Global Neural Activities Changes under Human Inhibitory Control Using Translational Scenario

Abstract
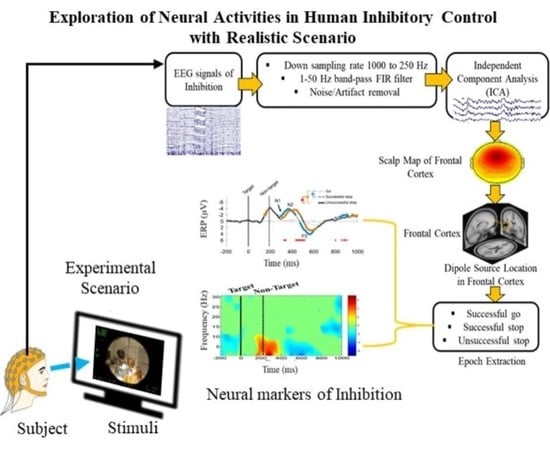
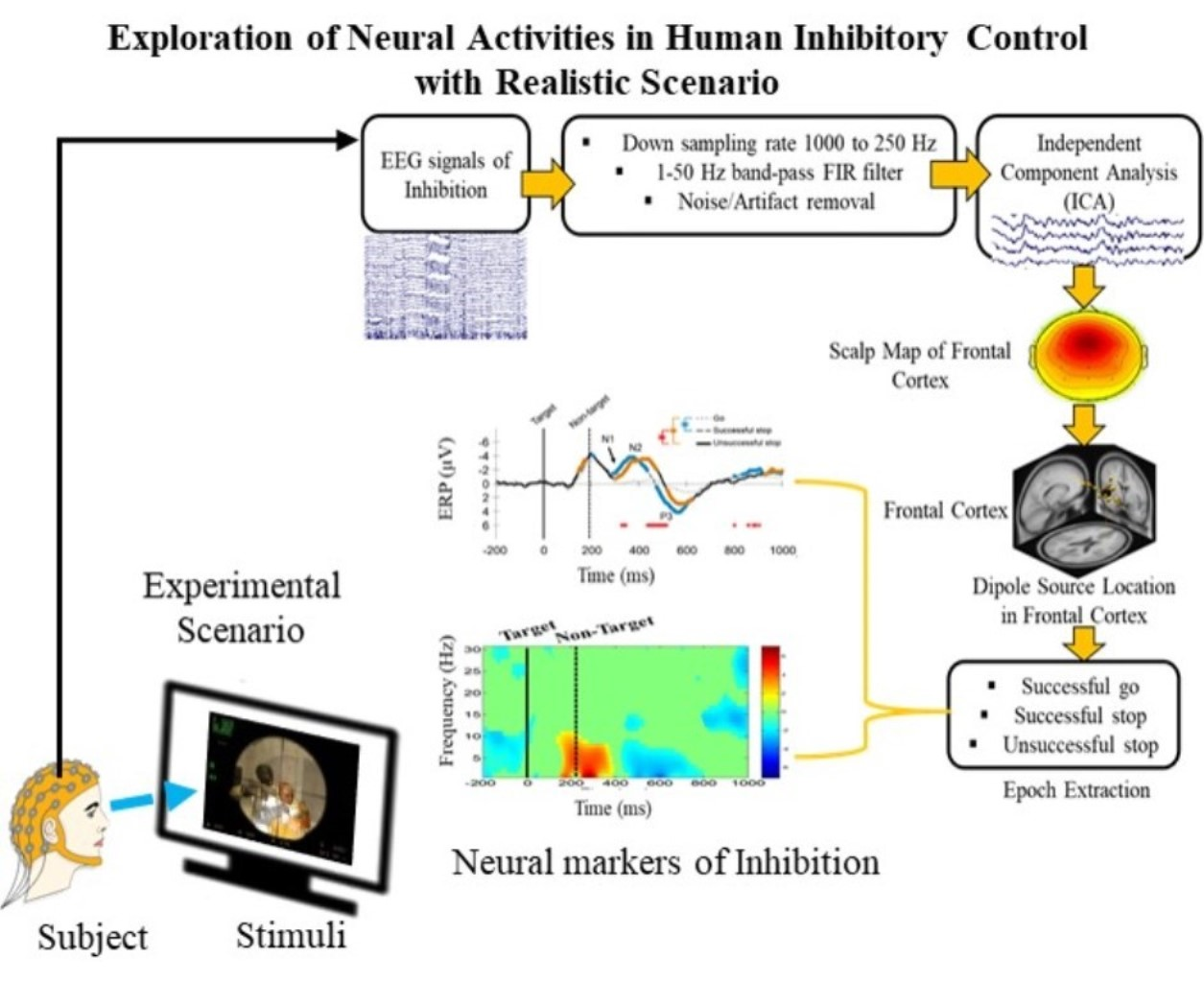{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Participants
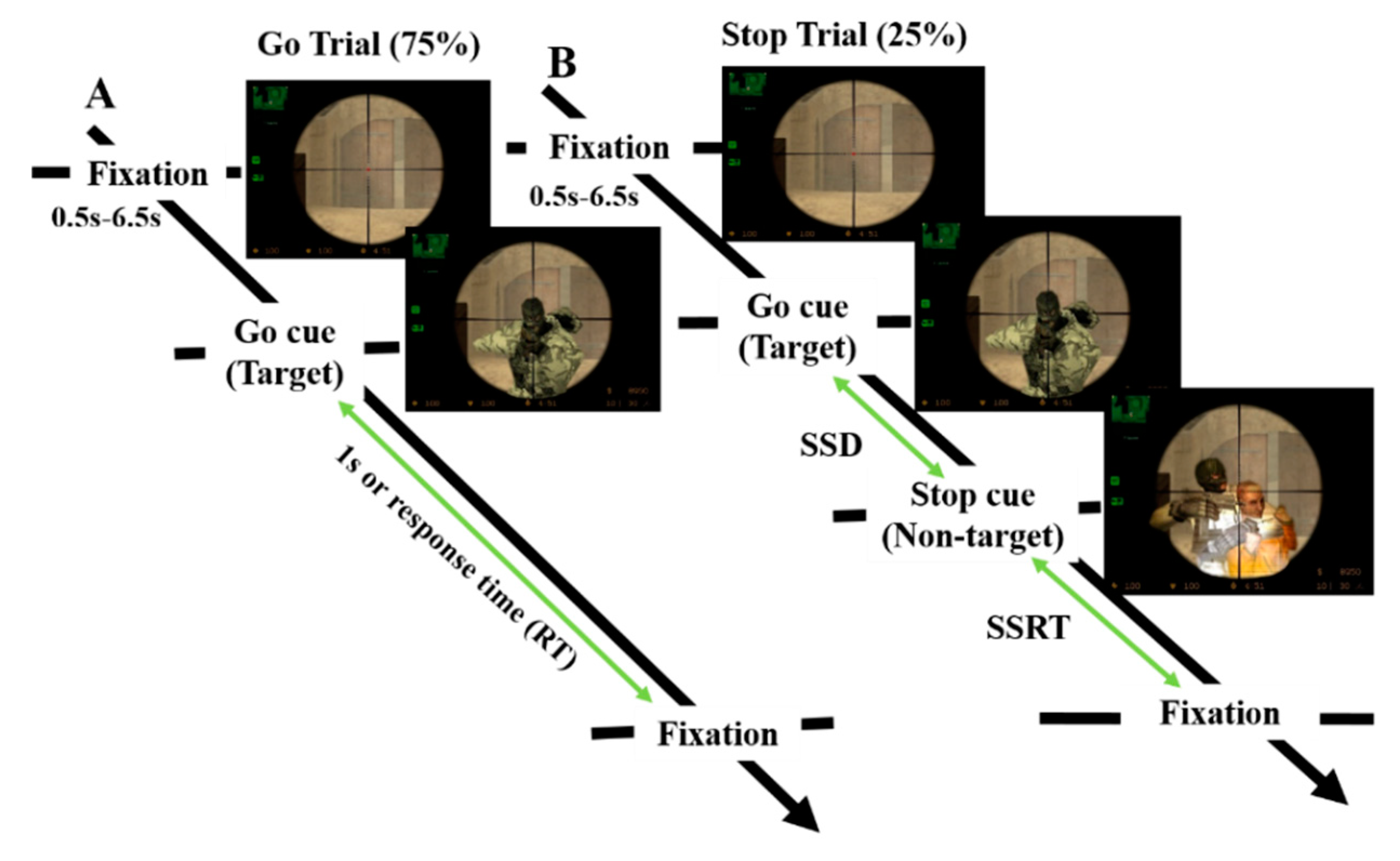
2.2. Experimental Design
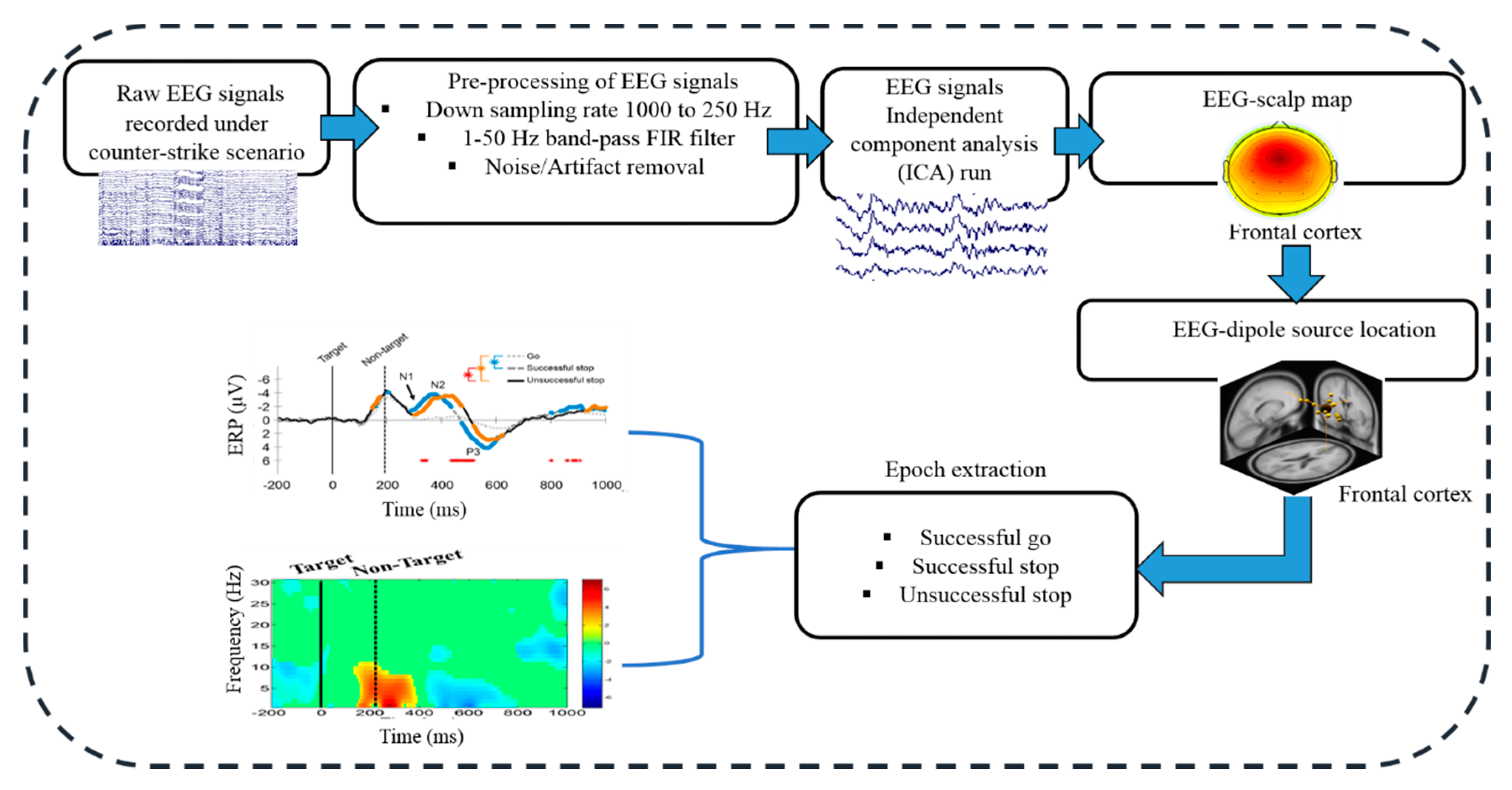
2.3. Acquisition and Preprocessing of EEG Signals
2.4. Independent Component Analysis (ICA)
2.5. Behavioral Analysis
2.6. Event-Related Spectral Perturbation (ERSP) Analysis
2.7. Statistical Analysis
3. Results
3.1. Behavioral Results
3.2. Electroencephalography (EEG) Results
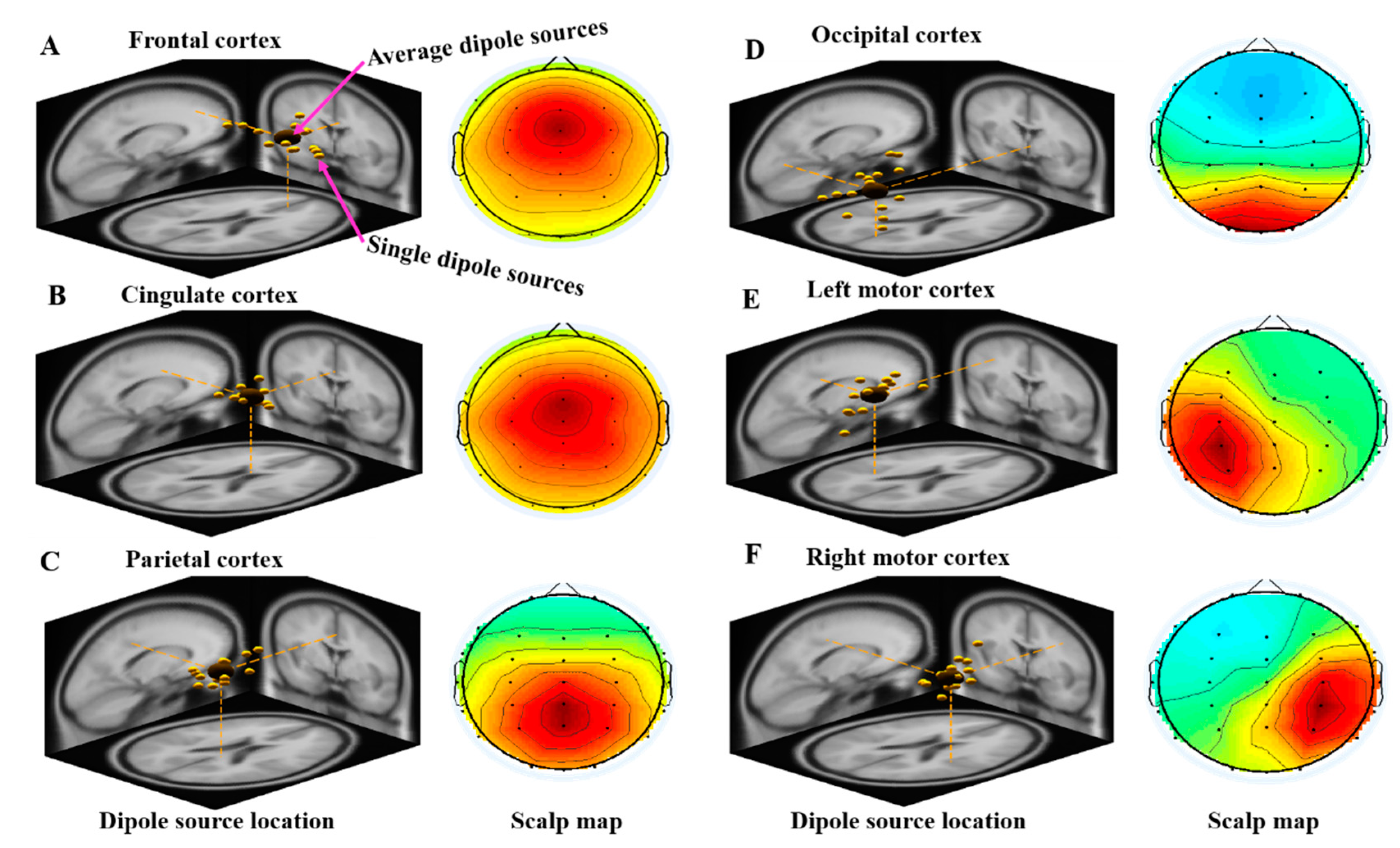
3.2.1. Independent Component Scalp Map and Location of Dipole Sources
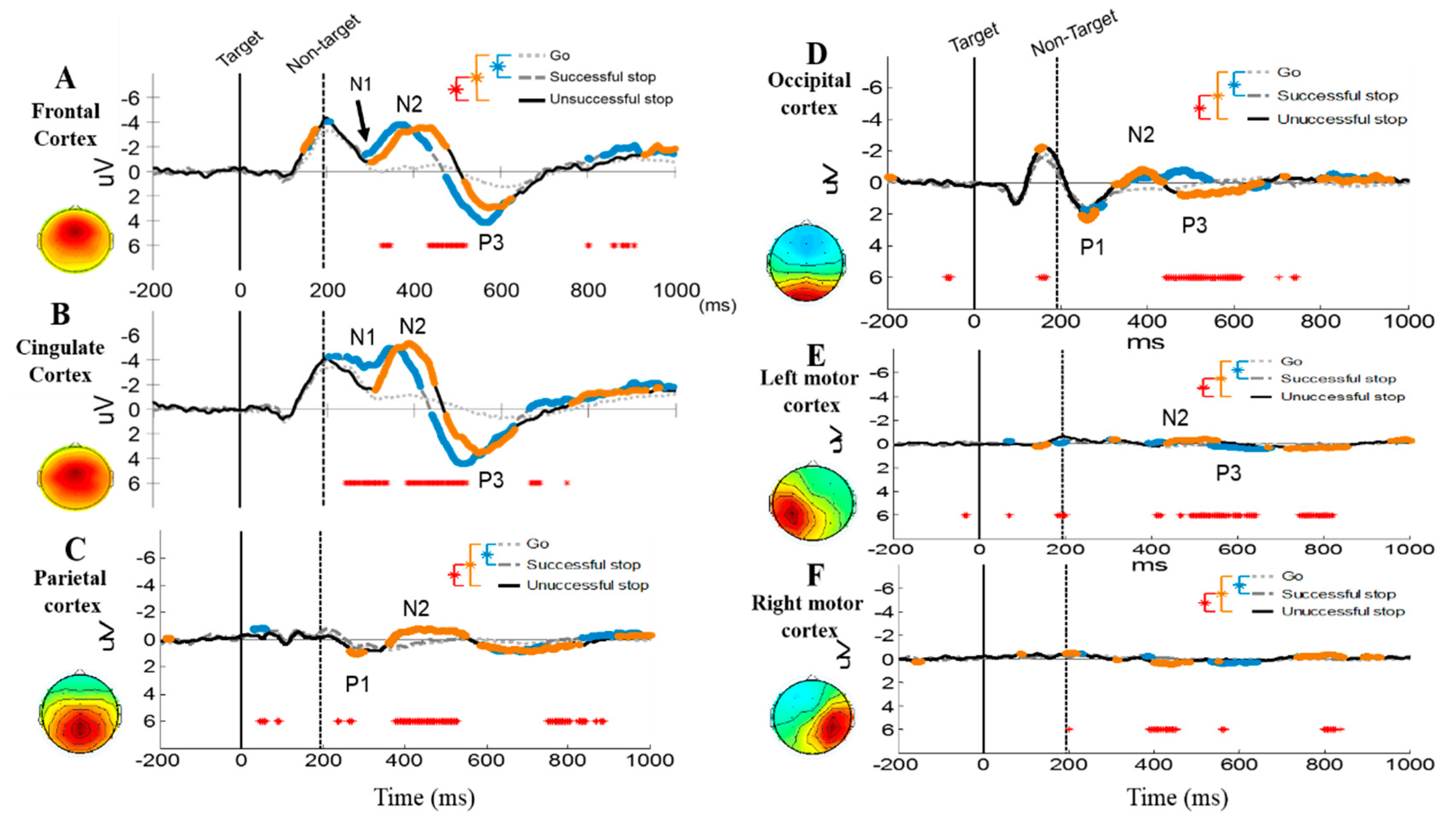
3.2.2. Event-Related Potentials (ERP) under Human Inhibition
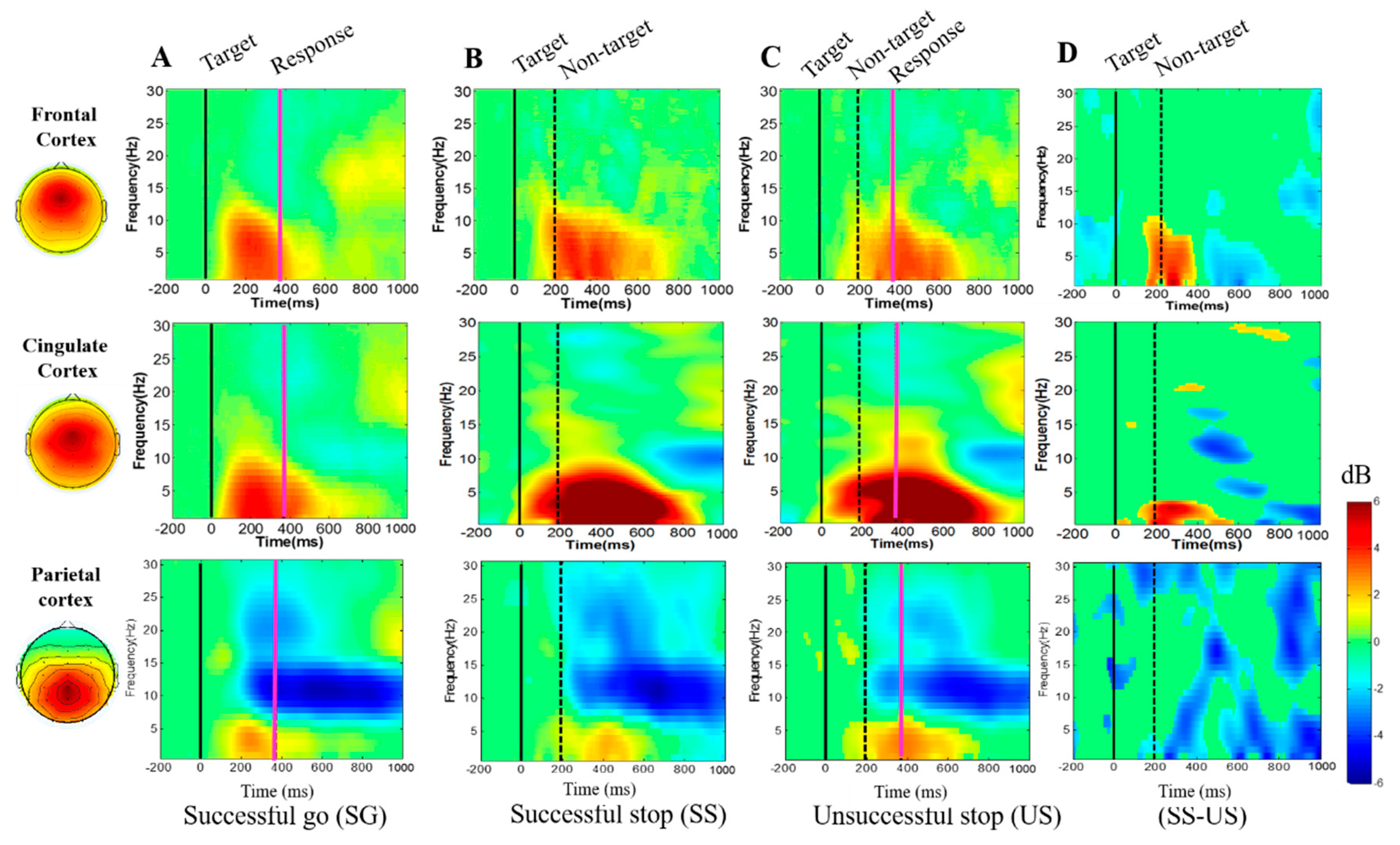
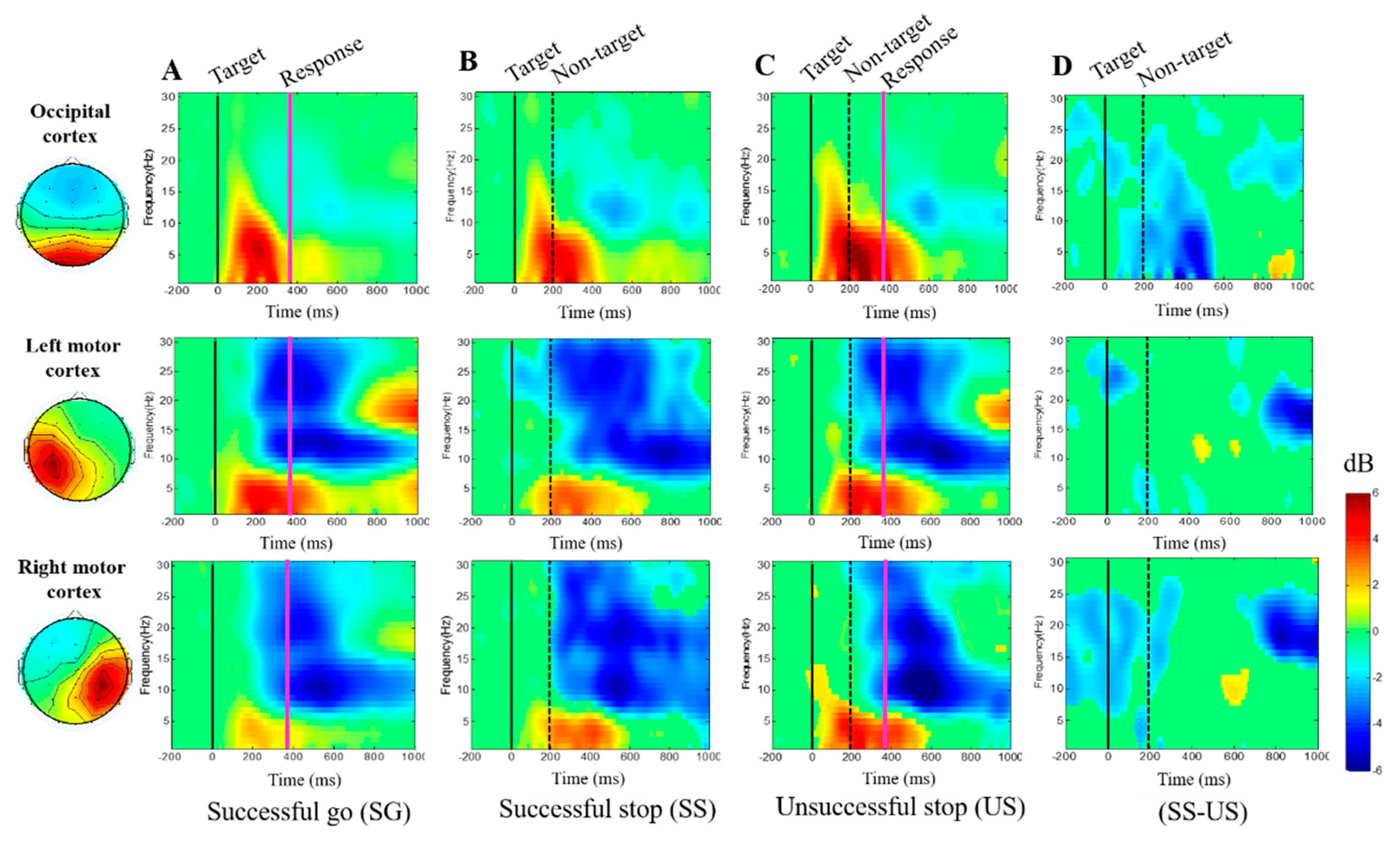
3.2.3. Event-Related Spectral Perturbation (ERSP) during Human Inhibition
4. Discussion
4.1. EEG-ERP N2 and P3 Waves under Human Inhibitory Control
4.2. EEG-Power Spectral Changes under Inhibitory Control
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lappin, J.S.; Eriksen, C.W. Use of a delayed signal to stop a visual reaction-time response. J. Exp. Psychol. 1966, 72, 805–811. [Google Scholar] [CrossRef]
- Logan, G.D.; Burkell, J. Dependence and independence in responding to double stimulation: A comparison of stop, change and dual-task paradigms. J. Exp. Psychol. Hum. Percept. Perform. 1986, 12, 549–563. [Google Scholar] [CrossRef]
- Logan, G.D.; Cowan, W.B. On the ability to inhibit thought and action: A theory of an act of control. Psychol. Rev. 1984, 91, 295–327. [Google Scholar] [CrossRef]
- Logan, G.D. On the ability to inhibit complex movements: A stop-signal study of typewriting. J. Exp. Psychol. Hum. Percept. Perform. 1982, 8, 778–792. [Google Scholar] [CrossRef]
