
Production and Chemical Characterization of Exopolysaccharides by Antarctic Yeasts Vishniacozyma victoriae and Tremellomycetes sp.

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Antarctic Yeasts
2.2. Growth Medium and Cultivation Conditions
2.3. Determination of the Biomass and EPS in the Culture Medium
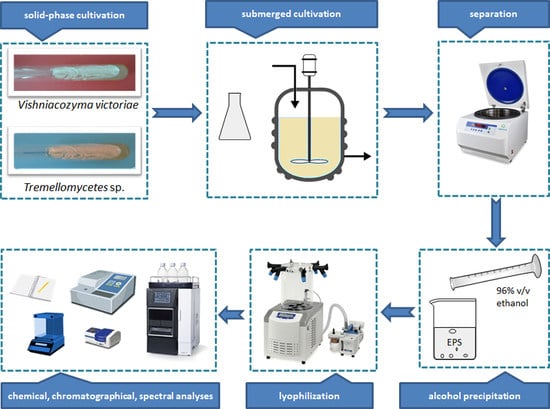
2.4. Isolation of EPSs
2.5. General Analytical Methods
2.6. Monosaccharide Composition Analysis
2.7. Molecular Weight Distribution Analysis
2.8. Fourier Transform Infrared (FT-IR) Spectroscopy
2.9. Statistics
3. Results
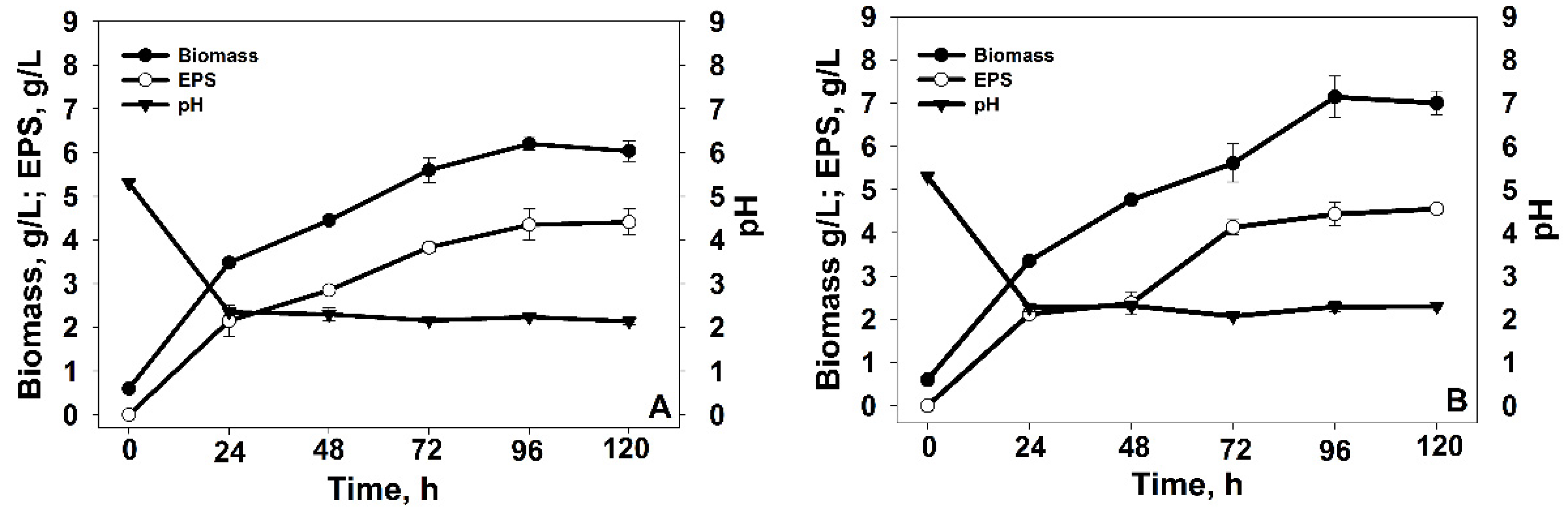
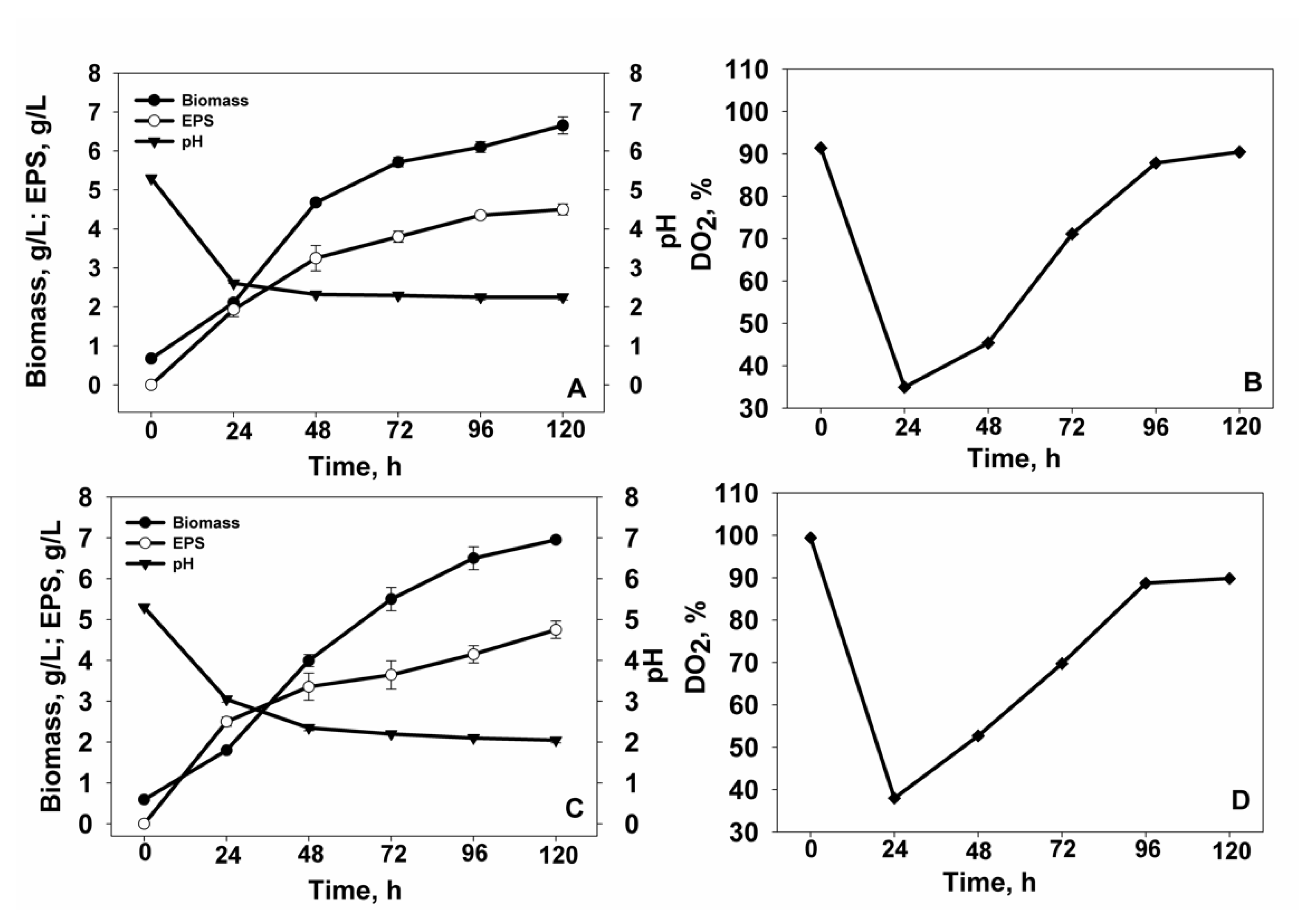
3.1. Identification and Cultivation of Yeast Strains
3.2. Chemical Characterization of EPSs
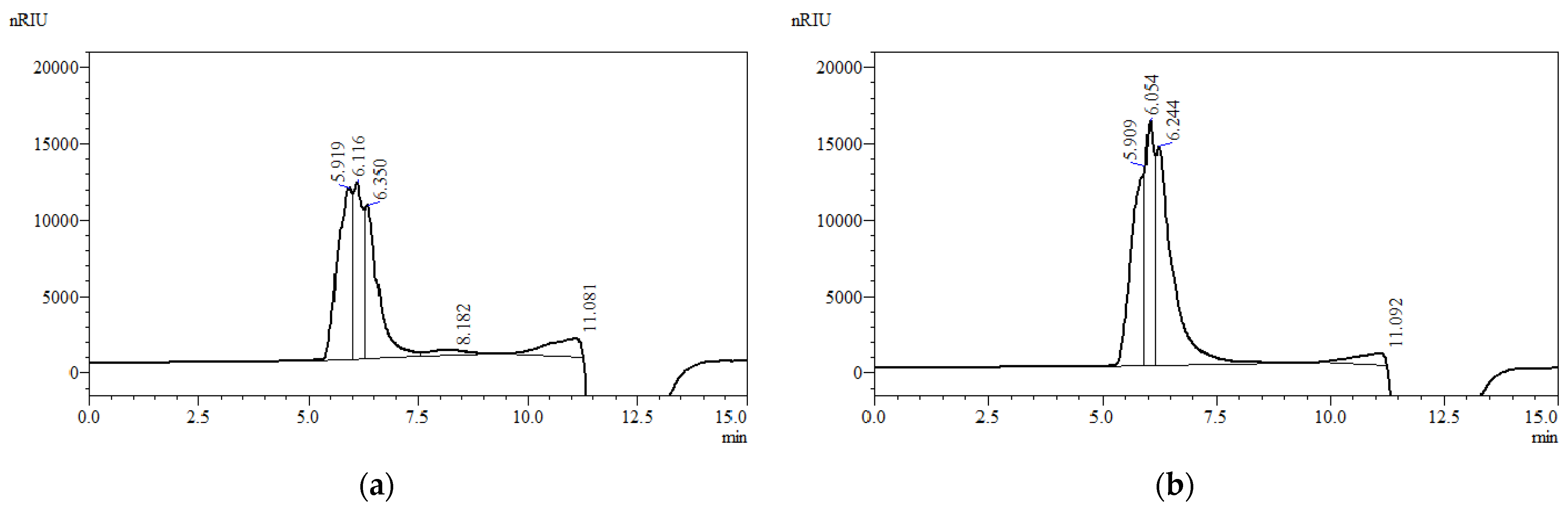
3.3. Molecular Weight Distribution Analysis of EPSs
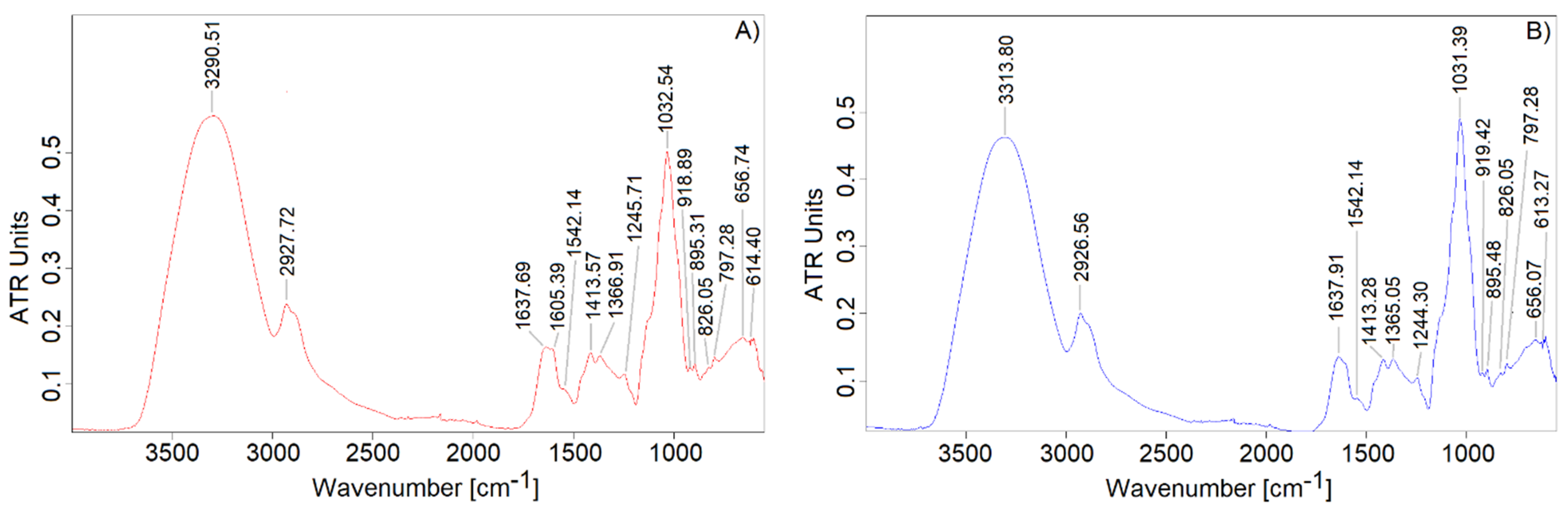
3.4. FT-IR Spectral Characterization of EPSs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buzzini, P.; Margesin, R. Cold-Adapted Yeasts: A Lesson from the Cold and a Challenge for the XXI Century. In Cold-Adapted Yeasts: Biodiversity, Adaptation Strategies and Biotechnological Significance; Buzzini, P., Margesin, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 3–22. ISBN 978-3-642-39681-6. [Google Scholar]
- Żymańczyk-Duda, E.; Brzezińska-Rodak, M.; Klimek-Ochab, M.; Duda, M.; Zerka, A. Yeast as a versatile tool in biotechnology. In Yeast: Industrial Applications; Morata, A., Loira, I., Eds.; IntechOpen: London, UK, 2017; pp. 3–40. [Google Scholar]
