A 5-Methylcytosine Site of Growth Differentiation Factor 9 (GDF9) Gene Affects Its Tissue-Specific Expression in Sheep


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Detection of GDF9 Expression by Real-Time PCR
2.3. Sequence Analysis of GDF9 5′-Flanking Sequence
2.4. DNA Isolation and Bisulfite Treatment
2.5. Bisulfite Sequencing
2.6. GDF9 Promoter Deletion Constructs
2.7. Site-Directed Mutation of Sp4 Binding Element
2.8. In Vitro-Methylation
2.9. Luciferase Assay
2.10. Statistical Analysis
3. Results
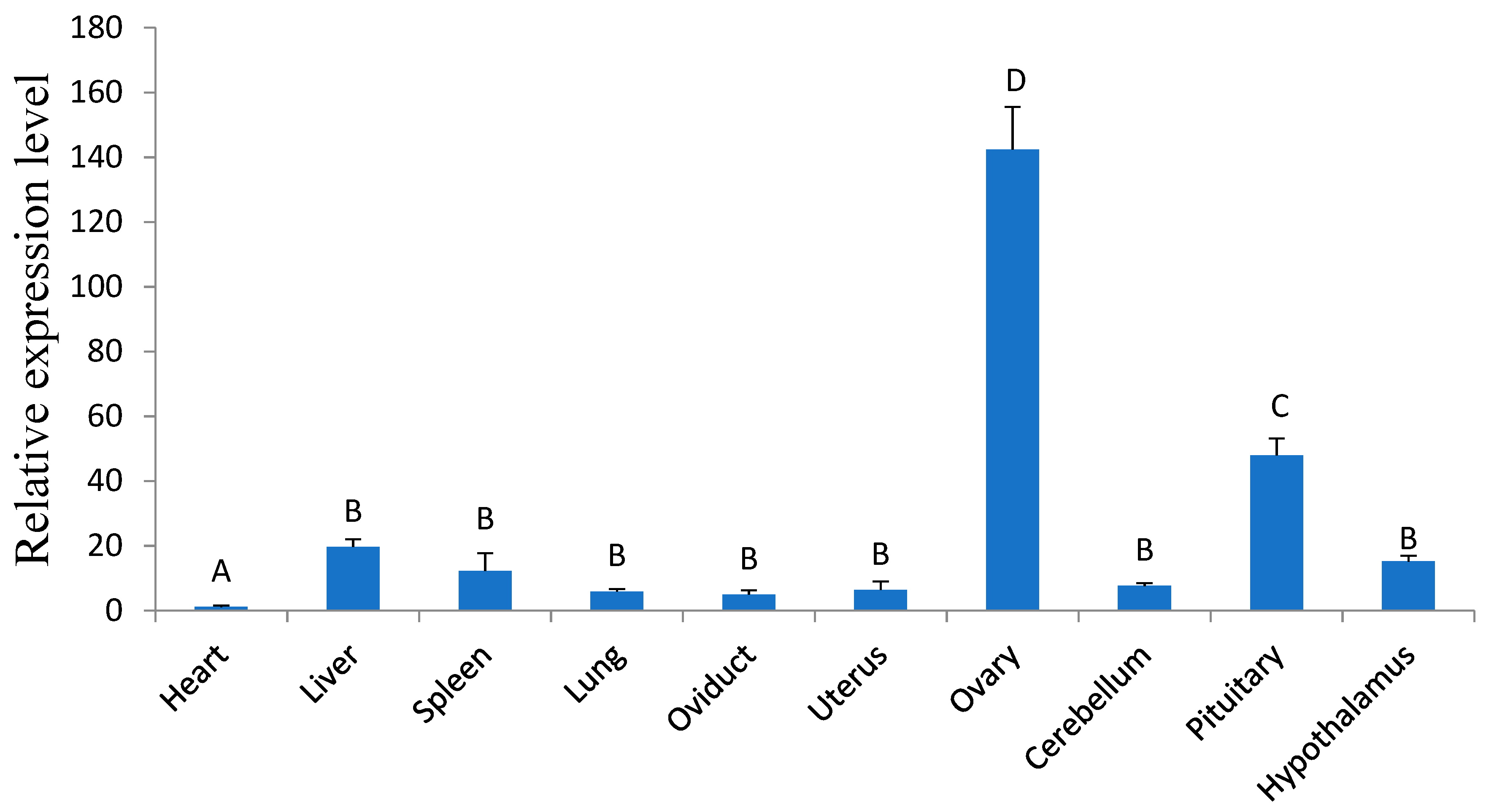
3.1. The Tissue Expression of Ovine GDF9
3.2. Analysis of the GDF9 Promoter Region
3.3. Methylation Level
3.4. Correlation between the Methylation Level and mRNA Expression
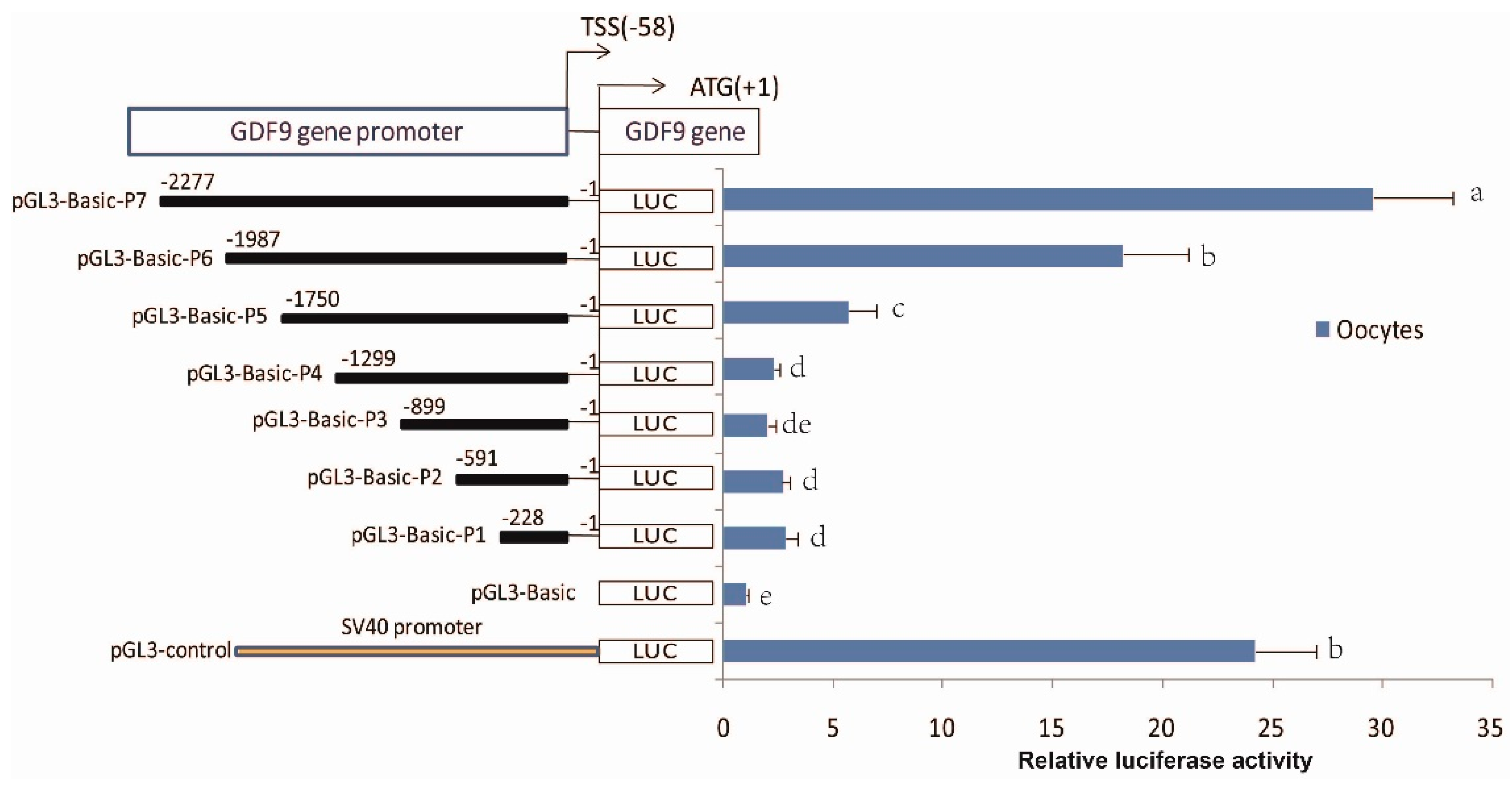
3.5. Identification of the Core Region of the Ovine GDF9
3.6. Verification of the Effect of the mC-4 Site
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McPherron, A.C.; Lee, S.J. GDF-3 and GDF-9: Two new members of the transforming growth factor-beta superfamily containing a novel pattern of cysteines. J. Biol. Chem. 1993, 268, 3444–3449. [Google Scholar] [PubMed]
- Dong, J.W.; Albertini, D.F.; Nishimori, K.; Kumar, T.R.; Lu, N.F.; Matzuk, M.M. Growth differentiation factor-9 is required during early ovarian folliculogenesis. Nature 1996, 383, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Melo, E.; Silva, B.; Castro, E.; Silva, T.; Paiva, S.; Sartori, R.; Franco, M.; Souza, C.J.; Neves, J. A novel mutation in the growth and differentiation factor 9 (GDF9) gene is associated, in homozygosis, with increased ovulation rate in Santa Ines sheep. Biol. Reprod. 2008, 78, 141. [Google Scholar] [CrossRef]
- Nicol, L.; Bishop, S.C.; Pong-Wong, R.; Bendixen, C.; Holm, L.E.; Rhind, S.M.; McNeilly, A.S. Homozygosity for a single base-pair mutation in the oocyte-specific GDF9 gene results in sterility in Thoka sheep. Reproduction 2009, 138, 921–933. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.D.M.; Castro, E.A.; Souza, C.J.H.; Paiva, S.R.; Sartori, R.; Franco, M.M.; Azevedo, H.C.; Silva, T.A.S.N.; Vieira, A.M.C.; Neves, J.P.; et al. A new polymorphism in the Growth and Differentiation Factor 9 (GDF9) gene is associated with increased ovulation rate and prolificacy in homozygous sheep. Anim. Genet. 