- Logan, G.D. Executive control of thought and action. Acta Psychol. 1985, 60, 193–210. [Google Scholar] [CrossRef]
- Band, G.P.H.; Van Der Molen, M.W.; Logan, G.D. Horse-race model simulations of the stop-signal procedure. Acta Psychol. 2003, 112, 105–142. [Google Scholar] [CrossRef]
- Tabu, H.; Mima, T.; Aso, T.; Takahashi, R.; Fukuyama, H. Functional relevance of pre-supplementary motor areas for the choice to stop during Stop signal task. Neurosci. Res. 2011, 70, 277–284. [Google Scholar] [CrossRef]
- Tabu, H.; Mima, T.; Aso, T.; Takahashi, R.; Fukuyama, H. Common inhibitory prefrontal activation during inhibition of hand and foot responses. NeuroImage 2012, 59, 3373–3378. [Google Scholar] [CrossRef]
- Verbruggen, F.; Logan, G.D. Response inhibition in the stop-signal paradigm. Trends Cogn. Sci. 2008, 12, 418–424. [Google Scholar] [CrossRef]
- Kok, A.; Ramautar, J.R.; De Ruiter, M.B.; Band, G.P.H.; Ridderinkhof, K.R. ERP components associated with successful and unsuccessful stopping in a stop-signal task. Psychophysiology 2004, 41, 9–20. [Google Scholar] [CrossRef]
- Mostofsky, S.H.; Schafer, J.G.B.; Abrams, M.T.; Goldberg, M.C.; Flower, A.A.; Boyce, A.; Courtney, S.M.; Calhoun, V.D.; Kraut, M.A.; Denckla, M.B.; et al. fMRI evidence that the neural basis of response inhibition is task-dependent. Cogn. Brain Res. 2003, 17, 419–430. [Google Scholar] [CrossRef]
- Rubia, K.; Russella, T.; Overmeyera, S.; Brammer, M.J.; Bullmore, E.T.; Sharmaa, T.; Simmonsa, A.; Williams, S.C.; Giampietro, V.; Andrew, C.M.; et al. Mapping motor inhibition: Conjunctive brain activations across different versions of go/no-go and stop tasks. NeuroImage 2001, 13, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Rubia, K.; Smith, A.B.; Brammer, M.J.; Taylor, E. Right inferior prefrontal cortex mediates response inhibition while mesial prefrontal cortex is responsible for error detection. NeuroImage 2003, 20, 351–358. [Google Scholar] [CrossRef]
