
Generation and Characterization of Novel Bioactive Peptides from Fish and Beef Hydrolysates



Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Extraction and Enzymatic Hydrolysis
2.2. Purification of Protein Hydrolysates
2.3. Mass Spectrometry and Amino Acid Sequencing
2.4. Sequence Analysis
2.5. Determination of Antimicrobial Activity
2.6. ACE Inhibition Activity Assay
2.7. Antioxidant Activity Assay
3. Results
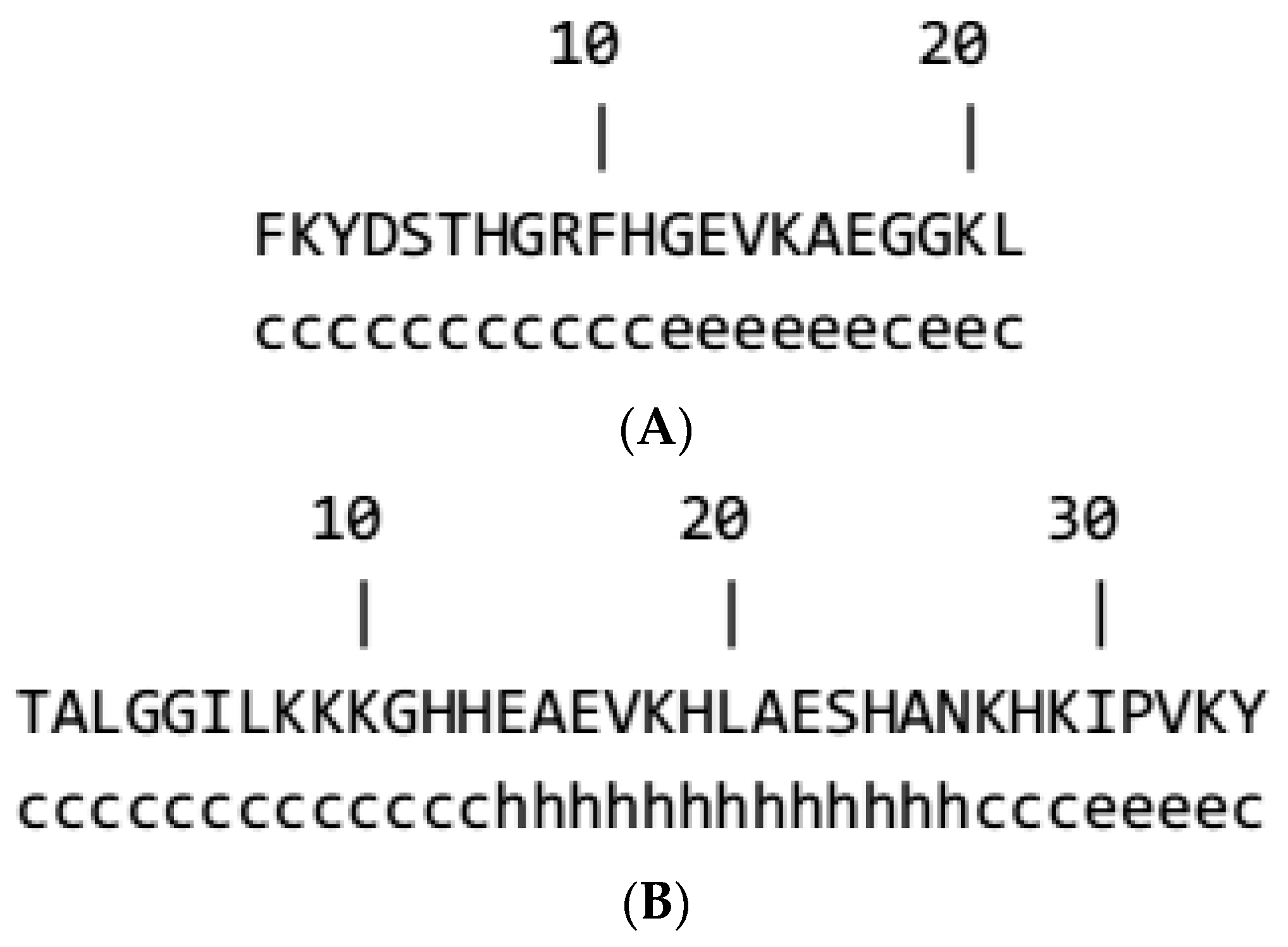
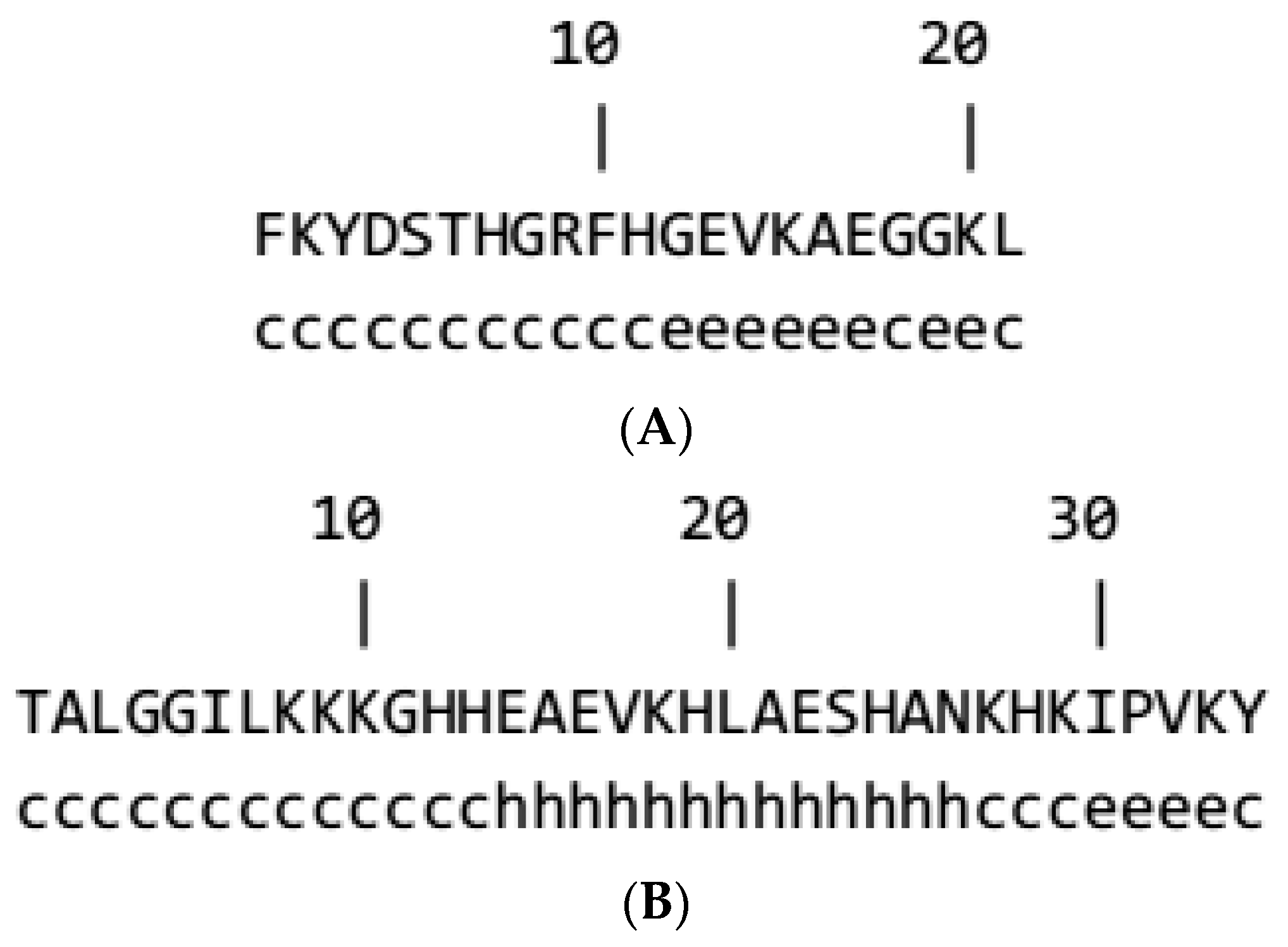
3.1. Generation, Purification, and Structural Analysis of Fish and Beef Peptides
3.2. Prediction of Secondary Structure
3.3. Antimicrobial Spectrum
3.4. ACE Inhibition and Antioxidant Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marqus, S.; Pirogova, E.; Piva, T.J. Evaluation of the use of therapeutic peptides for cancer treatment. Biomed. Sci. 2017, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halim, N.R.A.; Yusof, H.M.; Sarbon, N.M. Functional and bioactive properties of fish protein hydrolysates and peptides: A comprehensive review. Trends Food Sci. Technol. 2016, 51, 24–33. [Google Scholar] [CrossRef]
- Przybylski, R.; Firdaous, L.; Chataigne, G.; Dhulster, P.; Nedjar, N. Production of an antimicrobial peptide derived from slaughterhouse byproduct and its potential application on meat as preservative. Food Chem. 2016, 211, 306–313. [Google Scholar] [CrossRef]
- Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K.; Tomita, M. Identification of the bactericidal domain of lactoferrin. Biochim. Biophys. Acta 1992, 1121, 130–136. [Google Scholar] [CrossRef]
- McCann, K.B.; Shiell, B.J.; Michalski, W.P.; Lee, A.; Wan, J.; Roginsky, H.; Coventry, M.J. Isolation and characterization of antibacterial peptides derived from the f(164–207) region of bovine αS2-casein. Int. J. Dairy Sci. 2005, 15, 133–143. [Google Scholar] [CrossRef]
- Jones, F.S.; Simms, H.S. The bacterial growth inhibitor (lactenin) of milk. J. Exp. Med. 1929, 51, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Korhonen, H.; Pihlanto, A. Bioactive peptides from food proteins. In Handbook of Food Products Manufacturing; Hui, Y.H., Ed.; John Wiley and Sons Inc.: Hoboken, NJ, USA, 2007; pp. 5–37. [Google Scholar]
