Volatile Fatty Acids in Ruminal Fluid Can Be Used to Predict Methane Yield of Dairy Cows


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Model Development
2.2. Data
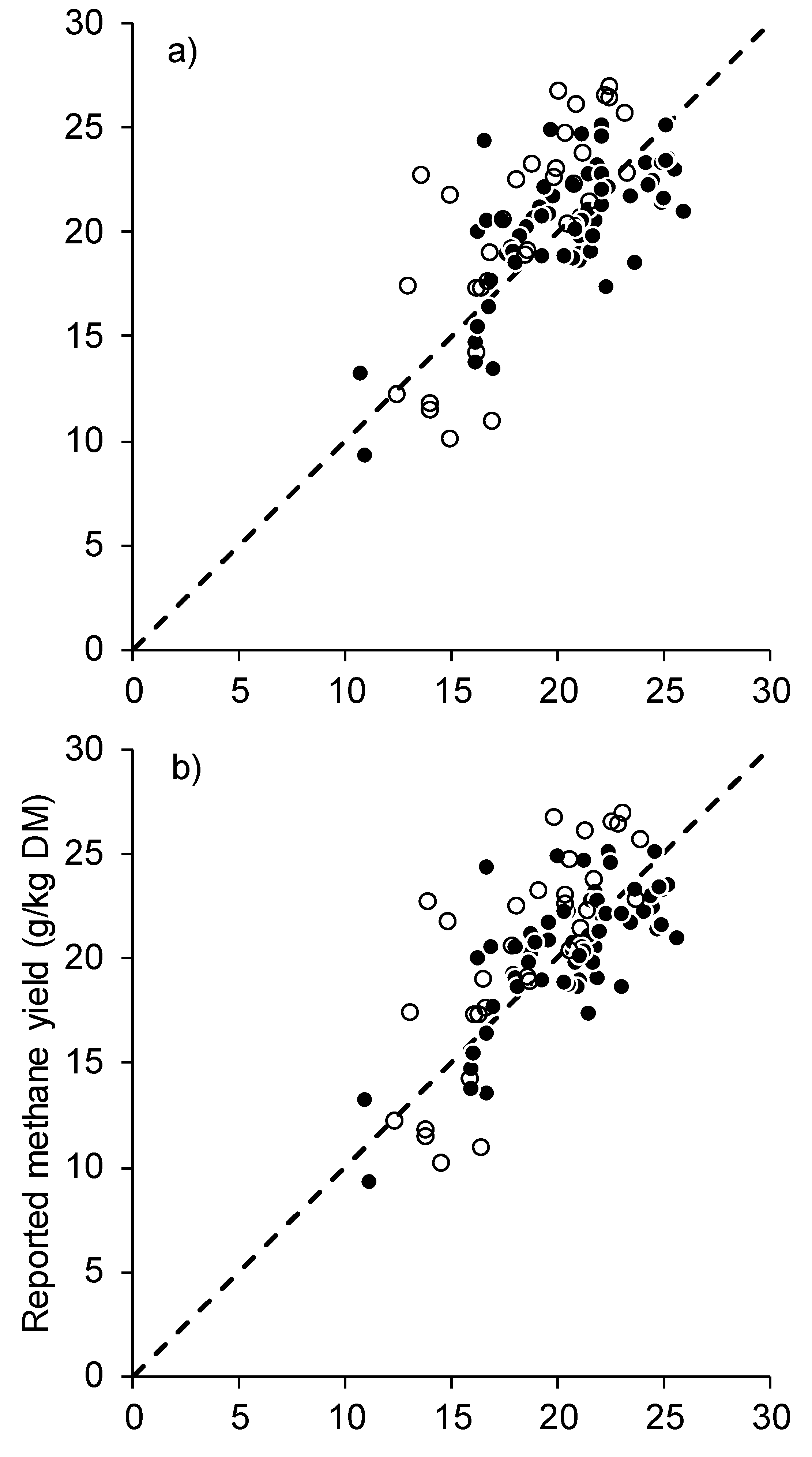
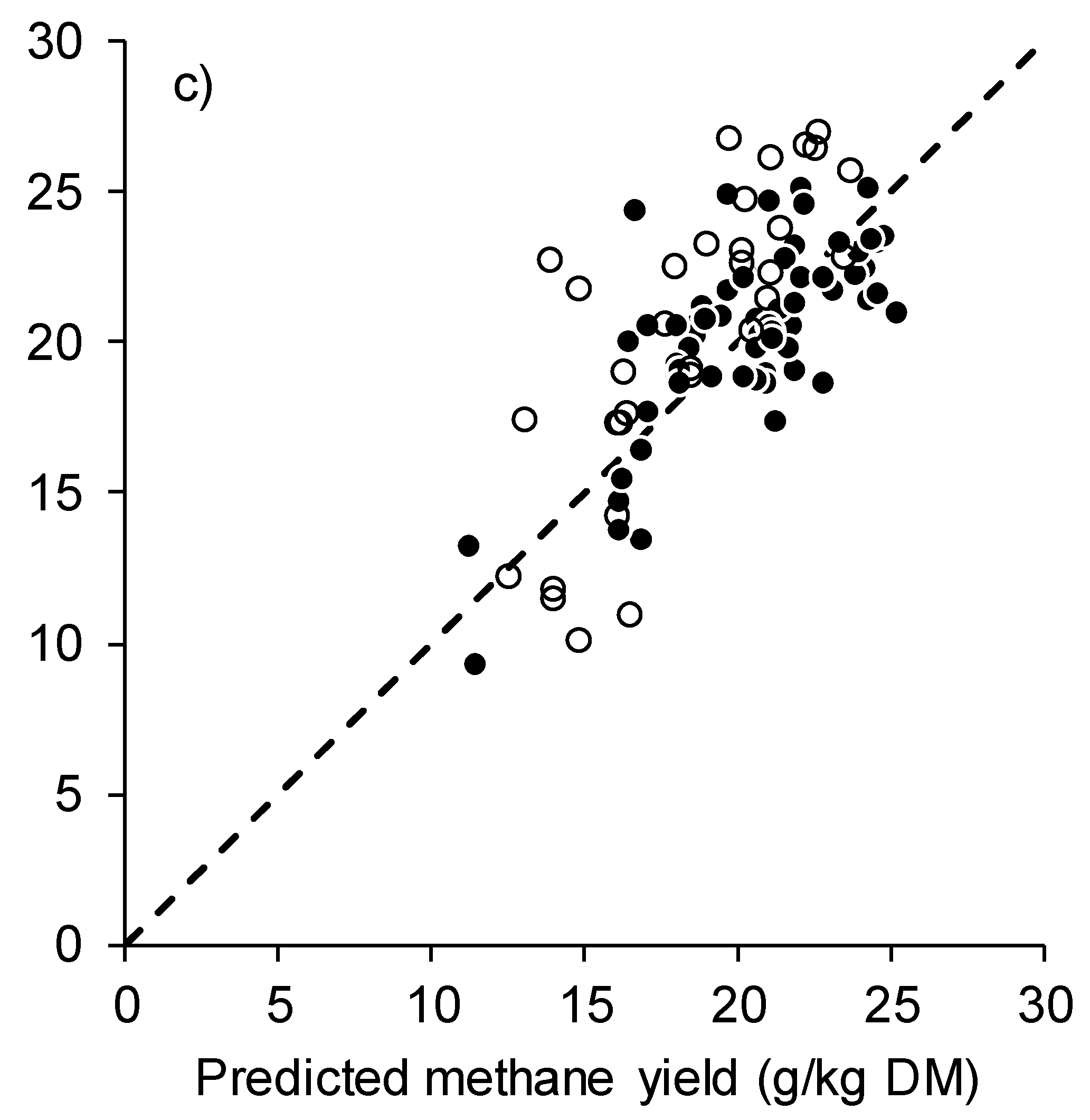
2.3. Statistical Analysis and Model Evaluation
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hammond, K.J.; Humphries, D.J.; Crompton, L.A.; Green, C.; Reynolds, C.K. Methane emissions from cattle: Estimates from short-term measurements using a GreenFeed system compared with measurements obtained using respiration chambers or sulphur hexafluoride tracer. Anim. Feed Sci. Technol. 2015, 203, 41–52. [Google Scholar] [CrossRef]
- Hegarty, R.S. Applicability of short-term emission measurements for on-farm quantification of enteric methane. Animal 2013, 7, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Negussie, E.; de Haas, Y.; Dehareng, F.; Dewhurst, R.J.; Dijkstra, J.; Gengler, N.; Morgavi, D.P.; Soyeurt, H.; van Gastelen, S.; Yan, T.; et al. Invited review: Large-scale indirect measurements for enteric methane emissions in dairy cattle: A review of proxies and their potential for use in management and breeding decisions. J. Dairy Sci. 2017, 100, 2433–2453. [Google Scholar] [CrossRef] [PubMed]
- Charmley, E.; Williams, S.R.O.; Moate, P.J.; Hegarty, R.S.; Herd, R.M.; Oddy, V.H.; Reyenga, P.; Staunton, K.M.; Anderson, A.; Hannah, M.C. A universal equation to predict methane production of forage-fed cattle in Australia. Anim. Prod. Sci. 2016, 56, 169–180. [Google Scholar] [CrossRef]
- Moate, P.J.; Deighton, M.H.; Williams, S.R.O.; Pryce, J.E.; Hayes, B.J.; Jacobs, J.L.; Eckard, R.J.; Hannah, M.C.; Wales, W.J. Reducing the carbon footprint of Australian milk production by mitigation of enteric methane emissions. Anim. Prod. Sci. 2016, 56, 1017–1034. [Google Scholar] [CrossRef]
- Moate, P.J.; Williams, S.R.O.; Jacobs, J.L.; Hannah, M.C.; Beauchemin, K.A.; Eckard, R.J.; Wales, W.J. Wheat is more potent than corn or barley for dietary mitigation of enteric methane emissions from dairy cows. J. Dairy Sci. 2017, 100, 7139–7153. [Google Scholar] [CrossRef]
- Sauvant, D.; Giger-Reverdin, S.; Serment, A.; Broudiscou, L. Influences des régimes et de leur fermentation dans le rumen sur la production de méthane par les ruminants. INRA Prod. Anim. 2011, 24, 433–446. [Google Scholar]
- Niu, M.; Kebreab, E.; Hristov, A.N.; Oh, J.; Arndt, C.; Bannink, A.; Bayat, A.R.; Brito, A.F.; Boland, T.; Casper, D.; et al. Prediction of enteric methane production, yield, and intensity in dairy cattle using an intercontinental database. Glob. Chang. Biol. 2018, 24, 3368–3389. [Google Scholar] [CrossRef]
