Exogenous Gonadotrophin Stimulation Induces Partial Maturation of Human Sertoli Cells in a Testicular Xenotransplantation Model for Fertility Preservation

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Human Testis Tissue Collection
2.3. Xenotransplantation Procedure
2.4. Treatment Regimens
2.5. Retrieval of Testicular Xenografts
2.6. Histological Analysis
2.7. Immunofluorescence and Confocal Microscopic Analysis
2.8. Statistical Analysis
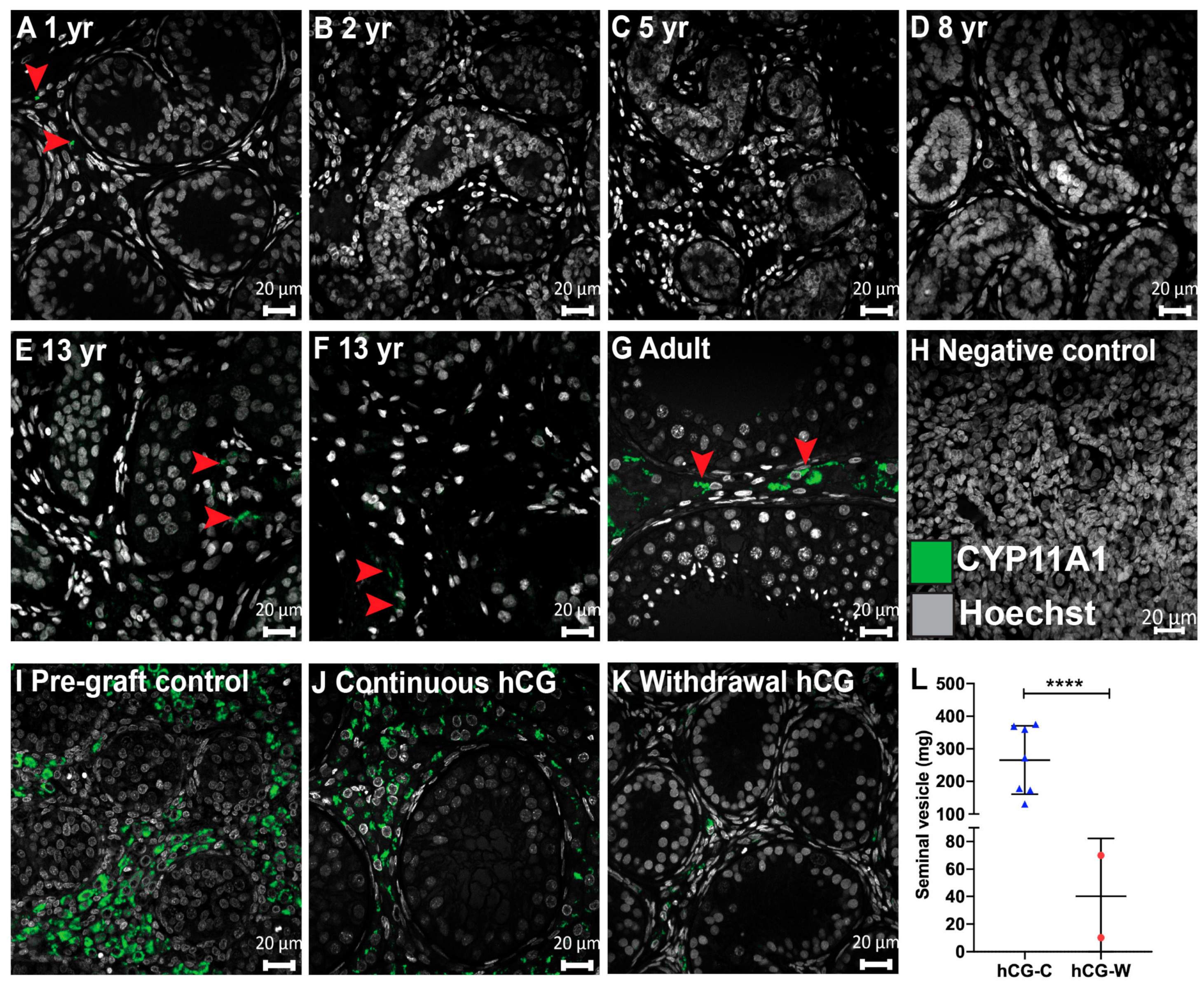
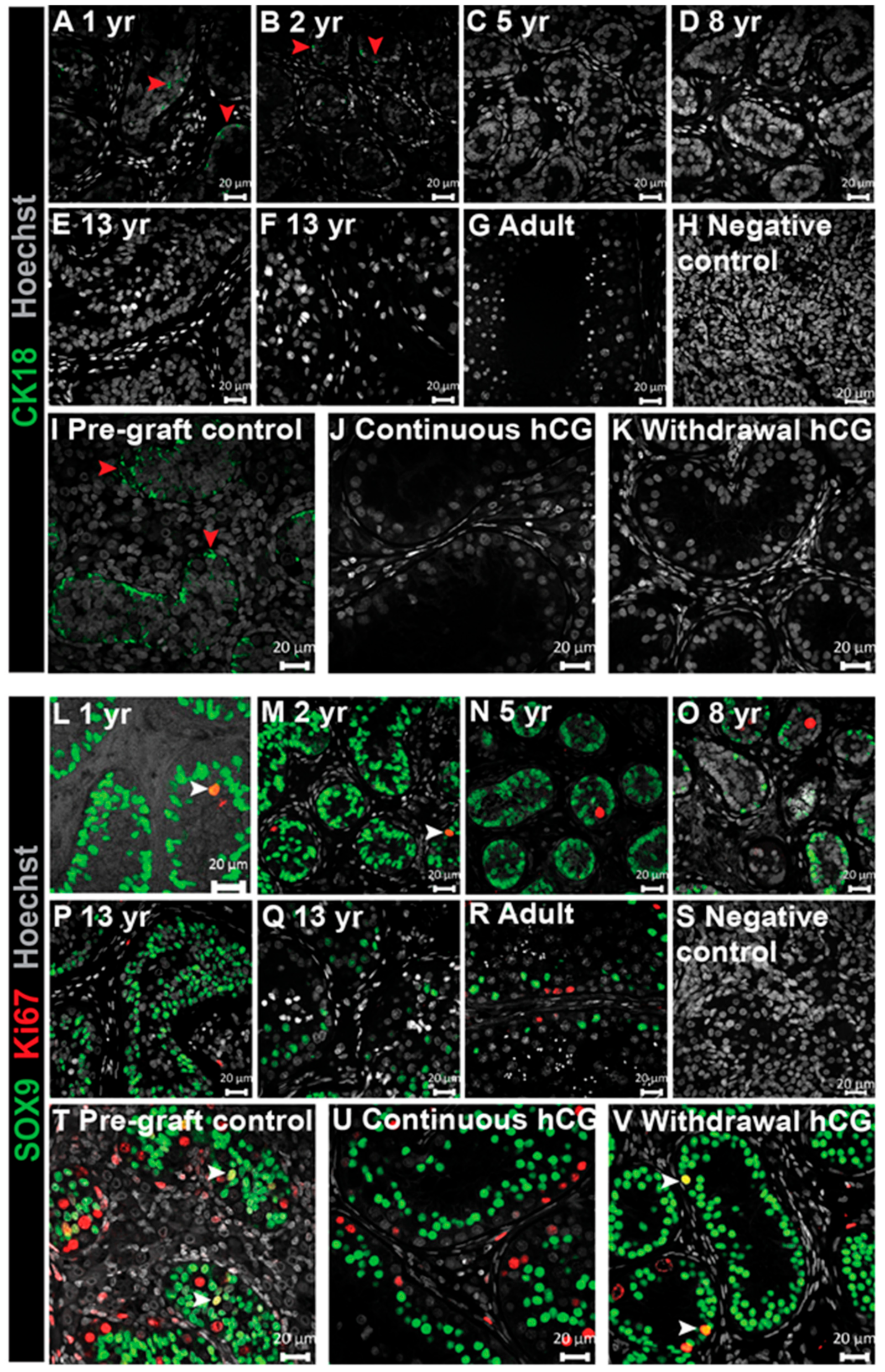
3. Results
3.1. Human Fetal Testis Xenograft Recovery
3.2. Lumen Development in Long-Term Continuously hCG-Exposed Testis Xenografts
3.3. Steroidogenesis in Long-Term Continuously hCG-Exposed Testis Xenografts
3.4. Expression of Immature Sertoli Cell Markers in Testis Xenografts
3.5. Analysis of Junctional Proteins in Testis Xenografts
3.6. Expression of AMH and AR in Testis Xenografts
3.7. Assessment of Germ Cell Survival and Differentiation in Long-Term Testis Xenografts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bhakta, N.; Force, L.M.; Allemani Catun, R.; Bray, F.; Coleman, M.P.; Steliarova-Foucher, E.; Frazier, A.L.; Robison, L.L.; Rodriguez-Galindo, C.; Fitzmaurice, C. Childhood Cancer Burden: A Review of Global Estimates. Lancet Oncol. 2019, 20, e42–e53. [Google Scholar] [CrossRef]
- Mitchell, R.T.; Saunders, P.T.; Sharpe, R.M.; Kelnar, C.J.; Wallace, W.H. Male Fertility and Strategies for Fertility Preservation Following Childhood Cancer Treatment. Endocr. Dev. 2009, 15, 101–134. [Google Scholar] [PubMed]
- Anderson, R.A.; Mitchell, R.T.; Kelsey, T.W.; Spears, N.; Telfer, E.E.; Wallace, W.H.B. Cancer Treatment and Gonadal Function: Experimental and Established Strategies for Fertility Preservation in Children and Young Adults. Lancet Diabet. Endocrinol. 2015, 3, 556–567. [Google Scholar] [CrossRef]
- Picton, H.M.; Wyns, C.; Anderson, R.A.; Goossens, E.; Jahnukainen, K.; Kliesch, S.; Mitchell, R.T.; Pennings, G.; Rives, N.; Tournaye, H.; et al. A European Perspective on Testicular Tissue Cryopreservation for Fertility Preservation in Prepubertal and Adolescent Boys†. Hum. Reprod. 2015, 30, 2463–2475. [Google Scholar] [CrossRef]
- Stukenborg, J.B.; Jahnukainen, K.; Hutka, M.; Mitchell, R.T. Cancer Treatment in Childhood and Testicular Function: The Importance of the Somatic Environment. Endocr. Connect. 2018, 7, R69–R87. [Google Scholar] [CrossRef]
