: 2012 update. Hum. Mutat. 2012, 33, 887–894. [Google Scholar] [CrossRef]
- Yeh, S.; Tsai, M.Y.; Xu, Q.; Mu, X.M.; Lardy, H.; Huang, K.E.; Lin, H.; Yeh, S.D.; Altuwaijri, S.; Zhou, X.; et al. Generation and characterization of androgen receptor knockout (ARKO) mice: An in vivo model for the study of androgen functions in selective tissues. Proc. Natl. Acad. Sci. USA 2002, 99, 13498–13503. [Google Scholar] [CrossRef]
- De Gendt, K.; Swinnen, J.V.; Saunders, P.T.; Schoonjans, L.; Dewerchin, M.; Devos, A.; Tan, K.; Atanassova, N.; Claessens, F.; Lécureuil, C.; et al. A Sertoli cell-selective knockout of the androgen receptor causes spermatogenic arrest in meiosis. Proc. Natl. Acad. Sci. USA 2004, 101, 1327–1332. [Google Scholar] [CrossRef]
- Edwards, A.; Hammond, H.A.; Jin, L.; Caskey, C.T.; Chakraborty, R. Genetic variation at five trimeric and tetrameric tandem repeat loci in four human population groups. Genomics 1992, 12, 241–253. [Google Scholar] [CrossRef]
- Tut, T.G.; Ghadessy, F.J.; Trifiro, M.A.; Pinsky, L.; Yong, E.L. Long polyglutamine tracts in the androgen receptor are associated with reduced trans-activation, impaired sperm production, and male infertility. J. Clin. Endocrinol. Metab. 1997, 82, 3777–3782. [Google Scholar] [CrossRef]
- Krausz, C.; Escamilla, A.R.; Chianese, C. Genetics of male infertility: From research to clinic. Reproduction 2015, 150, R159–R174. [Google Scholar] [CrossRef]
- Davis-Dao, C.A.; Tuazon, E.D.; Sokol, R.Z.; Cortessis, V.K. Male infertility and variation in CAG repeat length in the androgen receptor gene: A meta-analysis. J. Clin. Endocrinol. Metab. 2007, 92, 4319–4326. [Google Scholar] [CrossRef]
- Westerveld, H.; Visser, L.; Tanck, M.; van der Veen, F.; Repping, S. CAG repeat length variation in the androgen receptor gene is not associated with spermatogenic failure. Fertil. Steril. 2008, 89, 253–259. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schutz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The nuclear receptor superfamily: The second decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef]
- Ikeda, Y.; Lala, D.S.; Luo, X.; Kim, E.; Moisan, M.P.; Parker, K.L. Characterization of the mouse FTZ-F1 gene, which encodes a key regulator of steroid hydroxylase gene expression. Mol. Endocrinol. 1993, 7, 852–860. [Google Scholar]
- Morohashi, K.; Iida, H.; Nomura, M.; Hatano, O.; Honda, S.; Tsukiyama, T.; Niwa, O.; Hara, T.; Takakusu, A.; Shibata, Y.; et al. Functional difference between Ad4BP and ELP, and their distributions in steroidogenic tissues. Mol. Endocrinol. 1994, 8, 643–653. [Google Scholar]
- Barnhart, K.M.; Mellon, P.L. The orphan nuclear receptor, steroidogenic factor-1, regulates the glycoprotein hormone alpha-subunit gene in pituitary gonadotropes. Mol. Endocrinol. 1994, 8, 878–885. [Google Scholar] [PubMed]
- Ingraham, H.A.; Lala, D.S.; Ikeda, Y.; Luo, X.; Shen, W.H.; Nachtigal, M.W.; Abbud, R.; Nilson, J.H.; Parker, K.L. The nuclear receptor steroidogenic factor 1 acts at multiple levels of the reproductive axis. Genes Dev. 1994, 8, 2302–2312. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, K.; Lei, H.; Yoshii, H.; Nomura, M.; Nagano, M.; Shiba, H.; Sasaki, H.; Osawa, Y.; Ninomiya, Y.; Niwa, O.; et al. Developmental defects of the ventromedial hypothalamic nucleus and pituitary gonadotroph in the Ftz-F1 disrupted mice. Dev. Dyn. 1995, 204, 22–29. [Google Scholar] [CrossRef]
- Luo, X.; Ikeda, Y.; Parker, K.L. A cell-specific nuclear receptor is essential for adrenal and gonadal development and sexual differentiation. Cell 1994, 77, 481–490. [Google Scholar] [CrossRef]
- Jameson, J.L. Of mice and men: The tale of steroidogenic factor-1. J. Clin. Endocrinol. Metab. 2004, 89, 5927–5929. [Google Scholar] [CrossRef][Green Version]
- Sadovsky, Y.; Crawford, P.A.; Woodson, K.G.; Polish, J.A.; Clements, M.A.; Tourtellotte, L.M.; Simburger, K.; Milbrandt, J. Mice deficient in the orphan receptor steroidogenic factor 1 lack adrenal glands and gonads but express P450 side-chain-cleavage enzyme in the placenta and have normal embryonic serum levels of corticosteroids. Proc. Natl. Acad. Sci. USA 1995, 92, 10939–10943. [Google Scholar] [CrossRef]
- Ferraz-de-Souza, B.; Lin, L.; Achermann, J.C. Steroidogenic factor-1 (SF-1, NR5A1) and human disease. Mol. Cell. Endocrinol. 2011, 336, 198–205. [Google Scholar] [CrossRef]
- Bashamboo, A.; Ferraz-de-Souza, B.; Lourenco, D.; Lin, L.; Sebire, N.J.; Montjean, D.; Bignon-Topalovic, J.; Mandelbaum, J.; Siffroi, J.P.; Christin-Maitre, S.; et al. Human male infertility associated with mutations in NR5A1 encoding steroidogenic factor 1. Am. J. Hum. Genet. 2010, 87, 505–512. [Google Scholar] [CrossRef]
- Ropke, A.; Tewes, A.C.; Gromoll, J.; Kliesch, S.; Wieacker, P.; Tuttelmann, F. Comprehensive sequence analysis of the NR5A1 gene encoding steroidogenic factor 1 in a large group of infertile males. Eur. J. Hum. Genet. 2013, 21, 1012–1015. [Google Scholar] [CrossRef]
- Ferlin, A.; Rocca, M.S.; Vinanzi, C.; Ghezzi, M.; Di Nisio, A.; Foresta, C. Mutational screening of NR5A1 gene encoding steroidogenic factor 1 in cryptorchidism and male factor infertility and functional analysis of seven undescribed mutations. Fertil. Steril. 2015, 104, 163–169.e161. [Google Scholar] [CrossRef] [PubMed]
- Zare-Abdollahi, D.; Safari, S.; Mirfakhraie, R.; Movafagh, A.; Bastami, M.; Azimzadeh, P.; Salsabili, N.; Ebrahimizadeh, W.; Salami, S.; Omrani, M.D. Mutational screening of the NR5A1 in azoospermia. Andrologia 2015, 47, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, D.V.S.; Nizamuddin, S.; Manisha, G.; Devi, J.R.; Gupta, N.J.; Chakravarthy, B.N.; Deenadayal, M.; Singh, L.; Thangaraj, K. NR5A1 mutations are not associated with male infertility in Indian men. Andrologia 2018, 50. [Google Scholar] [CrossRef] [PubMed]
- Raymond, C.S.; Shamu, C.E.; Shen, M.M.; Seifert, K.J.; Hirsch, B.; Hodgkin, J.; Zarkower, D. Evidence for evolutionary conservation of sex-determining genes. Nature 1998, 391, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Volff, J.N.; Zarkower, D.; Bardwell, V.J.; Schartl, M. Evolutionary dynamics of the DM domain gene family in metazoans. J. Mol. Evol. 2003, 57 (Suppl. 1), S241–S249. [Google Scholar] [CrossRef]