2011, 42, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Vage, D.I.; Husdal, M.; Kent, M.P.; Klemetsdal, G.; Boman, I.A. A missense mutation in growth differentiation factor 9 (GDF9) is strongly associated with litter size in sheep. BMC Genet. 2013, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.J.H.; McNeilly, A.S.; Benavides, M.V.; Melo, E.O.; Moraes, J.C.F. Mutation in the protease cleavage site of GDF9 increases ovulation rate and litter size in heterozygous ewes and causes infertility in homozygous ewes. Anim. Genet. 2014, 45, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Hanrahan, J.P.; Gregan, S.M.; Mulsant, P.; Mullen, M.; Davis, G.H.; Powell, R.; Galloway, S.M. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol. Reprod. 2004, 70, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Pokharel, K.; Peippo, J.; Honkatukia, M.; Seppälä, A.; Rautiainen, J.; Ghanem, N.; Hamama, T.M.; Crowe, M.A.; Andersson, M.; Li, M.H. Integrated ovarian mRNA and miRNA transcriptome profiling characterizes the genetic basis of prolificacy traits in sheep (Ovis aries). BMC Genom. 2018, 19, 104. [Google Scholar] [CrossRef] [PubMed]
- Jackowska, M.; Kempisty, B.; Wozna, M.; Piotrowska, H.; Antosik, P.; Zawierucha, P.; Bukowska, D.; Nowicki, M.; Jaskowski, J.M.; Brussow, K.P. Differential expression of GDF9, TGFB1, TGFB2 and TGFB3 in porcine oocytes isolated from follicles of different size before and after culture in vitro. Acta Vet. Hung. 2013, 61, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.; Sharma, V.; Dubey, P.K.; Pratheesh, M.D.; Gade, N.E.; Saikumar, G.; Sharma, G.T. Impact of gonadotropin supplementation on the expression of germ cell marker genes (MATER, ZAR1, GDF9, and BMP15) during in vitro maturation of buffalo (Bubalus bubalis) oocyte. In Vitro Cell. Dev. Biol. Anim. 2013, 49, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Aaltonen, J.; Laitinen, M.P.; Vuojolainen, K.; Jaatinen, R.; Horelli-Kuitunen, N.; Seppa, L.; Louhio, H.; Tuuri, T.; Sjoberg, J.; Butzow, R.; et al. Human growth differentiation factor 9 (GDF-9) and its novel homolog GDF-9B are expressed in oocytes during early folliculogenesis. J. Clin. Endocrinol. Metab. 1999, 84, 2744–2750. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yuan, C.; Qin, F.; Hu, G.; Wang, Z. Molecular characterization of gdf9 and bmp15 genes in rare minnow Gobiocypris rarus and their expression upon bisphenol A exposure in adult females. Gene 2014, 546, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.Y.; Di, R.; Tang, Q.Q.; Jin, H.H.; Chu, M.X.; Huang, D.W.; He, J.N.; Liu, Q.Y.; Hu, W.P.; Wang, X.Y.; et al. Tissue-specific mRNA expression profiles of GDF9, BMP15, and BMPR1B genes in prolific and non-prolific goat breeds. Czech J. Anim. Sci. 2015, 60, 452–458. [Google Scholar] [CrossRef]