- Chilliard, Y.; Martin, C.; Rouel, J.; Doreau, M. Milk fatty acids in dairy cows fed whole crude linseed, extruded linseed, or linseed oil, and their relationship with methane output. J. Dairy Sci. 2009, 92, 5199–5211. [Google Scholar] [CrossRef]
- Dijkstra, J.; van Zijderveld, S.M.; Apajalahti, J.A.; Bannink, A.; Gerrits, W.J.J.; Newbold, J.R.; Perdok, H.B.; Berends, H. Relationships between methane production and milk fatty acid profiles in dairy cattle. Anim. Feed Sci. Technol. 2011, 166–167, 590–595. [Google Scholar] [CrossRef]
- Williams, S.R.O.; Moate, P.J.; Deighton, M.H.; Hannah, M.C.; Wales, W.J. Methane emissions of dairy cows cannot be predicted by the concentrations of C8:0 and total C18 fatty acids in milk. Anim. Prod. Sci. 2014, 54, 1757–1761. [Google Scholar] [CrossRef]
- Demeyer, D. Quantitative aspects of microbial metabolism in the rumen and hind gut. In Rumen Microbial Metabolism and Ruminant Digestion; Jouany, J.P., Ed.; INRA Editions: Paris, France, 1991; pp. 217–237. [Google Scholar]
- Moss, A.R.; Jouany, J.-P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Ann. Zoot. 2000, 49, 231–253. [Google Scholar] [CrossRef]
- Holtshausen, L.; Chaves, A.V.; Beauchemin, K.A.; McGinn, S.M.; McAllister, T.; Odongo, N. Feeding saponin-containing Yucca schidigera and Quillaja saponaria to decrease enteric methane production in dairy cows. J. Dairy Sci. 2009, 92, 2809–2821. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, N.; Ajisaka, N.; Lila, Z.A.; Hara, K.; Mikuni, K.; Kanda, S.; Itabashi, H. Effect of Japanese horseradish oil on methane production and ruminal fermentation in vitro and in steers. J. Anim. Sci. 2004, 82, 1839–1846. [Google Scholar] [CrossRef] [PubMed]
- Eugène, M.; Martin, C.; Mialon, M.M.; Krauss, D.; Renand, G.; Doreau, M. Dietary linseed and starch supplementation decreases methane production of fattening bulls. Anim. Feed Sci. Technol. 2011, 166–167, 330–337. [Google Scholar] [CrossRef]
- Danielsson, R.; Schnürer, A.; Arthurson, V.; Bertilsson, J. Methanogenic population and CH4 production in Swedish dairy cows fed different levels of forage. Appl. Environ. Microbiol. 2012, 78, 6172–6179. [Google Scholar] [CrossRef] [PubMed]
- Foley, P.A.; Kenny, D.A.; Callan, J.J.; Boland, T.M.; O’Mara, F.P. Effect of DL-malic acid supplementation on feed intake, methane emission, and rumen fermentation in beef cattle. J. Anim. Sci. 2009, 87, 1048–1057. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, R.; McGinn, S.M.; Beauchemin, K.A. Prediction of enteric methane output from milk fatty acid concentrations and rumen fermentation parameters in dairy cows fed sunflower, flax, or canola seeds. J. Dairy Sci. 2011, 94, 6057–6068. [Google Scholar] [CrossRef]
- NHMRC. Australian Code for the Care and Use of Animals for Scientific Purposes, 7th ed.; National Health and Medical Research Council: Canberra, Australia, 2004.