- Zarkower, D. Establishing sexual dimorphism: Conservation amidst diversity? Nat. Rev. Genet. 2001, 2, 175–185. [Google Scholar] [CrossRef]
- Moniot, B.; Berta, P.; Scherer, G.; Sudbeck, P.; Poulat, F. Male specific expression suggests role of DMRT1 in human sex determination. Mech. Dev. 2000, 91, 323–325. [Google Scholar] [CrossRef]
- Huang, S.; Ye, L.; Chen, H. Sex determination and maintenance: The role of DMRT1 and FOXL2. Asian J. Androl. 2017, 19, 619–624. [Google Scholar]
- Raymond, C.S.; Murphy, M.W.; O’Sullivan, M.G.; Bardwell, V.J.; Zarkower, D. Dmrt1, a gene related to worm and fly sexual regulators, is required for mammalian testis differentiation. Genes Dev. 2000, 14, 2587–2595. [Google Scholar] [CrossRef]
- Lopes, A.M.; Aston, K.I.; Thompson, E.; Carvalho, F.; Goncalves, J.; Huang, N.; Matthiesen, R.; Noordam, M.J.; Quintela, I.; Ramu, A.; et al. Human spermatogenic failure purges deleterious mutation load from the autosomes and both sex chromosomes, including the gene DMRT1. PLoS Genet. 2013, 9, e1003349. [Google Scholar] [CrossRef]
- Tewes, A.C.; Ledig, S.; Tuttelmann, F.; Kliesch, S.; Wieacker, P. DMRT1 mutations are rarely associated with male infertility. Fertil. Steril. 2014, 102, 816–820.e813. [Google Scholar] [CrossRef]
- Araujo, T.F.; Friedrich, C.; Grangeiro, C.H.P.; Martelli, L.R.; Grzesiuk, J.D.; Emich, J.; Wyrwoll, M.J.; Kliesch, S.; Simões, A.L.; Tüttelmann, F.; et al. Sequence analysis of 37 candidate genes for male infertility: Challenges in variant assessment and validating genes. Andrology 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.C.; Carvalho, F.; Goncalves, J.; Fernandes, S.; Marques, P.I.; Sousa, M.; Barros, A.; Seixas, S.; Amorim, A.; Conrad, D.F.; et al. Rare double sex and mab-3-related transcription factor 1 regulatory variants in severe spermatogenic failure. Andrology 2015, 3, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.J.; McCarrey, J.R.; Yang, F.; Page, D.C. An abundance of X-linked genes expressed in spermatogonia. Nat. Genet. 2001, 27, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Adelman, C.A.; Petrini, J.H. ZIP4H (TEX11) deficiency in the mouse impairs meiotic double strand break repair and the regulation of crossing over. PLoS Genet. 2008, 4, e1000042. [Google Scholar] [CrossRef] [PubMed]
- Yatsenko, A.N.; Georgiadis, A.P.; Ropke, A.; Berman, A.J.; Jaffe, T.; Olszewska, M.; Westernströer, B.; Sanfilippo, J.; Kurpisz, M.; Rajkovic, A.; et al. X-linked TEX11 mutations, meiotic arrest, and azoospermia in infertile men. N. Engl. J. Med. 2015, 372, 2097–2107. [Google Scholar] [CrossRef]
- Yang, F.; Silber, S.; Leu, N.A.; Oates, R.D.; Marszalek, J.D.; Skaletsky, H.; Brown, L.G.; Rozen, S.; Page, D.C.; Wang, P.J. TEX11 is mutated in infertile men with azoospermia and regulates genome-wide recombination rates in mouse. EMBO Mol. Med. 2015, 7, 1198–1210. [Google Scholar] [CrossRef]
- Nakamura, S.; Miyado, M.; Saito, K.; Katsumi, M.; Nakamura, A.; Kobori, Y.; Tanaka, Y.; Ishikawa, H.; Yoshida, A.; Okada, H.; et al. Next-generation sequencing for patients with non-obstructive azoospermia: Implications for significant roles of monogenic/oligogenic mutations. Andrology 2017, 5, 824–831. [Google Scholar] [CrossRef]
- Ben Khelifa, M.; Ghieh, F.; Boudjenah, R.; Hue, C.; Fauvert, D.; Dard, R.; Garchon, H.J.; Vialard, F.; et al. A MEI1 homozygous missense mutation associated with meiotic arrest in a consanguineous family. Hum. Reprod. 2018, 33, 1034–1037. [Google Scholar] [CrossRef]
- Sha, Y.; Zheng, L.; Ji, Z.; Mei, L.; Ding, L.; Lin, S.; Wang, X.; Yang, X.; Li, P. A novel TEX11 mutation induces azoospermia: A case report of infertile brothers and literature review. BMC Med. Genet. 2018, 19, 63. [Google Scholar] [CrossRef]
- Boroujeni, P.B.; Sabbaghian, M.; Totonchi, M.; Sodeifi, N.; Sarkardeh, H.; Samadian, A.; Sadighi-Gilani, M.A.; Gourabi, H. Expression analysis of genes encoding TEX11, TEX12, TEX14 and TEX15 in testis tissues of men with non-obstructive azoospermia. JBRA Assist. Reprod. 2018, 22, 185–192. [Google Scholar] [CrossRef]
- Greenbaum, M.P.; Yan, W.; Wu, M.H.; Lin, Y.N.; Agno, J.E.; Sharma, M.; Braun, R.E.; Rajkovic, A.; Matzuk, M.M. TEX14 is essential for intercellular bridges and fertility in male mice. Proc. Natl. Acad. Sci. USA 2006, 103, 4982–4987. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Yoon, J.; Matsuura, A.; Na, J.H.; Lee, W.K.; Kim, H.; Choi, J.W.; Park, J.E.; Park, S.J.; Kim, K.T.; et al. Structural and biochemical insights into the role of testis-expressed gene 14 (TEX14) in forming the stable intercellular bridges of germ cells. Proc. Natl. Acad. Sci. USA 2015, 112, 12372–12377. [Google Scholar] [CrossRef] [PubMed]
- Fakhro, K.A.; Elbardisi, H.; Arafa, M.; Robay, A.; Rodriguez-Flores, J.L.; Al-Shakaki, A.; Syed, N.; Mezey, J.G.; Abi Khalil, C.; Malek, J.A.; et al. Point-of-care whole-exome sequencing of idiopathic male infertility. Genet. Med. 2018, 20, 1365–1373. [Google Scholar] [CrossRef]
- Yang, F.; Eckardt, S.; Leu, N.A.; McLaughlin, K.J.; Wang, P.J. Mouse TEX15 is essential for DNA double-strand break repair and chromosomal synapsis during male meiosis. J. Cell Biol. 2008, 180, 673–679. [Google Scholar] [CrossRef]
- Gershoni, M.; Hauser, R.; Yogev, L.; Lehavi, O.; Azem, F.; Yavetz, H.; Pietrokovski, S.; Kleiman, S.E. A familial study of azoospermic men identifies three novel causative mutations in three new human azoospermia genes. Genet. Med. 2017, 19, 998–1006. [Google Scholar] [CrossRef]
- Okutman, O.; Muller, J.; Baert, Y.; Serdarogullari, M.; Gultomruk, M.; Piton, A.; Rombaut, C.; Benkhalifa, M.; Teletin, M.; Skory, V.; et al. Exome sequencing reveals a nonsense mutation in TEX15 causing spermatogenic failure in a Turkish family. Hum. Mol. Genet. 2015, 24, 5581–5588. [Google Scholar] [CrossRef]