- Buzzini, P.; Branda, E.; Goretti, M.; Turchetti, B. Psychrophilic yeasts from worldwide glacial habitats: Diversity, adaptation strategies and biotechnological potential. FEMS Microbiol. Ecol. 2012, 82, 217–241. [Google Scholar] [CrossRef] [PubMed]
- Zlatanov, M.; Pavlova, K.; Antova, G.; Angelova-Romova, M.; Georgieva, K.; Rusinova-Videva, S. Biomass production by Antarctic yeast strains: An investigation on lipid composition. Biotechnol. Biotechnol. Equip. 2010, 24, 2096–2101. [Google Scholar] [CrossRef]
- Dimitrova, S.; Pavlova, K.; Lukanov, L.; Korotkov, E.; Petrova, E.; Zagorchev, P.; Kuncheva, M. Production of metabolites with antioxidant and emulsifying properties by Antarctic strain Sporobolomyces salmonicolor AL1. Appl. Biochem. Biotechnol. 2013, 169, 301–311. [Google Scholar] [CrossRef]
- Debnath, A.; Das, B.; Devi, M.; Ram, R. Fungal exopolysaccharides: Types, production and application. In Microbial Polymers, Applications and Ecological Perspectives; Vaishnav, A., Choudhary, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 45–68. [Google Scholar]
- Elinov, N.; Ananieva, E.; Vitovskaya, G. Features of the biosynthesis and characteristics of the exoglycanns in yeasts of the genus Sporobolomyces. Microbiologiya 1992, 60, 466–470. [Google Scholar]
- Pavlova, K.; Rusinova-Videva, S.; Kuncheva, M.; Kratchanova, M.; Gocheva, M.; Dimitrova, S. Synthesis and characterization of an exopolysaccharide from Antarctic yeast strain Cryptococcus laurentii AL100. Appl. Biochem. Biotechnol. 2011, 163, 1038–1052. [Google Scholar] [CrossRef] [PubMed]
