Effects of Herbal Supplementation on Growth Performance of Kenguri Sheep Exposed to Heat Stress

 , , , ,
, , , ,  ,
,  and
and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Experimental Location, Climate, and Design
2.3. Blood Collection for Hormonal Analysis
2.4. Blood Collection for PBMC Isolation
2.5. Variables Studied
2.6. Data Analysis
3. Results
3.1. Effect of Heat Stress and Herbal Supplementation on Feed Intake and Water Intake in Kenguri Sheep
3.1.1. Results on Feed Intake
3.1.2. Results on Water Intake
3.2. Effect of Heat Stress and Herbal Supplementation on Body Weight in Kenguri Sheep
3.3. Effect of Heat Stress and Herbal Supplementation on Allometric Measurements in Kenguri Sheep
3.4. Effect of Heat Stress and Herbal Supplementation on Endocrine Variables in Kenguri Sheep
3.4.1. Growth Hormone (GH)
3.4.2. Insulin-like Growth Factor (IGF-1)
3.4.3. Tri-Iodo-Thyronine (T3)
3.4.4. Thyroxine (T4)
3.5. Effect of Heat Stress and Herbal Supplementation on Expression Pattern of Targeted Genes in Kenguri Sheep
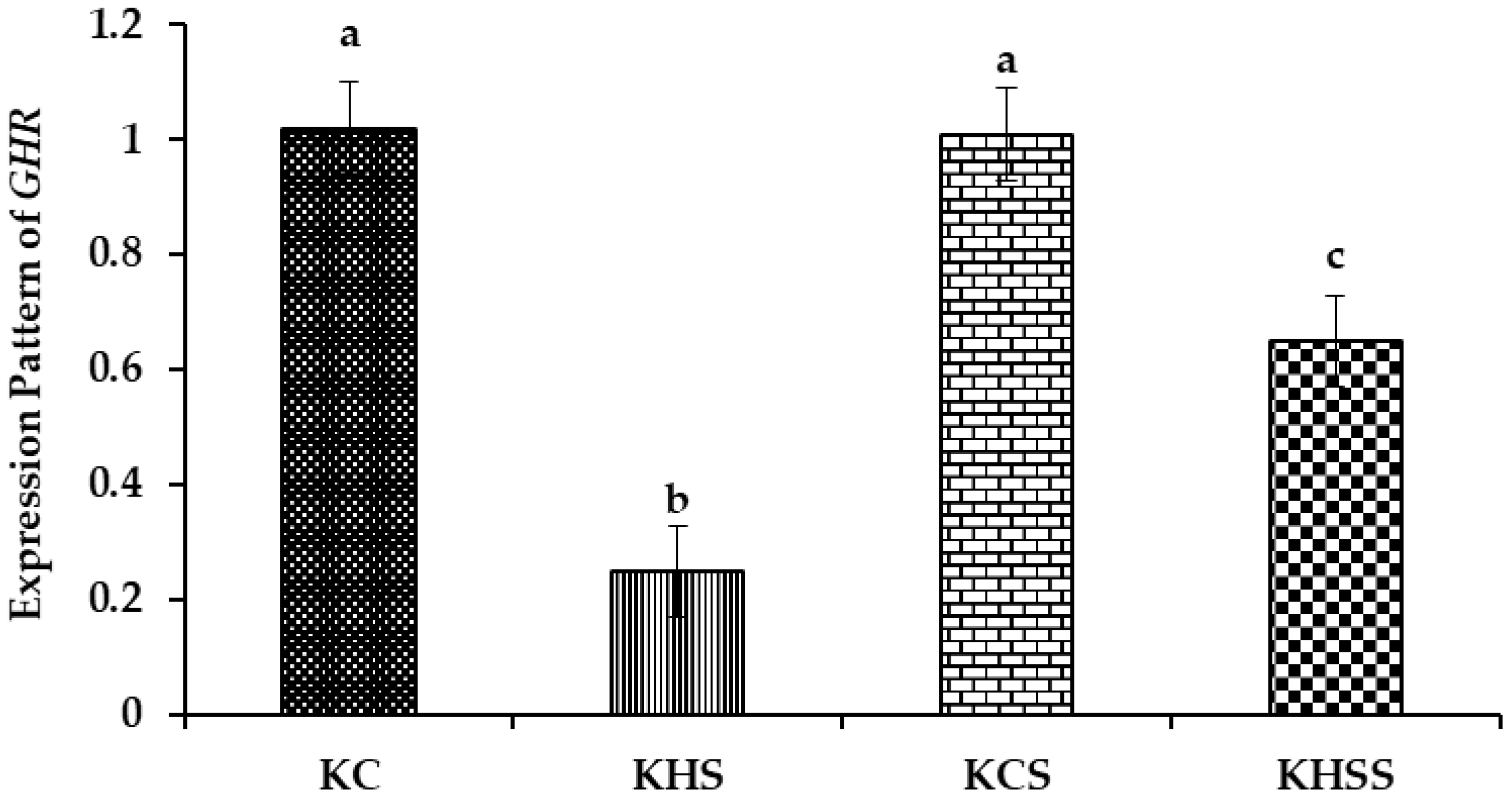