- Colombo, R.; Pontoglio, A.; Bini, M. Two Novel TEX15 Mutations in a Family with Nonobstructive Azoospermia. Gynecol. Obstet. Investig. 2017, 82, 283–286. [Google Scholar] [CrossRef]
- Zhou, Y.D.; Barnard, M.; Tian, H.; Li, X.; Ring, H.Z.; Francke, U.; Shelton, J.; Richardson, J.; Russell, D.W.; McKnight, S.L. Molecular characterization of two mammalian bHLH-PAS domain proteins selectively expressed in the central nervous system. Proc. Natl. Acad. Sci. USA 1997, 94, 713–718. [Google Scholar] [CrossRef]
- Reick, M.; Garcia, J.A.; Dudley, C.; McKnight, S.L. NPAS2: An analog of clock operative in the mammalian forebrain. Science 2001, 293, 506–509. [Google Scholar] [CrossRef]
- Ramasamy, R.; Bakircioglu, M.E.; Cengiz, C.; Karaca, E.; Scovell, J.; Jhangiani, S.N.; Akdemir, Z.C.; Bainbridge, M.; Yu, Y.; Huff, C.; et al. Whole-exome sequencing identifies novel homozygous mutation in NPAS2 in family with nonobstructive azoospermia. Fertil. Steril. 2015, 104, 286–291. [Google Scholar] [CrossRef]
- Choi, Y.; Jeon, S.; Choi, M.; Lee, M.H.; Park, M.; Lee, D.R.; Jun, K.Y.; Kwon, Y.; Lee, O.H.; Song, S.H.; et al. Mutations in SOHLH1 gene associate with nonobstructive azoospermia. Hum. Mutat. 2010, 31, 788–793. [Google Scholar] [CrossRef]
- Stouffs, K.; Lissens, W.; Tournaye, H.; Van Steirteghem, A.; Liebaers, I. Possible role of USP26 in patients with severely impaired spermatogenesis. Eur. J. Hum. Genet. 2005, 13, 336–340. [Google Scholar] [CrossRef]
- Ma, Q.; Li, Y.; Guo, H.; Li, C.; Chen, J.; Luo, M.; Jiang, Z.; Li, H.; Gui, Y. A Novel Missense Mutation in USP26 Gene Is Associated With Nonobstructive Azoospermia. Reprod. Sci 2016, 23, 1434–1441. [Google Scholar] [CrossRef]
- Miyamoto, T.; Hasuike, S.; Yogev, L.; Maduro, M.R.; Ishikawa, M.; Westphal, H.; Lamb, D.J. Azoospermia in patients heterozygous for a mutation in SYCP3. Lancet 2003, 362, 1714–1719. [Google Scholar] [CrossRef]
- Ayhan, O.; Balkan, M.; Guven, A.; Hazan, R.; Atar, M.; Tok, A.; Tolun, A. Truncating mutations in TAF4B and ZMYND15 causing recessive azoospermia. J. Med. Genet. 2014, 51, 239–244. [Google Scholar] [CrossRef]
- Maor-Sagie, E.; Cinnamon, Y.; Yaacov, B.; Shaag, A.; Goldsmidt, H.; Zenvirt, S.; Laufer, N.; Richler, C.; Frumkin, A. Deleterious mutation in SYCE1 is associated with non-obstructive azoospermia. J. Assist. Reprod. Genet. 2015, 32, 887–891. [Google Scholar] [CrossRef]
- Tenenbaum-Rakover, Y.; Weinberg-Shukron, A.; Renbaum, P.; Lobel, O.; Eideh, H.; Gulsuner, S.; Dahary, D.; Abu-Rayyan, A.; Kanaan, M.; Levy-Lahad, E.; et al. Minichromosome maintenance complex component 8 (MCM8) gene mutations result in primary gonadal failure. J. Med. Genet. 2015, 52, 391–399. [Google Scholar] [CrossRef]
- Mou, L.; Wang, Y.; Li, H.; Huang, Y.; Jiang, T.; Huang, W.; Li, Z.; Chen, J.; Xie, J.; Liu, Y.; et al. A dominant-negative mutation of HSF2 associated with idiopathic azoospermia. Hum. Genet. 2013, 132, 159–165. [Google Scholar] [CrossRef]
- Kherraf, Z.E.; Christou-Kent, M.; Karaouzene, T.; Amiri-Yekta, A.; Martinez, G.; Vargas, A.S.; Lambert, E.; Borel, C.; Dorphin, B.; Aknin-Seifer, I.; et al. SPINK2 deficiency causes infertility by inducing sperm defects in heterozygotes and azoospermia in homozygotes. EMBO Mol. Med. 2017, 9, 1132–1149. [Google Scholar] [CrossRef]
- Arafat, M.; Har-Vardi, I.; Harlev, A.; Levitas, E.; Zeadna, A.; Abofoul-Azab, M.; Dyomin, V.; Sheffield, V.C.; Lunenfeld, E.; Huleihel, M.; et al. Mutation in TDRD9 causes non-obstructive azoospermia in infertile men. J. Med. Genet. 2017, 54, 633–639. [Google Scholar] [CrossRef]
- Krausz, C. Male infertility: Pathogenesis and clinical diagnosis. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Tuttelmann, F.; Rajpert-De Meyts, E.; Nieschlag, E.; Simoni, M. Gene polymorphisms and male infertility--a meta-analysis and literature review. Reprod. Biomed. Online 2007, 15, 643–658. [Google Scholar] [CrossRef]
- Mobasseri, N.; Babaei, F.; Karimian, M.; Nikzad, H. Androgen receptor (AR)-CAG trinucleotide repeat length and idiopathic male infertility: A case-control trial and a meta-analysis. EXCLI J. 2018, 17, 1167–1179. [Google Scholar] [PubMed]
- Pan, B.; Li, R.; Chen, Y.; Tang, Q.; Wu, W.; Chen, L.; Lu, C.; Pan, F.; Ding, H.; Xia, Y.; et al. Genetic Association between Androgen Receptor Gene CAG Repeat Length Polymorphism and Male Infertility: A Meta-Analysis. Medicine 2016, 95, e2878. [Google Scholar] [CrossRef]
- Munoz, X.; Navarro, M.; Mata, A.; Bassas, L.; Larriba, S. Association of PIWIL4 genetic variants with germ cell maturation arrest in infertile Spanish men. Asian J. Androl. 2014, 16, 931–933. [Google Scholar]
- Kamaliyan, Z.; Pouriamanesh, S.; Amin-Beidokhti, M.; Rezagholizadeh, A.; Mirfakhraie, R. HIWI2 rs508485 Polymorphism is Associated with Non-obstructive Azoospermia in Iranian Patients. Rep. Biochem. Mol. Biol. 2017, 5, 108–111. [Google Scholar]
- Kamaliyan, Z.; Pouriamanesh, S.; Soosanabadi, M.; Gholami, M.; Mirfakhraie, R. Investigation of piwi-interacting RNA pathway genes role in idiopathic non-obstructive azoospermia. Sci. Rep. 2018, 8, 142. [Google Scholar] [CrossRef]
- Stuppia, L.; Gatta, V.; Scarciolla, O.; Colosimo, A.; Guanciali-Franchi, P.; Calabrese, G.; Palka, G. The methylenetethrahydrofolate reductase (MTHFR) C677T polymorphism and male infertility in Italy. J. Endocrinol. Investig. 2003, 26, 620–622. [Google Scholar] [CrossRef]
- Gava, M.M.; Kayaki, E.A.; Bianco, B.; Teles, J.S.; Christofolini, D.M.; Pompeo, A.C.; Glina, S.; Barbosa, C.P. Polymorphisms in folate-related enzyme genes in idiopathic infertile Brazilian men. Reprod. Sci. 2011, 18, 1267–1272. [Google Scholar] [CrossRef]
- Kim, S.Y.; Lim, J.W.; Kim, J.W.; Park, S.Y.; Seo, J.T. Association between genetic polymorphisms in folate-related enzyme genes and infertile men with non-obstructive azoospermia. Syst. Biol. Reprod. Med. 2015, 61, 286–292. [Google Scholar]
- Lee, H.C.; Jeong, Y.M.; Lee, S.H.; Cha, K.Y.; Song, S.H.; Kim, N.K.; Lee, K.W.; Lee, S. Association study of four polymorphisms in three folate-related enzyme genes with non-obstructive male infertility. Hum. Reprod. 2006, 21, 3162–3170. [Google Scholar] [CrossRef] [PubMed]