- Rusinova-Videva, S.; Pavlova, K.; Georgieva, K. Effect of different carbon sources on biosynthesis of exopolysaccharide from Antarctic strain Cryptococcus laurentii AL62. Biotechnol. Biotechnol. Equip. 2011, 25, 80–84. [Google Scholar] [CrossRef]
- Kuncheva, M.; Pavlova, K.; Panchev, I.; Dobreva, S. Emulsifying power of mannan and glucomannan produced by yeasts. Int. J. Cosmetic. Sci. 2007, 29, 377–384. [Google Scholar] [CrossRef]
- Williams, P.; Hickey, M. Fluid gels based on natural polymers for cosmetic applications. Cosmet. Toilet. 2003, 118, 51–59. [Google Scholar]
- Freitas, F.; Alves, V.; Reis, M. Advances in bacterial exopolysaccharides: From production to biotechnological applications. Trends Biotechnol. 2011, 29, 388–398. [Google Scholar] [CrossRef]
- Nishinari, K. Polysaccharide rheology and in-mouth perception. In Food Polysaccharides and Their Applications, 2nd ed.; Alistair, M., Glyn, O., Peter, A., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 541–588. [Google Scholar]
- Nicolaus, B.; Kambourova, M.; Oner, E. Exopolysaccharides from extremophiles: From fundamentals to biotechnology. Environ. Technol. 2010, 31, 1145–1158. [Google Scholar] [CrossRef]
- Chen, J.; Mao, D.; Yong, Y.; Li, J.; Wei, H.; Lu, L. Hepatoprotective and hypolipidemic effects of water-soluble polysaccharidic extract of Pleurotus eryngii. Food Chem. 2012, 130, 687–694. [Google Scholar] [CrossRef]