- Hampshire, A.; Chamberlain, S.R.; Monti, M.M.; Duncan, J.; Owen, A.M. The role of the right inferior frontal gyrus: Inhibition and attentional control. NeuroImage 2010, 50, 1313–1319. [Google Scholar] [CrossRef]
- Nakata, H.; Sakamoto, K.; Ferretti, A.; Perrucci, M.G.; Del Gratta, C.; Kakigi, R.; Romani, G.L. Somato-motor inhibitory processing in humans: An event-related functional MRI study. NeuroImage 2008, 39, 1858–1866. [Google Scholar] [CrossRef]
- Lansbergen, M.M.; Schutter, D.J.; Kenemans, J.L. Subjective impulsivity and baseline EEG in relation to stopping performance. Brain Res. 2007, 1148, 161–169. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- De Jong, R.; Coles, M.G.H.; Logan, G.D.; Gratton, G. In search of the point of no return: The control of response processes. J. Exp. Psychol. Hum. Percep. Perform. 1990, 16, 164–182. [Google Scholar] [CrossRef]
- Jong, D.R.; Coles, M.G.H.; Logan, G.D. Strategies and mechanisms in nonselective and selective inhibitory motor control. J. Exp. Psychol. Hum. Percep. Perform. 1995, 21, 498–511. [Google Scholar] [CrossRef]
- Naito, E.; Matsumura, M. Movement-related potentials associated with motor inhibition under different preparatory states during performance of two visual stop signal paradigms in humans. Neuropsychology 1996, 34, 565–573. [Google Scholar] [CrossRef]
- Gennady, G.K.; Evgenij, A.L.; Alexander, N.S. A failure to stop and attention fluctuations: An evoked oscillations study of the stop-signal paradigm. Clin. Neurophysiol. 2008, 119, 556–567. [Google Scholar]
- Pfefferbaum, A.; Ford, J.M.; Weller, B.J.; Kopell, B.S. ERPs to response production and inhibition. Electroencephalogr. Clin. Neurophysiol. 1985, 60, 423–434. [Google Scholar] [CrossRef]