- Fayomi, A.P.; Peters, K.; Sukhwani, M.; Valli-Pulaski, H.; Shetty, G.; Meistrich, M.L.; Houser, L.; Robertson, N.; Roberts, V.; Ramsey, C.; et al. Autologous Grafting of Cryopreserved Prepubertal Rhesus Testis Produces Sperm and Offspring. Science 2019, 363, 1314–1319. [Google Scholar] [CrossRef]
- Furman, W.L.; Fontanesi, J.; Hustu, O.; Dahl, G.V.; Kalwinsky, D.K.; Pui, C.H. Testicular Relapse in Children with Acute Nonlymphoblastic Leukemia. Cancer 1990, 66, 2095–2098. [Google Scholar] [CrossRef]
- Lal, A.E.; Kwan, M.; Al Mahr, L.; Zhou, D.; Ferrara, V.; Tobias, D.; O’Gorman, H.M.; Haber, M.; Norris, D.; Marshall, G.M. Molecular Detection of Acute Lymphoblastic Leukaemia in Boys with Testicular Relapse. Mol. Pathol. 1998, 51, 277–281. [Google Scholar] [CrossRef]
- Ding, L.-W.; Sun, Q.-Y.; Anand, M.; Tan, K.-T.; Chien, W.; Lin, D.-C.; Jiang, Y.-Y.; Xu, L.; Garg, M.; Lao, Z.-T.; et al. Mutational Profiling of Acute Lymphoblastic Leukemia with Testicular Relapse. J. Hematol. Oncol. 2017, 10, 65. [Google Scholar] [CrossRef]
- Barredo, J.C.; Hastings, C.; Lu, X.; Devidas, M.; Chen, Y.; Armstrong, D.; Winick, N.; Wood, B.L.; Yanofsky, R.; Loh, M.; et al. Isolated Late Testicular Relapse of B-Cell Acute Lymphoblastic Leukemia Treated with Intensive Systemic Chemotherapy and Response-Based Testicular Radiation: A Children’s Oncology Group Study. Pediatr. Blood Cancer 2018, 65, e26928. [Google Scholar] [CrossRef]
- Pui, C.H. Is Testicular Irradiation Necessary for Patients with Acute Lymphoblastic Leukemia and Testicular Relapse? Pediatr. Blood Cancer 2018, 65, e26977. [Google Scholar] [CrossRef] [PubMed]
- França, L.R.; Hess, R.A.; Dufour, J.M.; Hofmann, M.C.; Griswold, M.D. The Sertoli Cell: One Hundred Fifty Years of Beauty and Plasticity. Andrology 2016, 4, 189–212. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, L.; Stanton, P.; de Kretser, D.M. Endocrinology of the Male Reproductive System and Spermatogenesis. In Endotext; De Groot, L., Chrousos, J.G., Dungan, K., Feingold, K.R., Grossman, A., Hershman, J.M., Koch, C., Korbonits, M., McLachlan, R., New, M., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2017. [Google Scholar]
- Gow, A.; Southwood, C.M.; Li, J.S.; Pariali, M.G.; Riordan, P.; Brodie, S.E.; Danias, J.; Bronstein, J.M.; Kachar, B.; Lazzarini, R.A. Cns Myelin and Sertoli Cell Tight Junction Strands Are Absent in Osp/Claudin-11 Null Mice. Cell 1999, 99, 649–659. [Google Scholar] [CrossRef]