3.5.1. Expression Pattern of GHR
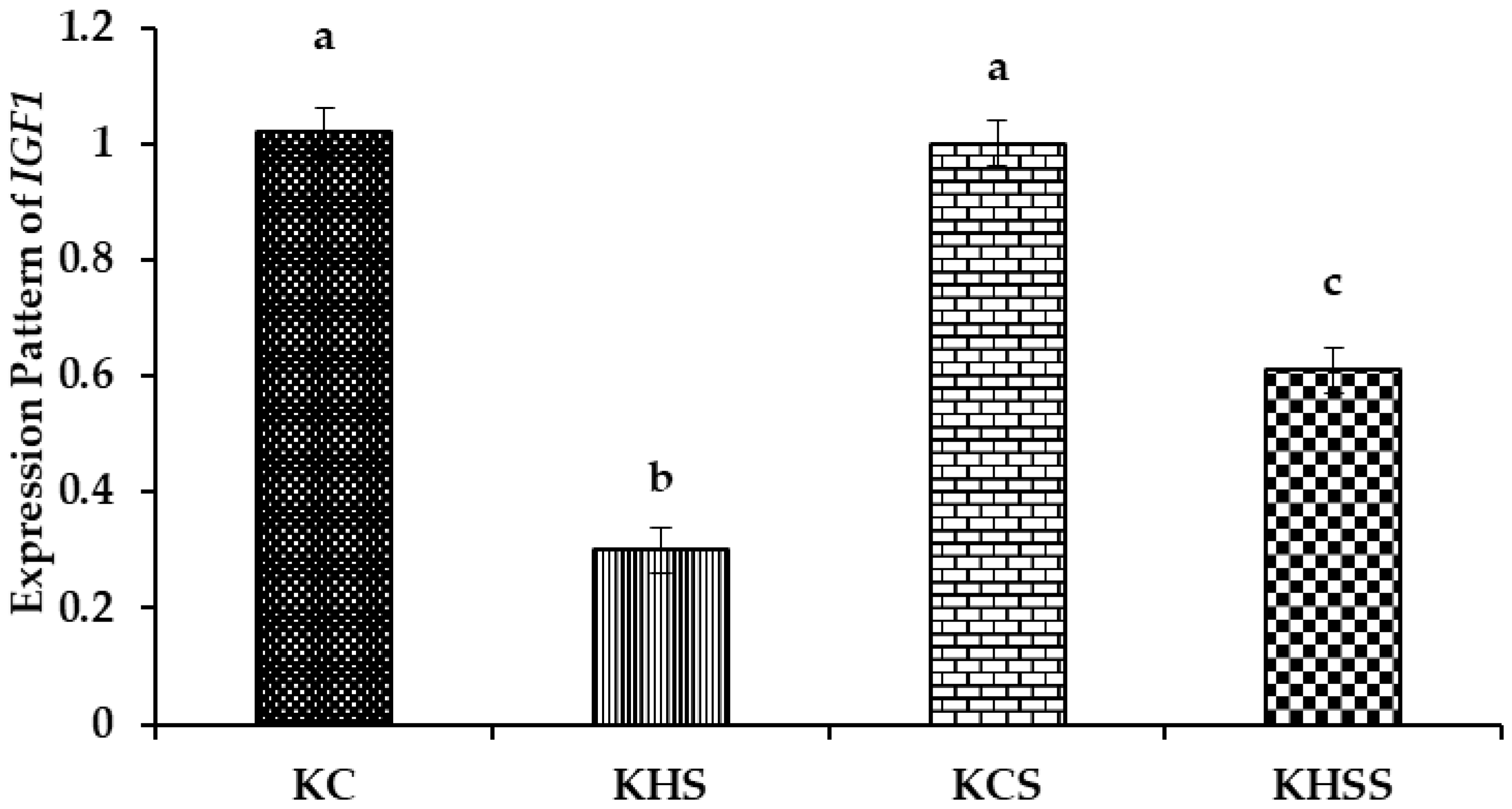
3.5.2. Expression Pattern of IGF1
3.5.3. Expression Pattern of PRLR
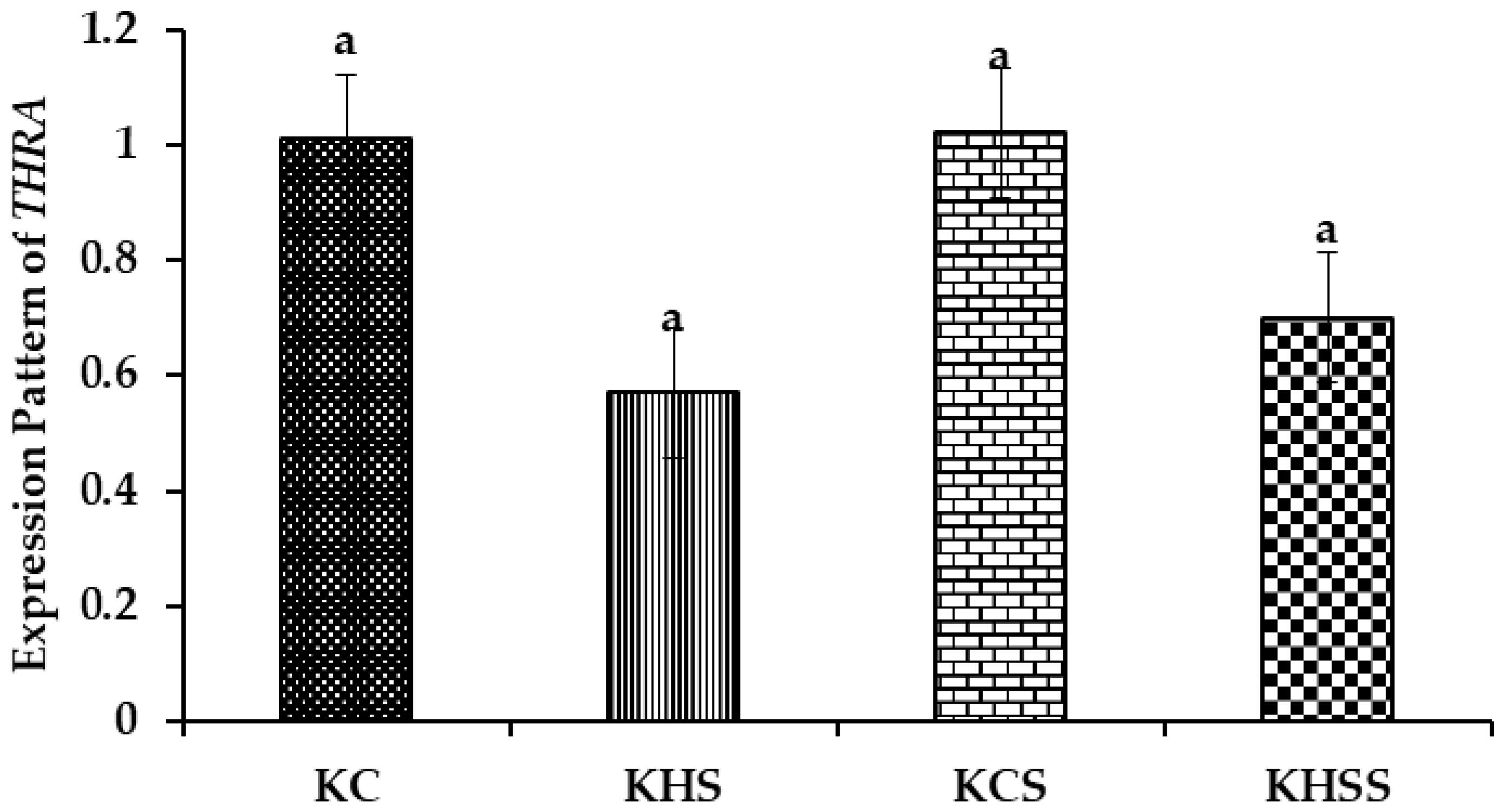
3.5.4. Expression Pattern of THRA
4. Discussion
4.1. Temperature–Humidity Index
4.2. Feed Intake
4.3. Water Intake
4.4. Body Weight
4.5. Allometric Measurements
4.6. Endocrine Variables
4.6.1. Plasma GH and IGF-1 Concentration
4.6.2. Plasma Thyroid Hormone Concentration
4.7. Gene Expression Patterns
4.7.1. The GHR and IGF-1 Expression Patterns
4.7.2. The PRLR Expression Pattern
4.7.3. The THR Expression Pattern
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Indu, S.; Pareek, A. A review: Growth and physiological adaptability of sheep to heat stress under semi–arid environment. Int. J. Emerg. Trends Sci. Technol. 2015, 2, 3188–3198. [Google Scholar] [CrossRef]
- IWTO. 2022 World Sheep Numbers & Wool Production. Available online: https://iwto.org/wp-content/uploads/2022/04/IWTO-Market-Information-Sample-Edition-17.pdf (accessed on 23 September 2024).
