
Astaxanthin: A Compound in the Prevention of Chronic Diseases and as a Potential Adjuvant Treatment Agent



Abstract
1. Introduction
1.1. Overview of Astaxanthin
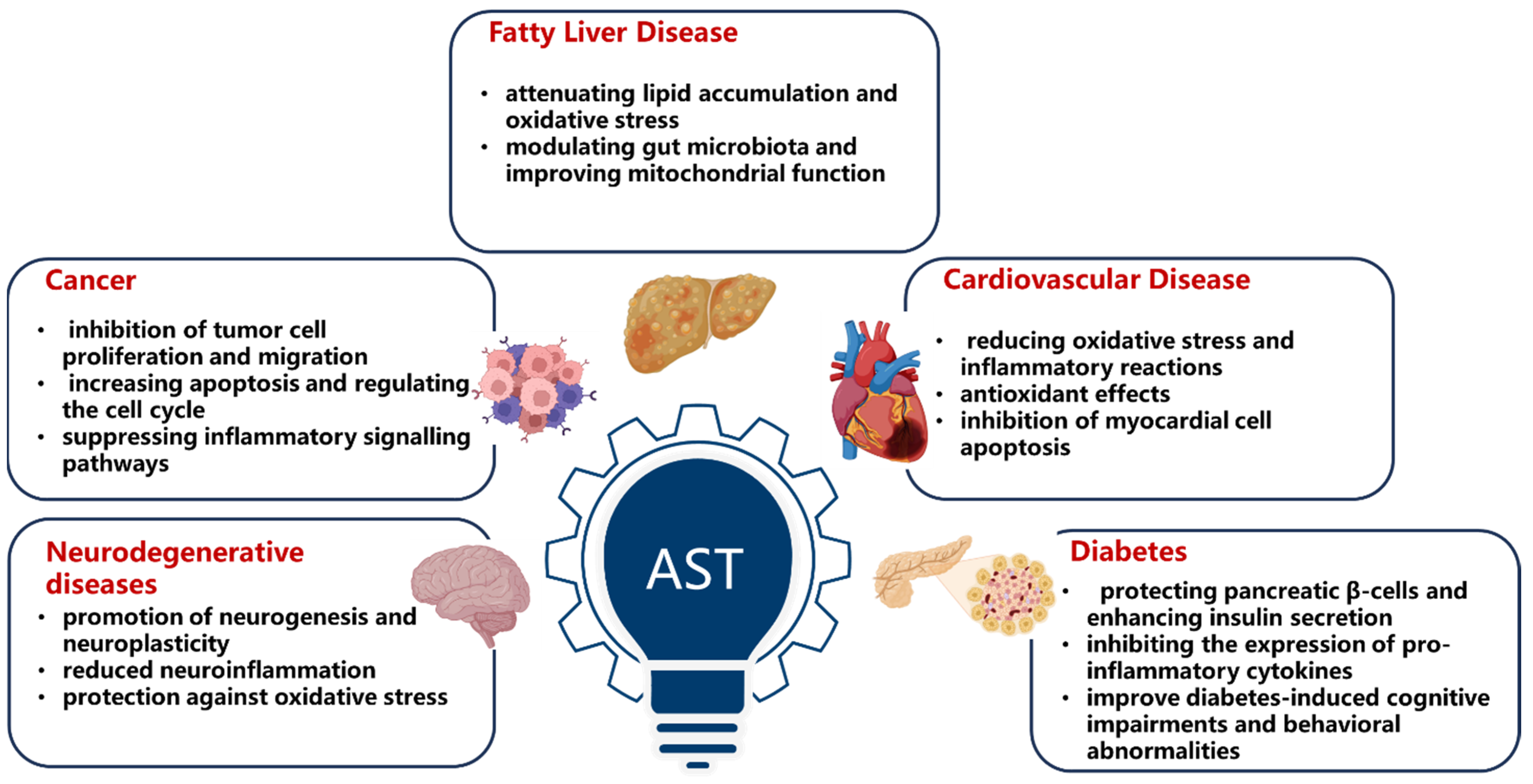
1.2. The Health Benefits of Astaxanthin
1.3. Molecular Targets of Astaxanthin
2. Antioxidant and Anti-Inflammatory Effects and Mitochondrial Protection
2.1. Antioxidant Properties
2.2. Anti-Inflammatory Properties
2.3. Mitochondrial Protection
3. Immune Regulatory Effect

{kind=link}
| Mechanism | Study Population/Model | Key Findings | Reference |
|---|---|---|---|
| Enhancement of both cellular and humoral immunity | SPF Kunming female mice | Significant improvement in delayed allergy reaction and NK cell activity | Fan et al. (2021) [26] |
| Inhibition in STING carbonylation to enhance antiviral responses | HSV-1-induced mouse primary peritoneal macrophages | Mitigating lipid peroxidation and inflammation, augments type I interferon production, restricting viral replication | Li et al. (2024) [32] |
| Downregulation of pro-inflammatory cytokines in autoimmune hepatitis | Concanavalin A-induced mouse model | Alleviating liver damage, downregulates pro-inflammatory cytokines, increases CD8+ T cells | He et al. (2024) [27] |
| Counteracting post-exercise decline in immune-related plasma proteins | Oplegnathus punctatus | Effective in counteracting the post-exercise decline in immune-related plasma proteins, particularly immunoglobulin IgM | Wu et al. (2023) [28] |
4. Anti-Apoptotic Effect and Nervous System Protection
| Mechanism | Study Population/Model | Key Findings | Reference |
|---|---|---|---|
| Improvement of brain aging | Six-month-old SAMP10 mice | Induction of autophagy by regulating IGF-1/Akt/mTOR and IGF-1/Akt/FoxO3a signaling pathways | Fu et al. (2023) [34,35] |
| Prevention of neurotoxicity | H2O2-induced SH-SY5Y cells | Inhibition H2O2-induced apoptosis in SH-SY5Y cells by ameliorating mitochondrial damage and enhancing cell survival | Yan et al. (2024) [34,35] |
| Promoting neurogenesis and neuroplasticity | Epidermal neural crest stem cells extracted from bulge hair follicle in adults | Enhances spatial memory performance, improves hippocampus-related spatial memory | Mohaghegh et al. (2020) [36] |
| Antioxidant properties | A mouse model of brain aging | Improvements in the learning, cognitive, and memory abilities of mice | Liu et al. (2021) [37] |
| Improvement of erythrocyte antioxidant status | Thirty middle-aged and senior subjects | Decreased PLOOH levels, which may contribute to the prevention of dementia | Nakagawa et al. (2011) [39] |
| Suppression of oxidative stress and apoptosis in neurons | PQ-induced SH-SY5Y cells and mice Parkinson’s disease model | Inhibition PQ-induced activation of MAPK signaling pathway | Wang et al. (2023) [44] |
| Anti-neuroapoptosis effects | Isofluorane-induced rat model | Reducing the isoflurane-induced neuroapoptosis via activation of the PI3K/Akt signaling pathway | Wang et al. (2016) [46] |
5. Anti-Tumor Effect
6. Liver Protection
7. Anti-Fibrotic Effect
8. Cardiovascular Health Improvement
| Mechanism | Study Population/Model | Key Findings | Reference |
|---|---|---|---|
| Antioxidant Properties | Human umbilical vein endothelial cell | Reducing ROS-induced lipid peroxidation and enhancing antioxidant enzyme activity | Nishigaki et al. (2010) [67] |
| U397 cell | Restoring SHP-1 expression and reducing NF-κB (p65) nuclear expression | Speranza et al. (2012) [25] | |
| U397 cell | Reducing LPS-induced toxicity and ROS production by decreasing intracellular O₂ (−) production | Franceschelli et al. (2014) [68] | |
| HepG2 cell | Enhancing cellular antioxidant capacity by inducing PON1 | Dose et al. (2016) [72] | |
| Hypercholesterolemic rabbits | Regulator of PON1 activity | Augusti et al. (2012) [73] | |
| Healthy subjects | Increasing PON1 activity | Baralic et al. (2013) [74] | |
| Rats | Increasing the expression of Nrf2 and its downstream target HO-1 | Tripathi et al. (2009) [79] | |
| Anti-inflammatory Effects | Diabetic rats | Reducing multiple inflammatory markers | Chan et al. (2012) [85] |
| Antihypertensive Effects | Spontaneously hypertensive rats | Modulating blood fluidity and improving vascular reactivity | Hussein et al. (2005) [88] |
| Metabolic syndrome rats (SHR/NDmcr-cp model) | Inducing a significant reduction in arterial blood pressure in metabolic syndrome rats | Hussein et al. (2007) [89] | |
| Male Sprague-Dawley rats | Enhancing nitric oxide, thereby inducing vasodilation of the rat aorta | Preuss et al. (2011) [90] | |
| Spontaneously hypertensive rats | Lowering the ratio of coronary artery vessel wall to lumen, reducing the increase in aortic elastin | Hussein et al. (2006) [91] | |
| Hypolipidemic Effects | Male C57BL/6J mice fed a high-fat diet | Lowering the plasma concentrations of TAG, ALT, and AST | Yang et al. (2014) [94] |
| Metabolic syndrome rats | Significantly increasing HDL-C levels and decreasing plasma TG and non-esterified fatty acid levels in a metabolic syndrome rat model | Yoshida et al. (2010) [95] | |
| APOE-knockout mice fed a high-fat, high-cholesterol diet | Increasing expression of liver cholesterol metabolism-related genes and major β-oxidation-related enzymes | Yang et al. (2011) [96] | |
| Antithrombotic Effects | Spontaneously hypertensive rats | Reduction in systolic blood pressure, a delay in cerebral vascular thrombosis, an increase in NO metabolic rate, a significant elevation in urinary NO₂/NO₃ levels, and a marked decrease in 8-OHdG levels | Sasaki et al. (2011) [97] |
| C57BL/6 mice | Substantially enhancing NO levels, reducing ONOO− levels, promoting vasodilation in the aortic and coronary arteries, augmenting blood flow, decreasing blood viscosity, and protecting LDL from oxidation | Khan et al. (2010) [98] | |
