
Comparative Expression Analysis of Innate Immune Markers and Phagocytic Activity in Peripheral Blood of Dogs with Mammary Tumors

 , , , ,
, , , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Experimental Samples
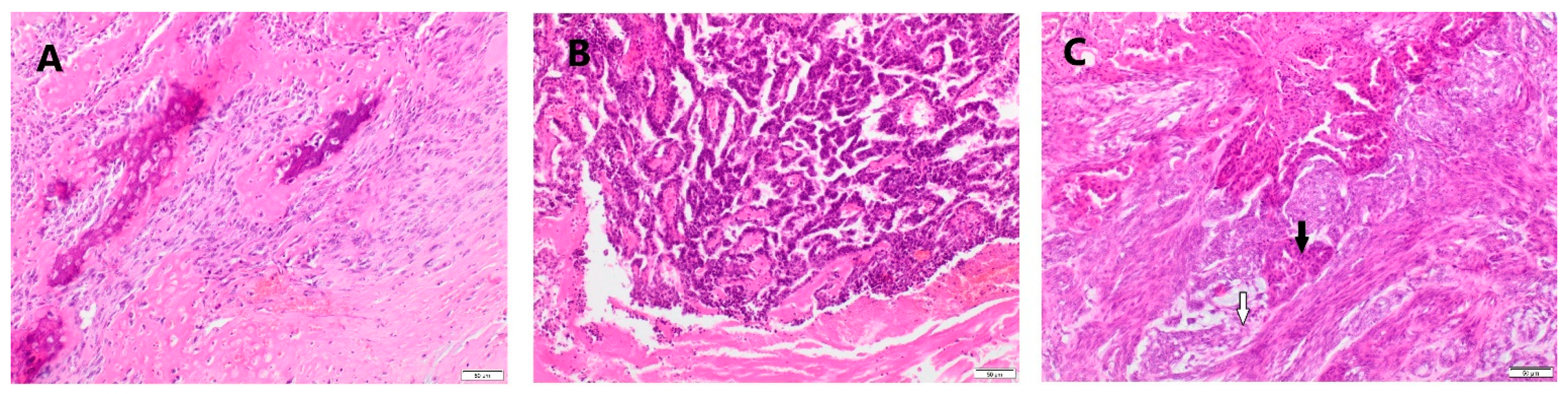
2.2. Histological Examinations of Tumor Samples
2.3. Flow Cytometric Immunophenotyping
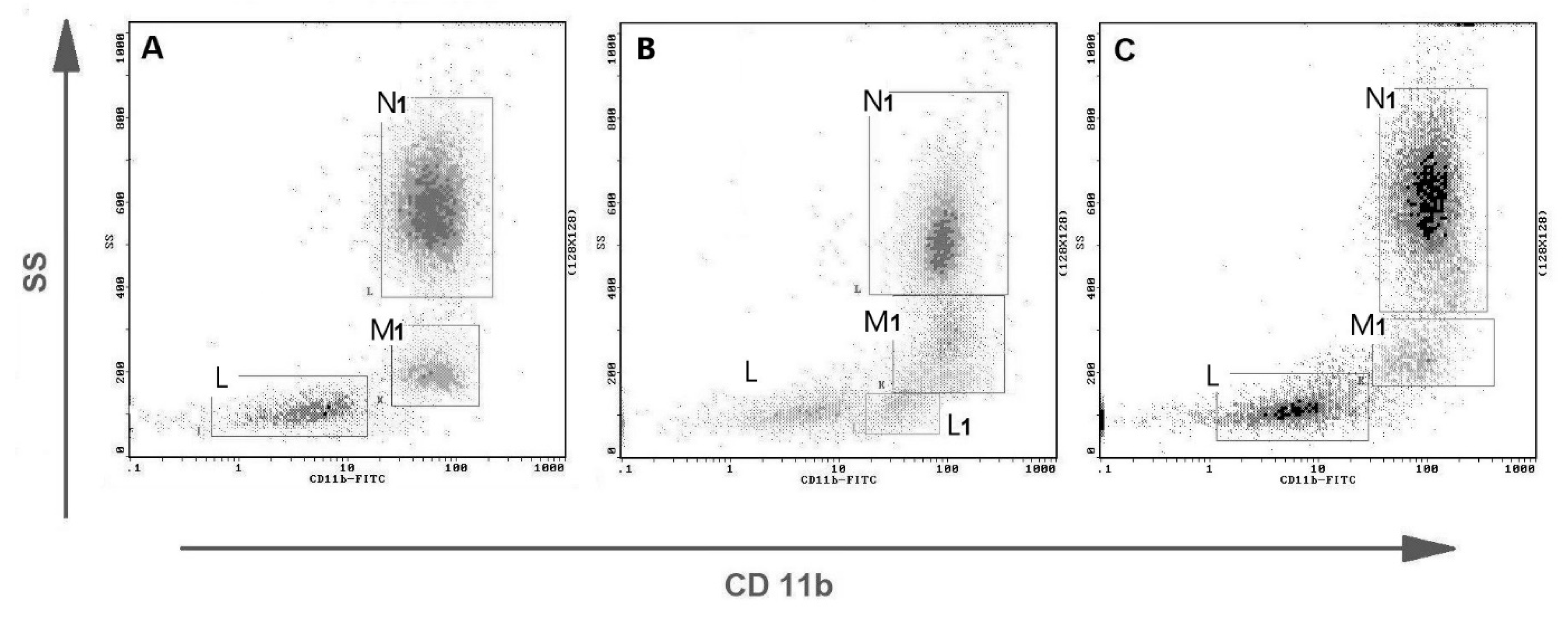
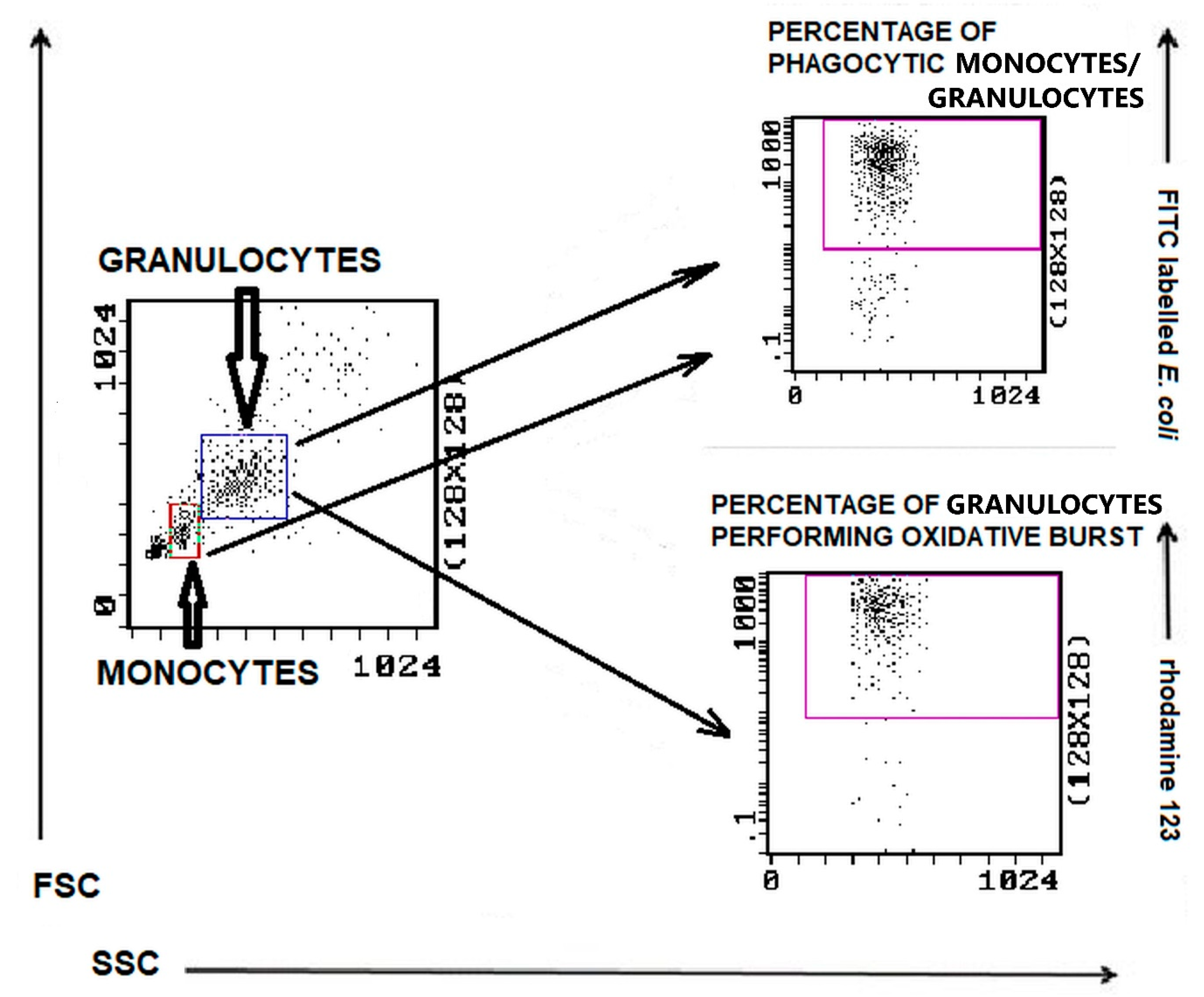
2.4. Phagocytosis Determination by Flow Cytometry
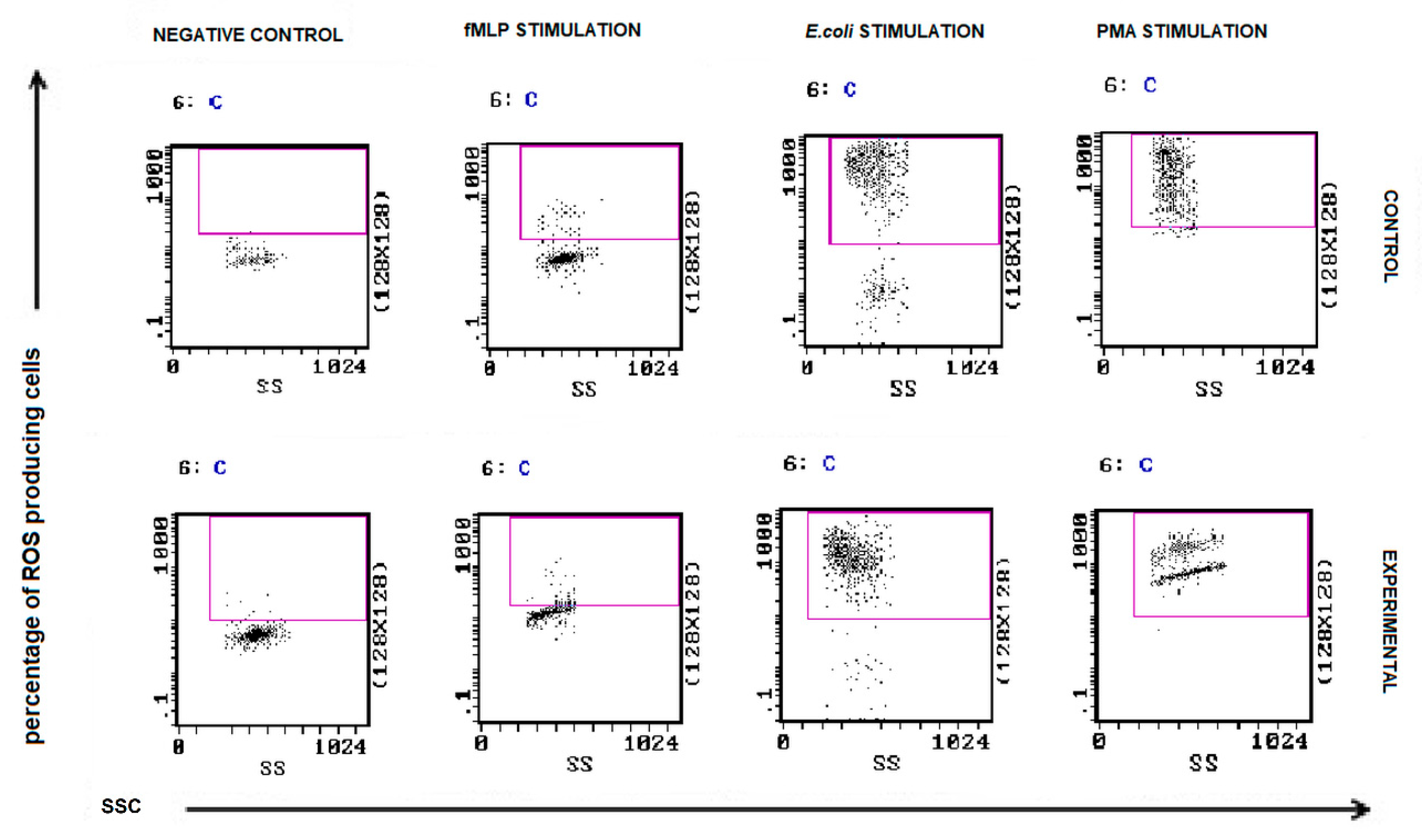
2.5. Measurement of Reactive Oxygen Species (ROS Assay) by Flow Cytometry
2.6. Statistical Analysis
3. Results
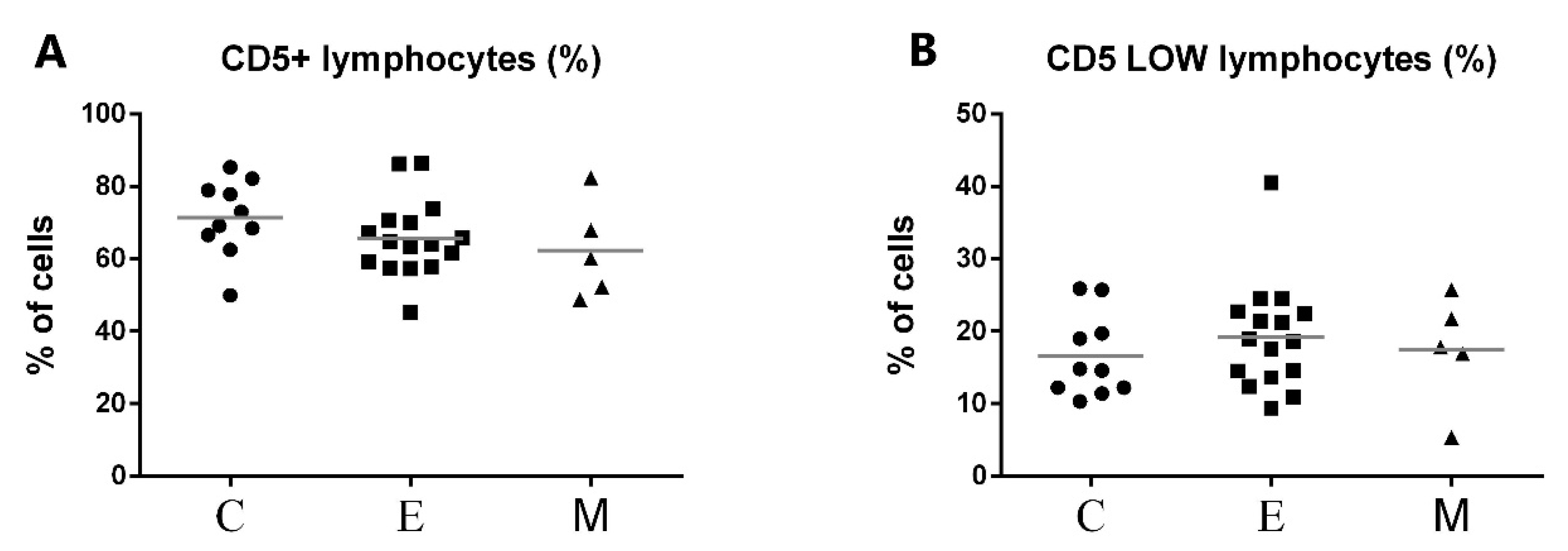
3.1. Immunophenotyping Results
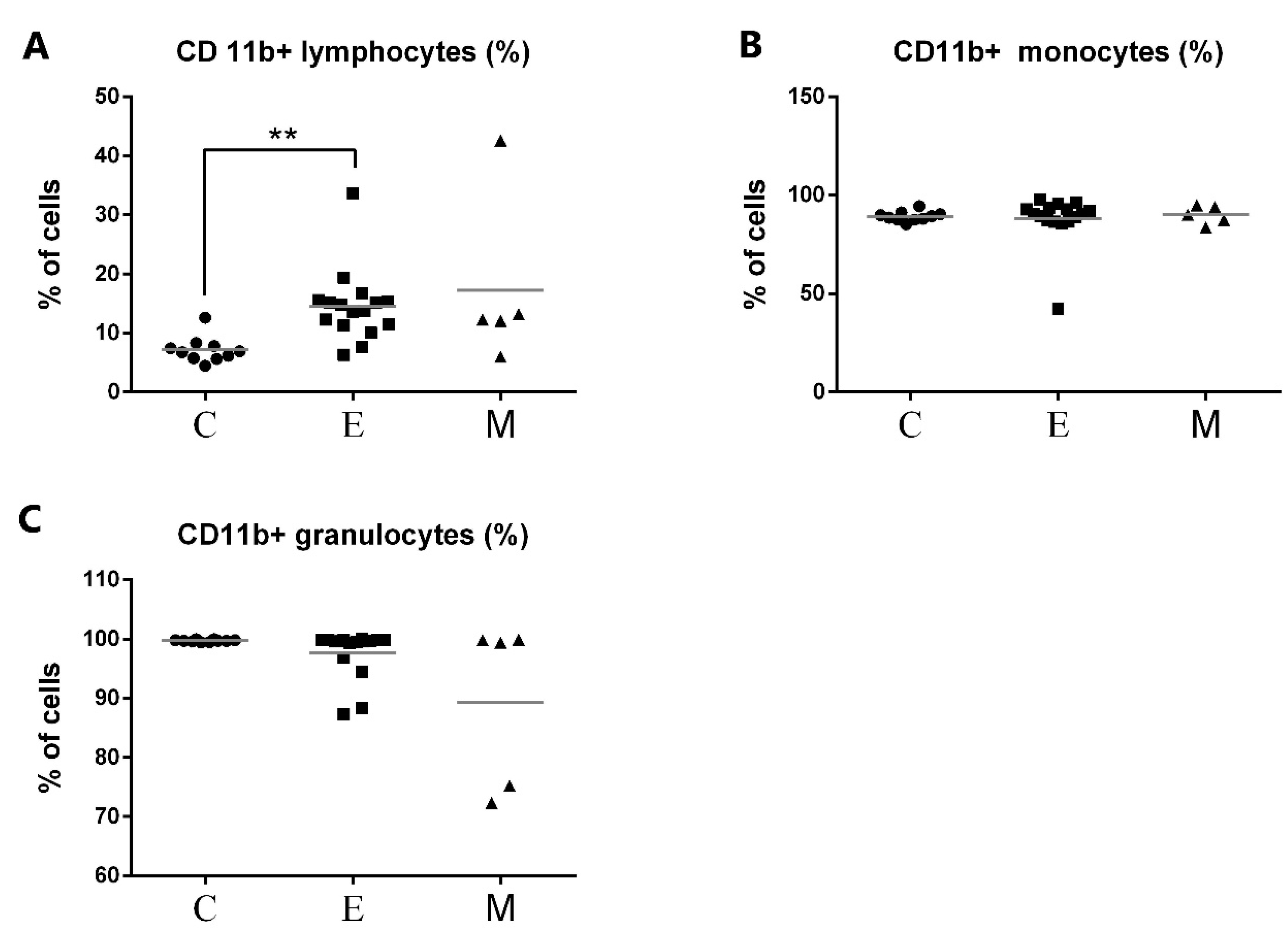
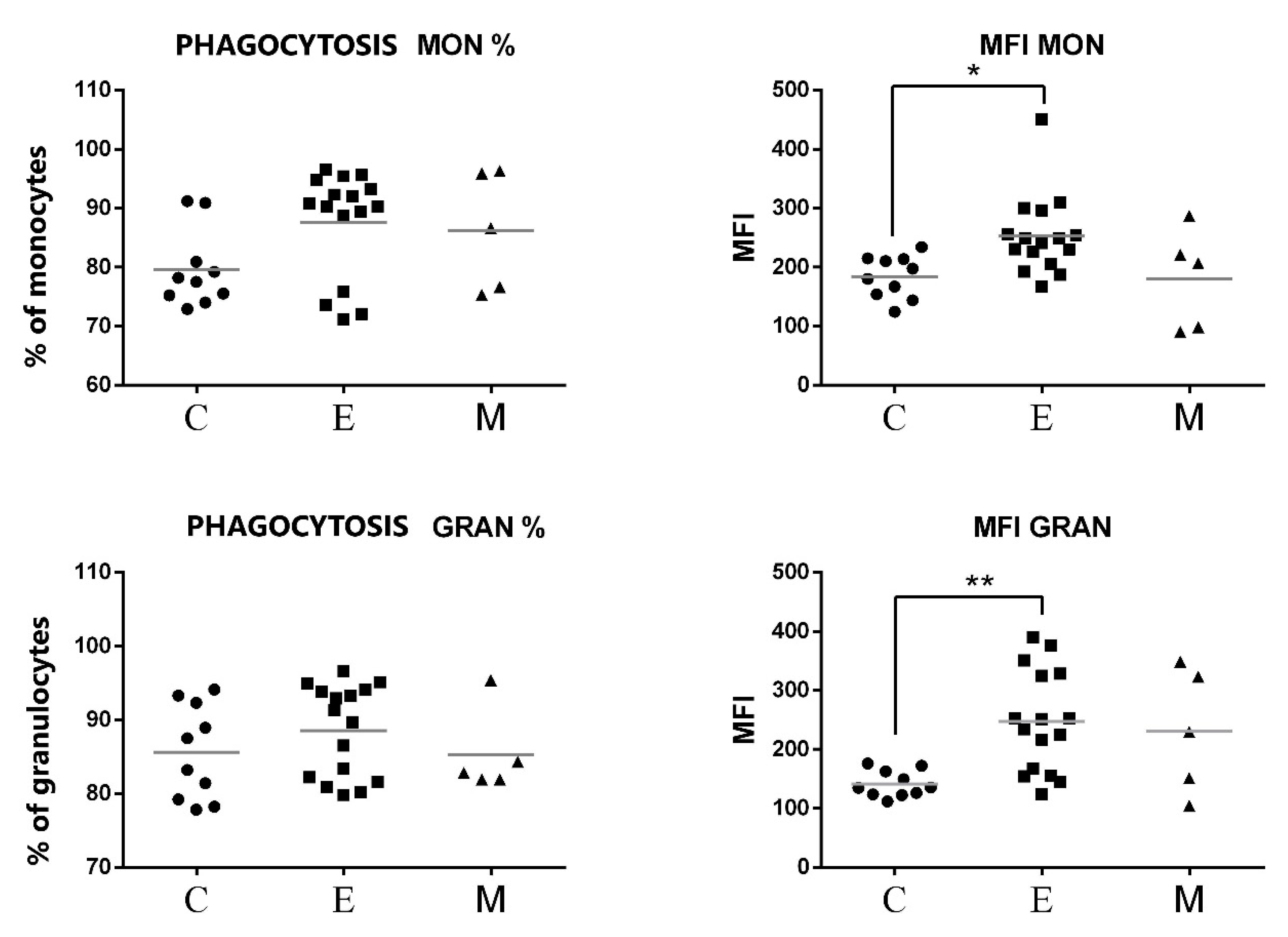
3.2. Phagocytic Activity of Granulocytes and Monocytes
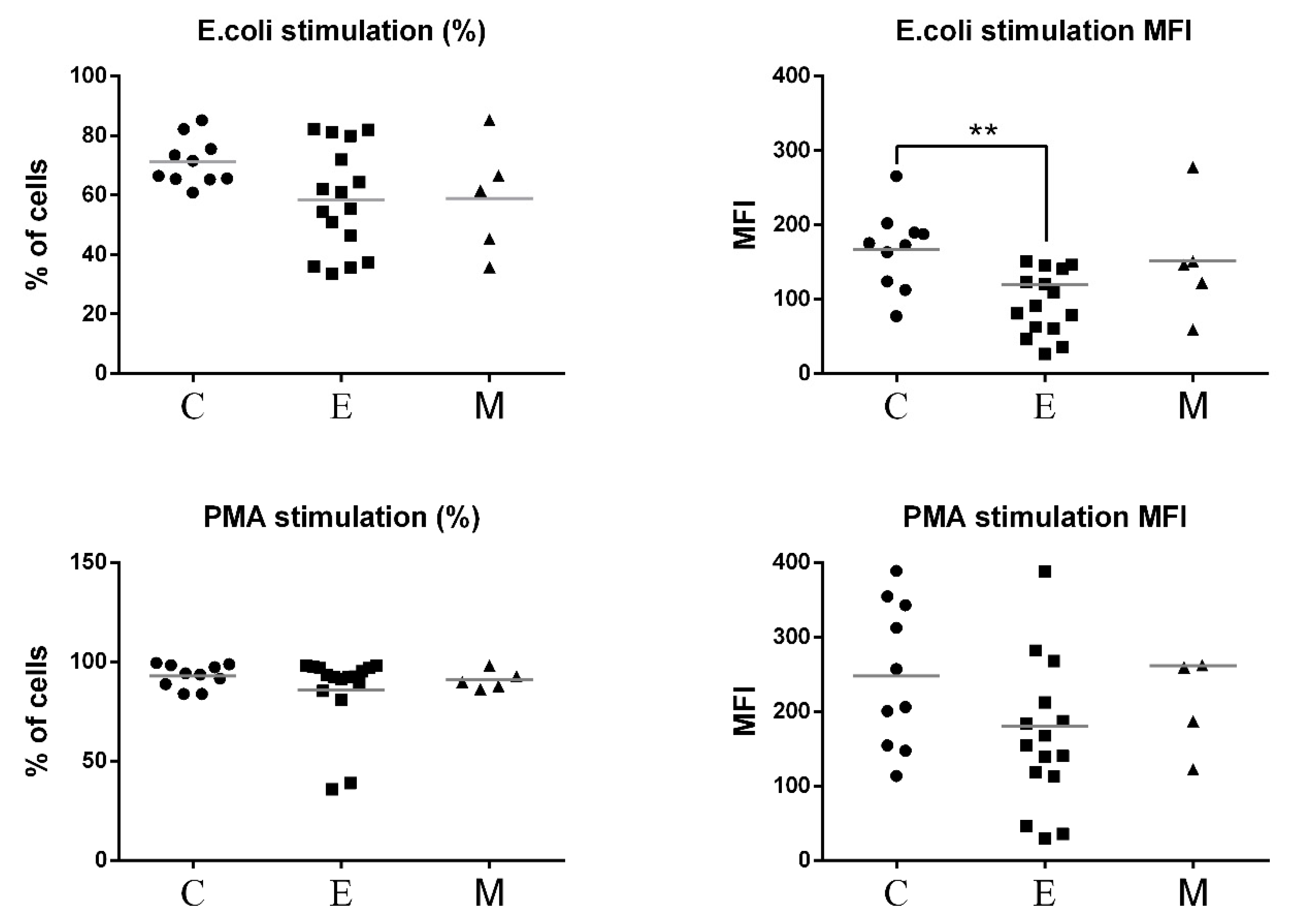
3.3. Neutrophil Oxidative Burst
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Peña, L.; Gama, A.; Goldschmidt, M.H.; Abadie, J.; Benazzi, C.; Castagnaro, M.; Díez, L.; Gärtner, F.; Hellmén, E.; Kiupel, M.; et al. Canine Mammary Tumors: A Review and Consensus of Standard Guidelines on Epithelial and Myoepithelial Phenotype Markers, HER2, and Hormone Receptor Assessment Using Immunohistochemistry. Vet. Pathol. 