- Khani, M.; Fattah, A.; Ebrahimi-Mahmoudabad, S.; Joezy-Shekalgorabi, S. Impact of dietary cation-anion difference on rumen fermentation, digestibility, and blood parameters in Zandi lambs under heat stress. Agrobiol. Rec. 2023, 12, 60–67. [Google Scholar] [CrossRef]
- Yue, S.; Qian, J.; Du, J.; Liu, X.; Xu, H.; Liu, H.; Chen, X. Heat stress negatively influence mammary blood flow, mammary uptake of amino acids and milk amino acids profile of lactating Holstein dairy cows. Pak. Vet. J. 2023, 43, 73–78. [Google Scholar]
- Li, J.B.; Wang, X.; Sun, A.; Li, H.B.; Luo, Y.; He, F.; Yi, K.L. Comparative transcriptomic analysis of spermatozoa from Xiangxi and Simmental bulls under heat stress: Implications for fertility prediction. Pak. Vet. J. 2023, 43, 184–188. [Google Scholar] [CrossRef]
- Hussan, S.J.; Al-hummsod, S.K.; Al-Asadi, M.H. The Impact of Astaxanthin Supplementation on the Production Performance of Laying Hens Exposed to Heat Stress. J. Glob. Innov. Agric. Sci. 2023, 11, 637–642. [Google Scholar]
- Demir, E.; Ceccobelli, S.; Bilginer, U.; Pasquini, M.; Attard, G.; Karsli, T. Conservation and selection of genes related to environmental adaptation in native small ruminant breeds: A review. Ruminants 2022, 2, 255–270. [Google Scholar] [CrossRef]
- Wanjala, G. Genome-Wide Analysis of the Genetic Diversity of Native Sheep Breeds and Determination of Potential Selection Signatures for Climate Change Adaptation. Ph.D. Thesis, Debreceni Egyetem, Debrecen, Hungary, 2024. [Google Scholar]
- Marai, I.F.M.; El-Darawany, A.A.; Fadiel, A.; Abdel-Hafez, M.A.M. Physiological traits as affected by heat stress in sheep—A review. Small Rumin. Res. 2007, 71, 1–12. [Google Scholar] [CrossRef]
- Silanikove, N. Effects of heat stress on the welfare of extensively managed domestic ruminants. Livest. Prod Sci. 2000, 67, 1–18. [Google Scholar] [CrossRef]
- Marai, I.F.M.; Habeeb, A.A.M.; Farghaly, H.M. Productive, physiological and biochemical changes in imported and locally born Friesian and Holstein lactating cows under hot summer conditions of Egypt. Trop. Anim. Health Prod. 1999, 31, 233–243. [Google Scholar] [CrossRef]
- Kamanga-Sollo, E.; Pampusch, M.S.; White, M.E.; Hathaway, M.R.; Dayton, W.R. Effects of heat stress on proliferation, protein turnover, and levels of heat shock protein mRNA in cultured porcine muscle satellite cells. J. Anim. Sci. 2011, 89, 3473–3480. [Google Scholar] [CrossRef]
- McManus, C.M.; Lucci, C.M.; Maranhão, A.Q.; Pimentel, D.; Pimentel, F.; Paiva, S.R. Response to heat stress for small ruminants: Physiological and genetic aspects. Livest. Sci. 2022, 263, 105028. [Google Scholar] [CrossRef]
- West, J.W. Effects of heat-stress on production in dairy cattle. J. Dairy Sci. 2003, 86, 2131–2144. [Google Scholar] [CrossRef]
- Angel, S.P.; Bagath, M.; Sejian, V.; Krishnan, G.; Bhatta, R. Expression patterns of candidate genes reflecting the growth performance of goats subjected to heat stress. Mol. Biol. Rep. 2018, 45, 2847–2856. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Song, L.; Dong, Q.; Zhang, A.; Zhang, L.; Wang, Y.; Feng, M.; Li, X.; Li, F.; Sun, X.; et al. Effects of herbal formula on growth performance, apparent digestibility, antioxidant capacity, and rumen microbiome in fattening lambs under heat stress. Environ. Sci. Pollut. Res. 2024, 31, 51364–51380. [Google Scholar] [CrossRef]
- Saleh, A.A.; Soliman, M.M.; Yousef, M.F.; Eweedah, N.M.; El-Sawy, H.B.; Shukry, M.; Wadaan, M.A.; Kim, I.H.; Cho, S.; Eltahan, H.M. Effects of herbal supplements on milk production quality and specific blood parameters in heat-stressed early lactating cows. Front. Vet. Sci. 2023, 10, 1180539. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Li, L.; Yan, S.; Shi, B. Plant Extracts to Alleviating Heat Stress in Dairy Cows. Animals 2023, 13, 2831. [Google Scholar] [CrossRef]
- Lakhani, P.; Jindal, R.; Nayyar, S. Effect of heat stress on humoral immunity and its amelioration by amla powder (Emblica officinalis) supplementation in buffaloes. J. Anim. Res. 2016, 6, 401–404. [Google Scholar] [CrossRef]
- Verma, P.; Jain, A.K.; Mishra, A.; Jesse, D.D.; Mandal, S.; Gattani, A.; Patel, P.; Singh, P.; Jatav, M. Ameliorative effect of Withania somnifera on haemato-biochemical and hormonal parameters during heat stress in murrah buffalo calves. Int. J. Adv. Biochem. Res. 2024, 8, 80–85. [Google Scholar] [CrossRef]
- Saleh, A.A.K.; Abozed, G.F. Impact of using chamomile flower as a feed additive on reproductive performance and physiological responses of Farafra ewes during heat stress conditions. Egypt. J. Nutr. Feed. 2018, 21, 635–643. [Google Scholar] [CrossRef]
- Kalaitsidis, K.; Sidiropoulou, E.; Tsiftsoglou, O.; Mourtzinos, I.; Moschakis, T.; Basdagianni, Z.; Vasilopoulos, S.; Chatzigavriel, S.; Lazari, D.; Giannenas, I. Effects of cornus and its mixture with oregano and thyme essential oils on dairy sheep performance and milk, yoghurt and cheese quality under heat stress. Animals 2021, 11, 1063. [Google Scholar] [CrossRef]