| Cardioprotective Effects | Healthy subjects | Reducing the oxidation of fatty acids and LDL | Fassett et al. (2012) [4] |
| Overweight and obese adults | Decreasing lipid peroxidation markers in the blood significantly and increasing plasma superoxide dismutase levels and total antioxidant levels | Choi et al. (2011) [99] | |
| Young, healthy women | Lower levels of C-reactive protein (an inflammatory marker used to determine CVD risk), improving immune cell responses and cytokine status | Park et al. (2010) [100] | |
| Renal transplant patients | Having no significant effect on plasma C-reactive protein levels | Coombes et al. (2016) [101] |
9. Anti-Diabetes Effect
10. Strategies for Improving Astaxanthin Bioavailability
| Strategy | Specific Measures | Results | Reference |
|---|---|---|---|
| Lipid-based Carriers | Co-intake of AST with dietary fat | AUC of the serum concentration of AST taken after meals was significantly higher than that taken on an empty stomach | Okada et al. (2009) [107] |
| Using lipid carriers like microemulsions and liposomes | AST liposomes coated with soybean lecithin significantly improved their biological accessibility in simulated digestion | Chang et al. (2022) [108] | |
| Three lipid-based formulations of AST all showed enhanced bioavailability, ranging from 1.7 to 3.7 times that of the reference formulation | Mercke et al. (2003) [109] | ||
| Nano Delivery and SR System | Using nanoemulsions, solid lipid nanoparticles, chitosan or PLGA nanoparticles | AST was encapsulated in PLGA nanoparticles coated with chitosan oligosaccharides, with good dispersibility and stability in aqueous solutions, as well as high cytocompatibility | Liu et al. (2019) [110] |
| PLGA nanoparticles can achieve sustained release and improve bioavailability of AST | Xue et al. (2023) [112] | ||
| SR formulation | The bioavailability of AST with an SR matrix formulation is 3.6 times higher than that of AST oil without formulation | Madhavi et al. (2018) [113] | |
| Microencapsulation | Microencapsulation technology like spray drying (e.g., whey protein microcapsules, soybean lecithin microcapsules) | WP encapsulation can effectively improve the stability, water solubility, and bioavailability of AST esters | Yang et al. (2022) [114] |
| AST was microencapsulated with soluble polymers using spray drying to improve its solubility and bioavailability | Nalawade et al. (2015) [115] | ||
| Isomer Optimization | Studying molecular structures (optical stereoisomers and geometric isomers) | 13Z-AST showed higher bioaccessibility than 9Z- and all-E-ASTs during in vitro digestion, and 9Z-AST exhibited higher transport efficiency than all-E- and 13Z-ASTs | Yang et al. (2016) [116] |
| Using Haematococcus pluvialis-derived AST (all-E-3S-3′ S form) | A selective process increases the relative proportion of AST Z-isomers compared to the all-E-AST before uptake in blood and that the AST esters are hydrolyzed selectively during absorption | Coral-Hinostroza et al. (2004) [122] | |
| A selective process increases the relative proportion of AST Z-isomers compared to the all-E-AST during blood uptake and that AST E/Z isomers have similar pharmacokinetics | Østerlie et al. (2000) [123] | ||
| Esterified Form | Using esterified AST | AST ester (such as diethyl ASTA de, ASTA es) is more stable and has higher bioavailability than free AST | Yang et al. (2020) [117] |
| AST polyethylene glycol succinate (APGS) showed better solubility with enhanced bioavailability and stability compared to free AST | Sakayanathan et al. (2024) [102] | ||
| PEG−carotenoid ester conjugates also showed good antioxidant activity due to their water dispersibility | Háda et al. (2011) [118] | ||
| Other Formulations and Advances | Synthetic AST | Cardax, a novel synthetic AST disodium disuccinate derivative, exhibits higher water solubility and bioavailability than free AST | Lockwood et al. (2005) [119] |
| Micellar solubilization technology | In healthy men, the Novasol capsule, based on micellar solubilization technology, showed higher plasma concentration and better pharmacokinetic parameters | Khayyal et al. (2024) [120] | |
| Intestine-targeted delivery systems | A gut-responsive carrier remarkably increased muco-adhesion, deep mucus layer penetration, and AST absorption and bioavailability | Liu et al. (2024) [121] |
11. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| AFLD | alcoholic fatty liver disease |
| ALD | alcoholic liver disease |
| AMPK | AMP-activated protein kinase |
| APGS | astaxanthin polyethylene glycol succinate |
| AST | astaxanthin |
| AUC | area under the curve |
| bw | body weight |
| COPD | chronic obstructive pulmonary disease |
| COX | cyclooxygenase |
| CVD | cardiovascular diseases |
| EMT | epithelial–mesenchymal transition |
| FGF21 | fibroblast growth factor 21 |
| GPX | glutathione peroxidase |
| HDL | high-density lipoprotein cholesterol |
| HUVEC | human umbilical vein endothelial cell |
| IL-6 | interleukin-6 |
| iNOS | inducible nitric oxide synthase |
| LDL | low-density lipoprotein cholesterol |
| LPS | lipopolysaccharides |
| MAPK | mitogen-activated protein kinase |
| MASLD | metabolic dysfunction-associated steatotic liver disease |
| MCP-1 | monocyte chemoattractant protein-1 |
| mPTP | mitochondrial permeability transition pore |
| NO | nitric oxide |
| PD | Parkinson’s disease |
| PEG | polyethylene glycol |
| PGC-1α | peroxisome proliferator-activated receptor gamma coactivator 1α |
| PLGA | poly lactic-co-glycolic acid |
| PON1 | paraoxonase 1 |
| PPAR | peroxisome proliferator-activated receptors |
| ROS | reactive oxygen species |
| RNS | reactive nitrogen species |
| SOD | superoxide dismutase |
| SHR | spontaneously hypertensive rat |
| STING | Stimulator of Interferon Genes |
