Profiling of Omega-Polyunsaturated Fatty Acids and Their Oxidized Products in Salmon after Different Cooking Methods



Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Samples and Heat Treatment
- Boiling: Salmon filets were boiled individually on a heat plate in 800 mL of water initially at 99 ± 1 °C for 10 min.
- Pan-frying: Salmon filets were fried on a medium-sized frying pan on a heat plate to a center temperature of 200 ± 10 °C for 10 min and were flipped over half way (5 min) through cooking.
- Oven-baking: The oven was pre-heated to 200 °C. Salmon filets were individually placed on an aluminum foil-coated metal tray and baked at 200 °C for 15 min.
2.2. Extraction of Crude Oil
2.3. Measurement of Lipid Peroxidation
2.4. Extraction of Polyunsaturated Fatty Acids and Oxidized Products
2.5. Quantification of Polyunsaturated Fatty Acids and Its Oxidized Products
2.6. Extraction of Reactive Aldehydes, 4-Hydroxy-2(E)-Hexenal and 4-Hydroxy-2-Nonenal
2.7. Quantification of 4-HHE and 4-HNE
2.8. Statistical Analysis
3. Results
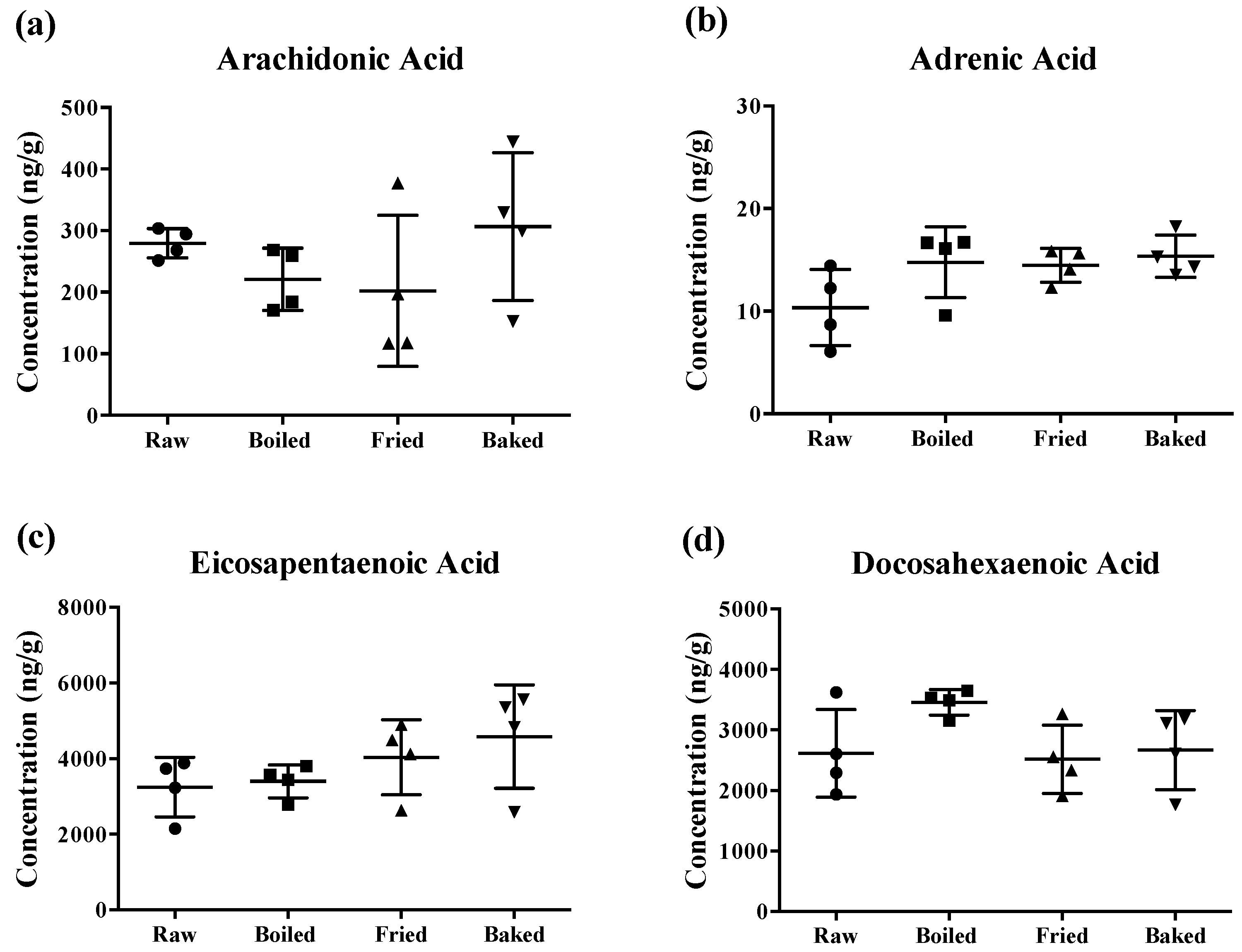
3.1. Different Cooking Methods Did Not Reduce PUFAs’ Levels in Salmon
3.2. Elevated Lipid Peroxidation Was Found in Pan-Fried and Oven-Baked Salmons
3.3. Elevated 4-HHE and 4-HNE Were Found in Cooked Salmons
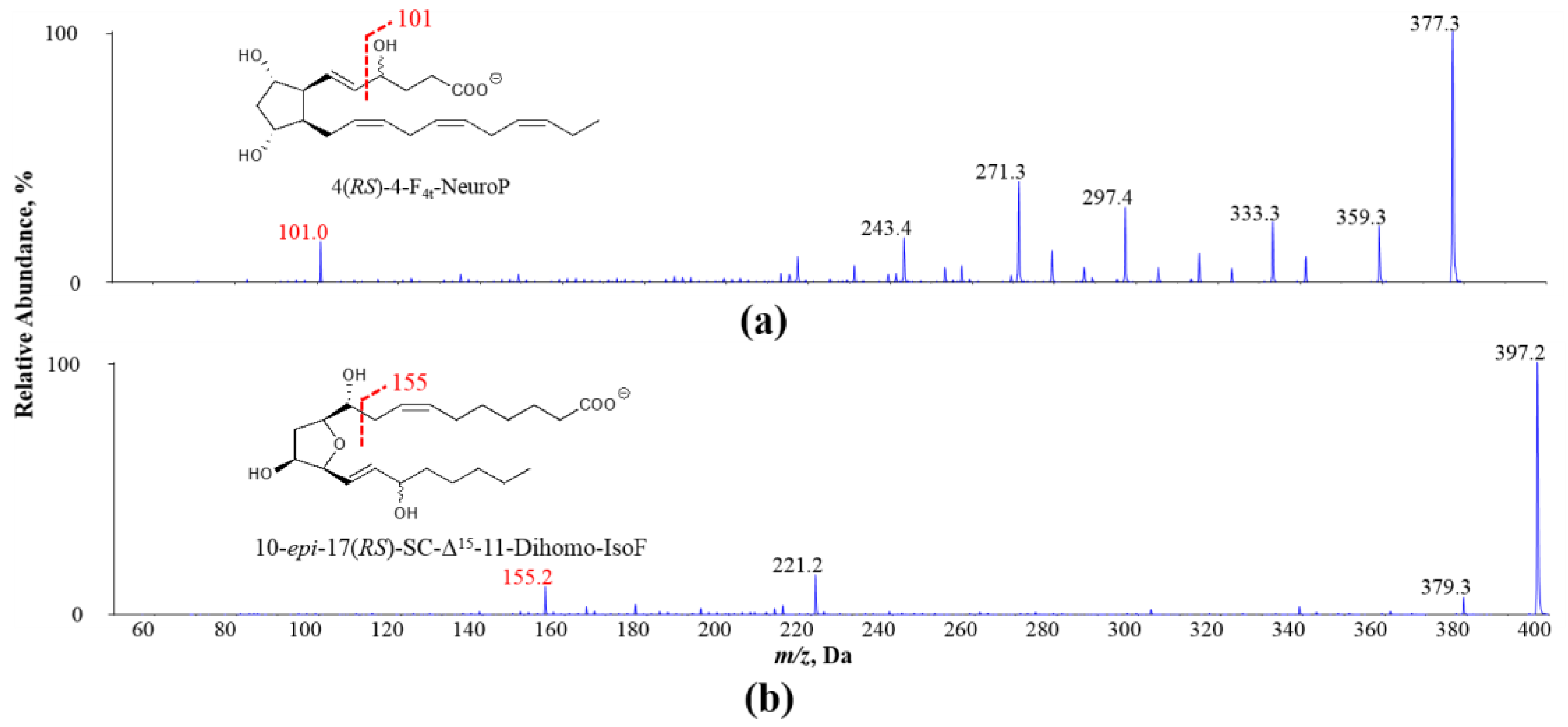
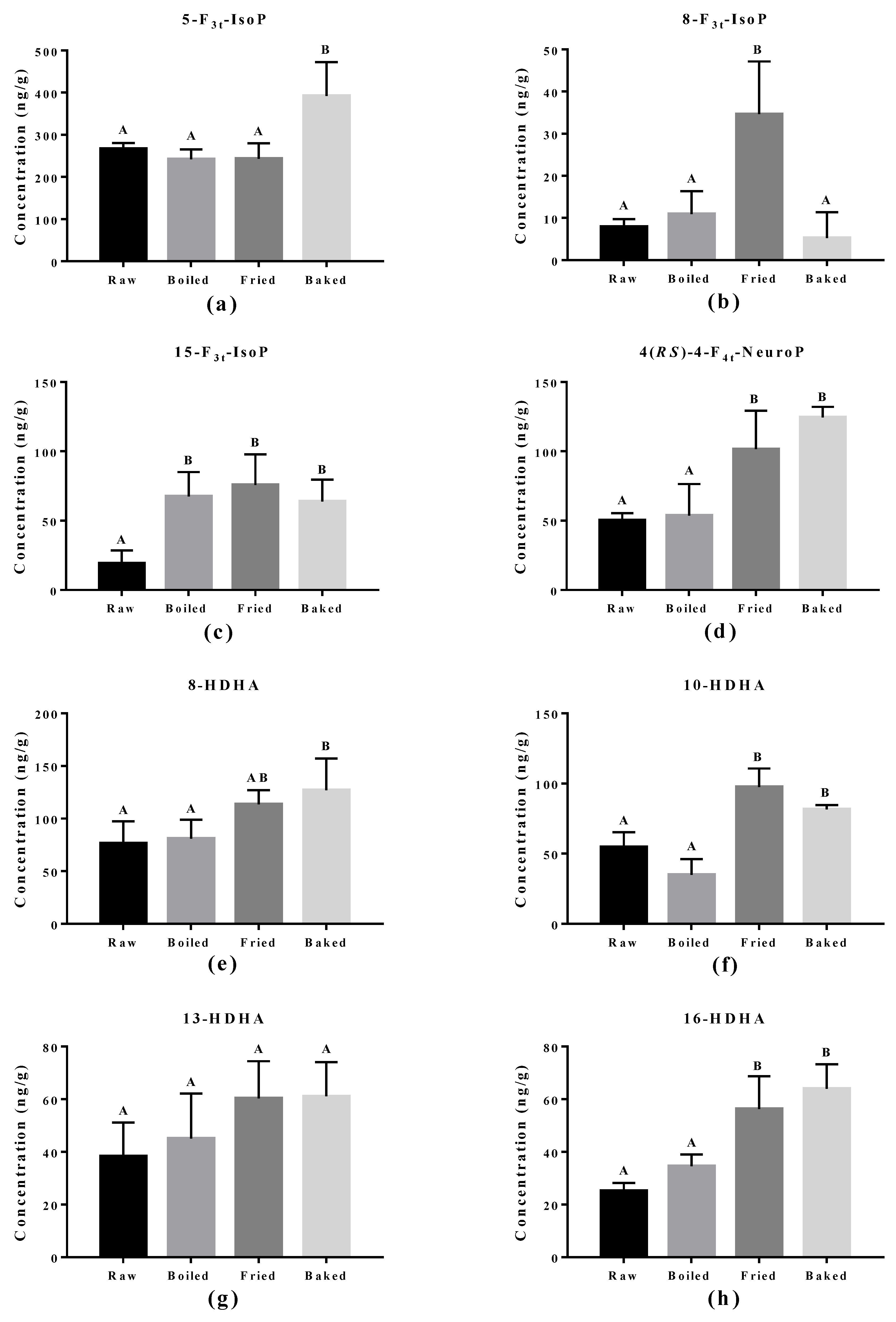
3.4. Elevated Oxidized PUFA Products Were Found in Cooked Salmons
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rajaram, S. Health benefits of plant-derived α-linolenic acid. Am. J. Clin. Nutr. 2014, 100 (Suppl. 1), 443S–448S. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, H.; Ba, G.N.; Couvreur, P.; Tew, K.D. Polyunsaturated fatty acids (PUFA) and eicosanoids in human health and pathologies. Biomed. Pharmacother. 2002, 56, 215–222. [Google Scholar] [CrossRef]