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I converting enzyme inhibitory peptides derived from phycobiliproteins of dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vercruysse, L.; Camp, J.V.; Smagghe, G. ACE inhibitory peptides derived from enzymatic hydrolysates of animal muscle protein: A review. J. Agric. Food Chem. 2005, 53, 8106–8115. [Google Scholar] [CrossRef]
- Maruyama, S.; Nakagomi, K.; Tomizuka, N.; Suzuki, H. Angiotensin I-converting enzyme inhibitor derived from an enzymatic hydrolysate of casein. Agric. Biol. Chem. 1985, 49, 1405–1409. [Google Scholar] [CrossRef]
- Okamoto, A.; Hanagata, H.; Kawamura, Y.; Yanagida, F. Anti-hypertensive substances in fermented soybean, natto. Plant Foods Hum. Nutr. 1995, 47, 39–47. [Google Scholar] [CrossRef]
- Arihara, K.; Nakashima, Y.; Mukai, T.; Ishikawa, S.; Itoh, M. Peptide inhibitors for angiotensin I-converting enzyme from enzymatic hydrolysates of porcine skeletal muscle proteins. Meat Sci. 2001, 57, 319–324. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imaida, K.; Fukushima, S.; Shirai, T.; Ohtani, M.; Nakanishi, K.; Ito, N. Promoting activities of butylated hydroxyanisole and butylated hydroxytoluene on 2-stage urinary carcinogenesis and inhibition of gamma-glutamyl transpeptidase-positivefor development in the liver of rats. Carcinogenesis 1983, 4, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Sohaib, M.; Anjum, F.M.; Sahar, A.; Arshad, M.S.; Rahman, U.U.; Imran, A.; Hussain, S. Antioxidant proteins and peptides to enhance the oxidative stability of meat and meat products: A comprehensive review. Int. J. Food Prop. 2017, 20, 2581–2593. [Google Scholar] [CrossRef]
- Jang, A.; Lee, M. Purification and identification of angiotensin converting enzyme inhibitory peptides from beef hydrolysates. Meat Sci. 2005, 69, 653–661. [Google Scholar] [CrossRef]
