
Protective Effect and Potential Antioxidant Role of Kakadu Plum Extracts on Alcohol-Induced Oxidative Damage in HepG2 Cells

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Collection of Fruit Materials
2.3. Extract Preparation
2.4. HPLC System and Condition for Analysis of Chemical Contents of Fruits
2.5. Cell Culture and Treatment
2.6. Cell Viability Test
2.7. DPPH Scavenging Assay
2.8. Protocol for Reducing Power Activity
2.9. Measurement of Cellular ROS
2.10. Measurement of Cellular ADH Activity
2.11. Measurement of Cellular ALDH Activity
2.12. Statistical Analysis
3. Results and Discussion
3.1. Analysis of Vitamin C, Gallic Acid, Ellagic Acid, and Daidzin Using HPLC
3.2. Total Phenolic and Flavonoid Contents
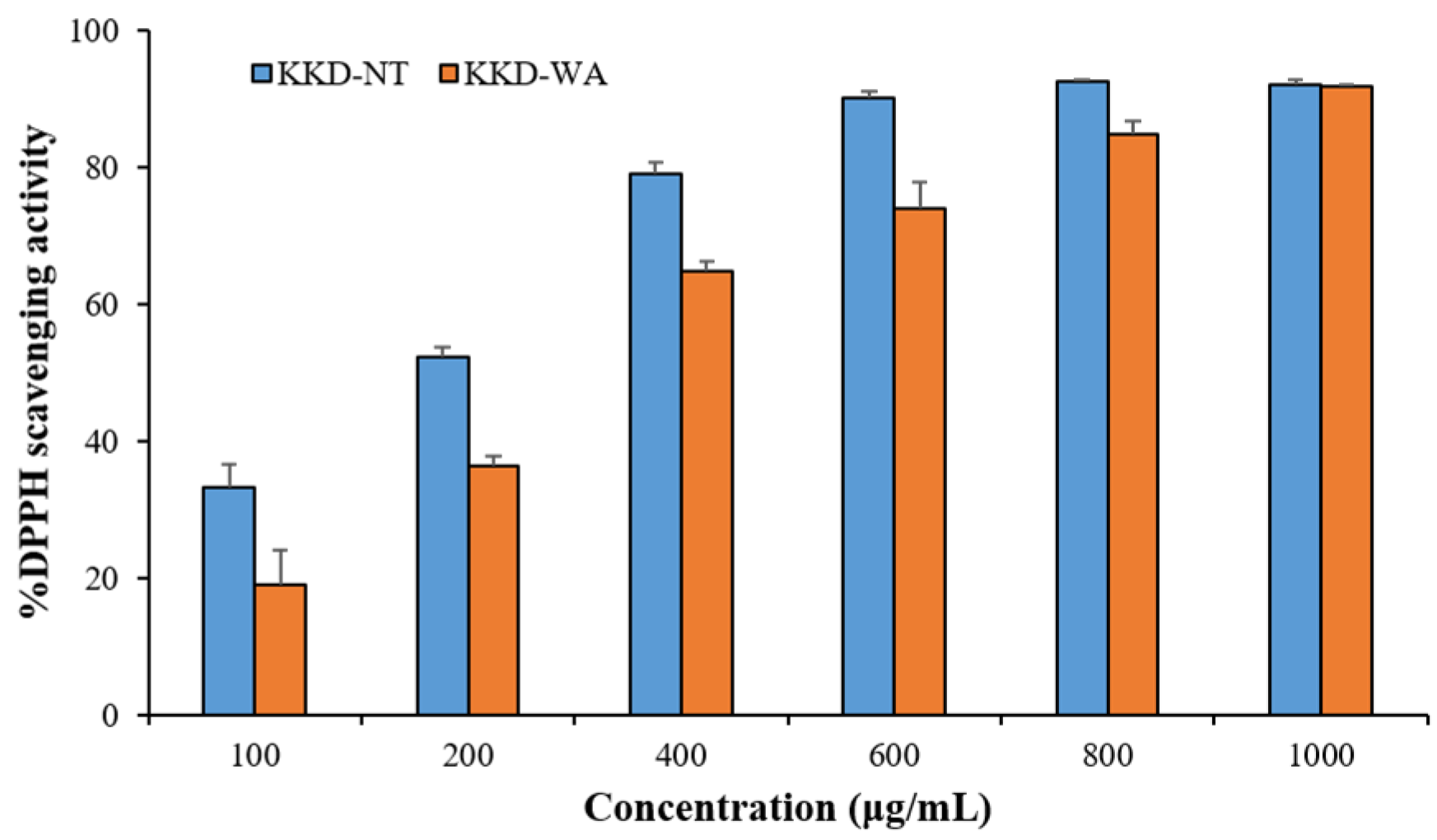
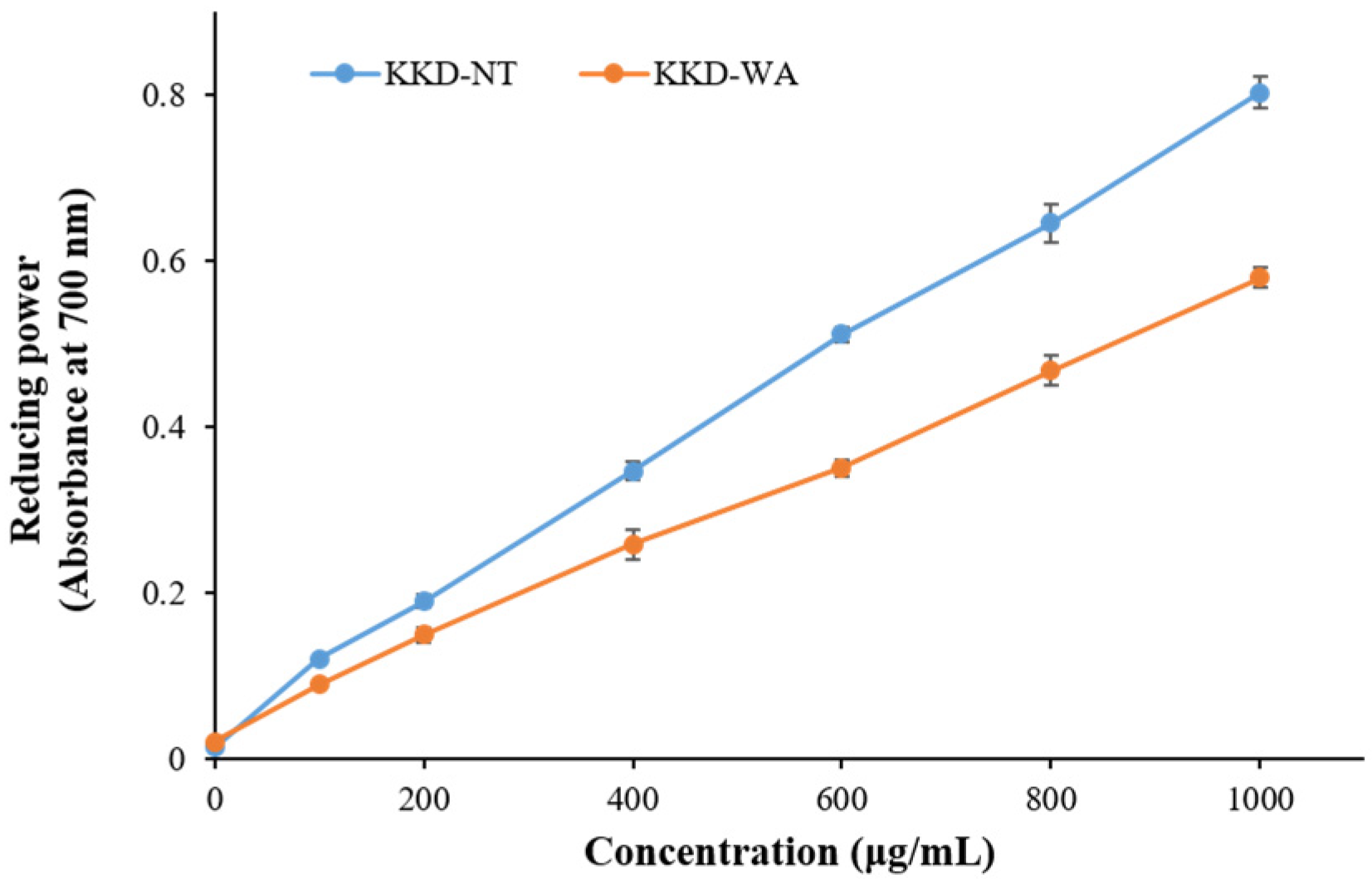
3.3. Antioxidant Activity of Plant Extracts: DPPH and Reducing Power Assays
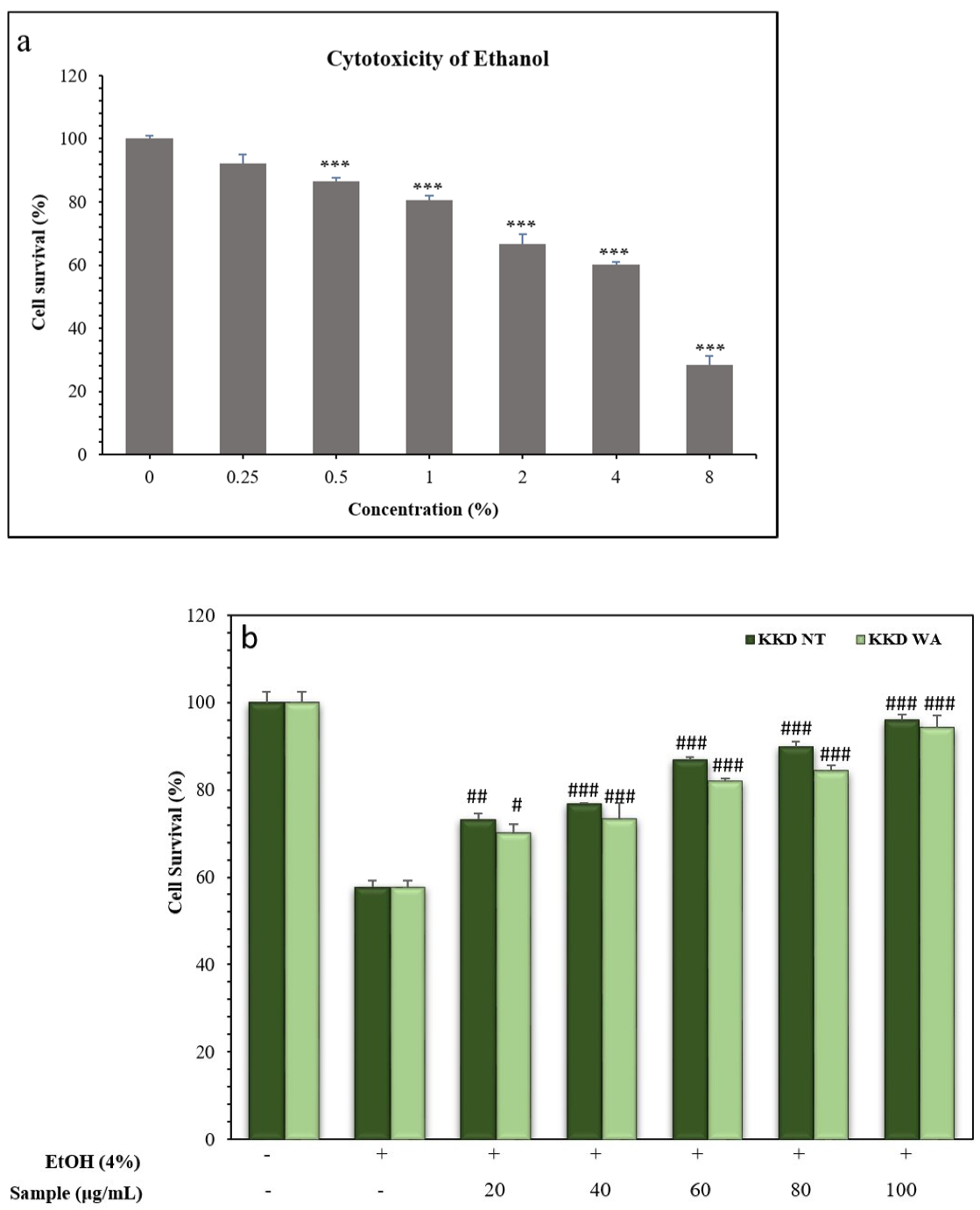
3.4. Ethanol Induces Cell Death
3.5. Kakadu Plum Protects HepG2 Cells from Ethanol-Induced Cell Death
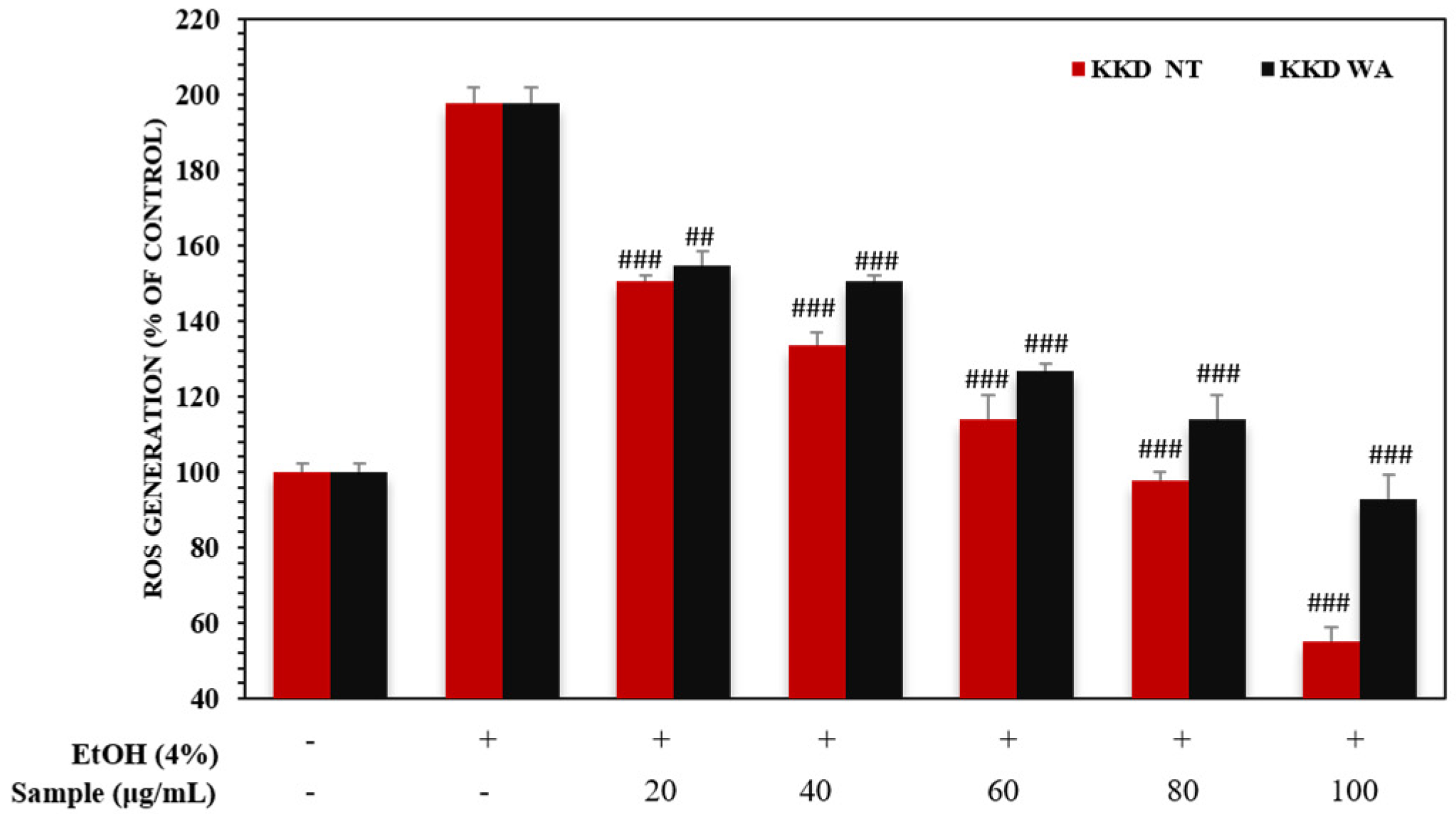
3.6. Kakadu Plum Suppresses Elevated Levels of ROS in HepG2 Cells
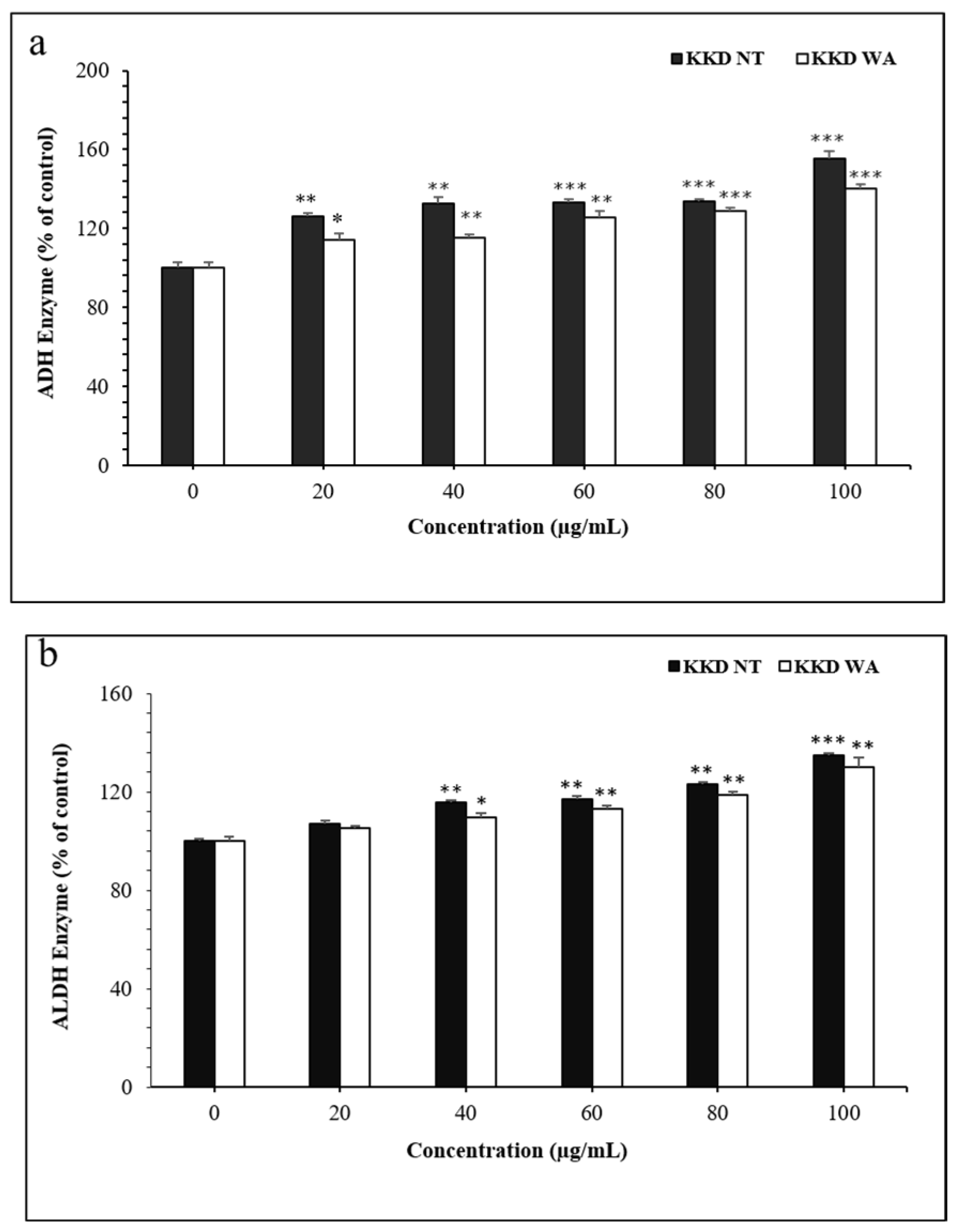
3.7. Effect of KKD-NT and KKD-WA on ADH and ALDH Activities
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Park, J.H.; Kim, Y.; Kim, S.H. Green tea extract (Camellia sinensis) fermented by Lactobacillus fermentum attenuates alcohol-induced liver damage. Biosci. Biotechnol. Biochem. 2012, 76, 2294–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudeep, H.; Venkatakrishna, K.; Sundeep, K.; Vasavi, H.; Raj, A.; Chandrappa, S.; Shyamprasad, K. Turcuron: A standardized bisacurone-rich turmeric rhizome extract for the prevention and treatment of hangover and alcohol-induced liver injury in rats. Pharmacogn. Mag. 2020, 16, 263. [Google Scholar] [CrossRef]
- Bobasa, E.M.; Phan, A.D.T.; Manolis, C.; Netzel, M.; Smyth, H.; Cozzolino, D.; Sultanbawa, Y. Effect of sample presentation on the near infrared spectra of wild harvest Kakadu plum fruits (Terminalia ferdinandiana). Infrared Phys. Technol. 2020, 111, 103560. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, T.; Zhang, H.; Sang, S.; Chen, H.; Zuo, X. Temporal trend of gastric cancer burden along with its risk factors in China from 1990 to 2019, and projections until 2030: Comparison with Japan, South Korea, and Mongolia. Biomark. Res. 2021, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Eshak, E.S.; Shirai, K.; Liu, K.; Dong, J.Y.; Iso, H.; Tamakoshi, A.; JACC Study Group. Alcohol Consumption and Risk of Gastric Cancer: The Japan collaborative cohort study. J. Epidemiol. 2021, 31, 30–36. [Google Scholar]