- Calder, P.C. N-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, 1505S–1519S. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, P.; Hu, X.; Zhang, F.; Chen, J.; Gao, Y. Omega-3 polyunsaturated fatty acids in the brain: Metabolism and neuroprotection. Front. Biosci. 2011, 16, 2653–2670. [Google Scholar] [CrossRef]
- Thomas, J.; Thomas, C.J.; Radcliffe, J.; Itsiopoulos, C. Omega-3 fatty acids in early prevention of inflammatory neurodegenerative disease: A focus on Alzheimer’s disease. Biomed. Res. Int. 2015, 2015, 172801. [Google Scholar] [CrossRef] [PubMed]
- Eser, P.O.; Vanden Heuvel, J.P.; Araujo, J.; Thompson, J.T. Marine- and plant-derived omega-3 fatty acids differentially regulate prostate cancer cell proliferation. Mol. Clin. Oncol. 2013, 1, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Calder, P.C. Conversion of α-linolenic acid to longer-chain polyunsaturated fatty acids in human adults. Reprod. Nutr. Dev. 2005, 45, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, T.; Shozen, K.; Ushio, H.; Koizumi, C. Effects of grilling on formation of cholesterol oxides in seafood products rich in polyunsaturated fatty acids. Food Sci. Technol. 1996, 29, 94–99. [Google Scholar] [CrossRef]
- Sampaio, G.R.; Bastos, D.H.M.; Soares, R.A.M.; Queiroz, Y.S.; Torres, E.A.F.S. Fatty acids and cholesterol oxidation in salted and dried shrimp. Food Chem. 2006, 95, 344–351. [Google Scholar] [CrossRef]
- Sant’Ana, L.S.; Mancini, J. Influence of the addition of antioxidants in vivo on the fatty acid composition of fish fillets. Food Chem. 2000, 68, 175–178. [Google Scholar] [CrossRef]
- Tarley, C.R.T.; Visentainer, J.V.; Matsushita, M.; de Souza, N.E. Proximate composition, cholesterol and fatty acids profile of canned sardines (Sardinella brasiliensis) in soybean oil and tomato sauce. Food Chem. 2004, 88, 1–6. [Google Scholar] [CrossRef]
- Echarte, M.; Zulet, M.A.; Astiasaran, I. Oxidation process affecting fatty acids and cholesterol in fried and roasted salmon. J. Agric. Food Chem. 2001, 49, 5662–5667. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, M.I.; Sushchik, N.N.; Gubanenko, G.A.; Demirchieva, S.M.; Kalachova, G.S. Effect of way of cooking on content of essential polyunsaturated fatty acids in muscle tissue of humpback salmon (Oncorhynchus gorbuscha). Food Chem. 2006, 96, 446–451. [Google Scholar] [CrossRef]