- Zendo, T.; Nakayama, J.; Fujita, K.; Sonomoto, K. Bacteriocin detection by liquid chromatography/mass spectrometry for rapid identification. J. Appl. Microbiol. 2008, 104, 499–507. [Google Scholar] [CrossRef]
- Edman, P. Method for determination of the amino acid sequence in peptides. Acta Chem. Scand. 1950, 4, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Maky, M.A.; Ishibashi, N.; Nakayama, J.; Zendo, T. Characterization of the biosynthetic gene cluster of enterocin F4-9, a glycosylated bacteriocin. Microorganisms 2021, 9, 2276. [Google Scholar] [CrossRef]
- Abuine, R.; Rathnayake, A.U.; Byun, H. Biological activity of peptides purified from fish skin hydrolysates. Fish. Aquat. Sci. 2019, 22, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.; Lee, M.J.; Go, H.; Park, T.H.; Park, N.G. Purification and characterization of YFGAP, a GAPDH-related novel antimicrobial peptide, from the skin of yellow fin tuna, Thunnus albacares. Fish Shellfish Immunol. 2012, 33, 743–752. [Google Scholar] [CrossRef]
- Seo, J.; Lee, M.J.; Go, H.; Kim, Y.J.; Park, N.G. Antimicrobial function of the GAPDH related antimicrobial peptide in the skin of skipjack tuna, Katsuwonus pelamis. Fish Shellfish Immunol. 2014, 36, 571–581. [Google Scholar] [CrossRef]
- Keska, P.; Wojciak, K.M.; Stadnik, J. Bioactive peptides from beef products fermented by acid whey—In vitro and in silico study. Sci. Agric. 2019, 76, 311–320. [Google Scholar] [CrossRef]
- Ryan, J.T.; Ross, R.P.; Bolton, D.; Fitzgerald, G.F.; Stanton, C. Bioactive peptides from muscle Sources: Meat and fish. Nutrients 2011, 3, 765–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, L.; Liu, R.; Cao, S.; Zhang, W.; Guanghong, Z. Meat protein based bioactive peptides and their potential functional activity: A review. Int. J. Food Sci. 2019, 54, 1956–1966. [Google Scholar] [CrossRef] [Green Version]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shai, Y. Mechanism of the binding, insertion and destabilization of phospholipid bilayer membranes by alpha-helical antimicrobial and cell non-selective membrane-lytic peptides. Biochim. Biophys. Acta 1999, 1462, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Mahmoodani, F.; Ghassem, M.; Babji, A.S.; Yusop, S.M.; Khosrokhavar, R. ACE inhibitory activity of pangasius catfish (Pangasiussutchi) skin and bone gelatin hydrolysate. J. Food Sci. Technol. 2014, 51, 1847–1856. [Google Scholar] [CrossRef] [Green Version]