- NHMRC. Australian Code for the Care and Use of Animals for Scientific Purposes, 8th ed.; National Health and Medical Research Council: Canberra, Australia, 2013.
- Bannink, A.; France, J.; Lopez, S.; Gerrits, W.J.J.; Kebreab, E.; Tamminga, S.; Dijkstra, J. Modelling the implications of feeding strategy on rumen fermentation and functioning of the rumen wall. Anim. Feed Sci. Technol. 2008, 143, 3–26. [Google Scholar] [CrossRef]
- Robinson, D.L.; Goopy, J.; Hegarty, R.S. Can rumen methane production be predicted from volatile fatty acid concentrations? Anim. Prod. Sci. 2010, 50, 630–636. [Google Scholar] [CrossRef]
- Williams, S.R.O.; Chaves, A.V.; Deighton, M.H.; Jacobs, J.L.; Hannah, M.C.; Ribaux, B.E.; Morris, G.L.; Wales, W.J.; Moate, P.J. Influence of feeding supplements of almond hulls and ensiled citrus pulp on the milk production, milk composition, and methane emissions of dairy cows. J. Dairy Sci. 2018, 101, 2072–2083. [Google Scholar] [CrossRef] [PubMed]
- Moate, P.J.; Williams, S.R.O.; Hannah, M.C.; Eckard, R.J.; Auldist, M.J.; Ribaux, B.E.; Jacobs, J.L.; Wales, W.J. Effects of feeding algal meal high in docosahexaenoic acid on feed intake, milk production, and methane emissions in dairy cows. J. Dairy Sci. 2013, 96, 3177–3188. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.R.O.; Hannah, M.C.; Eckard, R.J.; Wales, W.J.; Moate, P.J. Supplementing the diet of dairy cows with fat or tannin reduces methane yield, and additively when fed in combination. Animal. Under review.
- Williams, S.R.O.; Moate, P.J.; Deighton, M.H.; Hannah, M.C.; Wales, W.J.; Jacobs, J.L. Milk production and composition, and methane emissions from dairy cows fed lucerne hay with forage brassica or chicory. Anim. Prod. Sci. 2016, 56, 304–311. [Google Scholar] [CrossRef]
- Moate, P.J.; Williams, S.R.O.; Deighton, M.H.; Hannah, M.C.; Ribaux, B.E.; Morris, G.L.; Jacobs, J.L.; Hill, J.; Wales, W.J. Effects of feeding wheat or corn and of rumen fistulation on milk production and methane emissions of dairy cows. Anim. Prod. Sci. 2019, 59, 891–905. [Google Scholar] [CrossRef]
- Moate, P.J.; Deighton, M.H.; Jacobs, J.L.; Ribaux, B.E.; Morris, G.L.; Hannah, M.C.; Mapleson, D.; Islam, M.S.; Wales, W.J.; Williams, S.R.O. Influence of proportion of wheat in a pasture-based diet, on milk yield, methane emissions, methane yield and ruminal protozoa of dairy cows. J. Dairy Sci. Under review.
- Moate, P.J.; (Agriculture Victoria Research, Ellinbank, Victoria, Australia); Williams, S.R.O.; (Agriculture Victoria Research, Ellinbank, Victoria, Australia). Personal communication, 2018.