- Hu, S.; Liang, Z.; Chia, Y.; Lien, J.; Chen, K.; Lee, M.; Wang, J. Antihyperlipidemic effect of polysaccharide from fermented broth of Pleurotus citrinopileatus. Appl. Microbiol. Biotechnol. 2006, 70, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Hristova, D.; Rusinova-Videva, S.; Konstantinov, S. Biologically active substances and extracts of fungal original. Pharmacong. Rev. 2021, 15, 12–19. [Google Scholar]
- Vaz, A.B.; Rosa, L.H.; Vieira, M.L.; Garcia, V.D.; Brandao, L.R.; Teixeira, L.C.; Rosa, C.A. The diversity, extracellular enzymatic activities and photoprotectivecompounds of yeasts isolated in Antarctica. Braz. J. Microbiol. 2011, 42, 937–947. [Google Scholar] [CrossRef]
- Ogaki, M.B.; Teixeira, D.R.; Vieira, R.; Lírio, J.M.; Felizardo, J.P.; Abuchacra, R.C.; Cardoso, R.P.; Zani, C.L.; Alves, T.M.; Junior, P.A.; et al. Diversity and bioprospecting of cultivable fungal assemblages in sediments of likes in the Antarctic of lakes in the Antarctic Peninsula. Fungal Biol. 2020, 124, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Baeza, M.; Alcaíno, J.; Cifuentes, V.; Turchetti, B.; Buzzini, P. Cold active enzymes from cold-adapted yeasts. In Biotechnology of Yeasts and Filamentous Fungi; Sibirny, A.A., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 297–324. [Google Scholar] [CrossRef]
- Joseph, B.; Ramteke, P.W.; Thomas, G. Cold active microbial lipases: Some hot issues and recent developments. Biotechnol. Adv. 2008, 26, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, P.; Carrasco, M.; Barahona, S.; Alcaíno, J.; Cifuentes, V.; Baeza, M. Antarctic yeasts: Analysis of their freeze-thaw tolerance and production of antifreeze proteins, fatty acids and ergosterol. BMC Microbiol. 2018, 18, 66. [Google Scholar] [CrossRef]