2014, 51, 127–145. [Google Scholar] [CrossRef]
- Queiroga, F.L.; Raposo, T.; Carvalho, M.I.; Prada, J.; Pires, I. Canine Mammary Tumours as a Model to Study Human Breast Cancer: Most Recent Findings. In Vivo 2011, 25, 455–465. [Google Scholar] [PubMed]
- Hiam-Galvez, K.J.; Allen, B.M.; Spitzer, M.H. Systemic immunity in cancer. Nat. Rev. Cancer 2021, 21, 345–359. [Google Scholar] [CrossRef]
- Bremnes, R.M.; Al-Shibli, K.; Donnem, T.; Sirera, R.; Al-Saad, S.; Andersen, S.; Stenvold, H.; Camps, C.; Busund, L.T. The Role of Tumor-Infiltrating Immune Cells and Chronic Inflammation at the Tumor Site on Cancer Development, Progression, and Prognosis: Emphasis on Non-small Cell Lung Cancer. J. Thorac. Oncol. 2011, 6, 824–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dongre, A.; Rashidian, M.; Reinhardt, F.; Bagnato, A.; Keckesova, Z.; Ploegh, H.L.; Weinberg, R.A. Epithelial-to-Mesenchymal Transition Contributes to Immunosuppression in Breast Carcinomas. Cancer Res. 