- Hu, D.L.; Li, Q.F.; Xu, Y.F.; Li, E.L.; Han, Y.D.; Tu, F.; Xie, Z. The tissue expression profile, mRNA expression level and SNPs analysis on GDF9 gene in Hu sheep. J. Agric. Biotechnol. 2010, 18, 533–538. [Google Scholar]
- Clark, E.L.; Bush, S.J.; Mcculloch, M.; Farquhar, I.L.; Young, R.; Lefevre, L.; Pridans, C.; Tsang, H.G.; Wu, C.; Afrasiabi, C. A high resolution atlas of gene expression in the domestic sheep (Ovis aries). PLoS Genet. 2017, 13, e1006997. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Xie, M.; Chen, W.; Talbot, R.; Maddox, J.F.; Faraut, T.; Wu, C.; Muzny, D.M.; Li, Y.; Zhang, W. The sheep genome illuminates biology of the rumen and lipid metabolism. Science 2014, 344, 1168–1173. [Google Scholar] [CrossRef] [PubMed]
- Kona, S.S.; Praveen Chakravarthi, V.; Siva Kumar, A.V.; Srividya, D.; Padmaja, K.; Rao, V.H. Quantitative expression patterns of GDF9 and BMP15 genes in sheep ovarian follicles grown in vivo or cultured in vitro. Theriogenology 2016, 85, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Feary, E.S.; Juengel, J.L.; Smith, P.; French, M.C.; O’Connell, A.R.; Lawrence, S.B.; Galloway, S.M.; Davis, G.H.; McNatty, K.P. Patterns of expression of messenger RNAs encoding GDF9, BMP15, TGFBR1, BMPR1B, and BMPR2 during follicular development and characterization of ovarian follicular populations in ewes carrying the Woodlands FecX2W mutation. Biol. Reprod. 2007, 77, 990–998. [Google Scholar] [CrossRef] [PubMed]
- Abdoli, R.; Zamani, P.; Deljou, A.; Rezvan, H. Association of BMPR-1B and GDF9 genes polymorphisms and secondary protein structure changes with reproduction traits in Mehraban ewes. Gene 2013, 524, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Varnosfaderani, S.R.; Ostadhosseini, S.; Hajian, M.; Hosseini, S.M.; Khashouei, E.A.; Abbasi, H.; Hosseinnia, P.; Nasr-Esfahani, M.H. Importance of the GDF9 signaling pathway on cumulus cell expansion and oocyte competency in sheep. Theriogenology 2013, 80, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Bodensteiner, K.J.; McNatty, K.P.; Clay, C.M.; Moeller, C.L.; Sawyer, H.R. Expression of growth and differentiation factor-9 in the ovaries of fetal sheep homozygous or heterozygous for the inverdale prolificacy gene (FecX(I)). Biol. Reprod. 