- Chaturvedi, I.; Dutta, T.K.; Singh, P.K.; Chatterjee, A.; Mandal, D.K.; Bhakat, C.; Mohammad, A.; Das, A.K. Effect of supplementation of phytogenic feed additives on intake, in vitro fermentation, growth performance and carcass traits in weaned Barbari kids reared under intensive feeding. Trop. Anim. Health Prod. 2022, 54, 150. [Google Scholar] [CrossRef] [PubMed]
- Maurya, P.K.; Singh, R.K.; Chandra, G.; Ranjan, K.; Roy, D.; Kumar, P.; Fahim, A.; Sejian, V. The Effect of Silymarin and Selenium-yeast Supplementation on Antioxidant Status, Lipid Peroxidation and Metabolic Hormones in Heat-Stressed Barbari Goats. Res. Sq. 2024. [Google Scholar] [CrossRef]
- Lavanya, M.; Reddy, G.B.; Reddy, M.K.; Anjana, N.S.; Babu, D.S.; Ganguly, B. Anti-Inflammatory Effect of Polyherbal Feed Supplements in Heat Stressed Pigs. Indian Vet. J. 2023, 100, 32–35. [Google Scholar]
- Rajaei-Sharifabadi, H.; Ellestad, L.; Porter, T.; Donoghue, A.; Bottje, W.G.; Dridi, S. Noni (Morinda citrifolia) modulates the hypothalamic expression of stress-and metabolic-related genes in broilers exposed to acute heat stress. Front. Genet. 2017, 8, 192. [Google Scholar] [CrossRef]
- Flees, J.; Rajaei-Sharifabadi, H.; Greene, E.; Beer, L.; Hargis, B.M.; Ellestad, L.; Porter, T.; Donoghue, A.; Bottje, W.G.; Dridi, S. Effect of Morinda citrifolia (noni)-enriched diet on hepatic heat shock protein and lipid metabolism-related genes in heat stressed broiler chickens. Front. Physiol. 2017, 8, 919. [Google Scholar] [CrossRef]
- Selvam, R.; Suresh, S.; Saravanakumar, M.; Chandrasekaran, C.V. Alleviation of heat stress by a polyherbal formulation, Phytocee™: Impact on zootechnical parameters, cloacal temperature, and stress markers. Pharmacogn. Res. 2018, 10, 1. [Google Scholar]
- Suryadi, U.; Kustiawan, E.; Prasetyo, A.F.; Imam, S. Effect of agarwood leaf extract on production performance of broilers experiencing heat stress. Vet. World 2021, 14, 1971. [Google Scholar] [CrossRef]
- Al-Hoshani, N.; Almahallawi, R.; Al-Nabati, E.A.; Althubyani, S.A.; Negm, S.; El-lkott, A.F.; Majed, A.; Bajaber, M.; Soliman, A.E. Anthelmintic effects of herbal mixture of selected plants of Apiaceae on Strongylus vulgaris and Fasciola hepatica. Pak. Vet. J. 2024, 44, 437–441. [Google Scholar] [CrossRef]
- Mohammad, L.M.; Kamil, A.M.; Tawfeeq, R.K.; Ahmed, S.J. Ameliorating effects of herbal mixture for dexamethasone induced histological changes in mice. Int. J. Vet. Sci. 2023, 12, 126–131. [Google Scholar] [CrossRef]
- Gavzan, H.; Araghi, A.; Abbasabadi, B.M.; Talebpour, N.; Golshahi, H. Antidepressant effects of a Persian herbal formula on mice with chronic unpredictable mild stress. Avicenna J. Phytomed. 2023, 13, 562. [Google Scholar] [CrossRef]
- Bhatt, N. Herbs and Herbal Supplements, a Novel Nutritional Approach in Animal Nutrition. Iran. J. Appl. Anim. Sci. 2015, 5, 497–516. [Google Scholar]
- Li, Y.; Fang, L.; Xue, F.; Mao, S.; Xiong, B.; Ma, Z.; Jiang, L. Effects of bamboo leaf extract on the production performance, rumen fermentation parameters, and rumen bacterial communities of heat-stressed dairy cows. Anim. BioSci. 2021, 34, 1784. [Google Scholar]
- Zhao, S.; Shan, C.; Wu, Z.; Feng, M.; Song, L.; Wang, Y.; Gao, Y.; Guo, J.; Sun, X. Fermented Chinese herbal preparation: Impacts on milk production, nutrient digestibility, blood biochemistry, and antioxidant capacity of late-lactation cows under heat stress. Anim. Feed. Sci. Technol. 2022, 292, 115448. [Google Scholar] [CrossRef]
- Hashemzadeh, F.; Rafeie, F.; Hadipour, A.; Rezadoust, M.H. Supplementing a phytogenic-rich herbal mixture to heat-stressed lambs: Growth performance, carcass yield, and muscle and liver antioxidant status. Small Rumin. Res. 2022, 206, 106596. [Google Scholar] [CrossRef]
- McDowell, R.E. Improvement of Livestock Production in Warm Climates; W. H. Freeman: New York, NY, USA, 1972. [Google Scholar]
- de Sousa Silva, R.; Furtado, D.A.; Ribeiro, N.L.; Neto, J.P.L.; Rodrigues, R.C.M.; de Oliveira, A.G.; da Costa Silva, J.A.P.; da Silva, M.R.; Mascarenhas, N.M.H.; Marques, J.I.; et al. Physiological variables and estimates of heat exchange in sheep kept at thermoneutral and thermal stress temperatures. Small Rumin. Res. 2024, 237, 107320. [Google Scholar] [CrossRef]
- Mullakkalparambil Velayudhan, S.; Sejian, V.; Devaraj, C.; Manjunathareddy, G.B.; Ruban, W.; Kadam, V.; König, S.; Bhatta, R. Novel insights to assess climate resilience in goats using a holistic approach of skin-based advanced NGS technologies. Int. J. Mol. Sci. 2023, 24, 10319. [Google Scholar] [CrossRef] [PubMed]
- Aleena, J. Comparative assessment of the adaptive capacity of different indigenous breed goats to summer heat stress based on changes in phenotypic traits. Ph.D. Thesis, Academy of Climate Change Education and Research Vellanikkara, Kerala Agricultural University, Kerala, India, 2017. Available online: http://krishikosh.egranth.ac.in/handle/1/5810144116 (accessed on 28 April 2025).