| TC | total cholesterol |
| TNF-α | tumor necrosis factor-alpha |
| VE-cadherin | vascular endothelial cadherin |
References
- Shah, M.M.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-Producing Green Microalga Haematococcus pluvialis: From Single Cell to High Value Commercial Products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.P.; Peng, J.; Yin, K.; Wang, J.H. Potential health-promoting effects of astaxanthin: A high-value carotenoid mostly from microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Ambati, R.R.; Phang, S.M.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications--a review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef]
- Fassett, R.G.; Coombes, J.S. Astaxanthin in cardiovascular health and disease. Molecules 2012, 17, 2030–2048. [Google Scholar] [CrossRef]
- Hussein, G.; Sankawa, U.; Goto, H.; Matsumoto, K.; Watanabe, H. Astaxanthin, a carotenoid with potential in human health and nutrition. J. Nat. Prod. 2006, 69, 443–449. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed]
- Yucetepe, M.; Tugba Ozaslan, Z.; Karakus, M.S.; Akalan, M.; Karaaslan, A.; Karaaslan, M.; Basyigit, B. Unveiling the multifaceted world of anthocyanins: Biosynthesis pathway, natural sources, extraction methods, copigmentation, encapsulation techniques, and future food applications. Food Res. Int. 2024, 187, 114437. [Google Scholar] [CrossRef]
- Vitale, M.; Gomez-Estaca, J.; Chung, J.; Chua, S.C.; Pampanin, D.M. Encapsulation Techniques to Enhance Astaxanthin Utilization as Functional Feed Ingredient. Mar. Drugs 2025, 23, 143. [Google Scholar] [CrossRef]
- Alugoju, P.; Krishna Swamy, V.K.D.; Anthikapalli, N.V.A.; Tencomnao, T. Health benefits of astaxanthin against age-related diseases of multiple organs: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2023, 63, 10709–10774. [Google Scholar] [CrossRef]
- Yin, Y.; Xu, N.; Shi, Y.; Zhou, B.; Sun, D.; Ma, B.; Xu, Z.; Yang, J.; Li, C. Astaxanthin Protects Dendritic Cells from Lipopolysaccharide-Induced Immune Dysfunction. Mar. Drugs 2021, 19, 346. [Google Scholar] [CrossRef]
- Wu, Y.; Bashir, M.A.; Shao, C.; Wang, H.; Zhu, J.; Huang, Q. Astaxanthin targets IL-6 and alleviates the LPS-induced adverse inflammatory response of macrophages. Food Funct. 2024, 15, 4207–4222. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Chu, A.; Luo, Q.; Wu, M.; Shi, X.; Chen, Y. The Protective Effect of Astaxanthin on Cognitive Function via Inhibition of Oxidative Stress and Inflammation in the Brains of Chronic T2DM Rats. Front. Pharmacol. 2018, 9, 748. [Google Scholar] [CrossRef]
- Faraone, I.; Sinisgalli, C.; Ostuni, A.; Armentano, M.F.; Carmosino, M.; Milella, L.; Russo, D.; Labanca, F.; Khan, H. Astaxanthin anticancer effects are mediated through multiple molecular mechanisms: A systematic review. Pharmacol. Res. 2020, 155, 104689. [Google Scholar] [CrossRef]
- Inoue, M.; Tanabe, H.; Matsumoto, A.; Takagi, M.; Umegaki, K.; Amagaya, S.; Takahashi, J. Astaxanthin functions differently as a selective peroxisome proliferator-activated receptor gamma modulator in adipocytes and macrophages. Biochem. Pharmacol. 2012, 84, 692–700. [Google Scholar] [CrossRef] [PubMed]
- Lewis Luján, L.M.; McCarty, M.F.; Di Nicolantonio, J.J.; Gálvez Ruiz, J.C.; Rosas-Burgos, E.C.; Plascencia-Jatomea, M.; Iloki Assanga, S.B. Nutraceuticals/Drugs Promoting Mitophagy and Mitochondrial Biogenesis May Combat the Mitochondrial Dysfunction Driving Progression of Dry Age-Related Macular Degeneration. Nutrients 2022, 14, 1985. [Google Scholar] [CrossRef]
- Baburina, Y.; Krestinin, R.; Odinokova, I.; Sotnikova, L.; Kruglov, A.; Krestinina, O. Astaxanthin Inhibits Mitochondrial Permeability Transition Pore Opening in Rat Heart Mitochondria. Antioxidants 2019, 8, 576. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Z.; Peng, P.; Gong, Z.; Huang, J.; Peng, H. Astaxanthin attenuates osteoarthritis progression via inhibiting ferroptosis and regulating mitochondrial function in chondrocytes. Chem. Biol. Interact. 2022, 366, 110148. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, Y.; Chen, Q.; Yang, H.; Xie, X. Astaxanthin Promotes Nrf2/ARE Signaling to Alleviate Renal Fibronectin and Collagen IV Accumulation in Diabetic Rats. J. Diabetes Res. 2018, 2018, 6730315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, X.; Hua, Z.; Xing, S.; Li, J.; Fei, S.; Tan, M. ROS-triggered self-disintegrating and pH-responsive astaxanthin nanoparticles for regulating the intestinal barrier and colitis. Biomaterials 2023, 292, 121937. [Google Scholar] [CrossRef]
- Zhang, Q.; Luo, C.; Li, Z.; Huang, W.; Zheng, S.; Liu, C.; Shi, X.; Ma, Y.; Ni, Q.; Tan, W.; et al. Astaxanthin activates the Nrf2/Keap1/HO-1 pathway to inhibit oxidative stress and ferroptosis, reducing triphenyl phosphate (TPhP)-induced neurodevelopmental toxicity. Ecotoxicol. Environ. Saf. 2024, 271, 115960. [Google Scholar] [CrossRef]
- Nishida, Y.; Berg, P.C.; Shakersain, B.; Hecht, K.; Takikawa, A.; Tao, R.; Kakuta, Y.; Uragami, C.; Hashimoto, H.; Misawa, N.; et al. Astaxanthin: Past, Present, and Future. Mar. Drugs 2023, 21, 514. [Google Scholar] [CrossRef]
- Mei, S.; Song, X.; Wang, Y.; Wang, J.; Su, S.; Zhu, J.; Geng, Y. Studies on Protection of Astaxanthin from Oxidative Damage Induced by H(2)O(2) in RAW 264.7 Cells Based on (1)H NMR Metabolomics. J. Agric. Food Chem. 2019, 67, 13568–13576. [Google Scholar] [CrossRef]
- Baburina, Y.; Krestinin, R.; Fedorov, D.; Odinokova, I.; Pershina, E.; Sotnikova, L.; Krestinina, O. The Improvement of Functional State of Brain Mitochondria with Astaxanthin in Rats after Heart Failure. Int. J. Mol. Sci. 2022, 24, 31. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.M.; Kim, Y.J. Astaxanthin Treatment Induces Maturation and Functional Change of Myeloid-Derived Suppressor Cells in Tumor-Bearing Mice. Antioxidants 2020, 9, 350. [Google Scholar] [CrossRef]