- De Gendt, K.; Swinnen, J.V.; Saunders, P.T.; Schoonjans, L.; Dewerchin, M.; Devos, A.; Tan, K.; Atanassova, N.; Claessens, F.; Lecureuil, C.; et al. A Sertoli Cell-Selective Knockout of the Androgen Receptor Causes Spermatogenic Arrest in Meiosis. Proc. Natl. Acad. Sci. USA 2004, 101, 1327–1332. [Google Scholar] [CrossRef] [PubMed]
- Brehm, R.; Zeiler, M.; Rüttinger, C.; Herde, K.; Kibschull, M.; Winterhager, E.; Willecke, K.; Guillou, F.; Lécureuil, C.; Steger, K.; et al. A Sertoli Cell-Specific Knockout of Connexin43 Prevents Initiation of Spermatogenesis. Am. J. Pathol. 2007, 171, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Maymon, B.B.; Yogev, L.; Paz, G.; Kleiman, S.E.; Schreiber, L.; Botchan, A.; Hauser, R.; Yavetz, H. Sertoli Cell Maturation in Men with Azoospermia of Different Etiologies. Fertil. Steril. 2002, 77, 904–909. [Google Scholar] [CrossRef]
- Hutka, M.; Smith, L.B.; Mitchell, R.T. Xenotransplantation as a Model for Human Testicular Development. Differentiation 2017, 97, 44–53. [Google Scholar] [CrossRef]
- Mitchell, R.T.; Saunders, P.T.; Childs, A.J.; Cassidy-Kojima, C.; Anderson, R.A.; Wallace, W.H.; Kelnar, C.J.; Sharpe, R.M. Xenografting of Human Fetal Testis Tissue: A New Approach to Study Fetal Testis Development and Germ Cell Differentiation. Hum. Reprod. 2010, 25, 2405–2414. [Google Scholar] [CrossRef]
- Nistal, M.; Paniagua, R.; Gonzalez-Peramato, P.; Reyes-Mugica, M. Perspectives in Pediatric Pathology, Chapter 3. Testicular Development from Birth to Puberty: Systematic Evaluation of the Prepubertal Testis. Pediatr. Dev. Pathol. 2015, 18, 173–186. [Google Scholar] [CrossRef]
- Yu, J.; Cai, Z.M.; Wan, H.J.; Zhang, F.T.; Ye, J.; Fang, J.Z.; Gui, Y.T.; Ye, J.X. Development of Neonatal Mouse and Fetal Human Testicular Tissue as Ectopic Grafts in Immunodeficient Mice. Asian J. Androl. 2006, 8, 393–403. [Google Scholar] [CrossRef]
- Lo, K.C.; Yildiz, C.; Zhu, Y.; Lambourne, M.D.; Mullen, J.B.; Samplaski, M.K.; Jarvi, K.A.; McKerlie, C. Human Fetal Testicular Tissue Xenotransplantation: A Platform to Study the Effect of Gonadotropins on Human Germ Cell Development in Utero. J. Urol. 2015, 194, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Rathi, R.; Zeng, W.; Megee, S.; Conley, A.; Meyers, S.; Dobrinski, I. Maturation of Testicular Tissue from Infant Monkeys after Xenografting into Mice. Endocrinology 2008, 149, 5288–5296. [Google Scholar] [CrossRef] [PubMed]