- Ying, H.Q.; Pu, X.Y.; Liu, S.R.; A, Z.-C. Genetic variants of eNOS gene may modify the susceptibility to idiopathic male infertility. Biomarkers 2013, 18, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Pan, F.; Tang, Q.; Wu, W.; Chen, M.; Lu, C.; Ding, H.; Hu, L.; Chen, D.; Xia, Y.; et al. eNOS gene T786C, G894T and 4a4b polymorphisms and male infertility susceptibility: A meta-analysis. Andrologia 2017, 49. [Google Scholar] [CrossRef]
- Ying, H.Q.; Scott, M.B.; Zhou-Cun, A. Relationship of SNP of H2BFWT gene to male infertility in a Chinese population with idiopathic spermatogenesis impairment. Biomarkers 2012, 17, 402–406. [Google Scholar] [CrossRef]
- Rafatmanesh, A.; Nikzad, H.; Ebrahimi, A.; Karimian, M.; Zamani, T. Association of the c.-9C>T and c.368A>G transitions in H2BFWT gene with male infertility in an Iranian population. Andrologia 2018, 50. [Google Scholar] [CrossRef]
- Haji Ebrahim Zargar, H.; Mohseni Meybodi, A.; Sabbaghian, M.; Shahhoseini, M.; Asadpor, U.; Sadighi Gilani, M.A.; Chehrazi, M.; Farhangniya, M.; Shahzadeh Fazeli, S.A. Association of Two Polymorphisms in H2B.W Gene with Azoospermia and Severe Oligozoospermia in An Iranian Population. Int. J. Fertil. Steril. 2015, 9, 205–214. [Google Scholar]
- Yoshida, K.I.; Yano, M.; Chiba, K.; Honda, M.; Kitahara, S. CAG repeat length in the androgen receptor gene is enhanced in patients with idiopathic azoospermia. Urology 1999, 54, 1078–1081. [Google Scholar] [CrossRef]
- Sato, H.; Miyamoto, T.; Yogev, L.; Namiki, M.; Koh, E.; Hayashi, H.; Sasaki, Y.; Ishikawa, M.; Lamb, D.J.; Matsumoto, N.; et al. Polymorphic alleles of the human MEI1 gene are associated with human azoospermia by meiotic arrest. J. Hum. Genet. 2006, 51, 533–540. [Google Scholar] [CrossRef][Green Version]
- Wilson, G.R.; Sim, M.L.; Brody, K.M.; Taylor, J.M.; McLachlan, R.I.; O’Bryan, M.K.; Delatycki, M.B.; Lockhart, P.J. Molecular analysis of the PArkin co-regulated gene and association with male infertility. Fertil. Steril. 2010, 93, 2262–2268. [Google Scholar] [CrossRef]
- Ma, J.; Lu, H.Y.; Xia, Y.K.; Dong, H.B.; Gu, A.H.; Li, Z.Y.; Li, Z.; Chen, A.M.; Wang, X.R.; Wang, S.L. BCL2 Ala43Thr is a functional variant associated with protection against azoospermia in a Han-Chinese population. Biol. Reprod. 2010, 83, 656–662. [Google Scholar] [CrossRef][Green Version]
- Aston, K.I.; Krausz, C.; Laface, I.; Ruiz-Castane, E.; Carrell, D.T. Evaluation of 172 candidate polymorphisms for association with oligozoospermia or azoospermia in a large cohort of men of European descent. Hum. Reprod. 2010, 25, 1383–1397. [Google Scholar] [CrossRef]
- Hu, Z.; Xia, Y.; Guo, X.; Dai, J.; Li, H.; Hu, H.; Jiang, Y.; Lu, F.; Wu, Y.; Yang, X.; et al. A genome-wide association study in Chinese men identifies three risk loci for non-obstructive azoospermia. Nat. Genet. 2011, 44, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Xu, J.; Zhang, H.; Sun, J.; Sun, Y.; Wang, Z.; Liu, J.; Ding, Q.; Lu, S.; Shi, R.; et al. A genome-wide association study reveals that variants within the HLA region are associated with risk for nonobstructive azoospermia. Am. J. Hum. Genet. 2012, 90, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, J.; Zhang, T.; Li, H.; Ni, Y. rs189037, a functional variant in ATM gene promoter, is associated with idiopathic nonobstructive azoospermia. Fertil. Steril. 2013, 100, 1536–1541.e1531. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Li, Z.; Yu, J.; Tong, C.; Lin, Y.; Guo, X.; Lu, F.; Dong, J.; Xia, Y.; Wen, Y.; et al. Association analysis identifies new risk loci for non-obstructive azoospermia in Chinese men. Nat. Commun. 2014, 5, 3857. [Google Scholar] [CrossRef]
- Qin, Y.; Ji, J.; Du, G.; Wu, W.; Dai, J.; Hu, Z.; Sha, J.; Hang, B.; Lu, C.; Xia, Y.; et al. Comprehensive pathway-based analysis identifies associations of BCL2, GNAO1 and CHD2 with non-obstructive azoospermia risk. Hum. Reprod. 2014, 29, 860–866. [Google Scholar] [CrossRef][Green Version]
- Ni, B.; Ma, H.; Lin, Y.; Dai, J.; Guo, X.; Xia, Y.; Sha, J.; Hu, Z. Genetic variants in Ser-Arg protein-coding genes are associated with the risk of nonobstructive azoospermia in Chinese men. Fertil. Steril. 2014, 101, 1711-7 e1–1711-7 e2. [Google Scholar] [CrossRef]
- Ghalkhani, E.; Sheidai, M.; Gourabi, H.; Noormohammadi, Z.; Bakhtari, N.; Malekasgar, A.M. Study of single nucleotide polymorphism (rs28368082) in SPO11 gene and its association with male infertility. J. Assist. Reprod. Genet. 2014, 31, 1205–1210. [Google Scholar] [CrossRef]
- Xiong, D.K.; Chen, H.H.; Ding, X.P.; Zhang, S.H.; Zhang, J.H. Association of polymorphisms in glutathione S-transferase genes (GSTM1, GSTT1, GSTP1) with idiopathic azoospermia or oligospermia in Sichuan, China. Asian J. Androl. 2015, 17, 481–486. [Google Scholar] [CrossRef]
- Zhang, Y.; He, X.J.; Song, B.; Ye, L.; Xie, X.S.; Ruan, J.; Zhou, F.S.; Zuo, X.B.; Cao, Y.X.; Du, W.D. Association of single nucleotide polymorphisms in the USF1, GTF2A1L and OR2W3 genes with non-obstructive azoospermia in the Chinese population. J. Assist. Reprod. Genet. 2015, 32, 95–101. [Google Scholar] [CrossRef]
- Sato, Y.; Tajima, A.; Tsunematsu, K.; Nozawa, S.; Yoshiike, M.; Koh, E.; Kanaya, J.; Namiki, M.; Matsumiya, K.; Tsujimura, A.; et al. An association study of four candidate loci for human male fertility traits with male infertility. Hum. Reprod. 2015, 30, 1510–1514. [Google Scholar] [CrossRef] [PubMed]
- Gharesi-Fard, B.; Ghasemi, Z.; Shakeri, S.; Behdin, S.; Aghaei, F.; Malek-Hosseini, Z. The frequency of follicle stimulating hormone receptor gene polymorphisms in Iranian infertile men with azoospermia. Iran. J. Reprod. Med. 2015, 13, 673–678. [Google Scholar] [PubMed]