- Speranza, L.; Pesce, M.; Patruno, A.; Franceschelli, S.; Lutiis, M.A.; Grilli, A.; Felaco, M. Astaxanthin treatment reduced oxidative induced pro-inflammatory cytokines secretion in U937: SHP-1 as a novel biological target. Mar. Drugs 2012, 10, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Chen, Z.; Wu, Y.; Zhu, J.; Yu, Z. Study on the Enhancement of Immune Function of Astaxanthin from Haematococcus pluvialis. Foods 2021, 10, 1847. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Ding, M.; Zhang, J.; Huang, C.; Shi, J.; Wang, Y.; Tao, R.; Wu, Z.; Guo, W. Astaxanthin Alleviates Autoimmune Hepatitis by Modulating CD8(+) T Cells: Insights From Mass Cytometry and Single-Cell RNA Sequencing Analyses. Adv. Sci. 2024, 11, e2403148. [Google Scholar] [CrossRef]
- Wu, Y.; Xiao, Y.; Li, W.; Yang, C.; Ma, W.; Pang, Z.; Zhang, J.; Xiao, Z.; Li, J. Tea polyphenols, astaxanthin, and melittin can significantly enhance the immune response of juvenile spotted knifejaw (Oplegnathus punctatus). Fish. Shellfish. Immunol. 2023, 138, 108817. [Google Scholar] [CrossRef]
- Meng, X.; Yang, F.; Zhu, L.; Zhan, L.; Numasawa, T.; Deng, J. Effects of dietary astaxanthin supplementation on growth performance, antioxidant status, immune response, and intestinal health of rainbow trout (Oncorhynchus mykiss). Anim. Nutr. 2024, 17, 387–396. [Google Scholar] [CrossRef]
- Aziz, M.M.; El-Sheikh, M.M.; Mohamed, M.A.; Abdelrahman, S.S.; Mekkawy, M.H. The senomorphic impact of astaxanthin on irradiated rat spleen: STING, TLR4 and mTOR contributed pathway. Int. J. Immunopathol. Pharmacol. 2024, 38, 3946320241297342. [Google Scholar] [CrossRef]
- Chen, L.; Wang, L.; Li, Y.; Wu, X.; Long, X. Comprehensive metabolomics and transcriptomics analyses investigating the regulatory effects of different sources of dietary astaxanthin on the antioxidant and immune functions of commercial-sized rainbow trout. Front. Immunol. 2024, 15, 1408168. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Jia, M.; Song, H.; Peng, J.; Zhao, W.; Zhang, W. Astaxanthin Inhibits STING Carbonylation and Enhances Antiviral Responses. J. Immunol. 2024, 212, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wen, C.; Yang, M.; Li, A.; Fan, C.; Gan, D.; Li, Q.; Zhao, J.; Zhu, L.; Lu, D. Astaxanthin Improved the Cognitive Deficits in APP/PS1 Transgenic Mice Via Selective Activation of mTOR. J. Neuroimmune Pharmacol. 2021, 16, 609–619. [Google Scholar] [CrossRef]
- Fu, M.; Liang, X.; Zhang, X.; Yang, M.; Ye, Q.; Qi, Y.; Liu, H.; Zhang, X. Astaxanthin delays brain aging in senescence-accelerated mouse prone 10: Inducing autophagy as a potential mechanism. Nutr. Neurosci. 2023, 26, 445–455. [Google Scholar] [CrossRef]
- Yan, T.; Ding, F.; Zhang, Y.; Wang, Y.; Wang, Y.; Zhang, Y.; Zhu, F.; Zhang, G.; Zheng, X.; Jia, G.; et al. Astaxanthin Inhibits H(2)O(2)-Induced Excessive Mitophagy and Apoptosis in SH-SY5Y Cells by Regulation of Akt/mTOR Activation. Mar. Drugs 2024, 22, 57. [Google Scholar] [CrossRef]
- Mohaghegh Shalmani, L.; Valian, N.; Pournajaf, S.; Abbaszadeh, F.; Dargahi, L.; Jorjani, M. Combination therapy with astaxanthin and epidermal neural crest stem cells improves motor impairments and activates mitochondrial biogenesis in a rat model of spinal cord injury. Mitochondrion 2020, 52, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Zeng, L.; Zhang, Y.M.; Pan, W.; Lai, H. Astaxanthin alleviates pathological brain aging through the upregulation of hippocampal synaptic proteins. Neural Regen. Res. 2021, 16, 1062–1067. [Google Scholar] [CrossRef]
- Chik, M.W.; Meor Mohd Affandi, M.M.R.; Mohd Nor Hazalin, N.A.; Surindar Singh, G.K. Astaxanthin nanoemulsion improves cognitive function and synaptic integrity in Streptozotocin-induced Alzheimer’s disease model. Metab. Brain Dis. 2025, 40, 136. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Kiko, T.; Miyazawa, T.; Carpentero Burdeos, G.; Kimura, F.; Satoh, A.; Miyazawa, T. Antioxidant effect of astaxanthin on phospholipid peroxidation in human erythrocytes. Br. J. Nutr. 2011, 105, 1563–1571. [Google Scholar] [CrossRef]
- Alghazwi, M.; Smid, S.; Musgrave, I.; Zhang, W. In vitro studies of the neuroprotective activities of astaxanthin and fucoxanthin against amyloid beta (Aβ(1-42)) toxicity and aggregation. Neurochem. Int. 2019, 124, 215–224. [Google Scholar] [CrossRef]
- Wang, S.; Qi, X. The Putative Role of Astaxanthin in Neuroinflammation Modulation: Mechanisms and Therapeutic Potential. Front. Pharmacol. 2022, 13, 916653. [Google Scholar] [CrossRef]
- Zhang, X.S.; Zhang, X.; Wu, Q.; Li, W.; Wang, C.X.; Xie, G.B.; Zhou, X.M.; Shi, J.X.; Zhou, M.L. Astaxanthin offers neuroprotection and reduces neuroinflammation in experimental subarachnoid hemorrhage. J. Surg. Res. 2014, 192, 206–213. [Google Scholar] [CrossRef]
- Menozzi, E.; Schapira, A.H.V. Prospects for Disease Slowing in Parkinson Disease. Annu. Rev. Pharmacol. Toxicol. 2025, 65, 237–258. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lu, K.; Lou, X.; Zhang, S.; Song, W.; Li, R.; Geng, L.; Cheng, B. Astaxanthin ameliorates dopaminergic neuron damage in paraquat-induced SH-SY5Y cells and mouse models of Parkinson’s disease. Brain Res. Bull. 2023, 202, 110762. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.D.; Khanal, S.; Lee, E.; Choi, J.; Bohara, G.; Rimal, N.; Choi, D.Y.; Park, S. Astaxanthin-loaded brain-permeable liposomes for Parkinson’s disease treatment via antioxidant and anti-inflammatory responses. J. Nanobiotechnology 2025, 23, 78. [Google Scholar] [CrossRef]
- Wang, C.M.; Cai, X.L.; Wen, Q.P. Astaxanthin reduces isoflurane-induced neuroapoptosis via the PI3K/Akt pathway. Mol. Med. Rep. 2016, 13, 4073–4078. [Google Scholar] [CrossRef] [PubMed]