- Schlatt, S.; Honaramooz, A.; Boiani, M.; Scholer, H.R.; Dobrinski, I. Progeny from Sperm Obtained after Ectopic Grafting of Neonatal Mouse Testes. Biol. Reprod. 2003, 68, 2331–2335. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, R.M.; McKinnell, C.; Kivlin, C.; Fisher, J.S. Proliferation and Functional Maturation of Sertoli Cells, and Their Relevance to Disorders of Testis Function in Adulthood. Reproduction 2003, 125, 769–784. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, S.; Brehm, R.; Bergmann, M.; Cooke, P.S. Role of Connexin 43 in Sertoli Cells of Testis. Ann. N. Y. Acad. Sci. 2007, 1120, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Li, M.W.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Connexin 43 Is Critical to Maintain the Homeostasis of the Blood-Testis Barrier Via Its Effects on Tight Junction Reassembly. Proc. Natl. Acad. Sci. USA 2010, 107, 17998–18003. [Google Scholar] [CrossRef]
- Mazaud-Guittot, S.; Meugnier, E.; Pesenti, S.; Wu, X.; Vidal, H.; Gow, A.; Le Magueresse-Battistoni, B. Claudin 11 Deficiency in Mice Results in Loss of the Sertoli Cell Epithelial Phenotype in the Testis. Biol. Reprod. 2010, 82, 202–213. [Google Scholar] [CrossRef]
- Haverfield, J.T.; Stanton, P.G.; Meachem, S.J. 14—Adult Sertoli Cell Differentiation Status in Humans. In Sertoli Cell Biology, 2nd ed.; Griswold, M.D., Ed.; Academic Press: Oxford, UK, 2015; pp. 409–436. [Google Scholar]
- Chang, C.; Chen, Y.T.; Yeh, S.D.; Xu, Q.; Wang, R.S.; Guillou, F.; Lardy, H.; Yeh, S. Infertility with Defective Spermatogenesis and Hypotestosteronemia in Male Mice Lacking the Androgen Receptor in Sertoli Cells. Proc. Natl. Acad. Sci. USA 2004, 101, 6876–6881. [Google Scholar] [CrossRef]
- Tan, K.A.L.; De Gendt, K.; Atanassova, N.; Walker, M.; Sharpe, R.M.; Saunders, P.T.K.; Denolet, E.; Verhoeven, G. The Role of Androgens in Sertoli Cell Proliferation and Functional Maturation: Studies in Mice with Total or Sertoli Cell-Selective Ablation of the Androgen Receptor. Endocrinology 2005, 146, 2674–2683. [Google Scholar] [CrossRef]
- Haverfield, J.T.; Meachem, S.J.; O’Bryan, M.K.; McLachlan, R.I.; Stanton, P.G. Claudin-11 and Connexin-43 Display Altered Spatial Patterns of Organization in Men with Primary Seminiferous Tubule Failure Compared with Controls. Fertil. Steril. 2013, 100, 658–666. [Google Scholar] [CrossRef]
- Bressler, R.S.; Lustbader, I.J. Effect of Testosterone on Development of the Lumen in Seminiferous Tubules of the Rat. Andrologia 1978, 10, 291–298. [Google Scholar] [PubMed]