- Rezaharsamto, B.; Subroto, E. A review on bioactive peptides derived from various sources of meat and meat by-products. Int. J. Sci. Technol. Res. 2019, 8, 3151–3156. [Google Scholar]
- Jang, A.; Jo, C.; Kang, K.S.; Lee, M. Antimicrobial and human cancer cell cytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory peptides. Food Chem. 2008, 107, 327–336. [Google Scholar] [CrossRef]
- Keska, P.; Stadnik, J. Antimicrobial peptides of meat origin—An in silico and in vitro analysis. Protein Pept. Lett. 2017, 24, 165–173. [Google Scholar] [CrossRef]
- Katayama, K.; Mori, T.; Kawahara, S.; Miake, K.; Kodama, Y.; Sugiyama, M.; Kawamura, Y.; Nakayama, T.; Maruyama, M.; Muguruma, M. Angiotensin-I converting enzyme inhibitory peptide derived from porcine skeletal muscle myosin and its antihypertensive activity in spontaneously hypertensive rats. J. Food Sci. 2007, 72, S702–S706. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Hur, S.J. Purification of novel angiotensin converting enzyme inhibitory peptides from beef myofibrillar proteins and analysis of their effect in spontaneously hypertensive rat model. Biomed. Pharmacother. 2019, 116, 1–7. [Google Scholar] [CrossRef]
- Wang, R.; Lu, X.; Sun, Q.; Gao, J.; Ma, L.; Huang, J. Novel ACE inhibitory peptides derived from simulated gastrointestinal digestion in vitro of Sesame (Sesamumindicum L.) protein and molecular docking study. Int. J. Mol. Sci. 2020, 21, 1059. [Google Scholar] [CrossRef] [Green Version]
- Wijesekara, I.; Kim, S. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [Green Version]
- Jakubczyk, A.; Karaś, M.; Rybczyńska-Tkaczyk, K.; Zielińska, E.; Zieliński, D. Current trends of bioactive peptides—New sources and therapeutic effect. Foods 2020, 9, 846. [Google Scholar] [CrossRef]