- Das, R.; Sailo, L.; Verma, N.; Bharti, P.; Saikia, J.; Imtiwati; Kumar, R. Impact of heat stress on health and performance of dairy animals: A review. Vet. World 2016, 9, 260–268. [Google Scholar] [CrossRef]
- Avendaño-Reyes, L.; Macías-Cruz, U.; Correa-Calderón, A.; Mellado, M.; Corrales, J.L.; Corrales, G.; Ramirez-Bribiesca, E.; Guerra-Liera, J.E. Biological responses of hair sheep to a permanent shade during a short heat stress exposure in an arid region. Small Rumin. Res. 2020, 189, 106146. [Google Scholar] [CrossRef]
- Mehaba, N.; Coloma-Garcia, W.; Such, X.; Caja, G.; Salama, A.A. Heat stress affects some physiological and productive variables and alters metabolism in dairy ewes. J. Dairy Sci. 2021, 104, 1099–1110. [Google Scholar] [CrossRef]
- Li, F.K.; Yang, Y.; Jenna, K.; Xia, C.H.; Lv, S.J.; Wei, W.H. Effect of heat stress on the behavioral and physiological patterns of Small-tail Han sheep housed indoors. Trop. Anim. Health Prod. 2018, 50, 1893–1901. [Google Scholar] [CrossRef]
- Sejian, V.; Maurya, V.P.; Naqvi, S.M. Adaptability and growth of Malpura ewes subjected to thermal and nutritional stress. Trop. Anim. Health Prod. 2010, 42, 1763–1770. [Google Scholar] [CrossRef]
- Bernabucci, U.; Lacetera, N.; Danieli, P.P.; Bani, P.; Nardone, A.; Ronchi, B. Influence of different periods of exposure to hot environment on rumen function and diet digestibility in sheep. Int. J. Biometeorol. 2009, 53, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Barnes, A.; Beatty, D.; Taylor, E.; Stockman, C.; Maloney, S.; McCarthy, M. Physiology of heat stress in cattle and sheep. Meat Livest. Aust. 2004, 209, 1–36. [Google Scholar]
- Nicolás-López, P.; Macías-Cruz, U.; Mellado, M.; Correa-Calderón, A.; Meza-Herrera, C.A.; Avendaño-Reyes, L. Growth performance and changes in physiological, metabolic and hematological parameters due to outdoor heat stress in hair breed male lambs finished in feedlot. Int. J. Biometeorol. 2021, 65, 1451–1459. [Google Scholar] [CrossRef] [PubMed]
- Nardone, A.; Ronchi, B.; Valentini, A. Effect of solar radiation on water and food intake and weight gain in “Sarda” and “Comisana” female lambs. In Proceedings of the International Symposium on Animal Husbandry in Warm Climates, Viterbo, Italy, 25–27 October 1990; pp. 149–150. [Google Scholar]
- Ramana, D.B.V.; Pankaj, P.K.; Nikhila, M.; Rani, R.; Sudheer, D. Productivity and physiological responses of sheep exposed to heat stress. J. Agrometeorol. (Spec. Issue) 2013, 15, 71–76. [Google Scholar]
- Yancey, J.W.S.; Apple, J.K.; Kegley, E.B.; Godbee, R.G. Effects of Morinda citrifolia (Noni) pulp on growth performance and stress responses of growing cattle. Prof. Anim. Sci. 2013, 29, 420–425. [Google Scholar] [CrossRef]
- Riva, J.; Rizzi, R.; Marelli, S.; Cavalchini, L.G. Body measurements in Bergamasca sheep. Small Rumin. Res. 2004, 55, 221–227. [Google Scholar] [CrossRef]
- Olayode, B.A.; Sowande, O.S.; Bemiji, M.N.; Bawala, T.O.; Ajagbe, O.O.; Oni, A.O. Effect of feed quality restriction on changes in body measurements and carcass characteristics of West African dwarf sheep. Niger. J. Anim. Prod. 2014, 41, 124–138. [Google Scholar] [CrossRef]
- Rana, M.S.; Hashem, M.A.; Sakib, M.N.; Kumar, A. Effect of heat stress on blood parameters in indigenous sheep. J. Bangladesh Agric. Univ. 2014, 12, 91–94. [Google Scholar] [CrossRef]
- Khalil, F.; Yapati, H.; Al Blallam, Z.; Jose, R. Seasonal effects on growth, physiology, hematology and biochemicalprofiles of Naeemi sheep breed. Adv. Anim. Vet. 2022, 10, 2161–2170. [Google Scholar] [CrossRef]
- McManus, C.M.; Faria, D.A.; Lucci, C.M.; Louvandini, H.; Pereira, S.A.; Paiva, S.R. Heat stress effects on sheep: Are hair sheep more heat resistant? Theriogenology 2020, 155, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, M. From molecular and cellular to integrative heat defense during exposure to chronic heat. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 131, 475–483. [Google Scholar] [CrossRef]
- Bernabucci, U.; Lacetera, N.; Baumgard, L.H.; Rhoads, R.P.; Ronchi, B.; Nardone, A. Metabolic and hormonal acclimation to heat stress in domesticated ruminants. Animal 2010, 4, 1167–1183. [Google Scholar] [CrossRef]
- Colthorpe, & Curlewis. Hypothalamic dopamine D1 receptors are involved in the stimulation of prolactin secretion by high environmental temperature on the female sheep. J. Neuroendocrinol. 1998, 10, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, M.L.; Kim, J.W.; Collier, R.J.; Crooker, B.A.; Boisclair, Y.R.; Baumgard, L.H.; Rhoads, R.P. Effects of heat stress and nutrition on lactating Holstein cows: II. Aspects of hepatic growth hormone responsiveness. J. Dairy Sci. 2010, 93, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Leyva-Corona, J.C.; Reyna-Granados, J.R.; Zamorano-Algandar, R.; Sanchez-Castro, M.A.; Thomas, M.G.; Enns, R.M.; Speidel, S.E.; Medrano, J.F.; Rincon, G.; Luna-Nevarez, P. Polymorphisms within the prolactin and growth hormone/insulin-like growth factor-1 functional pathways associated with fertility traits in Holstein cows raised in a hot-humid climate. Trop. Anim. Health Prod. 2018, 50, 1913–1920. [Google Scholar] [CrossRef]