- Zhang, X.; Ding, M.; Ding, X.; Li, T.; Chen, H. Six polymorphisms in genes involved in DNA double-strand break repair and chromosome synapsis: Association with male infertility. Syst. Biol. Reprod. Med. 2015, 61, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lin, W.Q.; Cao, H.F.; Li, C.Y.; Li, F. Association of a miR-34b binding site single nucleotide polymorphism in the 3’-untranslated region of the methylenetetrahydrofolate reductase gene with susceptibility to male infertility. Genet. Mol. Res. 2015, 14, 12196–12204. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Zhang, Y.; He, X.J.; Du, W.D.; Ruan, J.; Zhou, F.S.; Wu, H.; Zha, X.; Xie, X.S.; Ye, L.; et al. Association of genetic variants in SOHLH1 and SOHLH2 with non-obstructive azoospermia risk in the Chinese population. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 184, 48–52. [Google Scholar] [CrossRef]
- Lu, J.; Gu, H.; Tang, Q.; Wu, W.; Yuan, B.; Guo, D.; Wei, Y.; Sun, H.; Xia, Y.; Ding, H.; et al. Common SNP in hsa-miR-196a-2 increases hsa-miR-196a-5p expression and predisposes to idiopathic male infertility in Chinese Han population. Sci. Rep. 2016, 6, 19825. [Google Scholar] [CrossRef]
- Chihara, M.; Yoshihara, K.; Ishiguro, T.; Adachi, S.; Okada, H.; Kashima, K.; Sato, T.; Tanaka, A.; Tanaka, K.; Enomoto, T. Association of NR3C1/Glucocorticoid Receptor gene SNP with azoospermia in Japanese men. J. Obstet. Gynaecol. Res. 2016, 42, 59–66. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, Z.; Zhang, M.; Gong, R.; Xu, Y.; Wang, B. Association of the methylenetetrahydrofolate reductase gene C677T polymorphism with the risk of male infertility: A meta-analysis. Ren. Fail. 2016, 38, 185–193. [Google Scholar] [CrossRef]
- Najafipour, R.; Rashvand, Z.; Alizadeh, A.; Aleyasin, A.; Moghbelinejad, S. Association of G/T(rs222859) polymorphism in Exon 1 of YBX2 gene with azoospermia, among Iranian infertile males. Andrologia 2016, 48, 956–960. [Google Scholar] [CrossRef]
- Ramgir, S.S.; Sekar, N.; Jindam, D.; Abilash, V.G. Association of CYP1A1*2A Polymorphism with Idiopathic Non-Obstructive Azoospermia in A South Indian Cohort. Int. J. Fertil. Steril. 2017, 11, 142–147. [Google Scholar]
- Asgari, R.; Mansouri, K.; Bakhtiari, M.; Bidmeshkipour, A.; Yari, K.; Shaveisi-Zadeh, F.; Vaisi-Raygani, A. Association of FAS-670A/G and FASL-844C/T polymorphisms with idiopathic azoospermia in Western Iran. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 218, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Jahantigh, D.; Hosseinzadeh Colagar, A. XRCC5 VNTR, XRCC6 -61C>G, and XRCC7 6721G>T Gene Polymorphisms Associated with Male Infertility Risk: Evidences from Case-Control and In Silico Studies. Int. J. Endocrinol. 2017, 2017, 4795076. [Google Scholar] [CrossRef] [PubMed]
- Ashrafzadeh, H.R.; Nazari, T.; Dehghan Tezerjani, M.; Khademi Bami, M.; Ghasemi-Esmailabad, S.; Ghasemi, N. Frequency of TNFR1 36 A/G gene polymorphism in azoospermic infertile men: A case-control study. Int. J. Reprod. Biomed. 2017, 15, 521–526. [Google Scholar] [CrossRef]
- Moghbelinejad, S.; Najafipour, R.; Momeni, A. Association of rs1057035polymorphism in microRNA biogenesis pathway gene (DICER1) with azoospermia among Iranian population. Genes Genom. 2018, 40, 17–24. [Google Scholar] [CrossRef]
- Sato, Y.; Hasegawa, C.; Tajima, A.; Nozawa, S.; Yoshiike, M.; Koh, E.; Kanaya, J.; Namiki, M.; Matsumiya, K.; Tsujimura, A.; et al. Association of TUSC1 and DPF3 gene polymorphisms with male infertility. J. Assist. Reprod. Genet. 2018, 35, 257–263. [Google Scholar] [CrossRef]
- Teimouri, M.; Najaran, H.; Hosseinzadeh, A.; Mazoochi, T. Association between two common transitions of H2BFWT gene and male infertility: A case-control, meta, and structural analysis. Andrology 2018, 6, 306–316. [Google Scholar] [CrossRef]
- Zamani-Badi, T.; Karimian, M.; Azami Tameh, A.; Nikzad, H. IL-1a C376A Transversion Variant and Risk of Idiopathic Male Infertility in Iranian Men: A Genetic Association Study. Int. J. Fertil. Steril. 2018, 12, 229–234. [Google Scholar]
- Zamani-Badi, T.; Nikzad, H.; Karimian, M. IL-1RA VNTR and IL-1alpha 4845G>T polymorphisms and risk of idiopathic male infertility in Iranian men: A case-control study and an in silico analysis. Andrologia 2018, 50, e13081. [Google Scholar] [CrossRef]
- Yu, C.H.; Xie, T.; Zhang, R.P.; A, Z.-C. Association of the common SNPs in RNF212, STAG3 and RFX2 gene with male infertility with azoospermia in Chinese population. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 221, 109–112. [Google Scholar] [CrossRef]
- Singh, V.; Bansal, S.K.; Sudhakar, D.V.S.; Neelabh Chakraborty, A.; Trivedi, S.; et al. SNPs in ERCC1, ERCC2, and XRCC1 genes of the DNA repair pathway and risk of male infertility in the Asian populations: Association study, meta-analysis, and trial sequential analysis. J. Assist. Reprod. Genet. 2019, 36, 79–90. [Google Scholar] [CrossRef]
- Busch, A.S.; Tuttelmann, F.; Cremers, J.F.; Schubert, M.; Nordhoff, V.; Schuring, A.N.; Zitzmann, M.; Gromoll, J.; Kliesch, S. FSHB -211 G>T Polymorphism as Predictor for TESE Success in Patients with Unexplained Azoospermia. J. Clin. Endocrinol. Metab. 2019, 104, 2315–2324. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Mu, C.; Ma, J.; Dai, X.; Jiao, H. The association of four SNPs in DNA mismatch repair genes with idiopathic male infertility in northwest China. Int. J. Immunogenet. 2019, 46, 451–458. [Google Scholar] [CrossRef]
- Huang, M.; Zhu, M.; Jiang, T.; Wang, Y.; Wang, C.; Jin, G.; Guo, X.; Sha, J.; Dai, J.; Wang, X.; et al. Fine mapping the MHC region identified rs4997052 as a new variant associated with nonobstructive azoospermia in Han Chinese males. Fertil. Steril. 2019, 111, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.; Kang, K.M.; Sung, S.R.; Park, J.E.; Shin, Y.J.; Song, S.H.; Seo, J.T.; Yoon, T.K.; Shim, S.H. The association of stromal antigen 3 (STAG3) sequence variations with spermatogenic impairment in the male Korean population. Asian J. Androl. 2020, 22, 106–111. [Google Scholar] [PubMed]
- Pennisi, E. Breakthrough of the year. Human genetic variation. Science 2007, 318, 1842–1843. [Google Scholar] [CrossRef]