2000, 62, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.D.; Parnaik, V.K. Identification of tissue-specific regulatory region in the zebrafish lamin A promoter. Gene 2015, 567, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Guske, K.; Schmitz, B.; Schelleckes, M.; Duning, K.; Kremerskothen, J.; Pavenstadt, H.J.; Brand, S.M.; Brand, E. Tissue-specific differences in the regulation of KIBRA gene expression involve transcription factor TCF7L2 and a complex alternative promoter system. J. Mol. Med. 2014, 92, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Gong, W.; Wang, C.; Gao, Y.; Hong, W.; Chen, S.X. Dynamic methylation pattern of cyp19a1a core promoter during zebrafish ovarian folliculogenesis. Fish Physiol. Biochem. 2016, 42, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Jing, W.; Yin, X.; Sun, S.; Chen, Z.; Zhu, G.; Wu, S.; Bao, W. Identification of a 5-Methylcytosine Site that may Regulate C/EBPβ Binding and Determine Tissue-Specific Expression of theBPIGene in Piglets. Sci. Rep. 2016, 6, 28506. [Google Scholar]
- Wu, W.; Ren, Z.; Liu, H.; Wang, L.; Huang, R.; Chen, J.; Zhang, L.; Li, P.; Xiong, Y. Core promoter analysis of porcine Six1 gene and its regulation of the promoter activity by CpG methylation. Gene 2013, 529, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.Y.; He, X.Y.; Liu, Q.Y.; Hu, W.P.; Wang, X.Y.; Guo, X.F.; Cao, X.H.; Di, R.; Chu, M.X. Cloning and genetic polymorphism analysis of mRNA, DNA and regulatory region of ovine GDF9 gene in 11 breeds. Acta Vet. Zootech. Sin. 2016, 47, 1555–1564. [Google Scholar]
- Tu, Y.R. The Sheep and Goat Breeds in China; Shanghai Science and Technology Press: Shanghai, China, 1989. [Google Scholar]
- Roy, B.; Rajput, S.; Raghav, S.; Kumar, P.; Verma, A.; Kumar, S.; De, S.; Goswami, S.L.; Datta, T.K. A reporter promoter assay confirmed the role of a distal promoter NOBOX binding element in enhancing expression of GDF9 gene in buffalo oocytes. Anim. Reprod. Sci. 2012, 135, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, S.L.; Sindoni, D.M.; Shughrue, P.J.; Lane, M.V.; Merchenthaler, I.J.; Frail, D.E. Expression of growth differentiation factor-9 messenger ribonucleic acid in ovarian and nonovarian rodent and human tissues. Endocrinology 1998, 139, 2571–2578. [Google Scholar] [CrossRef] [PubMed]