- Jung, K.-W.; Won, Y.-J.; Oh, C.-M.; Kong, H.-J.; Lee, D.H.; Lee, K.H. Cancer statistics in Korea: Incidence, mortality, survival, and prevalence in 2014. Cancer Res. Treat. 2017, 49, 292. [Google Scholar] [CrossRef]
- Panchenko, L.; Davydov, B.; Terebilina, N.; Baronets, V.Y.; Zhuravleva, A. Oxidative stress in the alcoholic liver disease. Biochem. (Mosc.) Suppl. Ser. B Biomed. Chem. 2014, 8, 73–76. [Google Scholar] [CrossRef]
- Ohashi, K.; Pimienta, M.; Seki, E. Alcoholic liver disease: A current molecular and clinical perspective. Liver Res. 2018, 2, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Goc, Z.; Kapusta, E.; Formicki, G.; Martiniaková, M.; Omelka, R. Effect of taurine on ethanol-induced oxidative stress in mouse liver and kidney. Chin. J. Physiol. 2019, 62, 148. [Google Scholar] [CrossRef]
- Lee, S.I.; Kim, H.J.; Boo, Y.C. Effect of green tea and (-)-epigallocatechin gallate on ethanol-induced toxicity in HepG2 cells. Phytother. Res. 2008, 22, 669–674. [Google Scholar] [CrossRef]
- Hyun, J.; Han, J.; Lee, C.; Yoon, M.; Jung, Y. Pathophysiological Aspects of Alcohol Metabolism in the Liver. Int. J. Mol. Sci. 2021, 22, 5717. [Google Scholar] [CrossRef] [PubMed]
- Zakhari, S. Overview: How is alcohol metabolized by the body? Alcohol Res. Health 2006, 29, 245. [Google Scholar]
- Vairappan, B. Cholesterol Regulation by Leptin in Alcoholic Liver Disease. In Molecular Aspects of Alcohol and Nutrition; Elsevier: Amsterdam, The Netherlands, 2016; pp. 187–200. [Google Scholar]
- Ghazali, R.; Patel, V.B. Alcohol metabolism: General aspects. In Molecular Aspects of Alcohol and Nutrition; Elsevier: Amsterdam, The Netherlands, 2016; pp. 17–21. [Google Scholar]
- Sha, K.; Choi, S.-H.; Im, J.; Lee, G.G.; Loeffler, F.; Park, J.H. Regulation of ethanol-related behavior and ethanol metabolism by the Corazonin neurons and Corazonin receptor in Drosophila melanogaster. PLoS ONE 2014, 9, e87062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Cederbaum, A.I. Alcohol, oxidative stress, and free radical damage. Alcohol Res. Health 2003, 27, 277. [Google Scholar] [PubMed]