- Lutz, M.C.; Lopes, C.A.; Sosa, M.C.; Sangorrín, M.P. Semi-commercial testing of regional yeasts selected from North Patagonia Argentina for the biocontrol of pear postharvest decays. Biol. Control 2020, 150, 104246. [Google Scholar] [CrossRef]
- Andrade, P.; de Sousa, J.; Lira, P.; Assis, A.; Lerlinck, S.; Andreote, D. The bacterial and fungal communities associated with Anthurium ssp. leaves: Insight into plant endemism and microbe association. Microbiol. Res. 2021, 244, 126667. [Google Scholar] [CrossRef]
- Rusinova-Videva, S.; Radchenkova, N.; Dobrev, G.; Pavlova, K. Purification of arabinomannan synthesized by Cryptococcus laurentii AL100. Acta Microbiol. Bulg. 2015, 31, 141–144. [Google Scholar]
- Rusinova-Videva, S.; Nachkova, S.; Adamov, A.; Dimitrova-Dyulgerova, I. Antarctic yeast Cryptococcus laurentii (AL65): Biomass and exopolysaccharide production and biosorption of metals. J. Chem. Technol. Biotechnol. 2020, 95, 1372–1379. [Google Scholar] [CrossRef]
- Vlaev, S.; Pavlova, K.; Rusinova-Videva, S.; Georgieva, K.; Georgiev, D. Agitation effects and kinetic constants of exoglucomannan production by Antarctic yeast strain in a stirred tank bioreactor. Chem. Biochem. Eng. Q. 2016, 30, 393–400. [Google Scholar] [CrossRef]
- Bradstreet, R.B. The Kjeldahl Method for Organic Nitrogen; Academic Press Inc.: New York, NY, USA, 1965; pp. 9–145. [Google Scholar]
- GB 5009.5-2016; Determination of Protein in Foods. National Food Safety Standard (NFSS) of the People’s Republic of China. China National Center for Food Safety Risk Assessment: Beijing, China, 2016.