- Stolyhwo, A.; Kolodziejska, I.; Sikorski, Z.E. Long chain polyunsaturated fatty acids in smoked atlantic mackerel and baltic sprats. Food Chem. 2006, 94, 589–595. [Google Scholar] [CrossRef]
- Milne, G.L.; Dai, Q.; Roberts, L.J., 2nd. The isoprostanes—25 years later. Biochim. Biophys. Acta 2015, 1851, 433–445. [PubMed]
- Alkazemi, D.; Jackson, R.L., 2nd; Chan, H.M.; Kubow, S. Increased F3-isoprostanes in the canadian inuit population could be cardioprotective by limiting F2-isoprostane production. J. Clin. Endocrinol. Metab. 2016, 101, 3264–3271. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Oger, C.; Thireau, J.; Roussel, J.; Mercier-Touzet, O.; Faure, D.; Pinot, E.; Farah, C.; Taber, D.F.; Cristol, J.P.; et al. Nonenzymatic lipid mediators, neuroprostanes, exert the antiarrhythmic properties of docosahexaenoic acid. Free Radic. Biol. Med. 2015, 86, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Fauconnier, J.; Oger, C.; Farah, C.; Angebault-Prouteau, C.; Thireau, J.; Bideaux, P.; Scheuermann, V.; Bultel-Ponce, V.; Demion, M.; et al. Non-enzymatic oxidized metabolite of DHA, 4(RS)-4-F4T-neuroprostane protects the heart against reperfusion injury. Free Radic. Biol. Med. 2017, 102, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Gladine, C.; Newman, J.W.; Durand, T.; Pedersen, T.L.; Galano, J.M.; Demougeot, C.; Berdeaux, O.; Pujos-Guillot, E.; Mazur, A.; Comte, B. Lipid profiling following intake of the omega 3 fatty acid DHA identifies the peroxidized metabolites F4-neuroprostanes as the best predictors of atherosclerosis prevention. PLoS ONE 2014, 9, e89393. [Google Scholar] [CrossRef] [PubMed]