- Poljsak, B.; Milisav, I. NAD+ as the link between oxidative stress, inflammation, caloric restriction, exercise, DNA repair, longevity, and health span. Rejuvenation Res. 2016, 19, 406–413. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Cederbaum, A.I. CYP2E1 and oxidative liver injury by alcohol. Free. Radic. Biol. Med. 2008, 44, 723–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- She, X.; Wang, F.; Ma, J.; Chen, X.; Ren, D.; Lu, J. In vitro antioxidant and protective effects of corn peptides on ethanol-induced damage in HepG2 cells. Food Agric. Immunol. 2016, 27, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Butterfield, D.A. Oxidative stress in neurodegenerative disorders. Antioxid. Redox Signal. 2006, 8, 1971–1973. [Google Scholar] [CrossRef]
- Clarke, P.A. Aboriginal People and Their Plants; Rosenberg Publishing: Kenthurst, NSW, Australia, 2007. [Google Scholar]
- Akter, S.; Netzel, M.E.; Fletcher, M.T.; Tinggi, U.; Sultanbawa, Y. Chemical and nutritional composition of Terminalia ferdinandiana (kakadu plum) kernels: A novel nutrition source. Foods 2018, 7, 60. [Google Scholar] [CrossRef] [Green Version]
- Gorman, J.T.; Wurm, P.A.; Vemuri, S.; Brady, C.; Sultanbawa, Y. Kakadu Plum (Terminalia ferdinandiana) as a sustainable indigenous agribusiness. Econ. Bot. 2020, 74, 74–91. [Google Scholar] [CrossRef]
- Brock, J. Native Plants of Northern Australia; Reed New Holland: Sydney, NSW, Australia, 2001. [Google Scholar]
- Phan, A.D.T.; Damyeh, M.S.; Chaliha, M.; Akter, S.; Fyfe, S.; Netzel, M.E.; Cozzolino, D.; Sultanbawa, Y. The effect of maturity and season on health-related bioactive compounds in wild harvested fruit of Terminalia ferdinandiana (Exell). Int. J. Food Sci. Technol. 2021, 56, 6431–6442. [Google Scholar] [CrossRef]
- Singh, A.; Bajpai, V.; Kumar, S.; Kumar, B.; Srivastava, M.; Rameshkumar, K. Comparative profiling of phenolic compounds from different plant parts of six Terminalia species by liquid chromatography–tandem mass spectrometry with chemometric analysis. Ind. Crops Prod. 2016, 87, 236–246. [Google Scholar] [CrossRef]