- Tyrrell, H.F.; Reid, J.T. Prediction of the Energy Value of Cow’s Milk. J. Dairy Sci. 1965, 48, 1215–1223. [Google Scholar] [CrossRef]
- Williams, S.R.O.; Clarke, T.; Hannah, M.C.; Marett, L.C.; Moate, P.J.; Auldist, M.J.; Wales, W.J. Energy partitioning in herbage-fed dairy cows offered supplementary grain during an extended lactation. J. Dairy Sci. 2013, 96, 484–494. [Google Scholar] [CrossRef]
- Deighton, M.H.; Williams, S.R.O.; Hannah, M.C.; Eckard, R.J.; Boland, T.M.; Wales, W.J.; Moate, P.J. A modified sulphur hexafluoride tracer technique enables accurate determination of enteric methane emissions from ruminants. Anim. Feed Sci. Technol. 2014, 197, 47–63. [Google Scholar] [CrossRef]
- Geishauser, T. An instrument for collection and transfer of ruminal fluid and for administration of water soluble drugs in adult cattle. Bov. Pract. 1993, 27, 38–42. [Google Scholar]
- Moate, P.J.; Williams, S.R.O.; Torok, V.; Hannah, M.C.; Ribaux, B.E.; Tavendale, M.; Eckard, R.J.; Jacobs, J.L.; Auldist, M.J.; Wales, W.J. Grape marc reduces methane emissions when fed to dairy cows. J. Dairy Sci. 2014, 97, 5073–5087. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; McGinn, S.M. Methane emissions from feedlot cattle fed barley or corn diets. J. Anim. Sci. 2005, 83, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; McGinn, S.M. Methane emissions from beef cattle: Effects of fumaric acid, essential oil, and canola oil. J. Anim. Sci. 2006, 84, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; McGinn, S.M.; Martinez, T.F.; McAllister, T.A. Use of condensed tannin extract from quebracho trees to reduce methane emissions from cattle. J. Anim. Sci. 2007, 85, 1990–1996. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; McGinn, S.M.; Benchaar, C.; Holtshausen, L. Crushed sunflower, flax, or canola seeds in lactating dairy cow diets: Effects on methane production, rumen fermentation, and milk production. J. Dairy Sci. 2009, 92, 2118–2127. [Google Scholar] [CrossRef]
- Benchaar, C.; Hassanat, F.; Gervais, R.; Chouinard, P.Y.; Julien, C.; Petit, H.V.; Massé, D.I. Effects of increasing amounts of corn dried distillers grains with solubles in dairy cow diets on methane production, ruminal fermentation, digestion, N balance, and milk production. J. Dairy Sci. 2013, 96, 2413–2427. [Google Scholar] [CrossRef]
- Bougouin, A.; Ferlay, A.; Doreau, M.; Martin, C. Effects of carbohydrate type or bicarbonate addition to grass silage-based diets on enteric methane emissions and milk fatty acid composition in dairy cows. J. Dairy Sci. 2018, 101, 6085–6097. [Google Scholar] [CrossRef]
- Bougouin, A.; Martin, C.; Doreau, M.; Ferlay, A. Effects of starch-rich or lipid-supplemented diets that induce milk fat depression on rumen biohydrogenation of fatty acids and methanogenesis in lactating dairy cows. Animal 2018, 13, 1421–1431. [Google Scholar] [CrossRef]
- Guyader, J.; Eugène, M.; Meunier, B.; Doreau, M.; Morgavi, D.P.; Silberberg, M.; Rochette, Y.; Gerard, C.; Loncke, C.; Martin, C. Additive methane-mitigating effect between linseed oil and nitrate fed to cattle. J. Anim. Sci. 2015, 93, 3564–3577. [Google Scholar] [CrossRef]
- Hassanat, F.; Gervais, R.; Massé, D.I.; Petit, H.V.; Benchaar, C. Methane production, nutrient digestion, ruminal fermentation, N balance, and milk production of cows fed timothy silage- or alfalfa silage-based diets. J. Dairy Sci. 2014, 97, 6463–6474. [Google Scholar] [CrossRef]
- Hatew, B.; Podesta, S.C.; Van Laar, H.; Pellikaan, W.F.; Ellis, J.L.; Dijkstra, J.; Bannink, A. Effects of dietary starch content and rate of fermentation on methane production in lactating dairy cows. J. Dairy Sci. 2015, 98, 486–499. [Google Scholar] [CrossRef]