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Karkhanis, Y.D.; Zeltner, J.Y.; Jackson, J.J.; Carlo, D.J. A new and improved microassay to determine 2-keto-3-deoxyoctonate in lipopolysaccharide of gramnegative bacteria. Anal. Biochem. 1978, 85, 595–601. [Google Scholar] [CrossRef]
- Ognyanov, M.; Georgiev, Y.; Petkova, N.; Ivanov, I.; Vasileva, I.; Kratchanova, M. Isolation and characterization of pectic polysaccharide fraction from in vitro suspension culture of Fumaria officinalis L. Int. J. Polym. Sci. 2018, 2018, 5705036. [Google Scholar] [CrossRef]
- McComb, E.A.; McCready, R.M. Determination of acetyl in pectin and in acetylated carbohydrate polymers. Anal. Chem. 1957, 29, 819–821. [Google Scholar] [CrossRef]
- Synytsya, A.; Čopíková, J.; Matĕjka, P.; Machovič, V. Fourier transform Raman and infrared spectroscopy of pectins. Carbohydr. Polym. 2003, 54, 97–106. [Google Scholar] [CrossRef]
- Singh, B.R.; DeOliveira, D.B.; Fu, F.-N.; Fuller, M.P. Fourier transform infrared analysis of amide III bands of proteins for the secondary structure estimation. In Proceedings of the SPIE, Biomolecular Spectroscopy III, Los Angeles, CA, USA, 1 May 1993; Volume 1890, pp. 47–55. [Google Scholar]
- Farinha, I.; Duarte, P.; Pimentel, A.; Plotnikova, E.; Chagas, B.; Mafra, L.; Grandfils, C.; Freitas, F.; Fortunato, E.; Reis, M.A.M. Chitin–glucan complex production by Komagataella pastoris: Downstream optimization and product characterization. Carbohydr. Polym. 2015, 130, 455–464. [Google Scholar] [CrossRef]
- Kačuráková, M.; Capek, P.; Sasinková, V.; Wellner, N.; Ebringerová, A. FT-IR study of plant cell wall model compounds: Pectic polysaccharides and hemicelluloses. Carbohydr. Polym. 2000, 43, 195–203. [Google Scholar] [CrossRef]
- Šandula, J.; Kogan, G.; Kačuráková, M.; Machová, E. Microbial (1→3)-β-d-glucans, their preparation, physico-chemical characterization and immunomodulatory activity. Carbohydr. Polym. 1999, 38, 247–253. [Google Scholar] [CrossRef]
- Galichet, A.; Sockalingum, G.D.; Belarbi, A.; Manfait, M. FTIR spectroscopic analysis of Saccharomyces cerevisiae cell walls: Study of an anomalous strain exhibiting a pink-colored cell phenotype. FEMS Microbiol. Lett. 2001, 197, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Nitta, M.; Mizuno, T. Infrared spectroscopy of some mannans. Agric. Biol. Chem. 1973, 37, 433–435. [Google Scholar] [CrossRef]
- Gientka, I.; Bzducha-Wróbel, A.; Stasiak-Różańska, L.; Bednarska, A.A.; Błażejak, S. The exopolysaccharides biosynthesis by Candida yeast depends on carbon sources. Electron. J. Biotechnol. 2016, 22, 31–37. [Google Scholar] [CrossRef]
- Ragavan, M.L.; Das, N. Optimization of exopolysaccharide production by probiotic yeast Lipomyces starkeyi VIT-MN03 using response surface methodology and its applications. Ann. Microbiol. 2019, 69, 515–530. [Google Scholar] [CrossRef]
- Breierová, E.; Hromádková, Z.; Stratilová, E.; Sasinková, V.; Ebringerová, A. Effect of salt stress on the production and properties of extracellular polysaccharides produced by Cryptococcus laurentii. Z. Nat. C 2005, 60, 444–450. [Google Scholar] [CrossRef]
- Hamidi, M.; Gholipour, A.R.; Delattre, C.; Sesdighi, F.; Seveiri, R.M.; Pasdaran, A.; Kheirandish, S.; Pierre, G.; Kozani, P.S.; Kozani, P.S.; et al. Production, characterization and biological activities of exopolysaccharides from a new cold-adapted yeast: Rhodotorula mucilaginosa sp. GUMS16. Int. J. Biol. Macromol. 2020, 151, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Oluwa, S.W. Structure and foaming properties of viscous exopolysaccharides from a wild grape-associated basidiomycetous yeast Papiliotrema flavescens formerly known as Cryptococcus flavescens. J. Microbiol. Biotechnol. 2020, 30, 1739–1749. [Google Scholar] [CrossRef]