- Shami, A.-M.M.; Philip, K.; Muniandy, S. Synergy of antibacterial and antioxidant activities from crude extracts and peptides of selected plant mixture. BMC Complementary Altern. Med. 2013, 13, 360. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.J.; Edwards, D.; Pun, S.; Chaliha, M.; Burren, B.; Tinggi, U.; Sultanbawa, Y. Organic acids in Kakadu plum (Terminalia ferdinandiana): The good (ellagic), the bad (oxalic) and the uncertain (ascorbic). Food Res. Int. 2016, 89, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommano, S.; Caffin, N.; Kerven, G. Screening for antioxidant activity, phenolic content, and flavonoids from Australian native food plants. Int. J. Food Prop. 2013, 16, 1394–1406. [Google Scholar] [CrossRef] [Green Version]
- Tan, A.C.; Konczak, I.; Ramzan, I.; Sze, D.M.-Y. Native Australian fruit polyphenols inhibit cell viability and induce apoptosis in human cancer cell lines. Nutr. Cancer 2011, 63, 444–455. [Google Scholar] [CrossRef] [PubMed]
- Sirdaarta, J.; Matthews, B.; Cock, I. Kakadu plum fruit extracts inhibit growth of the bacterial triggers of rheumatoid arthritis: Identification of stilbene and tannin components. J. Funct. Foods 2015, 17, 610–620. [Google Scholar] [CrossRef]
- Tan, A.C.; Konczak, I.; Ramzan, I.; Zabaras, D.; Sze, D.M.-Y. Potential antioxidant, antiinflammatory, and proapoptotic anticancer activities of Kakadu plum and Illawarra plum polyphenolic fractions. Nutr. Cancer 2011, 63, 1074–1084. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.H.; Sirdaarta, J.; Matthews, B.; Greene, A.C.; Cock, I.E. Growth Inhibitory Activity of Kakadu Plum Extracts Against the Opportunistic Pathogenclostridium Perfringens: New Leads in the Prevention and Treatment of Clostridial Myonecrosis. Pharmacogn. J. 2016, 8, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Sprince, H.; Parker, C.M.; Smith, G.G.; Gonzales, L.J. Protective action of ascorbic acid and sulfur compounds against acetaldehyde toxicity: Implications in alcoholism and smoking. Agents Actions 1975, 5, 164–173. [Google Scholar] [CrossRef]
- Mohapatra, P.; Ray, A.; Jena, S.; Nayak, S.; Mohanty, S. Influence of extraction methods and solvent system on the chemical composition and antioxidant activity of Centella asiatica L. leaves. Biocatal. Agric. Biotechnol. 2021, 33, 101971. [Google Scholar] [CrossRef]