- Kok, A. Effects of degradation of visual stimulation on components of the event-related potential (ERP) in go/nogo reaction tasks. Biol. Psychol. 1986, 3, 21–38. [Google Scholar] [CrossRef]
- Dimoska, A.; Johnstone, S.J.; Barry, R.J. The auditory-evoked N2 and P3 components in the stop-signal task: Indices of inhibition, response-conflict or error-detection? Brain Cogn. 2006, 62, 98–112. [Google Scholar] [CrossRef]
- Chikara, R.K.; Su, K.; Ko, L.W. Classification of Brain Activities under Response Inhibition using Functional Connectivity. In Proceedings of the International Conference on Technologies and Applications of Artificial Intelligence (TAAI), Kaohsiung, Taiwan, 21–23 November 2019; pp. 1–5. [Google Scholar] [CrossRef]
- Huster, R.J.; Enriquez-Geppert, S.; Lavallee, C.F.; Falkenstein, M.; Herrmann, C.S. Electroencephalography of response inhibition tasks: Functional networks and cognitive contributions. Int. J. Psychophysiol. 2013, 87, 217–233. [Google Scholar] [CrossRef]
- Gennari, S.P.; Millman, R.; Hymers, M.; Mattys, S.L. Anterior paracingulate and cingulate cortex mediates the effects of cognitive load on speech sound discrimination. NeuroImage 2018, 178, 735–743. [Google Scholar] [CrossRef]
- Drevets, W.C.; Savitz, J.; Trimble, M. The subgenual anterior cingulate cortex in mood disorders. CNS Spectr. 2008, 13, 663–681. [Google Scholar] [CrossRef]
- Adams, R.; David, A.S. Patterns of anterior cingulate activation in schizophrenia: A selective review. Neuropsychiatr. Dis. Treat. 2007, 3, 87–101. [Google Scholar] [CrossRef]
- Ko, L.-W.; Shih, Y.-C.; Chikara, R.K.; Chuang, Y.-T.; Chang, E.C. Neural mechanisms of inhibitory response in a battlefield scenario: A simultaneous fMRI-EEG study. Front. Hum. Neurosci. 2016, 10, 185. [Google Scholar] [CrossRef]