- Bonilla, A.Q.; Hansen, P.J. 110 Developmental changes in actions of insulin-like growth factor-I in the preimplantation bovine embryo-receptor expression and thermotolerance. Reprod. Fertil. Dev. 2008, 21, 155. [Google Scholar] [CrossRef]
- Satrapa, R.A.; Razza, E.M.; Castilho, A.C.S.; Simões, R.A.L.; Silva, C.F.; Nabhan, T.; Pegorer, M.F.; Barros, C.M. Differential Expression of IGF Family Members in Heat-Stressed Embryos Produced In Vitro from OPU-Derived Oocytes of Nelore (Bos indicus) and Holstein (Bos taurus) Cows. Reprod. Domest. Anim. 2013, 48, 1043–1048. [Google Scholar] [CrossRef]
- De, K.; Kumar, D.; Singh, A.K.; Kumar, K.; Sahoo, A.; Naqvi, S.M.K. Effect of protection against hot climate on growth performance, physiological response and endocrine profile of growing lambs under semi-arid tropical environment. Trop. Anim. Health Prod. 2017, 49, 1317–1323. [Google Scholar] [CrossRef]
- Altin, T.; Yilmaz, M.; Kiral, F.; Yorulmaz, E.; Asici, G.S.E. Effects of the genotype and the seasons on physiological parameters related with adaptability in sheep in Mediterranean climate conditions. Vet. Zootech. 2018, 76, 1–8. [Google Scholar]
- Nazifi, S.; Saeb, M.; Rowghani, E.; Kaveh, K. The influences of thermal stress on serum biochemical parameters of Iranian fat-tailed sheep and their correlation with triiodothyronine (T3), thyroxine (T4) and cortisol concentrations. Comp. Clin. Path 2003, 12, 135–139. [Google Scholar] [CrossRef]
- Rathwa, S.D.; Vasava, A.A.; Pathan, M.M.; Madhira, S.P.; Patel, Y.G.; Pande, A.M. Effect of season on physiological, biochemical, hormonal, and oxidative stress parameters of indigenous sheep. Vet. World 2017, 10, 650–654. [Google Scholar] [CrossRef]
- Pragna, P.; Sejian, V.; Bagath, M.; Krishnan, G.; Archana, P.R.; Soren, N.M.; Beena, V.; Bhatta, R. Comparative assessment of growth performance of three different indigenous goat breeds exposed to summer heat stress. J. Anim. Physiol. Anim. Nutr. 2018, 102, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Le Roith, D.; Bondy, C.; Yakar, S.; Liu, J.L.; Butler, A. The somatomedin hypothesis: 2001. Endoc. Rev. 2001, 22, 53–74. [Google Scholar] [CrossRef]
- Del Vesco, A.P.; Gasparino, E.; Grieser, D.O.; Zancanela, V.; Voltolini, D.M.; Khatlab, A.S.; Guimarães, S.E.; Soares, M.A.; Neto, A.R.O. Effects of methionine supplementation on the expression of protein deposition-related genes in acute heat stress-exposed broilers. PLoS ONE 2015, 10, e0115821. [Google Scholar] [CrossRef]
- Madhusoodan, A.P.; Sejian, V.; Afsal, A.; Bagath, M.; Krishnan, G.; Savitha, S.T.; Rashamol, V.P.; Devaraj, C.; Bhatta, R. Differential expression patterns of candidate genes pertaining to productive and immune functions in hepatic tissue of heat-stressed Salem Black goats. Biol. Rhythm Res. 2021, 52, 809–820. [Google Scholar] [CrossRef]
- Collier, R.J.; Gebremedhin, K.; Macko, A.R.; Roy, K.S. Genes involved in the thermal tolerance of livestock. In Environmental Stress and Amelioration in Livestock Production; Springer Nature: Heidelberg, Germany, 2012; pp. 379–410. [Google Scholar] [CrossRef]
- Alamer, M. The role of prolactin in thermoregulation and water balance during heat stress in domestic ruminants. Asian J. Anim. Vet. Adv. 2011, 6, 1153–1169. [Google Scholar] [CrossRef]
- Hooper, H.B. Physiological Implications, Cellular Responses and Lactational Performance of Saanen Goats Under Heat Stress. Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2019. [Google Scholar] [CrossRef]
- Mullur, R.; Liu, Y.Y.; Brent, G.A. Thyroid hormone regulation of metabolism. Physiol. Rev. 2014. [Google Scholar] [CrossRef]
- Weitzel, J.M.; Viergutz, T.; Albrecht, D.; Bruckmaier, R.; Schmicke, M.; Tuchscherer, A.; Koch, F.; Kuhla, B. Hepatic thyroid signaling of heat-stressed late pregnant and early lactating cows. J. Endocrinol. 2017, 234, 129–141. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time of Recording | DBT (°C) | WBT (°C) | THI | |
|---|---|---|---|---|
| Thermoneutral zone chamber | Morning (8:00) | 21.78 ± 0.09 | 20.80 ± 0.10 | 71.25 ± 0.13 |
| Afternoon (14:00) | 20.15 ± 0.05 | 19.19 ± 0.07 | 68.93 ± 0.07 | |
| Heating chamber | Morning (8:00) | 22.49 ± 0.08 | 21.49 ± 0.08 | 72.27 ± 0.12 |
| Afternoon (14:00) | 40.44 ± 0.18 | 32.81 ± 0.31 | 93.34 ± 0.34 |
| Attribute | Concentrate Mixture (kg/100 kg) | Napier Hay (Pennisetum purpureum) |