- Simu, S.Y.; Ahn, S.; Castro-Aceituno, V.; Singh, P.; Mathiyalangan, R.; Jiménez-Pérez, Z.E.; Yang, D. Gold nanoparticles synthesized with fresh panax ginseng leaf extract suppress adipogenesis by downregulating PPAR/CEBP signaling in 3T3-L1 mature adipocytes. J. Nanosci. Nanotechnol 2018, 18, 1–8. [Google Scholar]
- Markus, J.; Mathiyalagan, R.; Kim, Y.-J.; Abbai, R.; Singh, P.; Ahn, S.; Perez, Z.E.J.; Hurh, J.; Yang, D.C. Intracellular synthesis of gold nanoparticles with antioxidant activity by probiotic Lactobacillus kimchicus DCY51T isolated from Korean kimchi. Enzym. Microb. Technol. 2016, 95, 85–93. [Google Scholar] [CrossRef]
- Bhalodia, N.R.; Nariya, P.B.; Shukla, V.; Acharya, R. In vitro antioxidant activity of hydro alcoholic extract from the fruit pulp of Cassia fistula Linn. AYU (An Int. Q. J. Res. Ayurveda) 2013, 34, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Castro-Aceituno, V.; Ahn, S.; Simu, S.Y.; Singh, P.; Mathiyalagan, R.; Lee, H.A.; Yang, D.C. Anticancer activity of silver nanoparticles from Panax ginseng fresh leaves in human cancer cells. Biomed. Pharmacother. 2016, 84, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Ciuclan, L.; Ehnert, S.; Ilkavets, I.; Weng, H.-L.; Gaitantzi, H.; Tsukamoto, H.; Ueberham, E.; Meindl-Beinker, N.M.; Singer, M.V.; Breitkopf, K.; et al. TGF-β enhances alcohol dependent hepatocyte damage via down-regulation of alcohol dehydrogenase I. J. Hepatol. 2010, 52, 407–416. [Google Scholar] [CrossRef]
- Kim, B.Y.; Cui, Z.G.; Lee, S.R.; Kim, S.J.; Kang, H.K.; Lee, Y.K.; Park, D.B. Effects of Asparagus officinalis extracts on liver cell toxicity and ethanol metabolism. J. Food Sci. 2009, 74, H204–H208. [Google Scholar] [CrossRef] [PubMed]
- Bobasa, E.M.; Phan, A.D.T.; Netzel, M.E.; Cozzolino, D.; Sultanbawa, Y. Hydrolysable tannins in Terminalia ferdinandiana Exell fruit powder and comparison of their functional properties from different solvent extracts. Food Chem. 2021, 358, 129833. [Google Scholar] [CrossRef]
- Cozzolino, D.; Phan, A.D.T.; Netzel, M.E.; Smyth, H.; Sultanbawa, Y. The use of vibrational spectroscopy to predict vitamin C in Kakadu plum powders (Terminalia ferdinandiana Exell, Combretaceae). J. Sci. Food Agric. 2021, 101, 3208–3213. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, D.; Phan, A.D.T.; Aker, S.; Smyth, H.E.; Sultanbawa, Y. Can Infrared Spectroscopy Detect Adulteration of Kakadu Plum (Terminalia ferdinandiana) Dry Powder with Synthetic Ascorbic Acid? Food Anal. Methods 2021, 14, 1936–1942. [Google Scholar] [CrossRef]