- Van Bogaert, I.N.A.; De Maeseneire, S.L.; Vandamme, E.J. Extracellular polysaccharides produced by yeasts and yeast-like fungi. In Yeast Biotechnology: Diversity and Applications; Satyanarayana, T., Kunze, G., Eds.; Springer Science + Business Media, B.V.: Berlin/Heidelberg, Germany, 2009; pp. 651–671. [Google Scholar]
- Sajna, K.V.; Sukumaran, R.K.; Gottumukkala, L.D.; Jayamurthy, H.; Dhar, K.S.; Pandey, A. Studies on structural and physical characteristics of a novel exopolysaccharide from Pseudozyma sp. NII 08165. Int. J. Biol. Macromol. 2013, 59, 84–89. [Google Scholar] [CrossRef]
- Panchev, I.N.; Dobreva, S.; Karashanova, D.; Pavlova, K.; Kuncheva, M.; Georgieva, K. Physical properties of cosmetic creams containing exopolysaccharides synthesized from Antarctic yeast. In Proceedings of the Food Science, Engineering and Technologies, Plovdiv, Bulgaria, 19–20 October 2012. [Google Scholar]
- Chatterjee, S.; Mukhopadhyay, S.K.; Gauri, S.S.; Dey, S. Sphingobactan, a new α-mannan exopolysaccharide from Arctic Sphingobacterium sp. IITKGP-BTPF3 capable of biological response modification. Int. Immunopharmacol. 2018, 60, 84–95. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.K.; Chatterjee, S.; Gauri, S.S.; Das, S.S.; Mishra, A.; Patra, M.; Ghosh, A.K.; Das, A.K.; Singh, S.M.; Dey, S. Isolation and characterization of extracellular polysaccharide Thelebolan produced by a newly isolated psychrophilic Antarctic fungus Thelebolus. Carbohydr. Polym. 2014, 104, 204–212. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| V-EPS | T-EPS | |
|---|---|---|
| Yield, g EPS/100 g 1 (g EPS/L) 2 | 18.4 (4.0) | 20.7(4.5) |
| Total carbohydrates 3, w/w% | 75.4 | 79.0 |
| Mannose | 30 (37)4 | 32 (36) |
| Glucose | 18 (22) | 22 (25) |
| Xylose | 13 (19) | 16 (21) |
| Galactose | 11 (14) | 11 (12) |
| Uronic acids | 7.0 (8) | 6.3 (7) |
| Rare sugars test | + | + |
| Acetyl content, w/w% | n.f | n.f. |
| Protein, w/w% (N × 5.78) | 11.8 | 11.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusinova-Videva, S.; Ognyanov, M.; Georgiev, Y.; Kambourova, M.; Adamov, A.; Krasteva, V. Production and Chemical Characterization of Exopolysaccharides by Antarctic Yeasts Vishniacozyma victoriae and Tremellomycetes sp. Appl. Sci. 2022, 12, 1805. https://doi.org/10.3390/app12041805
Rusinova-Videva S, Ognyanov M, Georgiev Y, Kambourova M, Adamov A, Krasteva V. Production and Chemical Characterization of Exopolysaccharides by Antarctic Yeasts Vishniacozyma victoriae and Tremellomycetes sp. Applied Sciences. 2022; 12(4):1805. https://doi.org/10.3390/app12041805
Chicago/Turabian StyleRusinova-Videva, Snezhana, Manol Ognyanov, Yordan Georgiev, Margarita Kambourova, Aleksandar Adamov, and Vasilena Krasteva. 2022. "Production and Chemical Characterization of Exopolysaccharides by Antarctic Yeasts Vishniacozyma victoriae and Tremellomycetes sp." Applied Sciences 12, no. 4: 1805. https://doi.org/10.3390/app12041805
APA StyleRusinova-Videva, S., Ognyanov, M., Georgiev, Y., Kambourova, M., Adamov, A., & Krasteva, V. (2022). Production and Chemical Characterization of Exopolysaccharides by Antarctic Yeasts Vishniacozyma victoriae and Tremellomycetes sp. Applied Sciences, 12(4), 1805. https://doi.org/10.3390/app12041805