- Yang, P.; Ke, H.; Hong, P.; Zeng, S.; Cao, W. Antioxidant activity of bigeye tuna (Thunnus obesus) head protein hydrolysate prepared with alcalase. Int. J. Food Sci. 2011, 46, 2460–2466. [Google Scholar] [CrossRef]
- Lassoued, I.; Mora, L.; Nasri, R.; Jridi, M.; Toldra, F.; Aristoy, M.; Barkia, A.; Nasri, M. Characterization and comparative assessment of antioxidant and ACE inhibitory activities of thornback ray gelatin hydrolysates. J. Funct. Foods 2015, 13, 225–238. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Indicator Strain | MIC (μg/mL) | |||
|---|---|---|---|---|
| Fish Hydrolysates | F21 | Beef Hydrolysates | B34 | |
| Enterococcus faecalis JCM 5803T | 800 | 46 | 800 | NA3 |
| Listeria innocua ATCC 33090T | 400 | 23 | 400 | NA3 |
| Escherichia coli JM109 | 400 | 46 | 400 | 184 |
| Bacillus coagulans JCM 2257T | 400 | 23 | 400 | NA3 |
| Pseudomonas putida ATCC 12633T | 800 | 23 | NA2 | NA3 |
| Salmonella enterica subsp. enterica NBRC 13245T | 400 | 184 | NA2 | NA3 |
| Proteus vulgaris F24B | 400 | NA1 | 800 | 91.6 |
| Sample | IC50 (µg/mL) | ||
|---|---|---|---|
| ACE | Antioxidant | TEAC | |
| Fish hydrolysates | 304.5 | 470.4 | 0.14 |
| F21 | 7.3 | 389.9 | 0.17 |
| Beef hydrolysates | 265.6 | 531.0 | 0.12 |
| B34 | 5.8 | negative | negative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maky, M.A.; Zendo, T. Generation and Characterization of Novel Bioactive Peptides from Fish and Beef Hydrolysates. Appl. Sci. 2021, 11, 10452. https://doi.org/10.3390/app112110452
Maky MA, Zendo T. Generation and Characterization of Novel Bioactive Peptides from Fish and Beef Hydrolysates. Applied Sciences. 2021; 11(21):10452. https://doi.org/10.3390/app112110452
Chicago/Turabian StyleMaky, Mohamed Abdelfattah, and Takeshi Zendo. 2021. "Generation and Characterization of Novel Bioactive Peptides from Fish and Beef Hydrolysates" Applied Sciences 11, no. 21: 10452. https://doi.org/10.3390/app112110452
APA StyleMaky, M. A., & Zendo, T. (2021). Generation and Characterization of Novel Bioactive Peptides from Fish and Beef Hydrolysates. Applied Sciences, 11(21), 10452. https://doi.org/10.3390/app112110452