- Jahan, I.; Harun-Ur-Rashid, M.; Islam, M.A.; Sharmin, F.; Al Jaouni, S.K.; Kaki, A.M.; Selim, S. Neuronal plasticity and its role in Alzheimer’s disease and Parkinson’s disease. Neural Regen. Res. 2024, 21, 107–125. [Google Scholar] [CrossRef]
- Li, M.Y.; Shi, Y.C.; Xu, W.X.; Zhao, L.; Zhang, A.Z. Exploring Cr(VI)-induced blood-brain barrier injury and neurotoxicity in zebrafish and snakehead fish, and inhibiting toxic effects of astaxanthin. Environ. Pollut. 2024, 355, 124280. [Google Scholar] [CrossRef]
- Wang, D.N.; Yu, C.X.; Feng, J.; Wei, L.J.; Chen, J.; Liu, Z.; Ouyang, L.; Zhang, L.; Liu, F.; Hua, Q. Comparative transcriptome analysis reveals the redirection of metabolic flux from cell growth to astaxanthin biosynthesis in Yarrowia lipolytica. Yeast 2024, 41, 369–378. [Google Scholar] [CrossRef]
- Copat, C.; Favara, C.; Tomasello, M.F.; Sica, C.; Grasso, A.; Dominguez, H.G.; Conti, G.O.; Ferrante, M. Astaxanthin in cancer therapy and prevention (Review). Biomed. Rep. 2025, 22, 66. [Google Scholar] [CrossRef]
- Song, X.; Wang, M.; Zhang, L.; Zhang, J.; Wang, X.; Liu, W.; Gu, X.; Lv, C. Changes in cell ultrastructure and inhibition of JAK1/STAT3 signaling pathway in CBRH-7919 cells with astaxanthin. Toxicol. Mech. Methods 2012, 22, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Kowshik, J.; Baba, A.B.; Giri, H.; Deepak Reddy, G.; Dixit, M.; Nagini, S. Astaxanthin inhibits JAK/STAT-3 signaling to abrogate cell proliferation, invasion and angiogenesis in a hamster model of oral cancer. PLoS ONE 2014, 9, e109114. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.S.; Daglia, M.; Rengasamy, K.R. Dietary carotenoids in cancer chemoprevention and chemotherapy: A review of emerging evidence. Pharmacol. Res. 2020, 157, 104830. [Google Scholar] [CrossRef]
- Sun, S.Q.; Zhao, Y.X.; Li, S.Y.; Qiang, J.W.; Ji, Y.Z. Anti-Tumor Effects of Astaxanthin by Inhibition of the Expression of STAT3 in Prostate Cancer. Mar. Drugs 2020, 18, 415. [Google Scholar] [CrossRef]
- Ni, X.; Yu, H.; Wang, S.; Zhang, C.; Shen, S. Astaxanthin Inhibits PC-3 Xenograft Prostate Tumor Growth in Nude Mice. Mar. Drugs 2017, 15, 66. [Google Scholar] [CrossRef]
- Maoka, T.; Tokuda, H.; Suzuki, N.; Kato, H.; Etoh, H. Anti-oxidative, anti-tumor-promoting, and anti-carcinogensis activities of nitroastaxanthin and nitrolutein, the reaction products of astaxanthin and lutein with peroxynitrite. Mar. Drugs 2012, 10, 1391–1399. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, M.; Zhou, Y.; Bao, S.; Wang, F.; Li, C. Protective Effects of Astaxanthin against Oxidative Stress: Attenuation of TNF-α-Induced Oxidative Damage in SW480 Cells and Azoxymethane/Dextran Sulfate Sodium-Induced Colitis-Associated Cancer in C57BL/6 Mice. Mar. Drugs 2024, 22, 469. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Jiang, C. Astaxanthin suppresses the malignant behaviors of nasopharyngeal carcinoma cells by blocking PI3K/AKT and NF-κB pathways via miR-29a-3p. Genes. Environ. 2024, 46, 10. [Google Scholar] [CrossRef]
- Cui, L.; Li, Z.; Xu, F.; Tian, Y.; Chen, T.; Li, J.; Guo, Y.; Lyu, Q. Antitumor Effects of Astaxanthin on Esophageal Squamous Cell Carcinoma by up-Regulation of PPARγ. Nutr. Cancer 2022, 74, 1399–1410. [Google Scholar] [CrossRef]
- Tsuji, S.; Nakamura, S.; Maoka, T.; Yamada, T.; Imai, T.; Ohba, T.; Yako, T.; Hayashi, M.; Endo, K.; Saio, M.; et al. Antitumour Effects of Astaxanthin and Adonixanthin on Glioblastoma. Mar. Drugs 2020, 18, 474. [Google Scholar] [CrossRef]
- Wu, L.; Mo, W.; Feng, J.; Li, J.; Yu, Q.; Li, S.; Zhang, J.; Chen, K.; Ji, J.; Dai, W.; et al. Astaxanthin attenuates hepatic damage and mitochondrial dysfunction in non-alcoholic fatty liver disease by up-regulating the FGF21/PGC-1α pathway. Br. J. Pharmacol. 2020, 177, 3760–3777. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, M.; Fu, X.; Zhang, Z.; Zhu, L.; Zheng, X.; Liu, J. Astaxanthin Prevents Alcoholic Fatty Liver Disease by Modulating Mouse Gut Microbiota. Nutrients 2018, 10, 1298. [Google Scholar] [CrossRef]
- Krestinina, O.; Odinokova, I.; Sotnikova, L.; Krestinin, R.; Zvyagina, A.; Baburina, Y. Astaxanthin Is Able to Prevent Alcohol-Induced Dysfunction of Liver Mitochondria. Antioxidants 2022, 11, 2019. [Google Scholar] [CrossRef]
- Diao, W.; Chen, W.; Cao, W.; Yuan, H.; Ji, H.; Wang, T.; Chen, W.; Zhu, X.; Zhou, H.; Guo, H.; et al. Astaxanthin protects against renal fibrosis through inhibiting myofibroblast activation and promoting CD8(+) T cell recruitment. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 1360–1370. [Google Scholar] [CrossRef]
- Zhao, J.; Meng, M.; Zhang, J.; Li, L.; Zhu, X.; Zhang, L.; Wang, C.; Gao, M. Astaxanthin ameliorates renal interstitial fibrosis and peritubular capillary rarefaction in unilateral ureteral obstruction. Mol. Med. Rep. 2019, 19, 3168–3178. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, J.; Li, R.; Lv, C.; Xu, P.; Wang, Y.; Song, X.; Zhang, J. Astaxanthin attenuates pulmonary fibrosis through lncITPF and mitochondria-mediated signal pathways. J. Cell Mol. Med. 2020, 24, 10245–10250. [Google Scholar] [CrossRef] [PubMed]
- Nishigaki, I.; Rajendran, P.; Venugopal, R.; Ekambaram, G.; Sakthisekaran, D.; Nishigaki, Y. Cytoprotective role of astaxanthin against glycated protein/iron chelate-induced toxicity in human umbilical vein endothelial cells. Phytother. Res. 2010, 24, 54–59. [Google Scholar] [CrossRef]
- Franceschelli, S.; Pesce, M.; Ferrone, A.; De Lutiis, M.A.; Patruno, A.; Grilli, A.; Felaco, M.; Speranza, L. Astaxanthin treatment confers protection against oxidative stress in U937 cells stimulated with lipopolysaccharide reducing O2− production. PLoS ONE 2014, 9, e88359. [Google Scholar] [CrossRef]
- Pereira, C.P.M.; Souza, A.C.R.; Vasconcelos, A.R.; Prado, P.S.; Name, J.J. Antioxidant and anti-inflammatory mechanisms of action of astaxanthin in cardiovascular diseases (Review). Int. J. Mol. Med. 2021, 47, 37–48. [Google Scholar] [CrossRef]