- Long, E.K.; Murphy, T.C.; Leiphon, L.J.; Watt, J.; Morrow, J.D.; Milne, G.L.; Howard, J.R.H.; Picklo, M.J. Trans-4-hydroxy-2-hexenal is a neurotoxic product of docosahexaenoic (22:6; n-3) acid oxidation. J. Neurochem. 2008, 105, 714–724. [Google Scholar] [CrossRef] [PubMed]
- Ishikado, A.; Morino, K.; Nishio, Y.; Nakagawa, F.; Mukose, A.; Sono, Y.; Yoshioka, N.; Kondo, K.; Sekine, O.; Yoshizaki, T.; et al. 4-hydroxy hexenal derived from docosahexaenoic acid protects endothelial cells via NRF2 activation. PLoS ONE 2013, 8, e69415. [Google Scholar] [CrossRef] [PubMed]
- Ishikado, A.; Nishio, Y.; Morino, K.; Ugi, S.; Kondo, H.; Makino, T.; Kashiwagi, A.; Maegawa, H. Low concentration of 4-hydroxy hexenal increases heme oxygenase-1 expression through activation of NRF2 and antioxidative activity in vascular endothelial cells. Biochem. Biophys. Res. Commun. 2010, 402, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Toru, T.; Yuichiro, M.; Mayumi, M. Determination of peroxide value by the colorimetric iodine method with protection of iodide as cadmium complex. Lipids 1978, 13, 147–151. [Google Scholar]
- Papastergiadis, A.; Mubiru, E.; Van Langenhove, H.; De Meulenaer, B. Malondialdehyde measurement in oxidized foods: Evaluation of the spectrophotometric thiobarbituric acid reactive substances (TBARS) test in various foods. J. Agric. Food Chem. 2012, 60, 9589–9594. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, A.; Le Faouder, P.; Vigor, C.; Oger, C.; Galano, J.M.; Dray, C.; Lee, J.C.; Valet, P.; Gladine, C.; Durand, T.; et al. Simultaneous quantitative profiling of 20 isoprostanoids from omega-3 and omega-6 polyunsaturated fatty acids by LC-MS/MS in various biological samples. Anal. Chim. Acta 2016, 921, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Jenner, A.M.; Halliwell, B. Rapid preparation of human urine and plasma samples for analysis of F2-isoprostanes by gas chromatography-mass spectrometry. Biochem. Biophys. Res. Commun. 