2017, 77, 3982–3989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.J.; Park, J.Y.; Jang, Y.Y.; Lee, J.J.; Lee, Y.K.; Shin, M.G.; Jung, J.Y.; Carson, W.E.; Cho, D.; Kim, S.K. Ex vivo expansion of canine cytotoxic large granular lymphocytes exhibiting characteristics of natural killer cells. Vet. Immunol. Immunopathol. 2013, 153, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Shin, D.J.; Kim, Y.; Kim, C.J.; Lee, J.J.; Yoon, M.S.; Uong, T.N.T.; Yu, D.; Jung, J.Y.; Cho, D.; et al. Comparison of Phenotypic and Functional Characteristics between Canine Non-B, Non-T Natural Killer Lymphocytes and CD3+CD5dimCD21− Cytotoxic Large Granular Lymphocytes. Front. Immunol. 2018, 9, 841. [Google Scholar] [CrossRef] [Green Version]
- Michael, H.T.; Ito, D.; McCullar, V.; Zhang, B.; Miller, J.S.; Modiano, J.F. Isolation and characterization of canine natural killer cells. Vet. Immunol. Immunopathol. 2013, 155, 211–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.S.; Chang, C.; Chiang, H.; Chuang, T.; Hsu, C.H.; Liu, C.C. Activating natural killer (NK) cytotoxicity of canine CD5−CD21− cells requires low surface CD5 density NK cells. Iran. J. Vet. Res. 2018, 19, 87–95. [Google Scholar] [PubMed]