- Klop, G.; Dijkstra, J.; Dieho, K.; Hendriks, W.H.; Bannink, A. Enteric methane production in lactating dairy cows with continuous feeding of essential oils or rotational feeding of essential oils and lauric acid. J. Dairy Sci. 2017, 100, 3563–3575. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Fernandez, G.; Duval, S.; Kindermann, M.; Schirra, H.J.; Denman, S.E.; McSweeney, C.S. 3-NOP vs. halogenated compound: Methane production, ruminal fermentation and microbial community response in forage fed cattle. Front. Microbiol. 2018, 9, 1582. [Google Scholar] [CrossRef] [PubMed]
- McGinn, S.M.; Beauchemin, K.A.; Coates, T.; Colombatto, D. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. J. Anim. Sci. 2004, 82, 3346–3356. [Google Scholar] [CrossRef] [PubMed]
- Olijhoek, D.W.; Løvendahl, P.; Lassen, J.; Hellwing, A.L.F.; Höglund, J.K.; Weisbjerg, M.R.; Noel, S.J.; McLean, F.; Højberg, O.; Lund, P. Methane production, rumen fermentation, and diet digestibility of Holstein and Jersey dairy cows being divergent in residual feed intake and fed at 2 forage-to-concentrate ratios. J. Dairy Sci. 2018, 101, 9926–9940. [Google Scholar] [CrossRef]
- Van Gastelen, S.; Antunes-Fernandes, E.C.; Hettinga, K.A.; Klop, G.; Alferink, S.J.J.; Hendriks, W.H.; Dijkstra, J. Enteric methane production, rumen volatile fatty acid concentrations, and milk fatty acid composition in lactating Holstein-Friesian cows fed grass silage- or corn silage-based diets. J. Dairy Sci. 2015, 98, 1915–1927. [Google Scholar] [CrossRef]
- Van Zijderveld, S.M.; Fonken, B.; Dijkstra, J.; Gerrits, W.J.J.; Perdok, H.B.; Fokkink, W.; Newbold, J.R. Effects of a combination of feed additives on methane production, diet digestibility, and animal performance in lactating dairy cows. J. Dairy Sci. 2011, 94, 1445–1454. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning; Springer: Berlin, Germany, 2012. [Google Scholar]
- Stevens, A.; Miralles, I.; Van Wesemael, B. Soil organic carbon predictions by airborne imaging spectroscopy: Comparing cross-validation and validation. Soil Sci. Soc. Am. J. 2012, 76, 2174–2183. [Google Scholar] [CrossRef]
- Lin, L.K. A concordance correlation coefficient to evaluate reproducibility. Biometrics 1989, 45, 255–268. [Google Scholar] [CrossRef]
- Bibby, J.; Toutenburg, H. Prediction and Improved Estimation in Linear Models; J. Wiley & Sons: Chichester, NY, USA, 1977. [Google Scholar]
- Gauch, H.G., Jr.; Hwang, J.T.G.; Fick, G.W. Model Evaluation by Comparison of Model-Based Predictions and Measured Values. Agron. J. 2003, 95, 1442–1446. [Google Scholar] [CrossRef]
- Lopes, J.C.; de Matos, L.F.; Harper, M.T.; Giallongo, F.; Oh, J.; Gruen, D.; Ono, S.; Kindermann, M.; Duval, S.; Hristov, A.N. Effect of 3-nitrooxipropanol on methane and hydrogen emissions, methane isotopic signature, and ruminal fermentation in dairy cows. J. Dairy Sci. 2016, 99, 5335–5344. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, P.J. Nutritional Ecology of the Ruminant; Cornell University Press: Ithaca, NY, USA, 1982; p. 373. [Google Scholar]
- Herd, R.; Bird, S.; Donoghue, K.; Arthur, P.; Hegarty, R. Phenotypic associations between methane production traits, volatile fatty acids and animal breeding traits. In Proceedings of the 20th conference of the Association for the Advancement of Animal Breeding and Genetics, Napier, New Zealand, 20–23 October 2013; pp. 286–289. [Google Scholar]
- Smith, H.C.; Herd, R.M.; Donoghue, K.; Bird-Gardiner, T.; Arthur, P.F.; Hegarty, R.F. Associations of rumen volatile fatty acids with phenotypic and genetic variation in methane production traits in Angus cattle. Proc. Assoc. Adv. Anim. Breed. Genet. 2015, 21, 93–96. [Google Scholar]
- Alemu, A.W.; Dijkstra, J.; Bannink, A.; France, J.; Kebreab, E. Rumen stoichiometric models and their contribution and challenges in predicting enteric methane production. Anim. Feed Sci. Technol. 2011, 166–167, 761–778. [Google Scholar] [CrossRef]