- Chikara, R.K.; Chang, E.C.; Lu, Y.-C.; Lin, D.-S.; Lin, C.-T.; Ko, L.-W. Monetary reward and punishment to response inhibition modulate activation and synchronization within the inhibitory brain network. Front. Hum. Neurosci. 2018, 12, 27. [Google Scholar] [CrossRef]
- Chikara, R.K.; Ko, L.-W. Modulation of the visual to auditory human inhibitory brain network: An EEG dipole source localization study. Brain Sci. 2019, 9, 216. [Google Scholar] [CrossRef]
- Mishkin, M.; Ungerleider, L.G. Contribution of striate inputs to the visuospatial functions of parieto-preoccipital cortex in monkeys. Behav. Brain Res. 1982, 6, 57–77. [Google Scholar] [CrossRef]
- Goodale, M.A.; Milner, A.D. Separate visual pathways for perception and action. Trends Neurosci. 1992, 15, 20–25. [Google Scholar] [CrossRef]
- Logan, G.D.; Cowan, W.B.; Davis, K.A. On the ability to inhibit simple and choice reaction time responses: A model and a method. J. Exp. Psychol. Hum. Percep. Perform 1984, 10, 275–291. [Google Scholar] [CrossRef]
- Jung, T.-P.; Makeig, S.; McKeown, M.J.; Bell, A.J.; Lee, T.-W.; Sejnowski, T. Imaging brain dynamics using independent component analysis. Proc. IEEE 2001, 89, 1107–1122. [Google Scholar] [CrossRef]
- Aron, A.R.; Robbins, T.; Poldrack, R.A. Inhibition and the right inferior frontal cortex. Trends Cogn. Sci. 2004, 8, 170–177. [Google Scholar] [CrossRef]
- Van Boxtel, G.; Van Der Molen, M.W.; Jennings, J.; Brunia, C.H. A psychophysiological analysis of inhibitory motor control in the stop-signal paradigm. Boil. Psychol. 2001, 58, 229–262. [Google Scholar] [CrossRef]
- Sharp, D.J.; Bonnelle, V.; De Boissezon, X.; Beckmann, C.F.; James, S.G.; Patel, M.C.; Mehta, M.A. Distinct frontal systems for response inhibition, attentional capture, and error processing. Proc. Natl. Acad. Sci. USA 2010, 107, 6106–6111. [Google Scholar] [CrossRef]