- Hofker, M.H.; Fu, J.; Wijmenga, C. The genome revolution and its role in understanding complex diseases. Biochim. Biophys. Acta 2014, 1842, 1889–1895. [Google Scholar] [CrossRef]
- Aston, K.I.; Carrell, D.T. Genome-wide study of single-nucleotide polymorphisms associated with azoospermia and severe oligozoospermia. J. Androl. 2009, 30, 711–725. [Google Scholar] [CrossRef]
- Johnson, R.C.; Nelson, G.W.; Troyer, J.L.; Lautenberger, J.A.; Kessing, B.D.; Winkler, C.A.; et al. Accounting for multiple comparisons in a genome-wide association study (GWAS). BMC Genom. 2010, 11, 724. [Google Scholar] [CrossRef]
- Pe’er, I.; Yelensky, R.; Altshuler, D.; Daly, M.J. Estimation of the multiple testing burden for genomewide association studies of nearly all common variants. Genet. Epidemiol. 2008, 32, 381–385. [Google Scholar] [CrossRef]
- Fadista, J.; Manning, A.K.; Florez, J.C.; Groop, L. The (in)famous GWAS P-value threshold revisited and updated for low-frequency variants. Eur. J. Hum. Genet. 2016, 24, 1202–1205. [Google Scholar] [CrossRef]
- Xu, M.; Qin, Y.; Qu, J.; Lu, C.; Wang, Y.; Wu, W.; Song, L.; Wang, S.; Chen, F.; Shen, H.; et al. Evaluation of five candidate genes from GWAS for association with oligozoospermia in a Han Chinese population. PLoS ONE 2013, 8, e80374. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Jinam, T.; Iwamoto, T.; Yamauchi, A.; Imoto, I.; Inoue, I.; Tajima, A. Replication study and meta-analysis of human nonobstructive azoospermia in Japanese populations. Biol. Reprod. 2013, 88, 87. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Li, H.; Chen, X.; Zhang, X.; Mei, F.; Jia, M.; Xiong, C. PEX10, SIRPA-SIRPG, and SOX5 gene polymorphisms are strongly associated with nonobstructive azoospermia susceptibility. J. Assist. Reprod. Genet. 2019, 36, 759–768. [Google Scholar] [CrossRef]
- Zhang, Y.; Qian, J.; Wu, M.; Liu, M.; Zhang, K.; Lin, Y.; Guo, X.; Zhou, Z.; Hu, Z.; Sha, J. A susceptibility locus rs7099208 is associated with non-obstructive azoospermia via reduction in the expression of FAM160B1. J. Biomed. Res. 2015, 29, 491–500. [Google Scholar]
- Zou, S.; Song, P.; Meng, H.; Chen, T.; Chen, J.; Wen, Z.; Li, Z.; Shi, Y.; Hu, H. Association and meta-analysis of HLA and non-obstructive azoospermia in the Han Chinese population. Andrologia 2017, 49. [Google Scholar] [CrossRef]
- Perez, C.V.; Theas, M.S.; Jacobo, P.V.; Jarazo-Dietrich, S.; Guazzone, V.A.; Lustig, L. Dual role of immune cells in the testis: Protective or pathogenic for germ cells? Spermatogenesis 2013, 3, e23870. [Google Scholar] [CrossRef]
- Jinam, T.A.; Nakaoka, H.; Hosomichi, K.; Mitsunaga, S.; Okada, H.; Tanaka, A.; Tanaka, K.; Inoue, I. HLA-DPB1*04:01 allele is associated with non-obstructive azoospermia in Japanese patients. Hum. Genet. 2013, 132, 1405–1411. [Google Scholar] [CrossRef]
- Kosova, G.; Scott, N.M.; Niederberger, C.; Prins, G.S.; Ober, C. Genome-wide association study identifies candidate genes for male fertility traits in humans. Am. J. Hum. Genet. 2012, 90, 950–961. [Google Scholar] [CrossRef]
- Ni, B.; Lin, Y.; Sun, L.; Zhu, M.; Li, Z.; Wang, H.; Yu, J.; Guo, X.; Zuo, X.; Dong, J.; et al. Low-frequency germline variants across 6p22.2-6p21.33 are associated with non-obstructive azoospermia in Han Chinese men. Hum. Mol. Genet. 2015, 24, 5628–5636. [Google Scholar]
- Tuttelmann, F.; Simoni, M.; Kliesch, S.; Ledig, S.; Dworniczak, B.; Wieacker, P.; Ropke, A. Copy number variants in patients with severe oligozoospermia and Sertoli-cell-only syndrome. PLoS ONE 2011, 6, e19426. [Google Scholar] [CrossRef]
- Krausz, C.; Giachini, C.; Lo Giacco, D.; Daguin, F.; Chianese, C.; Ars, E.; Ruiz-Castane, E.; Forti, G.; Rossi, E. High resolution X chromosome-specific array-CGH detects new CNVs in infertile males. PLoS ONE 2012, 7, e44887. [Google Scholar] [CrossRef]
- Stouffs, K.; Vandermaelen, D.; Massart, A.; Menten, B.; Vergult, S.; Tournaye, H.; Lissens, W. Array comparative genomic hybridization in male infertility. Hum. Reprod. 2012, 27, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Lo Giacco, D.; Chianese, C.; Ars, E.; Ruiz-Castane, E.; Forti, G.; Krausz, C. Recurrent X chromosome-linked deletions: Discovery of new genetic factors in male infertility. J. Med. Genet. 2014, 51, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Qin, Y.; Li, Z.; Dong, J.; Dai, J.; Lu, C.; Guo, X.; Zhao, Y.; Zhu, Y.; Zhang, W.; et al. Genome-wide microRNA expression profiling in idiopathic non-obstructive azoospermia: Significant up-regulation of miR-141, miR-429 and miR-7-1-3p. Hum. Reprod. 2013, 28, 1827–1836. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Wang, Y.; Zhu, M.; Huang, M.; Jin, G.; Guo, X.; Sha, J.; Dai, J.; Hu, Z. Transcriptome-wide association study revealed two novel genes associated with nonobstructive azoospermia in a Chinese population. Fertil. Steril. 2017, 108, 1056–1062.e1054. [Google Scholar] [CrossRef] [PubMed]
- Ferfouri, F.; Boitrelle, F.; Ghout, I.; Albert, M.; Molina Gomes, D.; Wainer, R.; Bailly, M.; Selva, J.; Vialard, F. A genome-wide DNA methylation study in azoospermia. Andrology 2013, 1, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Jiang, X.; Li, Z.M.; Zhuang, X.; Zhang, X.; Ouyang, W.M.; Liu, W.B.; Mao, C.Y.; Chen, Q.; Huang, C.S.; et al. Epigenetic Inactivation of SOX30 Is Associated with Male Infertility and Offers a Therapy Target for Non-obstructive Azoospermia. Mol. Ther. Nucleic Acids 2019, 19, 72–83. [Google Scholar] [CrossRef]
- Cannarella, R.; Condorelli, R.A.; Duca, Y.; La Vignera, S.; Calogero, A.E. New insights into the genetics of spermatogenic failure: A review of the literature. Hum. Genet. 2019, 138, 125–140. [Google Scholar] [CrossRef]
- Chalmel, F.; Lardenois, A.; Evrard, B.; Mathieu, R.; Feig, C.; Demougin, P.; Gattiker, A.; Schulze, W.; Jegou, B.; Kirchhoff, C.; et al. Global human tissue profiling and protein network analysis reveals distinct levels of transcriptional germline-specificity and identifies target genes for male infertility. Hum. Reprod. 2012, 27, 3233–3248. [Google Scholar] [CrossRef]