- Hoffmann, E.M.; Selje-Assmann, N.; Becker, K. Dose studies on anti-proteolytic effects of a methanol extract from Knautia arvensis on in vitro ruminal fermentation. Anim. Feed Sci. Technol. 2008, 145, 285–301. [Google Scholar] [CrossRef]
- Bentley, O.G.; Johnson, R.R.; Hershberger, T.V.; Cline, J.H.; Moxon, A.L. Cellulolytic-factor activity of certain short-chain fatty acids for rumen microorganisms in vitro. J. Nutr. 1955, 57, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Stewart, W.E.; Stewart, D.G.; Schultz, L.H. Rates of volatile fatty acid production in the bovine rumen. J. Anim. Sci. 1958, 17, 723–736. [Google Scholar] [CrossRef]
- Lee, M.R.F.; Merry, R.J.; Davies, D.R.; Moorby, J.M.; Humphreys, M.O.; Theodorou, M.K.; MacRae, J.C.; Scollan, N.D. Effect of increasing availability of water-soluble carbohydrates on in vitro rumen fermentation. Anim. Feed Sci. Technol. 2003, 104, 59–70. [Google Scholar] [CrossRef]
- Bryant, M.P. Microbial methane production—Theoretical aspects. J. Anim. Sci. 1979, 48, 193–201. [Google Scholar] [CrossRef]
- Hristov, A.N.; Kebreab, E.; Niu, M.; Oh, J.; Bannink, A.; Bayat, A.R.; Boland, T.B.; Brito, A.F.; Casper, D.P.; Crompton, L.A.; et al. Symposium review: Uncertainties in enteric methane inventories, measurement techniques, and prediction models. J. Dairy Sci. 2018, 101, 6655–6674. [Google Scholar] [CrossRef]
- Moate, P.J.; Deighton, M.H.; Hannah, M.C.; Williams, S.R.O. Intake effects on methane emissions from dairy cows. In Proceedings of the CCRSPI Conference, Melbourne, Australia, 27–29 November 2012; pp. 60–61. [Google Scholar]
- Ramin, M.; Huhtanen, P. Development of equations for predicting methane emissions from ruminants. J. Dairy Sci. 2013, 96, 2476–2493. [Google Scholar] [CrossRef]
- Moate, P.J.; Jacobs, J.L.; Hannah, M.C.; Morris, G.L.; Beauchemin, K.A.; Alvarez Hess, P.S.; Eckard, R.J.; Liu, Z.; Rochfort, S.; Wales, W.J.; et al. Adaptation responses in milk fat yield and methane emissions of dairy cows when wheat was included in their diet for 16 weeks. J. Dairy Sci. 2018, 101, 7117–7132. [Google Scholar] [CrossRef] [PubMed]
- Reeves, M.; Fulkerson, W.J.; Kellaway, R.C.; Dove, H. A comparison of three techniques to determine the herbage intake of dairy cows grazing kikuyu (Pennisetum clandestinum) pasture. Aust. J. Exp. Agric. 1996, 36, 23–30. [Google Scholar] [CrossRef]
- Swainson, N.; Muetzel, S.; Clark, H. Updated predictions of enteric methane emissions from sheep suitable for use in the New Zealand national greenhouse gas inventory. Anim. Prod. Sci. 2018, 58, 973–979. [Google Scholar] [CrossRef]
- Li, X.; Norman, H.C.; Kinley, R.D.; Laurence, M.; Wilmot, M.; Bender, H.; De Nys, R.; Tomkins, N. Asparagopsis taxiformis decreases enteric methane production from sheep. Anim. Prod. Sci. 2018, 58, 681–688. [Google Scholar] [CrossRef]
- van Zijderveld, S.M.; Gerrits, W.J.J.; Apajalahti, J.A.; Newbold, J.R.; Dijkstra, J.; Leng, R.A.; Perdok, H.B. Nitrate and sulfate: Effective alternative hydrogen sinks for mitigation of ruminal methane production in sheep. J. Dairy Sci. 2010, 93, 5856–5866. [Google Scholar] [CrossRef]
- Reynolds, C.K.; Humphries, D.J.; Kirton, P.; Kindermann, M.; Duval, S.; Steinberg, W. Effects of 3-nitrooxypropanol on methane emission, digestion, and energy and nitrogen balance of lactating dairy cows. J. Dairy Sci. 2014, 97, 3777–3789. [Google Scholar] [CrossRef]
- Kinley, R.D.; De Nys, R.; Vucko, M.J.; MacHado, L.; Tomkins, N.W. The red macroalgae Asparagopsis taxiformis is a potent natural antimethanogenic that reduces methane production during in vitro fermentation with rumen fluid. Anim. Prod. Sci. 2016, 56, 282–289. [Google Scholar] [CrossRef]