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; Monographs on Statistics & Applied Probability; Chapman Hall/CRC: Boca Raton, FL, USA, 1994; Volume 1, 456p. [Google Scholar]
- Makeig, S.; Debener, S.; Onton, J.; Delorme, A. Mining event-related brain dynamics. Trends Cogn. Sci. 2004, 8, 204–210. [Google Scholar] [CrossRef]
- Andrés, P. Frontal cortex as the central executive of working memory: Time to revise our view. Cortex 2003, 39, 871–895. [Google Scholar] [CrossRef]
- Aron, A.R. The neural basis of inhibition in cognitive control. Neuroscientist 2007, 13, 214–228. [Google Scholar] [CrossRef] [PubMed]
- Miyake, A.; Friedman, N.P.; Emerson, M.J.; Witzki, A.H.; Howerter, A.; Wager, T.D. The Unity and diversity of executive functions and their contributions to complex “frontal lobe” tasks: A latent variable analysis. Cogn. Psychol. 2000, 41, 49–100. [Google Scholar] [CrossRef] [PubMed]
- Stuphorn, V.; Schall, J.D. Executive control of countermanding saccades by the supplementary eye field. Nat. Neurosci. 2006, 9, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Ladefoged, P.; Silverstein, R.; Papçun, G. Interruptibility of speech. J. Acoust. Soc. Am. 1973, 54, 1105–1108. [Google Scholar] [CrossRef]
- Aron, A.R.; Poldrack, R.A. Cortical and subcortical contributions to stop signal response inhibition: Role of the subthalamic nucleus. J. Neurosci. 2006, 26, 2424–2433. [Google Scholar] [CrossRef]
- Chikara, R.K.; Lo, W.-C.; Ko, L.-W. Exploration of brain connectivity during human inhibitory control using inter-trial coherence. Sensors 2020, 20, 1722. [Google Scholar] [CrossRef]
- Nigg, J.T. Is ADHD a disinhibitory disorder. Psychol. Bull. 2001, 127, 571–598. [Google Scholar] [CrossRef]
- Schachar, R.; Logan, G.D. Impulsivity and inhibitory control in normal development and childhood psychopathology. Dev. Psychol. 1990, 26, 710–720. [Google Scholar] [CrossRef]