- Yan, W. Male infertility caused by spermiogenic defects: Lessons from gene knockouts. Mol. Cell. Endocrinol. 2009, 306, 24–32. [Google Scholar] [CrossRef]
- Guo, J.; Grow, E.J.; Mlcochova, H.; Maher, G.J.; Lindskog, C.; Nie, X.; Guo, Y.; Takei, Y.; Yun, J.; Cai, L.; et al. The adult human testis transcriptional cell atlas. Cell Res. 2018, 28, 1141–1157. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Metzler-Guillemain, C.; Toure, A.; Coutton, C.; Arnoult, C.; Ray, P.F. Single gene defects leading to sperm quantitative anomalies. Clin. Genet. 2017, 91, 208–216. [Google Scholar] [CrossRef]
- Ghieh, F.; Mitchell, V.; Mandon-Pepin, B.; Vialard, F. Genetic defects in human azoospermia. Basic Clin. Androl. 2019, 29, 4. [Google Scholar] [CrossRef] [PubMed]
- Asero, P.; Calogero, A.E.; Condorelli, R.A.; Mongioi, L.; Vicari, E.; Lanzafame, F.; et al. Relevance of genetic investigation in male infertility. J. Endocrinol. Investig. 2014, 37, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Punab, M.; Poolamets, O.; Paju, P.; Vihljajev, V.; Pomm, K.; Ladva, R.; et al. Causes of male infertility: A 9-year prospective monocentre study on 1737 patients with reduced total sperm counts. Hum. Reprod. 2017, 32, 18–31. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Year | Risk Loci | Variant ID | Variant Type | Position (GRCh38) | Functional Annotation | Population | Cohort Size (Case/Control) | p-Value | OR (CI 95%) | Associated Trait | Strategy | Reference | Replication |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1999 | AR | (CAG)n | CNV | X: 67545318 | Exonic | Japanese | 41/48 | 0.0013 | NA | NOA | Candidate gene | Yoshida et al. [157] | YES |
| 2003 | MTHFR | rs1801133 | SNV | 1:11796321 | Missense | Italian | 21/105 | NA | NA | NOA | Candidate gene | Stuppia et al. [148] | YES |
| 2006 | MEI1 | rs2050033 | SNV | 22:41763225 | Synonymous | European/Israeli | 26/121 | 0.027 *,*,*,* | NA | MA | Candidate gene | Sato et al. [158] | NO |
| 2006 | MTR | rs1805087 | SNV | 1:236885200 | Missense | South Korean | 174/325 | 0.0063 *,* | 4.63 (1.40–15.31) | NOA | Candidate gene | Lee et al. [151] | YES |
| 2010 | PACRG | rs9347683 | SNV | 6:162728023 | 5′-UTR | Australian | 206/156 | 0.009 | 1.60 (1.13–2.36) | NOA | Candidate gene | Wilson et al. [159] | NO |
| 2010 | BCL2 | rs1800477 | SNV | 18:63318540 | Missense | Han Chinese | 198/183 | 0.01 * | 0.45 (0.23–0.89) | NOA | Candidate gene | Ma et al. [160] | NO |
| 2010 | KDM3A | rs34605051 | SNV | 2:86466703 | Missense | European | 80/158 | 3.23 × 10−3 *,* | NA | NOA | GWAS follow-up | Aston et al. [161] | NO |
| 2010 | TAS2R38 | rs10246939 | SNV | 7:141972804 | Missense | European | 80/158 | 7.24 × 10−4 * | NA | NOA | GWAS follow-up | Aston et al. [161] | NO |
| 2011 | MTHFR | rs1801131 | SNV | 1:11794419 | Missense | Brazilian | 55/173 | 0.01 * | 0.34 (0.16–0.74) | NOA | Candidate gene | Gava et al. [149] | YES |
| 2011 | PEX10 | rs2477686 | SNV | 1:2461209 | Intergenic | Han Chinese | 2927/5734 | 5.65 × 10−12 | 1.39 (1.26–1.52) | NOA | GWAS | Hu et al. [162] | NO |
| 2011 | PRMT6 | rs12097821 | SNV | 1:106793679 | Intergenic | Han Chinese | 2927/5734 | 5.67 × 10−10 | 1.25 (1.17–1.34) | NOA | GWAS | Hu et al. [162] | YES |
| 2011 | SOX5 | rs10842262 | SNV | 12:24031610 | Intronic | Han Chinese | 2927/5734 | 2.32 × 10−9 | 1.23 (1.15–1.32) | NOA | GWAS | Hu et al. [162] | YES |
| 2012 | H2BFWT | rs7885967 | SNV | X:104013669 | 5′-UTR | Chinese | 204/209 | 0.001 | 1.89 (1.28–2.79) | NOA | Candidate gene | Ying et al. [154] | YES |
| 2012 | HLA-DRA | rs3129878 | SNV | 6:32440958 | Intronic | Han Chinese | 2226/4576 | 3.70 × 10−16 | 1.37 (NA) | NOA | GWAS | Zhao et al. [163] | YES |
| 2012 | TSBP1 | rs498422 | SNV | 6:32318984 | Intronic | Han Chinese | 2226/4576 | 2.43 × 10−12 | 1.42 (NA) | NOA | GWAS | Zhao et al. [163] | YES |
| 2013 | ATM | rs189037 | SNV | 11:108223106 | 5′-UTR | Chinese | 229/236 | 0.003 | 1.41 (1.11–1.78) | NOA | Candidate gene | Li et al. [164] | NO |
| 2013 | NOS3 | rs2070744 | SNV | 7:150992991 | Intronic | Chinese | 151/246 | <0.001 | 2.52 (1.56–4.06) | NOA | Candidate gene | Ying et al. [152] | YES |
| 2013 | NOS3 | rs61722009 | INDEL | 7:150997170 | Intronic | Chinese | 151/246 | 0.001 | 2.27 (1.39–3.72) | NOA | Candidate gene | Ying et al. [152] | YES |
| 2014 | ABLIM1 | rs7099208 | SNV | 10:114894815 | Intronic | Han Chinese | 3608/5909 | 6.41 × 10−14 | 1.41(1.29–1.54) | NOA | GWAS replication | Hu et al. [165] | YES |
| 2014 | BCL2 | rs7226979 | SNV | 18:63257737 | Intronic | Han Chinese | 1653/2329 | 4.50 × 10−5 | 1.21 (1.11–1.33) | NOA | GWAS replication | Qin et al. [166] | NO |
| 2014 | CHD2 | rs140671 | SNV | 15:26976951 | Intronic | Han Chinese | 1653/2329 | 1.70 × 10−4 | 0.78 (0.68–0.89) | NOA | GWAS replication | Qin et al. [166] | NO |
| 2014 | GNAO1 | rs2126986 | SNV | 16:56317795 | Intronic | Han Chinese | 1653/2329 | 2.30 × 10−6 | 1.28 (1.15–1.41) | NOA | GWAS replication | Qin et al. [166] | NO |
| 2014 | HLA-DRA | rs7194 | SNV | 6:32444703 | 3′-UTR | Han Chinese | 3608/5909 | 3.76 × 10−19 | 1.30 (1.23–1.38) | NOA | GWAS replication | Hu et al. [165] | YES |
| 2014 | IL17A | rs13206743 | SNV | 6:52152310 | Intergenic | Han Chinese | 3608/5909 | 3.69 × 10−8 | 1.35 (1.22–1.51) | NOA | GWAS replication | Hu et al. [165] | NO |
| 2014 | PIWIL4 | rs508485 | SNV | 11:94621313 | 3′-UTR | Spanish | 22/56 | 0.021 | NA | MA | Candidate gene | Muñoz et al. [145] | YES |
| 2014 | SFRS4 | rs12046213 | SNV | 1:29186811 | Intergenic | Chinese | 962/1931 | 0.021 | 0.88 (0.78–0.98) | NOA | Candidate gene | Ni et al. [167] | NO |
| 2014 | SFRS6 | rs6103330 | SNV | 20:43458814 | Intronic | Chinese | 962/1931 | 2.78 × 10−3 | 1.28 (1.09–1.50) | NOA | Candidate gene | Ni et al. [167] | NO |
| 2014 | SFRS9 | rs17431717 | SNV | 12:120474407 | Intronic | Chinese | 962/1931 | 0.035 | 0.73 (0.54–0.98) | NOA | Candidate gene | Ni et al. [167] | NO |
| 2014 | SFRS9 | rs10849753 | SNV | 12:120473010 | Intronic | Chinese | 962/1931 | 4.32 × 10−3 | 1.17 (1.05–1.31) | NOA | Candidate gene | Ni et al. [167] | NO |