- Oertel, P.; Küntzel, A.; Reinhold, P.; Köhler, H.; Schubert, J.K.; Kolb, J.; Miekisch, W. Continuous real-time breath analysis in ruminants: Effect of eructation on exhaled VOC profiles. J. Breath Res. 2018, 12. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Description | Units |
|---|---|---|
| A | acetate proportion | mol/100 mol total VFA |
| B | total butyrate proportion | mol/100 mol total VFA |
| C2 | acetate quantity | mol |
| C3 | propionate quantity | mol |
| C4 | total butyrate quantity | mol |
| CH4P | methane production, stoichiometry | mol |
| CH4Y | methane yield, stoichiometry | mol/100 mol total VFA |
| DMI | dry matter intake | kg/d |
| MY | methane yield, predicted | g methane/kg DMI |
| P | propionate proportion | mol/100 mol total VFA |
| [P] | propionate concentration | mmol/L |
| VFA | Volatile fatty acids | mol |
| Expt | Reference | Expt Design 1 | Dietary Treatment | n Cows | Methane Method | DIM | Diet Base | Season | Milk Yield | ECM 2 | Forage DMI | Concentrate DMI | Total DMI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (kg/Day) | (kg/Day) | (kg/Day) | (kg/Day) | (kg/Day) | |||||||||
| 1 | [24] | Randomized block | Control (lucerne) | 12 | SF6 | 179 | Lucerne | Autumn | 25.6 | 26.4 | 14.2 | 8.2 | 22.4 |
| Almond hulls | 10 | 177 | 23.2 | 24.6 | 14.4 | 8.2 | 22.6 | ||||||
| Citrus pulp | 10 | 176 | 25.9 | 25.4 | 13.2 | 7.8 | 21.0 | ||||||
| 2 | [25] | Randomized block | DHA-0 g | 8 | Calorimeter | 218 | Lucerne | Autumn | 22.2 | 25.4 | 18.2 | 5.9 | 24.1 |
| DHA-25 g | 7 | 215 | 26.0 | 24.0 | 18.0 | 6.1 | 24.1 | ||||||
| DHA-50 g | 8 | 216 | 23.1 | 21.9 | 16.5 | 6.2 | 22.7 | ||||||
| DHA-75 g | 7 | 215 | 22.3 | 21.6 | 15.3 | 6.2 | 21.5 | ||||||
| 3 | [26] | Full crossover | Control (water) | 8 | Calorimeter | 90 | Lucerne | Spring | 32.3 | 32.3 | 19.1 | 5.8 | 24.9 |
| Fat | 7 | 95 | 34.5 | 32.0 | 17.8 | 5.9 | 23.7 | ||||||
| Fat Plus Tannin | 8 | 90 | 33.7 | 31.3 | 17.9 | 5.9 | 23.8 | ||||||
| Tannin | 8 | 90 | 31.1 | 30.0 | 18.7 | 5.9 | 24.6 | ||||||
| 4 | [27] | Randomized block | Control (lucerne) | 12 | SF6 | 149 | Lucerne | Summer | 23.9 | 22.7 | 15.4 | 5.4 | 20.8 |
| Forage brassica | 10 | 150 | 27.5 | 25.4 | 15.2 | 5.4 | 20.6 | ||||||
| Chicory | 10 | 145 | 20.4 | 19.3 | 12.3 | 5.4 | 17.7 | ||||||
| 5 | [28] | Full crossover | Corn | 14 | Calorimeter | 191 | Lucerne | Autumn | 27.4 | 28.6 | 10.1 | 12.2 | 22.3 |
| Wheat | 14 | 191 | 29.1 | 23.8 | 9.0 | 11.5 | 20.5 | ||||||
| 6 | [29] | Randomized block | Wheat-0 kg | 8 | SF6 | 57 | Pasture | Spring | 29.9 | 29.5 | 17.1 | 2.1 | 19.2 |
| Wheat-3 kg | 8 | 57 | 31.3 | 32.4 | 15.4 | 5.0 | 20.4 | ||||||
| Wheat-6 kg | 8 | 57 | 32.3 | 33.0 | 12.3 | 7.9 | 20.2 | ||||||
| Wheat-9 kg | 8 | 57 | 34.7 | 32.9 | 8.9 | 10.9 | 19.8 | ||||||
| 7 | [30] | Randomized block | Corn 2 times/d | 8 | SF6 | 206 | Lucerne | Autumn | 21.2 | 20.6 | 8.7 | 10.2 | 18.9 |
| Wheat 2 times/d | 8 | 157 | 21.3 | 22.0 | 8.8 | 10.2 | 19.0 | ||||||
| Wheat 6 times/d | 8 | 161 | 24.0 | 22.6 | 8.7 | 10.1 | 18.8 | ||||||
| Wheat + buffer, 2 times/d | 8 | 197 | 20.5 | 22.4 | 8.8 | 10.4 | 19.2 |
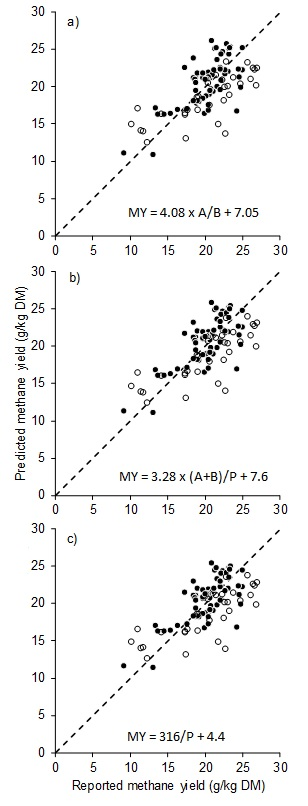
| Parameter | Methane Yield (g CH4/kg DMI) | Acetate (M%) | Propionate (M%) | Butyrate (M%) |
|---|---|---|---|---|
| Characteristics | ||||
| Mean (±s.d.) | 20.7 (4.81) | 63.3 (4.53) | 20.2 (5.44) | 9.9 (4.02) |
| Minimum | 6.9 | 51.3 | 12.9 | 0.7 |
| Maximum | 32.4 | 70.7 | 40.2 | 15.6 |
| Experiment | Experiment Design | N 1 | Animals | Methane Method | Days in Milk | Diet Base | Methane Yield | Acetate | Propionate | Butyrate |
|---|---|---|---|---|---|---|---|---|---|---|
| (g/kg DM) | (M%) | (M%) | (M%) | |||||||
| [6] | Randomized block | 32 | Dairy cows | SF6 | 71 | Lucerne hay | 10.1–26.9 | 51.5–67.4 | 16.3–38.5 | 6.5–15.3 |
| [14] | Full crossover | 3 | Dairy cows | Calorimeter | 155 | TMR, barley silage | 18.5–19.2 | 61.5–61.7 | 22.8–23.1 | 11.5–11.5 |