- van Himbergen, T.M.; van der Schouw, Y.T.; Voorbij, H.A.; van Tits, L.J.; Stalenhoef, A.F.; Peeters, P.H.; Roest, M. Paraoxonase (PON1) and the risk for coronary heart disease and myocardial infarction in a general population of Dutch women. Atherosclerosis 2008, 199, 408–414. [Google Scholar] [CrossRef]
- Kresanov, P.; Vasankari, T.; Ahotupa, M.; Kaikkonen, J.; Hutri-Kähönen, N.; Juonala, M.; Kähönen, M.; Lehtimäki, T.; Viikari, J.; Raitakari, O.T. Paraoxonase-1 and oxidized lipoprotein lipids. The Cardiovascular Risk in Young Finns Study. Atherosclerosis 2015, 241, 502–506. [Google Scholar] [CrossRef] [PubMed]
- Dose, J.; Matsugo, S.; Yokokawa, H.; Koshida, Y.; Okazaki, S.; Seidel, U.; Eggersdorfer, M.; Rimbach, G.; Esatbeyoglu, T. Free Radical Scavenging and Cellular Antioxidant Properties of Astaxanthin. Int. J. Mol. Sci. 2016, 17, 103. [Google Scholar] [CrossRef] [PubMed]
- Augusti, P.R.; Quatrin, A.; Somacal, S.; Conterato, G.M.; Sobieski, R.; Ruviaro, A.R.; Maurer, L.H.; Duarte, M.M.; Roehrs, M.; Emanuelli, T. Astaxanthin prevents changes in the activities of thioredoxin reductase and paraoxonase in hypercholesterolemic rabbits. J. Clin. Biochem. Nutr. 2012, 51, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Baralic, I.; Djordjevic, B.; Dikic, N.; Kotur-Stevuljevic, J.; Spasic, S.; Jelic-Ivanovic, Z.; Radivojevic, N.; Andjelkovic, M.; Pejic, S. Effect of astaxanthin supplementation on paraoxonase 1 activities and oxidative stress status in young soccer players. Phytother. Res. 2013, 27, 1536–1542. [Google Scholar] [CrossRef]
- Choi, B.H.; Kang, K.S.; Kwak, M.K. Effect of redox modulating NRF2 activators on chronic kidney disease. Molecules 2014, 19, 12727–12759. [Google Scholar] [CrossRef]
- O’Connell, M.A.; Hayes, J.D. The Keap1/Nrf2 pathway in health and disease: From the bench to the clinic. Biochem. Soc. Trans. 2015, 43, 687–689. [Google Scholar] [CrossRef]
- Saw, C.L.; Yang, A.Y.; Guo, Y.; Kong, A.N. Astaxanthin and omega-3 fatty acids individually and in combination protect against oxidative stress via the Nrf2-ARE pathway. Food Chem. Toxicol. 2013, 62, 869–875. [Google Scholar] [CrossRef]
- Li, Z.; Dong, X.; Liu, H.; Chen, X.; Shi, H.; Fan, Y.; Hou, D.; Zhang, X. Astaxanthin protects ARPE-19 cells from oxidative stress via upregulation of Nrf2-regulated phase II enzymes through activation of PI3K/Akt. Mol. Vis. 2013, 19, 1656–1666. [Google Scholar]
- Tripathi, D.N.; Jena, G.B. Astaxanthin intervention ameliorates cyclophosphamide-induced oxidative stress, DNA damage and early hepatocarcinogenesis in rat: Role of Nrf2, p53, p38 and phase-II enzymes. Mutat. Res. 2010, 696, 69–80. [Google Scholar] [CrossRef]
- Gómez-Guerrero, C.; Mallavia, B.; Egido, J. Targeting inflammation in cardiovascular diseases. still a neglected field? Cardiovasc. Ther. 2012, 30, e189–e197. [Google Scholar] [CrossRef]
- Bäck, M.; Weber, C.; Lutgens, E. Regulation of atherosclerotic plaque inflammation. J. Intern. Med. 2015, 278, 462–482. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.D.; Cahú, T.B.; Firmino, G.O.; de Castro, C.C.; Carvalho, L.B., Jr.; Bezerra, R.S.; Filho, J.L. Shrimp waste extract and astaxanthin: Rat alveolar macrophage, oxidative stress and inflammation. J. Food Sci. 2012, 77, H141–H146. [Google Scholar] [CrossRef]
- Abdelzaher, L.A.; Imaizumi, T.; Suzuki, T.; Tomita, K.; Takashina, M.; Hattori, Y. Astaxanthin alleviates oxidative stress insults-related derangements in human vascular endothelial cells exposed to glucose fluctuations. Life Sci. 2016, 150, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Bhuvaneswari, S.; Yogalakshmi, B.; Sreeja, S.; Anuradha, C.V. Astaxanthin reduces hepatic endoplasmic reticulum stress and nuclear factor-κB-mediated inflammation in high fructose and high fat diet-fed mice. Cell Stress. Chaperones 2014, 19, 183–191. [Google Scholar] [CrossRef]
- Chan, K.C.; Pen, P.J.; Yin, M.C. Anticoagulatory and antiinflammatory effects of astaxanthin in diabetic rats. J. Food Sci. 2012, 77, H76–H80. [Google Scholar] [CrossRef]
- Takahashi, K.; Takimoto, T.; Sato, K.; Akiba, Y. Effect of dietary supplementation of astaxanthin from Phaffia rhodozyma on lipopolysaccharide-induced early inflammatory responses in male broiler chickens (Gallus gallus) fed a corn-enriched diet. Anim. Sci. J. 2011, 82, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Chew, B.P.; Mathison, B.D.; Hayek, M.G.; Massimino, S.; Reinhart, G.A.; Park, J.S. Dietary astaxanthin enhances immune response in dogs. Vet. Immunol. Immunopathol. 2011, 140, 199–206. [Google Scholar] [CrossRef]
- Hussein, G.; Goto, H.; Oda, S.; Iguchi, T.; Sankawa, U.; Matsumoto, K.; Watanabe, H. Antihypertensive potential and mechanism of action of astaxanthin: II. Vascular reactivity and hemorheology in spontaneously hypertensive rats. Biol. Pharm. Bull. 2005, 28, 967–971. [Google Scholar] [CrossRef]
- Hussein, G.; Nakagawa, T.; Goto, H.; Shimada, Y.; Matsumoto, K.; Sankawa, U.; Watanabe, H. Astaxanthin ameliorates features of metabolic syndrome in SHR/NDmcr-cp. Life Sci. 2007, 80, 522–529. [Google Scholar] [CrossRef]
- Preuss, H.G.; Echard, B.; Yamashita, E.; Perricone, N.V. High dose astaxanthin lowers blood pressure and increases insulin sensitivity in rats: Are these effects interdependent? Int. J. Med. Sci. 2011, 8, 126–138. [Google Scholar] [CrossRef]
- Hussein, G.; Goto, H.; Oda, S.; Sankawa, U.; Matsumoto, K.; Watanabe, H. Antihypertensive potential and mechanism of action of astaxanthin: III. Antioxidant and histopathological effects in spontaneously hypertensive rats. Biol. Pharm. Bull. 2006, 29, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.C.; Huang, S.C.; Chang, W.T.; Chen, S.C.; Hsu, C.L. Effect of Astaxanthin on the Inhibition of Lipid Accumulation in 3T3-L1 Adipocytes via Modulation of Lipogenesis and Fatty Acid Transport Pathways. Molecules 2020, 25, 3598. [Google Scholar] [CrossRef] [PubMed]