- Courtney, R.; Sirdaarta, J.; Matthews, B.; Cock, I. Tannin components and inhibitory activity of Kakadu plum leaf extracts against microbial triggers of autoimmune inflammatory diseases. Pharmacogn. J. 2015, 7, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Phan, A.D.T.; Adiamo, O.; Akter, S.; Netzel, M.E.; Cozzolino, D.; Sultanbawa, Y. Effects of drying methods and maltodextrin on vitamin C and quality of Terminalia ferdinandiana fruit powder, an emerging Australian functional food ingredient. J. Sci. Food Agric. 2021, 101, 5132–5141. [Google Scholar] [CrossRef]
- Richmond, R.; Bowyer, M.; Vuong, Q. Australian native fruits: Potential uses as functional food ingredients. J. Funct. Foods 2019, 62, 103547. [Google Scholar] [CrossRef]
- Kaurinovic, B.; Vastag, D. Flavonoids and Phenolic Acids as Potential Natural Antioxidants; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Netzel, M.; Netzel, G.; Tian, Q.; Schwartz, S.; Konczak, I. Native Australian fruits—a novel source of antioxidants for food. Innov. Food Sci. Emerg. Technol. 2007, 8, 339–346. [Google Scholar] [CrossRef]
- Warinhomhoun, S.; Muangnoi, C.; Buranasudja, V.; Mekboonsonglarp, W.; Rojsitthisak, P.; Likhitwitayawuid, K.; Sritularak, B. Antioxidant Activities and Protective Effects of Dendropachol, a New Bisbibenzyl Compound from Dendrobium pachyglossum, on Hydrogen Peroxide-Induced Oxidative Stress in HaCaT Keratinocytes. Antioxidants 2021, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Jadid, N.; Hidayati, D.; Hartanti, S.R.; Arraniry, B.A.; Rachman, R.Y.; Wikanta, W. Antioxidant activities of different solvent extracts of Piper retrofractum Vahl. using DPPH assay. In AIP Conference Proceedings, Proceeding of the International Biology Conference 2016: Biodiversity and Biotechnology for Human Welfare, Surabaya, Indonesia, 15 October 2016; Murkovic, M., Risuleo, G., Eds.; AIP Publishing LLC: Melville, NY, USA, 2017. [Google Scholar]
- Farshori, N.N.; Al-Sheddi, E.S.; Al-Oqail, M.M.; Hassan, W.H.; Al-Khedhairy, A.A.; Musarrat, J.; Siddiqui, M.A. Hepatoprotective potential of Lavandula coronopifolia extracts against ethanol induced oxidative stress-mediated cytotoxicity in HepG2 cells. Toxicol. Ind. Health 2015, 31, 727–737. [Google Scholar] [CrossRef]