- Kim, Y.; Lee, S.; Kim, C.; Lee, J.; Yu, D.; Ahn, S.; Shin, D.; Kim, S. Canine non-B, non-T NK lymphocytes have a potential antibody-dependent cellular cytotoxicity function against antibody-coated tumor cells. BMC Vet. Res. 2019, 15, 339. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.C.; Hung, S.W.; Jan, T.R.; Liao, K.W.; Cheng, C.H.; Wang, Y.S.; Chu, R.M. CD5-low expression lymphocytes in canine peripheral blood show characteristics of natural killer cells. J. Leukoc. Biol. 2008, 84, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Knapp, D.H.; Leibnitz, R.R.; DeNicola, B.D.; Turek, J.J.; Teclaw, R.; Shaffer, L.; Chan, C.K. Measurement of NK activity in effector cells purified from canine peripheral lymphocytes. Vet. Immunol. Immunopathol. 1993, 35, 239–251. [Google Scholar] [CrossRef]
- Krneta, T.; Gillgrass, A.; Chew, M.; Ashkar, A.A. The breast tumor microenvironment alters the phenotype and function of natural killer cells. Cell. Mol. Immunol. 2016, 13, 628–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gingrich, A.A.; Modiano, J.F.; Canter, R.J. Characterization and Potential Applications of Dog Natural Killer Cells in Cancer Immunotherapy. J. Clin. Med. 2019, 8, 1802. [Google Scholar] [CrossRef] [Green Version]
- Jia, W.; Li, H.; He, Y.W. The extracellular matrix protein mindin serves as an integrin ligand and is critical for inflammatory cell recruitment. Blood 2005, 106, 3854–3859. [Google Scholar] [CrossRef]
- Taszkun, I. Assessment of CD11b and CD11/18 integrin expression on leukocytes of dogs with atopic dermatitis. Bull. Vet. Inst. Pulawy 2014, 58, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Goulart, M.R.; Pluhar, G.E.; Ohlfest, J.R. Identification of myeloid derived suppressor cells in dogs with naturally occurring cancer. PLoS ONE. 2012, 7, e33274. [Google Scholar] [CrossRef] [Green Version]
- Geraghty, T.; Rajagopalan, A.; Aslam, R.; Pohlman, A.; Venkatesh, I.; Zloza, A.; Cimbaluk, D.; DeNardo, D.G.; Gupta, V. Positive Allosteric Modulation of CD11b as a Novel Therapeutic Strategy Against Lung Cancer. Front Oncol. 2020, 10, 748. [Google Scholar] [CrossRef]