- Galema-Boers, J.M.; van Lennep, J.E. Dyslipidemia testing: Why, for whom and when. Maturitas 2015, 81, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Pham, T.X.; Wegner, C.J.; Kim, B.; Ku, C.S.; Park, Y.K.; Lee, J.Y. Astaxanthin lowers plasma TAG concentrations and increases hepatic antioxidant gene expression in diet-induced obesity mice. Br. J. Nutr. 2014, 112, 1797–1804. [Google Scholar] [CrossRef]
- Yoshida, H.; Yanai, H.; Ito, K.; Tomono, Y.; Koikeda, T.; Tsukahara, H.; Tada, N. Administration of natural astaxanthin increases serum HDL-cholesterol and adiponectin in subjects with mild hyperlipidemia. Atherosclerosis 2010, 209, 520–523. [Google Scholar] [CrossRef]
- Yang, Y.; Seo, J.M.; Nguyen, A.; Pham, T.X.; Park, H.J.; Park, Y.; Kim, B.; Bruno, R.S.; Lee, J. Astaxanthin-rich extract from the green alga Haematococcus pluvialis lowers plasma lipid concentrations and enhances antioxidant defense in apolipoprotein E knockout mice. J. Nutr. 2011, 141, 1611–1617. [Google Scholar] [CrossRef]
- Sasaki, Y.; Kobara, N.; Higashino, S.; Giddings, J.C.; Yamamoto, J. Astaxanthin inhibits thrombosis in cerebral vessels of stroke-prone spontaneously hypertensive rats. Nutr. Res. 2011, 31, 784–789. [Google Scholar] [CrossRef]
- Khan, S.K.; Malinski, T.; Mason, R.P.; Kubant, R.; Jacob, R.F.; Fujioka, K.; Denstaedt, S.J.; King, T.J.; Jackson, H.L.; Hieber, A.D.; et al. Novel astaxanthin prodrug (CDX-085) attenuates thrombosis in a mouse model. Thromb. Res. 2010, 126, 299–305. [Google Scholar] [CrossRef]
- Choi, H.D.; Kim, J.H.; Chang, M.J.; Kyu-Youn, Y.; Shin, W.G. Effects of astaxanthin on oxidative stress in overweight and obese adults. Phytother. Res. 2011, 25, 1813–1818. [Google Scholar] [CrossRef]
- Park, J.S.; Chyun, J.H.; Kim, Y.K.; Line, L.L.; Chew, B.P. Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutr. Metab. 2010, 7, 18. [Google Scholar] [CrossRef]
- Coombes, J.S.; Sharman, J.E.; Fassett, R.G. Astaxanthin has no effect on arterial stiffness, oxidative stress, or inflammation in renal transplant recipients: A randomized controlled trial (the XANTHIN trial). Am. J. Clin. Nutr. 2016, 103, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Sakayanathan, P.; Loganathan, C.; Thayumanavan, P. Astaxanthin-S-Allyl Cysteine Ester Protects Pancreatic β-Cell From Glucolipotoxicity by Suppressing Oxidative Stress, Endoplasmic Reticulum Stress and mTOR Pathway Dysregulation. J. Biochem. Mol. Toxicol. 2024, 38, e70058. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, L.; Guo, Y.; Zhang, T.; Qiao, X.; Wang, J.; Xu, J.; Xue, C. Hydrophilic Astaxanthin: PEGylated Astaxanthin Fights Diabetes by Enhancing the Solubility and Oral Absorbability. J. Agric. Food Chem. 2020, 68, 3649–3655. [Google Scholar] [CrossRef]
- Ying, C.J.; Zhang, F.; Zhou, X.Y.; Hu, X.T.; Chen, J.; Wen, X.R.; Sun, Y.; Zheng, K.Y.; Tang, R.X.; Song, Y.J. Anti-inflammatory Effect of Astaxanthin on the Sickness Behavior Induced by Diabetes Mellitus. Cell Mol. Neurobiol. 2015, 35, 1027–1037. [Google Scholar] [CrossRef]
- Kanwugu, O.N.; Glukhareva, T.V.; Danilova, I.G.; Kovaleva, E.G. Natural antioxidants in diabetes treatment and management: Prospects of astaxanthin. Crit. Rev. Food Sci. Nutr. 2022, 62, 5005–5028. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Ma, D.; Mulati, A.; Zhao, B.; Liu, F.; Liu, X. Development of astaxanthin-loaded layer-by-layer emulsions: Physicochemical properties and improvement of LPS-induced neuroinflammation in mice. Food Funct. 2021, 12, 5333–5350. [Google Scholar] [CrossRef]
- Okada, Y.; Ishikura, M.; Maoka, T. Bioavailability of astaxanthin in Haematococcus algal extract: The effects of timing of diet and smoking habits. Biosci. Biotechnol. Biochem. 2009, 73, 1928–1932. [Google Scholar] [CrossRef]
- Chang, H.I.; Shao, C.W.; Huang, E.; Huang, K.Y. Development of Astaxanthin-Loaded Nanosized Liposomal Formulation to Improve Bone Health. Pharmaceuticals 2022, 15, 490. [Google Scholar] [CrossRef]
- Mercke Odeberg, J.; Lignell, A.; Pettersson, A.; Hoglund, P. Oral bioavailability of the antioxidant astaxanthin in humans is enhanced by incorporation of lipid based formulations. Eur. J. Pharm. Sci. 2003, 19, 299–304. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, S.; McClements, D.J.; Wang, D.; Xu, Y. Design of Astaxanthin-Loaded Core-Shell Nanoparticles Consisting of Chitosan Oligosaccharides and Poly(lactic- co-glycolic acid): Enhancement of Water Solubility, Stability, and Bioavailability. J. Agric. Food Chem. 2019, 67, 5113–5121. [Google Scholar] [CrossRef]
- Abdol Wahab, N.R.; Meor Mohd Affandi, M.M.R.; Fakurazi, S.; Alias, E.; Hassan, H. Nanocarrier System: State-of-the-Art in Oral Delivery of Astaxanthin. Antioxidants 2022, 11, 1676. [Google Scholar] [CrossRef]
- Xue, Y.; Liao, Y.; Wang, H.; Li, S.; Gu, Z.; Adu-Frimpong, M.; Yu, J.; Xu, X.; Smyth, H.D.C.; Zhu, Y. Preparation and evaluation of astaxanthin-loaded 2-hydroxypropyl-beta-cyclodextrin and Soluplus(R) nanoparticles based on electrospray technology. J. Sci. Food Agric. 2023, 103, 3628–3637. [Google Scholar] [CrossRef]
- Madhavi, D.; Kagan, D.; Seshadri, S. A Study on the Bioavailability of a Proprietary, Sustained-release Formulation of Astaxanthin. Integr. Med. 2018, 17, 38–42. [Google Scholar]
- Yang, L.; Li, F.; Cao, X.; Qiao, X.; Xue, C.; Xu, J. Stability and bioavailability of protein matrix-encapsulated astaxanthin ester microcapsules. J. Sci. Food Agric. 2022, 102, 2144–2152. [Google Scholar] [CrossRef] [PubMed]