- Kim, T.-J.; Byun, J.-S.; Kwon, H.S.; Kim, D.-Y. Cellular toxicity driven by high-dose vitamin C on normal and cancer stem cells. Biochem. Biophys. Res. Commun. 2018, 497, 347–353. [Google Scholar] [CrossRef]
- Kumar, K.S.; Chu, F.-H.; Hsieh, H.-W.; Liao, J.-W.; Li, W.-H.; Lin, J.C.-C.; Shaw, J.-F.; Wang, S.-Y. Antroquinonol from ethanolic extract of mycelium of Antrodia cinnamomea protects hepatic cells from ethanol-induced oxidative stress through Nrf-2 activation. J. Ethnopharmacol. 2011, 136, 168–177. [Google Scholar] [CrossRef]
- Chaliha, M.; Sultanbawa, Y. Terminalia ferdinandiana, a traditional medicinal plant of Australia, alleviates hydrogen peroxide induced oxidative stress and inflammation, in vitro. J. Complementary Integr. Med. 2020, 17, 20190008. [Google Scholar] [CrossRef]
- Hur, J.M.; Park, J.C. Effects of the aerial parts of Orostachys japonicus and its bioactive component on hepatic alcohol-metabolizing enzyme system. J. Med. Food 2006, 9, 336–341. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System/Condition | Compound | ||
|---|---|---|---|
| Vitamin C | Phenolic Acids (Gallic Acid and Ellagic Acid) | Isoflavones (Daidzin) | |
| Flow rate | 1.0 mL/min | 1.0 mL/min | 0.8 mL/min |
| Wavelength | 254 nm | 260 nm | 254 nm |
| Injection Volume | 10 µL | 5 µL | 10 µL |
| Solvents | Gradient eluent: A: 0.1% Acetic acid in water B: 0.1% Acetic acid in methanol/water (90:10) | Gradient eluent: A: Methanol B: 0.1% Acetic acid in water | Gradient eluent: A: 0.1% Acetic acid in water B: Acetonitrile |
| Column Temperature | 25 °C | 35 °C | 35 °C |
| Samples | Vitamin C (L-Ascorbic Acid) (mg/g DW) | Gallic Acid (mg/g DW) | Ellagic Acid (mg/g DW) | Daidzin (mg/g DW) | Total Phenolics (mg GAE/g DW *) | Total Flavonoids (mg QE/g DW **) |
|---|---|---|---|---|---|---|
| KKD-NT | 88.66 ± 2.92 | 3.67 ± 0.16 | 0.11 ± 0.02 | 0.05 ± 0.01 | 147.2 ± 0.70 | 1.30 ± 0.01 |
| KKD-WA | 52.83 ± 1.85 | 5.10 ± 0.03 | 0.36 ± 0.01 | 0.04 ± 0.00 | 115.2 ± 2.12 | 1.10 ± 0.02 |
| Pomegranate | T | 0.13 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | 16.66 ± 1.32 | 0.32 ± 0.02 |
| Orange | 0.03 ± 0.00 | T | ND | ND | 27.01 ± 1.14 | 0.94 ± 0.03 |
| Lemon | 0.11 ± 0.00 | T | T | ND | 19.52 ± 0.84 | 0.66 ± 0.01 |
| Blueberry | T | T | ND | T | 21.96 ± 1.58 | 0.86 ± 0.06 |
| Raspberry | 0.01 ± 0.00 | T | 0.02 ± 0.00 | T | 25.79 ± 2.47 | 0.63 ± 0.03 |