- Franke, F.E.; Pauls, K.; Rey, R.; Marks, A.; Bergmann, M.; Steger, K. Differentiation Markers of Sertoli Cells and Germ Cells in Fetal and Early Postnatal Human Testis. Anat. Embryol. 2004, 209, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Stosiek, P.; Kasper, M.; Karsten, U. Expression of Cytokeratins 8 and 18 in Human Sertoli Cells of Immature and Atrophic Seminiferous Tubules. Differentiation 1990, 43, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Edelsztein, N.Y.; Rey, R.A. Importance of the Androgen Receptor Signaling in Gene Transactivation and Transrepression for Pubertal Maturation of the Testis. Cells 2019, 8, 861. [Google Scholar] [CrossRef]
- Hazra, R.; Jimenez, M.; Desai, R.; Handelsman, D.J.; Allan, C.M. Sertoli Cell Androgen Receptor Expression Regulates Temporal Fetal and Adult Leydig Cell Differentiation, Function, and Population Size. Endocrinology 2013, 154, 3410–3422. [Google Scholar] [CrossRef]
- Wyns, C.; Curaba, M.; Martinez-Madrid, B.; Van Langendonckt, A.; François-Xavier, W.; Donnez, J. Spermatogonial Survival after Cryopreservation and Short-Term Orthotopic Immature Human Cryptorchid Testicular Tissue Grafting to Immunodeficient Mice. Hum. Reprod. 2007, 22, 1603–1611. [Google Scholar] [CrossRef]
- Goossens, E.; Geens, M.; De Block, G.; Tournaye, H. Spermatogonial Survival in Long-Term Human Prepubertal Xenografts. Fertil. Steril. 2008, 90, 2019–2022. [Google Scholar] [CrossRef]
- Van Saen, D.; Goossens, E.; Bourgain, C.; Ferster, A.; Tournaye, H. Meiotic Activity in Orthotopic Xenografts Derived from Human Postpubertal Testicular Tissue. Hum. Reprod. 2011, 26, 282–293. [Google Scholar] [CrossRef]
- Wyns, C.; Van Langendonckt, A.; Wese, F.X.; Donnez, J.; Curaba, M. Long-Term Spermatogonial Survival in Cryopreserved and Xenografted Immature Human Testicular Tissue. Hum. Reprod. 2008, 23, 2402–2414. [Google Scholar] [CrossRef]
- Jonathan, P.; Van Langendonckt, A.; Many, M.C.; Wese, F.X.; Wyns, C. Vitrification Preserves Proliferation Capacity in Human Spermatogonia. Hum. Reprod. 2013, 28, 578–589. [Google Scholar]
- Van Saen, D.; Goossens, E.; Haentjens, P.; Baert, Y.; Tournaye, H. Exogenous Administration of Recombinant Human Fsh Does Not Improve Germ Cell Survival in Human Prepubertal Xenografts. Reprod. Biomed. Online 2013, 26, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, A.A.; Raivio, T.; Pitteloud, N. Gonadotrophin Replacement for Induction of Fertility in Hypogonadal Men. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.A.; Ramasamy, R. Indications for the Use of Human Chorionic Gonadotropic Hormone for the Management of Infertility in Hypogonadal Men. Transl. Androl. Urol. 2018, 7, S348–S352. [Google Scholar] [CrossRef] [PubMed]