| [36] | Randomized block | 4 | Beef heifers | Calorimeter | - | TMR, barley | 9.2–24.8 | 42.6–64.0 | 20.5–45.7 | 7.1–12.6 |
| [37] | Latin square | 4 | Beef heifers | Calorimeter | - | Barley silage | 19.9–21.6 | 59.0–65.0 | 20.6–25.9 | 9.2–11.4 |
| [38] | Latin square | 3 | Beef cattle | Calorimeter | - | Barley silage | 18.5–18.8 | 64.6–65.8 | 19.0–19.6 | 11.5–11.6 |
| [39] | Crossover | 4 | Dairy cows | Calorimeter | 96 | Barley silage and grain | 13.4–16.3 | 59.9–61.7 | 25.1–26.7 | 8.7–10.3 |
| [40] | Latin square | 4 | Dairy cows | Calorimeter | 99 | Alfalfa and corn silage | 18.9–20.6 | 60.1–63.4 | 21.8–23.1 | 12.3–14.4 |
| [41] | Latin square | 4 | Dairy cows | Calorimeter | 61 | Grass silage | 17.3–21.3 | 69.5–70.4 | 15.1–18.6 | 8.1–11.8 |
| [42] | Latin square | 4 | Dairy cows | Calorimeter | 61 | Corn silage | 18.7–20.2 | 64.7–66.1 | 18.0–19.8 | 10.4–12.8 |
| [43] | Randomized block | 2 | Dairy cows | Calorimeter | - | Grass hay | 20.7–25.0 | 67.0–70.2 | 158–19.4 | 10.1–10.4 |
| [44] | Latin square | 3 | Dairy cows | Calorimeter | 92 | Silage | 19.7–20.1 | 66.6–67.8 | 19.2–19.5 | 10.2–11.0 |
| [45] | Randomized block | 4 | Dairy cows | Calorimeter | 215 | TMR, grass silage | 21.5–22.4 | 67.9–69.1 | 15.6–16.8 | 11.1–11.8 |
| [46] | Randomized block | 8 | Dairy cows | Calorimeter | 139 | TMR, corn | 19.7–23.1 | 64.2–66.7 | 18.0–20.8 | 11.5–12.4 |
| [47] | Randomized block | 2 | Beef steers | Calorimeter | - | Grass hay | 22.9–23.2 | 70.0–73.5 | 16.1–16.6 | 7.6–10.2 |
| [48] | Latin square | 4 | Beef steers | Calorimeter | - | TMR, barley silage | 18.8–22.6 | 64.0–68.0 | 18.3–21.6 | 8.6–10.4 |
| [49] | Crossover | 8 | Dairy cows | Calorimeter | 187 | Grass clover silage | 15.4–23.4 | 60.2–69.7 | 15.4–26.5 | 9.3–13.5 |
| [50] | Randomized block | 4 | Dairy cows | Calorimeter | 192 | Grass or corn silage | 22.0–25.0 | 63.6–66.0 | 17.1–18.9 | 12.8–16.3 |
| [51] | Randomized block | 2 | Dairy cows | Calorimeter | 176 | TMR, grass silage | 20.5–22.1 | 58.6–60.3 | 19.8–23.0 | 13.6–15.3 |
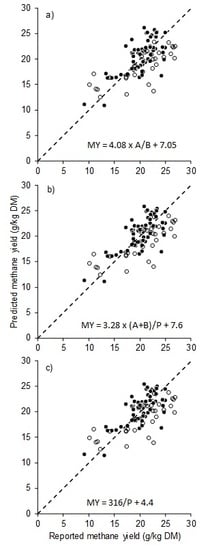
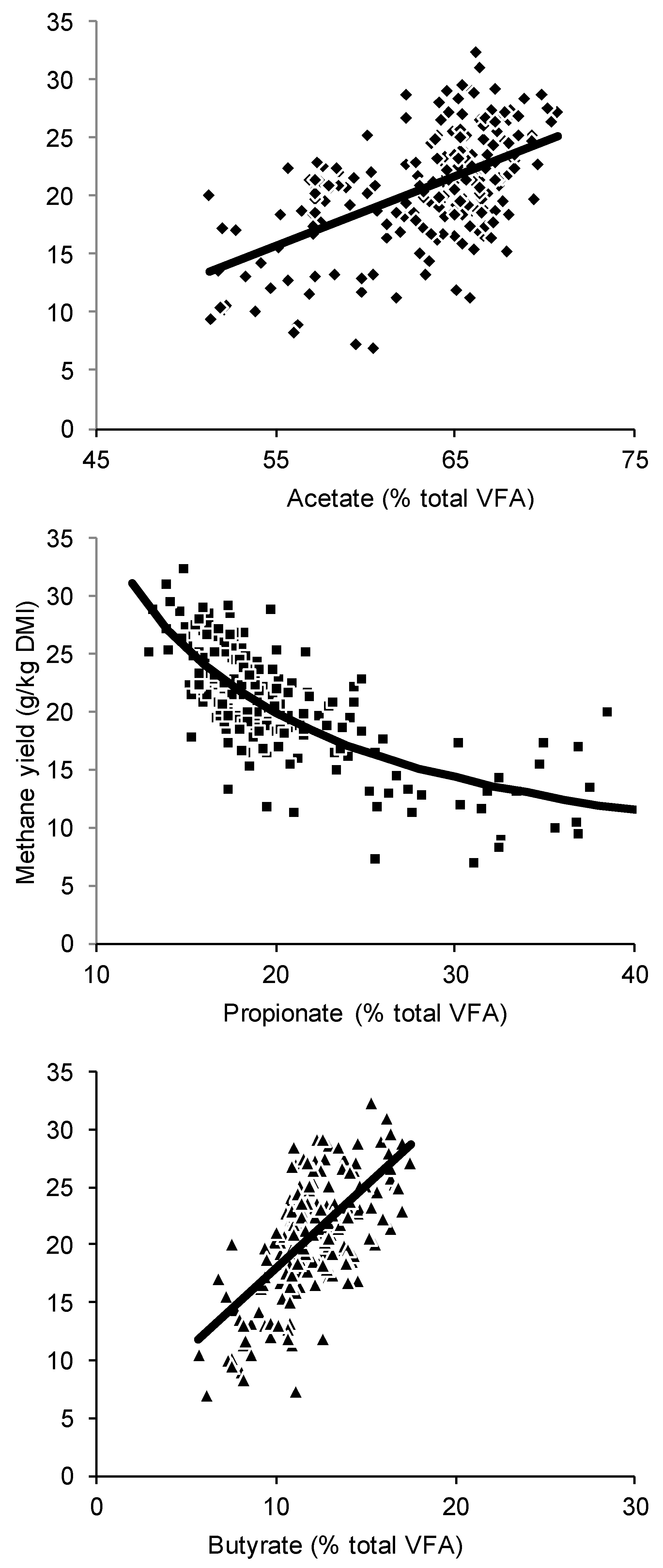
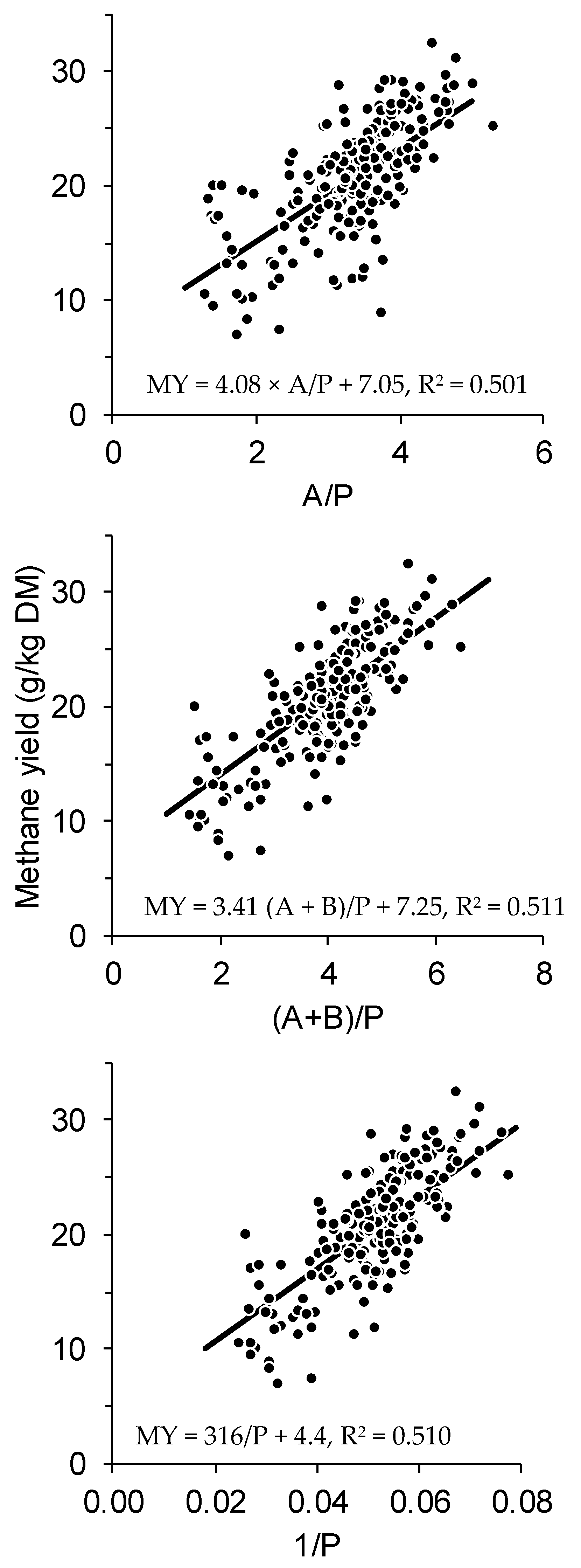
| Equation Number | Model | Estimates ± S.E. | RMSEP | CCC | Rank |
|---|---|---|---|---|---|
| (1c) | MY = 16 × (0.50A − 0.25P + 0.50B) × c/100 | c = 3.98 ± 0.15 | 3.7 | 0.52 | 4 |
| (2) | MY = dA − eP + fB | d = 0.30 ± 0.042, e = 0.22 ± 0.055, f = 0.48 ± 0.16 | 3.6 | 0.63 | 4 |
| (3) | MY = g(A/P) + h | g = 4.08 ± 0.36, h = 7.05 ± 1.40 | 3.2 | 0.69 | 1 |
| (4) | MY = i(A+B)/P + j | i = 3.28 ± 0.29. j = 7.60 ± 1.28 | 3.2 | 0.70 | 1 |
| (5) | MY = kP + m | k =−0.57 ± 0.057, m = 32.3 ± 1.4 | 3.5 | 0.63 | 4 |
| (6) | MY = n[P] + q | n =−0.24 ± 0.035, q = 26.1 ± 1.3 | 4.2 | 0.36 | 7 |
| (7) | MY = s/P + t | s = 316 ± 28, t = 4.4 ± 1.5 | 3.2 | 0.70 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, S.R.O.; Hannah, M.C.; Jacobs, J.L.; Wales, W.J.; Moate, P.J. Volatile Fatty Acids in Ruminal Fluid Can Be Used to Predict Methane Yield of Dairy Cows. Animals 2019, 9, 1006. https://doi.org/10.3390/ani9121006
Williams SRO, Hannah MC, Jacobs JL, Wales WJ, Moate PJ. Volatile Fatty Acids in Ruminal Fluid Can Be Used to Predict Methane Yield of Dairy Cows. Animals. 2019; 9(12):1006. https://doi.org/10.3390/ani9121006
Chicago/Turabian StyleWilliams, S. Richard O., Murray. C. Hannah, Joe L. Jacobs, William J. Wales, and Peter J. Moate. 2019. "Volatile Fatty Acids in Ruminal Fluid Can Be Used to Predict Methane Yield of Dairy Cows" Animals 9, no. 12: 1006. https://doi.org/10.3390/ani9121006
APA StyleWilliams, S. R. O., Hannah, M. C., Jacobs, J. L., Wales, W. J., & Moate, P. J. (2019). Volatile Fatty Acids in Ruminal Fluid Can Be Used to Predict Methane Yield of Dairy Cows. Animals, 9(12), 1006. https://doi.org/10.3390/ani9121006