| Standard | Solvent | Regression Equations | R2 | LOD (mg/mL) | LOQ (mg/mL) | Recovery (%) | RSD (%) |
|---|---|---|---|---|---|---|---|
| L-Ascorbic acid | 0.1% (v/v) 1 M HCL | y = 20,641x + 99 | 0.99998 | 2.01 | 6.71 | 99.13 | 0.24 |
| Gallic acid | Methanol | y = 14,948x + 113.86 | 0.99971 | 1.75 | 5.84 | 99.87 | 0.08 |
| Ellagic acid | y = 43,616x − 34.482 | 0.99999 | 0.57 | 1.90 | 99.35 | 0.12 | |
| Daidzin | y = 86,955x − 72.593 | 0.99981 | 0.39 | 1.31 | 98.50 | 0.45 |
| Samples | DPPH | Reducing Power | ||
|---|---|---|---|---|
| (mg GAE/g DW) | (mg AAE/g DW) | (mg GAE/g DW *) | (mg AAE/g DW **) | |
| KKD-NT KKD-WA Pomegranate | 239.7 ± 8.17 215.9 ± 16.2 2.19 ± 0.02 | 124.4 ± 4.34 111.1 ± 8.60 5.27 ± 0.04 | 179.6 ± 5.35 140.6 ± 7.80 7.22 ± 0.12 | 74.4 ± 2.06 60.5 ± 3.08 15.29 ± 0.23 |
| Orange | 1.16 ± 0.02 | 2.60 ± 0.04 | 8.75 ± 0.30 | 18.14 ± 0.56 |
| Lemon | 1.66 ± 0.08 | 2.47 ± 0.05 | 6.65 ± 0.44 | 14.08 ± 0.83 |
| Blueberry | 2.84 ± 0.01 | 6.86 ± 0.03 | 8.94 ± 0.33 | 18.86 ± 0.61 |
| Raspberry | 3.29 ± 0.03 | 8.13 ± 0.07 | 11.76 ± 0.20 | 23.79 ± 0.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akter, R.; Kwak, G.-Y.; Ahn, J.C.; Mathiyalagan, R.; Ramadhania, Z.M.; Yang, D.C.; Kang, S.C. Protective Effect and Potential Antioxidant Role of Kakadu Plum Extracts on Alcohol-Induced Oxidative Damage in HepG2 Cells. Appl. Sci. 2022, 12, 236. https://doi.org/10.3390/app12010236
Akter R, Kwak G-Y, Ahn JC, Mathiyalagan R, Ramadhania ZM, Yang DC, Kang SC. Protective Effect and Potential Antioxidant Role of Kakadu Plum Extracts on Alcohol-Induced Oxidative Damage in HepG2 Cells. Applied Sciences. 2022; 12(1):236. https://doi.org/10.3390/app12010236
Chicago/Turabian StyleAkter, Reshmi, Gi-Young Kwak, Jong Chan Ahn, Ramya Mathiyalagan, Zelika Mega Ramadhania, Deok Chun Yang, and Se Chan Kang. 2022. "Protective Effect and Potential Antioxidant Role of Kakadu Plum Extracts on Alcohol-Induced Oxidative Damage in HepG2 Cells" Applied Sciences 12, no. 1: 236. https://doi.org/10.3390/app12010236
APA StyleAkter, R., Kwak, G.-Y., Ahn, J. C., Mathiyalagan, R., Ramadhania, Z. M., Yang, D. C., & Kang, S. C. (2022). Protective Effect and Potential Antioxidant Role of Kakadu Plum Extracts on Alcohol-Induced Oxidative Damage in HepG2 Cells. Applied Sciences, 12(1), 236. https://doi.org/10.3390/app12010236