- Prior, M.; Stewart, J.; McEleny, K.; Dwyer, A.A.; Quinton, R. Fertility Induction in Hypogonadotropic Hypogonadal Men. Clin. Endocrinol. 2018, 89, 712–718. [Google Scholar] [CrossRef]
- Fallarino, F.; Luca, G.; Calvitti, M.; Mancuso, F.; Nastruzzi, C.; Fioretti, M.C.; Grohmann, U.; Becchetti, E.; Burgevin, A.; Kratzer, R.; et al. Therapy of Experimental Type 1 Diabetes by Isolated Sertoli Cell Xenografts Alone. J. Exp. Med. 2009, 206, 2511–2526. [Google Scholar] [CrossRef]
- Hansen, B.C.; Cameron, D.; Calafiore, R.; Luca, G.; Linden, E.H.; Szekeres, C. Effects of Sertoli Cells Implantation on Type 2 Diabetes in Nonhuman Primates. FASEB J. 2013, 27, 1154–1163. [Google Scholar]
- Chiappalupi, S.; Salvadori, L.; Luca, G.; Riuzzi, F.; Calafiore, R.; Donato, R.; Sorci, G. Employment of Microencapsulated Sertoli Cells as a New Tool to Treat Duchenne Muscular Dystrophy. J. Funct. Morphol. Kinesiol. 2017, 2, 47. [Google Scholar] [CrossRef]
- Luca, G.; Arato, I.; Sorci, G.; Cameron, D.F.; Hansen, B.C.; Baroni, T.; Donato, R.; White, D.G.J.; Calafiore, R. Sertoli Cells for Cell Transplantation: Pre-Clinical Studies and Future Perspectives. Andrology 2018, 6, 385–395. [Google Scholar] [CrossRef]
- Das, M.; Howell, M.; Foran, E.A.; Iyre, R.; Mohapatra, S.S.; Mohapatra, S. Sertoli Cells Loaded with Doxorubicin in Lipid Micelles Reduced Tumor Burden and Dox-Induced Toxicity. Cell Transplant. 2017, 26, 1694–1702. [Google Scholar] [CrossRef]
- Halley, K.; Dyson, E.L.; Kaur, G.; Mital, P.; Uong, P.M.; Dass, B.; Crowell, S.N.; Dufour, J.M. Delivery of a Therapeutic Protein by Immune-Privileged Sertoli Cells. Cell Transplant. 2010, 19, 1645–1657. [Google Scholar] [CrossRef]
- Cupp, A.S. Sertoli Cell-Based Gene Therapy? Biol. Reprod. 2014, 91, 40. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Thompson, L.A.; Babcock, R.L.; Mueller, K.; Dufour, J.M. Sertoli Cells Engineered to Express Insulin to Lower Blood Glucose in Diabetic Mice. DNA Cell Biol. 2018, 37, 680–690. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Cell Type | Supplier | Cat. No. | Dilution |
|---|---|---|---|---|
| CK18 (DC-10) | Sertoli cell (immature) | Santa Cruz Biotechnology | sc-6259 | 1:200 |