|---|---|---|
| Ingredients | ||
| Maize | 36 | - |
| Wheat bran | 37 | - |
| Soybean meal | 25 | - |
| Mineral mixture | 1.5 | - |
| Salt | 0.5 | - |
| Chemical composition (%) | ||
| Dry matter | 90.83 | 92.32 |
| Organic matter | 94.02 | 88.59 |
| Crude protein | 23.15 | 6.84 |
| Ether extract | 3.77 | 2.72 |
| Crude fibre | 6.12 | 32.58 |
| Total ash | 5.98 | 11.41 |
| Fibre fractions (%) | ||
| Neutral detergent fibre | 29.13 | 71.51 |
| Acid detergent fibre | 8.97 | 43.94 |
| Nutritive value | ||
| Total digestible nutrients (%) | 71 | 53.20 |
| Digestible energy (kJ/kg) | 14.24 | 9.78 |
| Metabolisable energy (kJ/kg) | 11.78 | 7.98 |
| Nutrient | Composition (100 g) |
|---|---|
| Ocimum sanctum | 30 g |
| Emblica officinalis | 30 g |
| Morinda citrifolia | 20 g |
| Withania somnifera | 10 g |
| Phyllostachys edulis | 10 g |
| Dose rate | 0.8 g/Kg BW/Day |
| Gene ID | Primer Sequence (5′–3′) | Product Size (bp) | Accession No. | |
|---|---|---|---|---|
| GAPDH | F R | GGTGATGCTGGTGCTGAGTAG ACGATGTCCACTTTGCCAGT | 147 | XM_005680968.3 |
| GHR | F R | AACCACCACCCAATACAG CAACGAGTACATCGGAAC | 134 | NM_001009323 |
| IGF1 | F R | AGCAGTCTTCCAACCCAA ACATCTCCAGCCTCCTCA | 85 | NM_001009774 |
| PRLR | F R | CCCCTTGTTCTCTGCTAAACCC CTATCCGTCACCCGAGACACC | 129 | O46561-1 |
| THRA | F R | TATCGCTGCATCACTTGCGA CTCAATCAGCTTGCGCTTGG | 225 | NM_001100919.3 |
| Variable | Temperature (T) | Diet | Day | Significance | ||||
|---|---|---|---|---|---|---|---|---|
| 0 | 15 | 30 | 45 | 60 | ||||
| Feed intake (g/w0.75/day) | Thermoneutral | Control | 50.5 ± 0.38 | 47.3 ± 1.33 | 49.1 ± 1.24 | 49.1 ± 1.17 | 50.3 ± 0.91 | Temperature.Day (p < 0.001); Diet.Day (p < 0.01), Temperature.Diet.Day (p < 0.01) |
| Herb | 51.9 ± 0.38 | 48.4 ± 1.33 | 51.5 ± 1.24 | 49.8 ± 1.17 | 51.4 ± 0.91 | |||
| Heat stress | Control | 50.9 ± 0.38 | 50.8 ± 1.33 | 51.7 ± 1.24 | 49.1 ± 1.17 | 51.4 ± 0.91 | ||
| Herb | 49.1 ± 0.38 | 49.3 ± 1.33 | 54.0 ± 1.24 | 54.1 ± 1.17 | 52.6 ± 0.91 | |||
| Day effect | 50.61 ± 0.19 | 48.95 ± 0.65 | 51.55 ± 0.59 | 50.54 ± 0.60 | 51.43 ± 0.39 | Day (p < 0.01) | ||
| Diet effect | 50.03 vs. 51.20 | Diet (p < 0.05) | ||||||
| Temperature effect | 49.93 vs. 51.30 | Temperature (p < 0.01) | ||||||
| Water intake (L/Kg DMI/day) | Thermoneutral | Control | 3.33 ± 0.61 | 5.00 ± 0.55 | 4.32 ± 0.49 | 4.23 ± 0.57 | 3.94 ± 0.47 | Temperature.Day (p < 0.001) |
| Herb | 3.50 ± 0.61 | 4.67 ± 0.55 | 4.67 ± 0.49 | 5.61 ± 0.57 | 4.98 ± 0.47 | |||
| Heat stress | Control | 3.47 ± 0.61 | 7.57 ± 0.55 | 7.28 ± 0.49 | 7.49 ± 0.57 | 6.60 ± 0.47 | ||
| Herb | 3.46 ± 0.61 | 7.27 ± 0.55 | 7.31 ± 0.49 | 7.33 ± 0.57 | 7.29 ± 0.47 | |||
| Day effect | 3.44 ± 0.30 | 6.13 ± 0.27 | 5.95 ± 0.25 | 6.17 ± 0.29 | 5.70 ± 0.24 | Day (p < 0.001) | ||
| Diet effect | 5.32 vs. 5.63 | NS | ||||||
| Temperature effect | 4.45 vs. 6.51 | Temperature (p < 0.001) | ||||||
| Variable | Temperature (T) | Diet | Day | Significance | ||||
| 0 | 15 | 30 | 45 | 60 | Diet.Day (p < 0.01) | |||
| Body weight (Kg) | Thermoneutral | Control | 18.8 ± 0.90 | 19.5 ± 0.80 | 20.0 ± 0.79 | 21.0 ± 0.80 | 22.2 ± 0.82 | |
| Herb | 19.1 ± 0.90 | 20.4 ± 0.80 | 21.4 ± 0.79 | 22.2 ± 0.80 | 23.7 ± 0.82 | |||
| Heat stress | Control | 19.2 ± 0.90 | 20.4 ± 0.80 | 20.6 ± 0.79 | 22.1 ± 0.80 | 22.6 ± 0.82 | ||
| Herb | 18.8 ± 0.90 | 19.7 ± 0.80 | 21.1 ± 0.79 | 21.7 ± 0.80 | 23.1 ± 0.82 | |||
| Day effect | 18.97 ± 0.45 | 19.98 ± 0.41 | 20.80 ± 0.40 | 21.75 ± 0.40 | 22.90 ± 0.42 | Day (p < 0.001) | ||
| Diet effect | 20.64 vs. 21.12 | NS | ||||||
| Temperature effect | 20.83 vs. 20.93 | NS | ||||||
| Variable | Temperature (T) | Diet | Day | Significance | ||||
|---|---|---|---|---|---|---|---|---|
| 0 | 15 | 30 | 45 | 60 | ||||
| Body Length (cm) | Thermoneutral | Control | 61.2 ± 2.25 | 57.8 ± 1.97 | 63.3 ± 2.13 | 61.8 ± 1.40 | 62.5 ± 1.39 | Diet.Day (p < 0.05) |
| Herb | 60.2 ± 2.25 | 63.0 ± 1.97 | 63.2 ± 2.13 | 61.8 ± 1.40 | 62.5 ± 1.39 | |||
| Heat stress | Control | 60.7 ± 2.25 | 58.3 ± 1.97 | 63.5 ± 2.13 | 64.5 ± 1.40 | 65.2 ± 1.39 | ||
| Herb | 63.8 ± 2.25 | 59.0 ± 1.97 | 61.2 ± 2.13 | 61.7 ± 1.40 | 62.0 ± 1.39 | |||
| Day effect | 61.46 ± 1.13 | 59.54 ± 0.99 | 62.80 ± 1.07 | 62.46 ± 0.70 | 63.04 ± 0.70 | Day (p < 0.001) | ||
| Diet effect | 61.88 vs. 61.83 | NS | ||||||
| Temperature effect | 61.73 vs. 61.98 | NS | ||||||
| Body Height (cm) | Thermoneutral | Control | 60.7 ± 2.11 | 64.0 ± 1.67 | 63.3 ± 1.56 | 64.3 ± 1.00 | 64.0 ± 1.00 | NS |
| Herb | 59.3 ± 2.11 | 63.2 ± 1.67 | 63.2 ± 1.56 | 64.7 ± 1.00 | 64.8 ± 1.00 | |||
| Heat stress | Control | 62.0 ± 2.11 | 65.0 ± 1.67 | 65.2 ± 1.56 | 64.7 ± 1.00 | 64.8 ± 1.00 | ||
| Herb | 61.3 ± 2.11 | 66.0 ± 1.67 | 63.0 ± 1.56 | 64.5 ± 1.00 | 64.7 ± 1.00 | |||