- Gauggel, S.; Rieger, M.; Feghoff, T.-A. Inhibition of ongoing responses in patients with Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 2004, 75, 539–544. [Google Scholar] [CrossRef]
- Chamberlain, S.R.; Fineberg, N.A.; Blackwell, A.D.; Robbins, T.W.; Sahakian, B.J. Motor inhibition and cognitive flexibility in obsessive–compulsive disorder and trichotillomania. Am. J. Psychiatry 2006, 163, 1282–1284. [Google Scholar] [CrossRef]
- Chikara, R.K.; Komarov, O.; Ko, L.W. Neural signature of event-related N200 and P300 modulation in parietal lobe during human response inhibition. Int. J. Comput. Biol. Drug Des. 2018, 11, 171–182. [Google Scholar] [CrossRef]
- Munro, G.E.; Dywan, J.; Harris, G.T.; McKee, S.; Unsal, A.; Segalowitz, S.J. Response inhibition in psychopathy: The frontal N2 and P3. Neurosci. Lett. 2007, 418, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Chikara, R.K.; Ko, L.-W. Neural Activities Classification of Human Inhibitory Control Using Hierarchical Model. Sensors 2019, 19, 3791. [Google Scholar] [CrossRef] [PubMed]
- Kirmizi-Alsana, E.; Bayraktaroglua, Z.; Gurvitb, H.; Keskina, Y.H.; Emreb, M.; Demiralpa, T. Comparative analysis of event-related potentials during go/no-go and CPT: Decomposition of response inhibition and sustained attention. Brain Res. 2006, 1104, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Harmony, T.; Alba, A.; Marroquin, J.L.; González-Frankenberger, B. Time-frequency-topographic analysis of induced power and synchrony of EEG signals during a Go/No-Go task. Int. J. Psychophysiol. 2009, 71, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Nigbur, R.; Ivanova, G.; Stürmer, B. Theta power as a marker for cognitive interference. Clin. Neurophysiol. 2011, 122, 2185–2194. [Google Scholar] [CrossRef] [PubMed]