| MIS (C-20) (AMH) | Sertoli cell (immature) | Santa Cruz Biotechnology | sc-6886 | 1:1000 |
| AR (N-20) | Sertoli cell * (mature) | Santa Cruz Biotechnology | sc-816 | 1:2000 |
| SOX9 | Sertoli cell (all stages) | Millipore | AB5535 | 1:10,000 |
| CX43 | Sertoli cell (BTB) | Cell Signaling Technology | 3512 | 1:300 |
| CLDN11 | Sertoli cell (BTB) | Thermo Fisher Scientific | 36-4500 | 1:500 |
| CYP11A1 | Leydig cell (steroidogenesis) | Sigma-Aldrich | HPA016436 | 1:5000 |
| AP2γ | Germ cell (Gonocyte) | Santa Cruz Biotechnology | sc-12762 | 1:75 |
| MAGE-A4 | Germ cell (Spermatogonia) | Gift # | - | 1:200 |
| γH2AX | Germ cell ** (meiotic) | Abcam | AB26350 | 1:1000 |
| DMC1 | Germ cell (meiotic) | Abcam | AB11054 | 1:4000 |
| BOLL | Germ cell (meiotic) | Novus Bio | H00066037-M03 | 1:1000 |
| Ki67 | Proliferating cell | Abcam | AB16667 | 1:100 |
| c-PARP | Apoptotic cell | Cell Signaling Technology | 5625 | 1:100 |
| c-Caspase-3 | Apoptotic cell | Cell Signaling Technology | 9661 | 1:100 |
| Hoechst | DNA counterstain | Thermo Fisher Scientific | 33342 | 1:4000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hutka, M.; Smith, L.B.; Goossens, E.; Wallace, W.H.B.; Stukenborg, J.-B.; Mitchell, R.T. Exogenous Gonadotrophin Stimulation Induces Partial Maturation of Human Sertoli Cells in a Testicular Xenotransplantation Model for Fertility Preservation. J. Clin. Med. 2020, 9, 266. https://doi.org/10.3390/jcm9010266
Hutka M, Smith LB, Goossens E, Wallace WHB, Stukenborg J-B, Mitchell RT. Exogenous Gonadotrophin Stimulation Induces Partial Maturation of Human Sertoli Cells in a Testicular Xenotransplantation Model for Fertility Preservation. Journal of Clinical Medicine. 2020; 9(1):266. https://doi.org/10.3390/jcm9010266
Chicago/Turabian StyleHutka, Marsida, Lee B. Smith, Ellen Goossens, W. Hamish B. Wallace, Jan-Bernd Stukenborg, and Rod T. Mitchell. 2020. "Exogenous Gonadotrophin Stimulation Induces Partial Maturation of Human Sertoli Cells in a Testicular Xenotransplantation Model for Fertility Preservation" Journal of Clinical Medicine 9, no. 1: 266. https://doi.org/10.3390/jcm9010266
APA StyleHutka, M., Smith, L. B., Goossens, E., Wallace, W. H. B., Stukenborg, J.-B., & Mitchell, R. T. (2020). Exogenous Gonadotrophin Stimulation Induces Partial Maturation of Human Sertoli Cells in a Testicular Xenotransplantation Model for Fertility Preservation. Journal of Clinical Medicine, 9(1), 266. https://doi.org/10.3390/jcm9010266