- Liu, J.; Lin, P.C.; Zhou, B.P. Inflammation fuels tumor progress and metastasis. Curr. Pharm. Des. 2015, 21, 3032–3040. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Saxena, S.; Awaji, M.; Singh, R.K. Tumor-Associated Neutrophils in Cancer: Going Pro. Cancers 2019, 11, 564. [Google Scholar] [CrossRef] [Green Version]
- Snoderly, H.T.; Boone, B.A.; Bennewitz, M.F. Neutrophil extracellular traps in breast cancer and beyond: Current perspectives on NET stimuli, thrombosis and metastasis, and clinical utility for diagnosis and treatment. Breast Cancer Res. 2019, 21, 145. [Google Scholar] [CrossRef] [Green Version]
- Cools-Lartigue, J.; Spicer, J.; McDonald, B.; Gowing, S.; Chow, S.; Giannias, B.; Bourdeau, F.; Kubes, P.; Ferri, L. Neutrophil extracellular traps sequester circulating tumor cells and promote metastasis. J. Clin. Investig. 2013, 123, 3446–3458. [Google Scholar] [CrossRef]
- Gentles, A.J.; Newman, A.M.; Liu, C.L.; Bratman, S.V.; Feng, W.; Kim, D.; Nair, V.S.; Xu, Y.; Khuong, A.; Hoang, C.D.; et al. The prognostic landscape of genes and infiltrating immune cells across human cancers. Nat. Med. 2015, 21, 938–945. [Google Scholar] [CrossRef]