- Nalawade, P.; Gajjar, A. Optimization of Astaxanthin microencapsulation in hydrophilic carriers using response surface methodology. Arch. Pharm. Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhang, H.; Liu, R.; Zhu, H.; Zhang, L.; Tsao, R. Bioaccessibility, Cellular Uptake, and Transport of Astaxanthin Isomers and their Antioxidative Effects in Human Intestinal Epithelial Caco-2 Cells. J. Agric. Food Chem. 2017, 65, 10223–10232. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Qiao, X.; Gu, J.; Li, X.; Cao, Y.; Xu, J.; Xue, C. Influence of molecular structure of astaxanthin esters on their stability and bioavailability. Food Chem. 2021, 343, 128497. [Google Scholar] [CrossRef]
- Háda, M.; Petrovics, D.; Nagy, V.; Böddi, K.; Deli, J.; Agócs, A. The first synthesis of PEG–carotenoid conjugates. Tetrahedron Lett. 2011, 52, 3195–3197. [Google Scholar] [CrossRef]
- Lockwood, S.F.; Gross, G.J. Disodium disuccinate astaxanthin (Cardax): Antioxidant and antiinflammatory cardioprotection. Cardiovasc. Drug Rev. 2005, 23, 199–216. [Google Scholar] [CrossRef]
- Khayyal, M.T.; Teaima, M.H.; Marzouk, H.M.; Hazek, R.M.E.; Behnam, F.; Behnam, D. Comparative Pharmacokinetic Study of Standard Astaxanthin and its Micellar Formulation in Healthy Male Volunteers. Eur. J. Drug Metab. Pharmacokinet. 2024, 49, 467–475. [Google Scholar] [CrossRef]
- Liu, A.; He, M.; Liu, C.; Ye, Z.; Tan, C.P.; Liu, Y.; Gong, J.; Lei, J.; He, Y.; Zhu, S.; et al. Prevention of Hypercholesterolemia with “Liposomes in Microspheres” Composite Carriers: A Promising Approach for Intestinal-Targeted Oral Delivery of Astaxanthin. J. Agric. Food Chem. 2024, 72, 6118–6132. [Google Scholar] [CrossRef] [PubMed]
- Coral-Hinostroza, G.N.; Ytrestøyl, T.; Ruyter, B.; Bjerkeng, B. Plasma appearance of unesterified astaxanthin geometrical E/Z and optical R/S isomers in men given single doses of a mixture of optical 3 and 3’R/S isomers of astaxanthin fatty acyl diesters. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 139, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Østerlie, M.; Bjerkeng, B.; Liaaen-Jensen, S. Plasma appearance and distribution of astaxanthin E/Z and R/S isomers in plasma lipoproteins of men after single dose administration of astaxanthin. J. Nutr. Biochem. 2000, 11, 482–490. [Google Scholar] [CrossRef]
| Target Category | Specific Molecular Target | Bioactivity | Reference |
|---|---|---|---|
| Antioxidant systems | ROS/RNS, SOD, GPX, Nrf2 | scavenge free radicals and enhance endogenous antioxidant capacity | Yin et al. (2021) [10] |
| Inflammatory signaling pathways | NF-κB, MAPK, COX-2, IL-6 and iNOS | inhibit the release of inflammatory factors and reduce tissue inflammation | Wu et al., 2024 [11], Feng et al. (2018) [12] |
| Apoptosis/proliferation | Bcl-2, p53 and cyclin D1 | regulate cell survival and inhibit abnormal proliferation | Faraone et al. (2020) [13] |
| Metabolic regulation/insulin signaling pathway | PPARs, AMPK, IRS/PI3K/Akt | improve glucose and lipid metabolism and enhance insulin sensitivity | Inoue et al. (2012) [14], Lewis et al. (2022) [15] |
| Mitochondrial protection | mPTP and mitochondrial membrane protein | protect mitochondrial membrane and inhibit the opening of mPTP | Baburina et al. (2019) [16] |
| Mechanism | Study Population/Model | Key Findings | Reference |
|---|---|---|---|
| Anti-tumor effect | Prostate cancer DU145 cell | Suppression of tumor cell proliferation and metastasis by inhibiting STAT3 expression | Sun et al. (2020) [54] |
| PC-3 prostate cancer xenograft mouse model cells | Validated inhibitory effects on tumor growth | Ni et al. (2017) [55] | |
| Mouse skin papillomas | Antioxidant properties and capacity to scavenge peroxynitrite | Maoka et al. (2012) [56] | |
| SW480 cell and colorectal cancer mouse model | Curtails tumor cell proliferation and migration in colorectal cancer by regulating MAPK and NF-κB signaling pathways | Zhang et al. (2024) [57] | |
| C666-1 cell | Inhibits proliferation, migration, and invasion in nasopharyngeal carcinoma by blocking PI3K/AKT and NF-κB pathways via miR-29a-3p | Xu et al. (2024) [58] | |
| F344 rats | Inhibits the proliferation and migration of esophageal cancer cells by upregulating PPARγ expression | Cui et al. (2022) [59] | |
| U251MG cell | Hormetic effect in glioblastoma multiforme, where low concentrations promote cell proliferation, while high concentrations induce apoptosis | Tsuji et al. (2020) [60] |
| Mechanism | Study Population/Model | Key Findings | Reference |
|---|---|---|---|
| Antioxidant Properties | Pancreatic β-cell | Protecting pancreatic β-cells, and enhancing insulin secretion | Sakayanathan et al. (2024) [102] |
| Anti-inflammation effect | Fat- and high-sucrose-diet-induced insulin-resistant mouse model | Strengthening endogenous antioxidant system, and mitigating oxidative damage | Liu et al. (2020) [103] |
| Diabetic mice model | Decreasing GFAP-positive cells in the brain, downregulating the cleaved caspase-3, IL-6, and IL-1β, and upregulating CBS in the frontal cortex | Ying et al. (2015) [104] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Chen, X.; Wang, M.; Hu, H. Astaxanthin: A Compound in the Prevention of Chronic Diseases and as a Potential Adjuvant Treatment Agent. Antioxidants 2025, 14, 715. https://doi.org/10.3390/antiox14060715
Zhu X, Chen X, Wang M, Hu H. Astaxanthin: A Compound in the Prevention of Chronic Diseases and as a Potential Adjuvant Treatment Agent. Antioxidants. 2025; 14(6):715. https://doi.org/10.3390/antiox14060715
Chicago/Turabian StyleZhu, Xiao, Xi Chen, Matthew Wang, and Honghua Hu. 2025. "Astaxanthin: A Compound in the Prevention of Chronic Diseases and as a Potential Adjuvant Treatment Agent" Antioxidants 14, no. 6: 715. https://doi.org/10.3390/antiox14060715
APA StyleZhu, X., Chen, X., Wang, M., & Hu, H. (2025). Astaxanthin: A Compound in the Prevention of Chronic Diseases and as a Potential Adjuvant Treatment Agent. Antioxidants, 14(6), 715. https://doi.org/10.3390/antiox14060715