2004, 320, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.Y.; Wong, C.K.; Oger, C.; Durand, T.; Galano, J.M.; Lee, J.C. Prenatal exposure to the contaminant perfluorooctane sulfonate elevates lipid peroxidation during mouse fetal development but not in the pregnant dam. Free Radic. Res. 2015, 49, 1015–1025. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.Y.; Crauste, C.; Wang, H.; Leung, H.H.; Vercauteren, J.; Galano, J.M.; Oger, C.; Durand, T.; Wan, J.M.; Lee, J.C. Extra virgin olive oil reduced polyunsaturated fatty acid and cholesterol oxidation in rodent liver: Is this accounted for hydroxytyrosol-fatty acid conjugation? Chem. Res. Toxicol. 2016, 29, 1689–1698. [Google Scholar] [CrossRef] [PubMed]
- Douny, C.; Bayram, P.; Brose, F.; Degand, G.; Scippo, M.L. Development of an LC-MS/MS analytical method for the simultaneous measurement of aldehydes from polyunsaturated fatty acids degradation in animal feed. Drug Test. Anal. 2016, 8, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Gluck, T.; Alter, P. Marine omega-3 highly unsaturated fatty acids: From mechanisms to clinical implications in heart failure and arrhythmias. Vasc. Pharmacol. 2016, 82, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J.; American Heart Association. Nutrition, C. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef] [PubMed]
- Bastias, J.M.; Balladares, P.; Acuna, S.; Quevedo, R.; Munoz, O. Determining the effect of different cooking methods on the nutritional composition of salmon (Salmo salar) and chilean jack mackerel (Trachurus murphyi) fillets. PLoS ONE 2017, 12, e0180993. [Google Scholar] [CrossRef] [PubMed]
- Flaskerud, K.; Bukowski, M.; Golovko, M.; Johnson, L.; Brose, S.; Ali, A.; Cleveland, B.; Picklo, M.; Raatz, S. Effects of cooking techniques on fatty acid and oxylipin content of farmed rainbow trout (Oncorhynchus mykiss). Food Sci. Nutr. 2017, 5, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Sioen, I.; Haak, L.; Raes, K.; Hermans, C.; De Henauw, S.; De Smet, S.; Van Camp, J. Effects of pan-frying in margarine and olive oil on the fatty acid composition of cod and salmon. Food Chem. 2006, 98, 609–617. [Google Scholar] [CrossRef]