- Hosoe, M.; Kaneyama, K.; Ushizawa, K.; Hayashi, K.G.; Takahashi, T. Quantitative analysis of bone morphogenetic protein 15 (BMP15) and growth differentiation factor 9 (GDF9) gene expression in calf and adult bovine ovaries. Reprod. Biol. Endocrinol. 2011, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Su, J.; Wan, Q.; Feng, X. CpG methylation in the 5′-flanking region of LGP2 gene lacks association with resistance/susceptibility to GCRV but contributes to the differential expression between muscle and spleen tissues in grass carp, Ctenopharyngodon idella. Fish Shellfish Immunol. 2014, 40, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Huse, S.M.; Gruppuso, P.A.; Boekelheide, K.; Sanders, J.A. Patterns of gene expression and DNA methylation in human fetal and adult liver. BMC Genom. 2015, 16, 981. [Google Scholar] [CrossRef] [PubMed]
- Lister, R.; Pelizzola, M.; Dowen, R.H.; Hawkins, R.D.; Hon, G.; Tontifilippini, J.; Nery, J.R.; Lee, L.K.; Ye, Z.; Ngo, Q. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature 2009, 462, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Supp, D.M.; Witte, D.P.; Branford, W.W.; Smith, E.P.; Potter, S.S. Sp4, a member of the Sp1-family of zinc finger transcription factors, is required for normal murine growth, viability, and male fertility. Dev. Biol. 1996, 176, 284–299. [Google Scholar] [CrossRef] [PubMed]
- Gollner, H.; Bouwman, P.; Mangold, M.; Karis, A.; Braun, H.; Rohner, I.; Del Rey, A.; Besedovsky, H.O.; Meinhardt, A.; van den Broek, M.; et al. Complex phenotype of mice homozygous for a null mutation in the Sp4 transcription factor gene. Genes Cells 2001, 6, 689–697. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′→3′) | Annealing Temperature (°C) | Amplified DNA Fragment (bp) |
|---|---|---|---|
| Primers used in real-time PCR | |||
| GDF9-YG-F | CAGACGCCACCTCTACAACA | 60 | 197 |
| GDF9-YG-R | CAGGAAAGGGAAAAGAAATGG | ||
| GAPDH-F | GAGAAACCTGCCAAGTATGA | 60 | 139 |
| GAPDH-R | CGAAGGTAGAAGAGTGAGTG | ||
| Primers used in promoter activity analysis | |||
| P1F | CGGGGTACCACTATATGGCCAAGTAAATCTGAATC | 58 | 228 |
| P2F | CGGGGTACCGTTCTCTGCTCTCTGGAATCTCAATTTC | 58 | 591 |
| P3F | CGGGGTACCCCTGATCTTAGCTCAGAGGCAAGAAC | 58 | 899 |
| P4F | CGGGGTACCTGTCATGTTGCCCACTGTTCACTGCC | 58 | 1299 |