| Day effect | 60.83 ± 1.06 | 64.54 ± 0.83 | 63.29 ± 0.78 | 64.54 ± 0.50 | 64.58 ± 0.50 | Day (p < 0.01) | ||
| Diet effect | 63.80 vs. 63.32 | NS | ||||||
| Temperature effect | 63.0 vs. 64.12 | NS | ||||||
| Chest Girth (cm) | Thermoneutral | Control | 67.7 ± 1.89 | 66.8 ± 1.84 | 67.3 ± 1.79 | 68.7 ± 1.73 | 69.2 ± 1.82 | NS |
| Herb | 69.7 ± 1.89 | 68.7 ± 1.84 | 68.3 ± 1.79 | 70.0 ± 1.73 | 70.0 ± 1.82 | |||
| Heat stress | Control | 68.8 ± 1.89 | 68.2 ± 1.84 | 69.3 ± 1.79 | 69.5 ± 1.73 | 70.2 ± 1.82 | ||
| Herb | 64.2 ± 1.89 | 68.7 ± 1.84 | 66.0 ± 1.79 | 67.5 ± 1.73 | 68.2 ± 1.82 | |||
| Day effect | 67.58 ± 0.95 | 67.17 ± 0.92 | 67.75 ± 0.89 | 68.92 ± 0.87 | 69.34 ± 0.91 | Day (p < 0.001) | ||
| Diet effect | 68.57 vs. 67.75 | NS | ||||||
| Temperature effect | 68.63 vs. 67.68 | NS | ||||||
| Variable | Temperature (T) | Diet | Day | Significance | ||||
|---|---|---|---|---|---|---|---|---|
| 0 | 15 | 30 | 45 | 60 | ||||
| GH (ng/mL) | Thermoneutral | Control | 2.70 ± 0.65 | 3.07 ± 0.48 | 2.82 ± 0.53 | 2.51 ± 0.47 | 2.74 ± 0.47 | Diet.Day (p < 0.001) |
| Herb | 2.90 ± 0.65 | 2.76 ± 0.48 | 3.71 ± 0.53 | 4.41 ± 0.47 | 5.14 ± 0.47 | |||
| Heat stress | Control | 2.74 ± 0.65 | 1.97 ± 0.48 | 2.07 ± 0.53 | 1.43 ± 0.47 | 1.17 ± 0.47 | ||
| Herb | 2.81 ± 0.65 | 2.87 ± 0.48 | 3.94 ± 0.53 | 4.54 ± 0.47 | 4.96 ± 0.47 | |||
| Day effect | 2.79 ± 0.33 | 2.67 ± 0.24 | 3.13 ± 0.26 | 3.22 ± 0.23 | 3.50 ± 0.24 | Day (p < 0.001) | ||
| Diet effect | 2.32 vs. 3.80 | Diet (p < 0.01) | ||||||
| Temperature effect | 3.28 vs. 2.85 | NS | ||||||
| IGF-1 (ng/mL) | Thermoneutral | Control | 48.8 ± 6.47 | 46.5 ± 6.81 | 59.0 ± 5.24 | 52.8 ± 5.32 | 44.8 ± 5.46 | Diet.Day (p < 0.001) |
| Herb | 49.7 ± 6.47 | 49.7 ± 6.81 | 62.1 ± 5.24 | 66.2 ± 5.32 | 73.6 ± 5.46 | |||
| Heat stress | Control | 47.1 ± 6.47 | 42.2 ± 6.81 | 39.7 ± 5.24 | 40.8 ± 5.32 | 34.7 ± 5.46 | ||
| Herb | 49.0 ± 6.47 | 51.9 ± 6.81 | 58.2 ± 5.24 | 66.0 ± 5.32 | 72.9 ± 5.46 | |||
| Day effect | 48.63 ± 3.25 | 48.71 ± 3.41 | 54.76 ± 2.63 | 56.45 ± 2.65 | 56.48 ± 2.72 | Day (p < 0.01) | ||
| Diet effect | 45.63 vs. 60.38 | Diet (p < 0.01) | ||||||
| Temperature effect | 55.77 vs. 50.25 | NS | ||||||
| T3 (nmol/L) | Thermoneutral | Control | 25.7 ± 1.55 | 27.5 ± 0.88 | 25.1 ± 1.09 | 27.5 ± 0.90 | 27.3 ± 0.81 | Temperature.Day (p < 0.001); Temperature.Diet (p < 0.001); Diet.Day (p < 0.001) |
| Herb | 26.3 ± 1.55 | 29.7 ± 0.88 | 30.7 ± 1.09 | 33.5 ± 0.90 | 34.3 ± 0.81 | |||
| Heat stress | Control | 25.6 ± 1.55 | 23.3 ± 0.88 | 19.9 ± 1.09 | 14.2 ± 0.90 | 11.2 ± 0.81 | ||
| Herb | 25.7 ± 1.55 | 29.9 ± 0.88 | 31.6 ± 1.09 | 32.6 ± 0.90 | 33.7 ± 0.81 | |||
| Day effect | 25.84 ± 0.89 | 27.61 ± 0.48 | 26.85 ± 0.54 | 26.97 ± 0.51 | 26.62 ± 0.56 | NS | ||
| Diet effect | 22.74 vs. 30.82 | Diet (p < 0.001) | ||||||
| Temperature effect | 28.77 vs. 24.79 | Temperature (p < 0.001) | ||||||
| T4 (nmol/L) | Thermoneutral | Control | 96.0 ± 6.33 | 108 ± 5.74 | 108 ± 5.94 | 112 ± 6.52 | 114 ± 6.88 | Temperature.Diet (p < 0.05); Temperature.Day (p < 0.01); Diet.Day (p < 0.001); Temperature.Diet.Day (p < 0.01) |
| Herb | 98.6 ± 6.33 | 115 ± 5.74 | 129 ± 5.94 | 147 ± 6.52 | 160 ± 6.88 | |||
| Heat stress | Control | 99.8 ± 6.33 | 88.8 ± 5.74 | 72.8 ± 5.94 | 69.9 ± 6.52 | 58.7 ± 6.88 | ||
| Herb | 97.7 ± 6.33 | 112 ± 5.74 | 124 ± 5.94 | 141 ± 6.52 | 153 ± 6.88 | |||
| Day effect | 98.02 ± 3.56 | 105.86 ± 2.91 | 108.42 ± 2.91 | 117.62 ± 3.23 | 121.28 ± 3.60 | Day (p < 0.001) | ||
| Diet effect | 92.76 vs. 127.71 | Diet (p < 0.001) | ||||||
| Temperature effect | 118.78 vs. 101.70 | Temperature (p < 0.05) | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebez, E.B.; Devaraj, C.; Ninan, J.; Silpa, M.V.; Perumal, S.V.; Sahoo, A.; Dunshea, F.R.; Sejian, V. Effects of Herbal Supplementation on Growth Performance of Kenguri Sheep Exposed to Heat Stress. Animals 2025, 15, 1285. https://doi.org/10.3390/ani15091285
Rebez EB, Devaraj C, Ninan J, Silpa MV, Perumal SV, Sahoo A, Dunshea FR, Sejian V. Effects of Herbal Supplementation on Growth Performance of Kenguri Sheep Exposed to Heat Stress. Animals. 2025; 15(9):1285. https://doi.org/10.3390/ani15091285
Chicago/Turabian StyleRebez, Ebenezer Binuni, Chinnasamy Devaraj, Jacob Ninan, Mullakkalparambil Velayudhan Silpa, Shanmugam Venkatesa Perumal, Artabandhu Sahoo, Frank Rowland Dunshea, and Veerasamy Sejian. 2025. "Effects of Herbal Supplementation on Growth Performance of Kenguri Sheep Exposed to Heat Stress" Animals 15, no. 9: 1285. https://doi.org/10.3390/ani15091285
APA StyleRebez, E. B., Devaraj, C., Ninan, J., Silpa, M. V., Perumal, S. V., Sahoo, A., Dunshea, F. R., & Sejian, V. (2025). Effects of Herbal Supplementation on Growth Performance of Kenguri Sheep Exposed to Heat Stress. Animals, 15(9), 1285. https://doi.org/10.3390/ani15091285