- Gladyshev, M.I.; Sushchik, N.N.; Gubanenko, G.A.; Demirchieva, S.M.; Kalachova, G.S. Effect of boiling and frying on the content of essential polyunsaturated fatty acids in muscle tissue of four fish species. Food Chem. 2007, 101, 1694–1700. [Google Scholar] [CrossRef]
- McGee, H. On Food and Cooking: The Science and Lore of the Kitchen; Scribner: New York, NY, USA, 2004. [Google Scholar]
- Roy, J.; Le Guennec, J.Y.; Galano, J.M.; Thireau, J.; Bultel-Ponce, V.; Demion, M.; Oger, C.; Lee, J.C.; Durand, T. Non-enzymatic cyclic oxygenated metabolites of omega-3 polyunsaturated fatty acid: Bioactive drugs? Biochimie 2016, 120, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Long, E.K.; Picklo, M.J. Trans-4-hydroxy-2-hexenal, a product of n-3 fatty acid peroxidation: Make some room HNE. Free Radic. Biol. Med. 2010, 49, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Larsson, K.; Harrysson, H.; Havenaar, R.; Alminger, M.; Undeland, I. Formation of malondialdehyde (MDA), 4-hydroxy-2-hexenal (HHE) and 4-hydroxy-2-nonenal (HNE) in fish and fish oil during dynamic gastrointestinal in vitro digestion. Food Funct. 2016, 7, 1176–1187. [Google Scholar] [CrossRef] [PubMed]
- Awada, M.; Soulage, C.O.; Meynier, A.; Debard, C.; Plaisancié, P.; Bérengère, B.; Picard, G.; Loizon, E.; Chauvin, M.A.; Estienne, M.; et al. Dietary oxidized n-3 PUFA induce oxidative stress and inflammation: Role of intestinal absorption of 4-HHE and reactivity in intestinal cells. J. Lipid Res. 2012, 53, 3069–3080. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw | Boiled | Fried | Baked | |
|---|---|---|---|---|
| Enzymatic oxidized lipid products derived from AA | ||||
| 5(S)-HETE | 134.12 ± 29.53 a | 150.46 ± 33.12 a | 265.91 ± 92.79 a | 201.24 ± 7.79 a |
| 8(S)-HETE | 19.98 ± 8.02 a | 21.30 ± 2.45 a | 56.83 ± 35.47 a | 29.80 ± 4.81 a |
| 11(S)-HETE | 32.15 ± 9.16 a | 30.73 ± 7.76 a | 81.76 ± 43.66 a | 55.31 ± 27.44 a |