- Pfurtscheller, G.; Neuper, C.; Flotzinger, D.; Pregenzer, M. EEG-based discrimination between imagination of right and left hand movement. Electroencephalogr. Clin. Neurophysiol. 1997, 103, 642–651. [Google Scholar] [CrossRef]
- Solis-Escalante, T.; Müller-Putz, G.R.; Pfurtscheller, G.; Neuper, C. Cue-induced beta rebound during withholding of overt and covert foot movement. Clin. Neurophysiol. 2012, 123, 1182–1190. [Google Scholar] [CrossRef]
- Cunnington, R.; Windischberger, C.; Deecke, L.; Moser, E. The Preparation and execution of self-initiated and externally-triggered movement: A study of event-related fMRI. NeuroImage 2002, 15, 373–385. [Google Scholar] [CrossRef]
- Ko, L.-W.; Chikara, R.K.; Lee, Y.-C.; Lin, W.-C. Exploration of user’s mental state changes during performing brain–computer interface. Sensors 2020, 20, 3169. [Google Scholar] [CrossRef]
- Banis, S.; Geerligs, L.; Lorist, M.M. Acute stress modulates feedback processing in men and women: Differential effects on the feedback-related negativity and theta and beta power. PLoS ONE 2014, 9, e95690. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chikara, R.K.; Ko, L.-W. Global Neural Activities Changes under Human Inhibitory Control Using Translational Scenario. Brain Sci. 2020, 10, 640. https://doi.org/10.3390/brainsci10090640
Chikara RK, Ko L-W. Global Neural Activities Changes under Human Inhibitory Control Using Translational Scenario. Brain Sciences. 2020; 10(9):640. https://doi.org/10.3390/brainsci10090640
Chicago/Turabian StyleChikara, Rupesh Kumar, and Li-Wei Ko. 2020. "Global Neural Activities Changes under Human Inhibitory Control Using Translational Scenario" Brain Sciences 10, no. 9: 640. https://doi.org/10.3390/brainsci10090640
APA StyleChikara, R. K., & Ko, L.-W. (2020). Global Neural Activities Changes under Human Inhibitory Control Using Translational Scenario. Brain Sciences, 10(9), 640. https://doi.org/10.3390/brainsci10090640