| P5F | CGGGGTACCGGCTTTCGTCGGAG | 58 | 1750 |
| P6F | CGGGGTACCCCACTTCCGGTAGATCGGACG | 58 | 1987 |
| P7F | CGGGGTACCCTGCTCGAAGGGCGACAAGCTAT | 58 | 2277 |
| PR | GTCGTCAAGCTTGGCTTGGAAGAATTAGCAAGG | - | - |
| Primers used in methylation analysis | |||
| M1F | GGGATTTGTCGTCGTTAAT | 55 | 352 |
| M1R | CAAAACCCGCCCAAAAAC | ||
| TFs | Description | Position | Sequence | Strand |
|---|---|---|---|---|
| AP-2 | Transcription factor AP-2, beta | −1669, −1655 | gctGCCCtagggtcg | + |
| SP4 | Sp4 transcription factor | −1796, −1780 | tcccgggGCGCCcccca | + |
| ATF | Activating transcription factor 1 | −1867, −1847 | cccacttcACGTcacgcggcg | − |
| EBOX | E-box binding factors | −1869, −1853 | cccgccgCGTGacgtga | + |
| USF | Upstream stimulating factor 1 | −1870, −1854 | cacgTCACgcggcgggg | − |
| CREB | cAMP-responsive element binding protein | −1916, −1896 | gtctccaggTGACggcgccat | + |
| ERRG | Estrogen-related receptor gamma binding site | −1960, −1938 | ccaggaggcggtgaGGTCacttc | + |
| ER2 | Estrogen receptor 2 (ER beta) | −2034, −2016 | cgagGTCActtcgcccact | + |
| ERRB | Estrogen-related receptor beta | −2045, −2023 | cactcacacaccgAGGTcacttc | + |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, Z.; Wang, X.; Di, R.; Liu, Q.; Hu, W.; Cao, X.; Guo, X.; He, X.; Lv, S.; Li, F.; et al. A 5-Methylcytosine Site of Growth Differentiation Factor 9 (GDF9) Gene Affects Its Tissue-Specific Expression in Sheep. Animals 2018, 8, 200. https://doi.org/10.3390/ani8110200
Pan Z, Wang X, Di R, Liu Q, Hu W, Cao X, Guo X, He X, Lv S, Li F, et al. A 5-Methylcytosine Site of Growth Differentiation Factor 9 (GDF9) Gene Affects Its Tissue-Specific Expression in Sheep. Animals. 2018; 8(11):200. https://doi.org/10.3390/ani8110200
Chicago/Turabian StylePan, Zhangyuan, Xiangyu Wang, Ran Di, Qiuyue Liu, Wenping Hu, Xiaohan Cao, Xiaofei Guo, Xiaoyun He, Shengjin Lv, Fukuan Li, and et al. 2018. "A 5-Methylcytosine Site of Growth Differentiation Factor 9 (GDF9) Gene Affects Its Tissue-Specific Expression in Sheep" Animals 8, no. 11: 200. https://doi.org/10.3390/ani8110200
APA StylePan, Z., Wang, X., Di, R., Liu, Q., Hu, W., Cao, X., Guo, X., He, X., Lv, S., Li, F., Wang, H., & Chu, M. (2018). A 5-Methylcytosine Site of Growth Differentiation Factor 9 (GDF9) Gene Affects Its Tissue-Specific Expression in Sheep. Animals, 8(11), 200. https://doi.org/10.3390/ani8110200