| 12(S)-HETE | 138.95 ± 50.97 a | 235.53 ± 198 a | 142.66 ± 86.35 a | 98.54 ± 42.05 a |
| 15(S)-HETE | 8.71 ± 4.42 a | 9.62 ± 4.32 a | 6.71 ± 2.77 a | 7.90 ± 2.69 a |
| Enzymatic oxidized lipid products derived from EPA | ||||
| RvE1 | 8.97 ± 2.94 a | 14.31 ± 2.84 a | 25.55 ± 3.86 b | 11.80 ± 3.41 a |
| Enzymatic oxidized lipid products derived from DHA | ||||
| 4-HDHA | 63.36 ± 17.08 a | 51.03 ± 10.09 a | 78.24 ± 15.16 a | 88.54 ± 17.42 a |
| 7-HDHA | 36.45 ± 6.32 a | 35.28 ± 7.88 a | 47.03 ± 11.73 a | 54.47 ± 16.39 a |
| 11-HDHA | 38.21 ± 4.84 a | 33.86 ± 8.98 a | 55.02 ± 7.74 a | 48.91 ± 13.09 a |
| 14-HDHA | 40.43 ± 3.42 a | 35.62 ± 10.38 a | 84.71 ± 22.08b | 89.26 ± 13.17 b |
| 17-HDHA | 391.58 ± 43.80 a | 539.14 ± 149.46 a | 489.99 ± 36.43 a | 790.34 ± 273.12 a |
| RvD1 | 19.46 ± 8.50 a | 19.11 ± 2.90 a | 23.82 ± 3.81 a | 37.43 ± 12.13 a |
| NPD1 | 11.41 ± 3.01 a | 15.44 ± 2.70 a | 24.68 ± 6.20 s | 11.93 ± 2.33 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leung, K.S.; Galano, J.-M.; Durand, T.; Lee, J.C.-Y. Profiling of Omega-Polyunsaturated Fatty Acids and Their Oxidized Products in Salmon after Different Cooking Methods. Antioxidants 2018, 7, 96. https://doi.org/10.3390/antiox7080096
Leung KS, Galano J-M, Durand T, Lee JC-Y. Profiling of Omega-Polyunsaturated Fatty Acids and Their Oxidized Products in Salmon after Different Cooking Methods. Antioxidants. 2018; 7(8):96. https://doi.org/10.3390/antiox7080096
Chicago/Turabian StyleLeung, Kin Sum, Jean-Marie Galano, Thierry Durand, and Jetty Chung-Yung Lee. 2018. "Profiling of Omega-Polyunsaturated Fatty Acids and Their Oxidized Products in Salmon after Different Cooking Methods" Antioxidants 7, no. 8: 96. https://doi.org/10.3390/antiox7080096
APA StyleLeung, K. S., Galano, J.-M., Durand, T., & Lee, J. C.-Y. (2018). Profiling of Omega-Polyunsaturated Fatty Acids and Their Oxidized Products in Salmon after Different Cooking Methods. Antioxidants, 7(8), 96. https://doi.org/10.3390/antiox7080096