| 2014 | SPO11 | rs28368082 | SNV | 20:57335452 | Missense | Iranian | 58/50 | 0.006 | 6.68 (NA) | NOA | Candidate gene | Ghalkhani et al. [168] | NO |
| 2015 | GSTP1 | rs1695 | SNV | 11:67585218 | Missense | Chinese | 361/234 | 0.023 * | 1.48 (1.06–2.07) | NOA | Candidate gene | Xiong et al. [169] | NO |
| 2015 | USF1 | rs2516838 | SNV | 1:161044580 | Intronic | Chinese | 361/368 | 0.02 | 1.43 (1.06–1.95) | NOA | Candidate gene | Zhang et al. [170] | NO |
| 2015 | EPSTI1 | rs12870438 | SNV | 13:42906069 | Intronic | Japanese | 76/791 | 0.0059 | 1.92 (1.21–3.05) | NOA | Candidate gene | Sato et al. [171] | NO |
| 2015 | FSHR | rs6165 | SNV | 2:48963902 | Missense | Iranian | 126/86 | 0.001 | 2.06 (1.36–3.12) | NOA | Candidate gene | Gharesi-Fard et al. [172] | NO |
| 2015 | MLH3 | rs175080 | SNV | 14:75047125 | Missense | Chinese | 244/614 | <0.001 | 1.75 (1.27–2.41) | NOA | Candidate gene | Zhang et al. [173] | NO |
| 2015 | MTHFR | rs55763075 | SNV | 1:11790377 | 3′-UTR | Chinese | 253/458 | 0.043 | 1.27 (1.01–1.58) | NOA | Candidate gene | Zhang et al. [174] | NO |
| 2015 | SOHLH2 | rs1328626 | SNV | 13:36204635 | Intronic | Chinese | 361/368 | 0.038 | 0.80 (0.65–0.99) | NOA | Candidate gene | Song et al. [175] | NO |
| 2015 | SOHLH2 | rs6563386 | SNV | 13:36202894 | Intronic | Chinese | 361/368 | 0.029 | 1.40 (1.03–1.90) | NOA | Candidate gene | Song et al. [175] | NO |
| 2016 | HSA-miR-196a | rs11614913 | SNV | 12:53991815 | Intergenic | Chinese | 140/486 | 0.009 * | 1.76 (1.15–2.70) | NOA | Candidate gene | Lu et al. [176] | NO |
| 2016 | NR3C1 | rs852977 | SNV | 5:143307929 | Intronic | Japanese | 335/410 | 5.70 × 10−15 | 3.20 (2.40–4.26) | NOA | Candidate gene | Chihara et al. [177] | NO |
| 2016 | TDRD1 | rs77559927 | SNV | 10:114179297 | 5′-UTR | Chinese | 342/493 | 0.03 * | 0.73 (0.56–0.97) | NOA | Candidate gene | Zhu et al. [178] | NO |
| 2016 | YBX2 | rs222859 | SNV | 17:7294475 | Missense | Iranian | 60/96 | <0.05 *,*,* | 0.23 (0.12–0.6) | NOA | Candidate gene | Najafipour et al. [179] | NO |
| 2017 | CYP1A1 | rs4646903 | SNV | 15:74719300 | Intergenic | South Indian | 120/80 | 0.0001 * | 3.71 (2.05–6.74) | NOA | Candidate gene | Ramgir et al. [180] | NO |
| 2017 | FASL | rs763110 | SNV | 1:172658358 | Intergenic | Iranian | 102/110 | 0.02 *,*,* | NA | NOA | Candidate gene | Asgari et al. [181] | NO |
| 2017 | PRKDC | rs7003908 | SNV | 8:47858141 | Intronic | Iranian | 102/214 | 0.03 | 1.51 (1.04–2.18) | NOA | Candidate gene | Jahantigh et al. [182] | NO |
| 2017 | TNFR1 | rs767455 | SNV | 12:6341779 | Synonymous | Iranian | 108/119 | <0.001 | 2.30 (1.58–3.36) | NOA | Candidate gene | Ashrafzadeh et al. [183] | NO |
| 2017 | XRCC5 | rs6147172 | VNTR | 2:216109147 | Promoter | Iranian | 102/214 | 0.001 | 0.43 (0.26–0.73) | NOA | Candidate gene | Jahantigh et al. [182] | NO |
| 2017 | XRCC6 | rs2267437 | SNV | 22:41620695 | Intronic | Iranian | 102/214 | 0.0002 | 1.94 (1.37–2.75) | NOA | Candidate gene | Jahantigh et al. [182] | NO |
| 2018 | DICER1 | rs1057035 | SNV | 14:95087805 | 3′-UTR | Iranian | 135/120 | <0.05 | 1.49 (1.27–1.88) | NOA | Candidate gene | Moghbelinejad et al. [184] | NO |
| 2018 | DPF3 | rs10129954 | SNV | 14:72683993 | Intronic | Japanese | 83/713 | 7.40 × 10−3 | 2.05 (1.21–3.46) | NOA | Candidate gene | Sato et al. [185] | NO |
| 2018 | H2BFWT | rs553509 | SNV | X:104013293 | Missense | Iranian | 120/250 | 0.019 | 1.69 (1.09–2.62) | NOA | Candidate gene | Teimouri et al. [186] | NO |
| 2018 | IL1A | rs2071376 | SNV | 2:112777818 | Intronic | Iranian | 230/230 | 0.034 | 1.67 (1.04–2.68) | NOA | Candidate gene | Zamani-Badi et al. [187] | NO |
| 2018 | IL1A | rs17561 | SNV | 2:112779646 | Missense | Iranian | 230/230 | <0.0001 | 2.59 (1.67–4.04) | NOA | Candidate gene | Zamani-Badi et al. [188] | NO |
| 2018 | RNF212 | rs4045481 | SNV | 4:1096837 | Synonymous | Chinese | 220/248 | 0.003 | 1.50 (1.15–1.95) | NOA | Candidate gene | Yu et al. [189] | NO |
| 2019 | ERCC2 | rs13181 | SNV | 19:45351661 | Missense | Indo-European | 541/416 | 0.03 *,*,*,* | 1.59 (1.04–2.42) | NOA | Candidate gene | Singh et al. [190] | NO |
| 2019 | FSHB | rs10835638 | SNV | 11:30230805 | Intergenic | German | 659 | 0.017 *,*,*,*,* | 0.20 (0.06–0.70) | TESE– | Candidate gene | Busch et al. [191] | NO |
| 2019 | MSH3 | rs26279 | SNV | 5:80873118 | Missense | Northwest Chinese | 131/201 | <0.05 *,* | 2.62 (1.05–6.57) | NOA | Candidate gene | Zhao et al. [192] | NO |
| 2019 | HLA-B | rs4997052 | SNV | 6:31356367 | Missense | Han Chinese | 981/1657 | 2.26 × 10−5 | 1.30 (1.15–1.46) | NOA | GWAS imputation | Huang et al. [193] | NO |
| 2020 | STAG3 | rs1727130 | SNV | 7:100213841 | Intronic | Korean | 77/245 | 0.039 * | 1.64 (1.03–2.61) | NOA | Candidate gene | Nam et al. [194] | NO |
| 2020 | STAG3 | rs1052482 | SNV | 7:100214213 | 3′-UTR | Korean | 77/245 | 0.039 * | 1.64 (1.03–2.61) | NOA | Candidate gene | Nam et al. [194] | NO |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerván-Martín, M.; Castilla, J.A.; Palomino-Morales, R.J.; Carmona, F.D. Genetic Landscape of Nonobstructive Azoospermia and New Perspectives for the Clinic. J. Clin. Med. 2020, 9, 300. https://doi.org/10.3390/jcm9020300
Cerván-Martín M, Castilla JA, Palomino-Morales RJ, Carmona FD. Genetic Landscape of Nonobstructive Azoospermia and New Perspectives for the Clinic. Journal of Clinical Medicine. 2020; 9(2):300. https://doi.org/10.3390/jcm9020300
Chicago/Turabian StyleCerván-Martín, Miriam, José A. Castilla, Rogelio J. Palomino-Morales, and F. David Carmona. 2020. "Genetic Landscape of Nonobstructive Azoospermia and New Perspectives for the Clinic" Journal of Clinical Medicine 9, no. 2: 300. https://doi.org/10.3390/jcm9020300
APA StyleCerván-Martín, M., Castilla, J. A., Palomino-Morales, R. J., & Carmona, F. D. (2020). Genetic Landscape of Nonobstructive Azoospermia and New Perspectives for the Clinic. Journal of Clinical Medicine, 9(2), 300. https://doi.org/10.3390/jcm9020300