- Matlung, H.L.; Babes, L.; Zhao, X.W.; van Houdt, M.; Treffers, L.W.; van Rees, D.J.; Franke, K.; Schornagel, K.; Verkuijlen, P.; Janssen, H.; et al. Neutrophils Kill Antibody-Opsonized Cancer Cells by Trogoptosis. Cell Rep. 2018, 23, 3946–3959. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Qin, W.; Song, M.; Liu, L.; Yu, Y.; Qi, X.; Sun, H. Neutrophil Suppresses Tumor Cell Proliferation via Fas /Fas Ligand Pathway Mediated Cell Cycle Arrested. Int. J. Biol. Sci. 2018, 14, 2103–2113. [Google Scholar] [CrossRef] [PubMed]
- Riondato, F.; Comazzi, S. Flow Cytometry in the Diagnosis of Canine B-Cell Lymphoma. Front. Vet. Sci. 2021, 8, 600986. [Google Scholar] [CrossRef]
- Cassali, G.D.; Salvador, A.; Freitas, C.; Dutra, A.P.; Schmitt, F.C. DNA flow cytometry of canine mammary tumors: Comparative aspects with human breast tumors. Arq. Bras. Med. Vet. Zootec. 2007, 59, 1163–1168. [Google Scholar] [CrossRef]
- Misdorp, W.; Else, R.W.; Hellmen, E.; Lipscom, T.P. Histological classification of the mammary tumours of the dog and the cat. In World Health Organization International Histologic Classification of Tumours of Domestic Animals; Shulman, F.O., Ed.; Armed Forces Institute of Pathology: Washington, DC, USA, 1999; pp. 16–29. [Google Scholar]
- Goldschmidt, M.; Peña, L.; Rasotto, R.; Zappulli, V. Classification and grading of canine mammary tumors. Vet. Pathol. 2011, 48, 117–131. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, C.J.; LeBlanc, A.K.; Jones, M.M.; Bartges, J.W.; Kania, S.A. Evaluation of peripheral blood neutrophil function intumor-bearing dogs. Vet. Clin. Pathol. 2010, 39, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Kzhyshkowska, J.; Bugert, P. Innate Immune System for Diagnostics and Therapy: Progress in Fundamental Knowledge and Clinical Application. Transfus. Med. Hemother. 2016, 43, 63–64. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.C.; Huang, Y.C.; Wang, Y.S.; Juang, R.H.; Liao, K.W.; Chu, R.M. Canine CD8 T cells showing NK cytotoxic activity express mRNAs for NK cell-associated surface molecules. Vet. Immunol. Immunopathol. 2010, 133, 144–153. [Google Scholar] [CrossRef]
- Saito, H.; Osaki, T.; Ikeguchi, M. Decreased NKG2D expression on NK cells correlates with impaired NK cell function in patients with gastric cancer. Gastric Cancer 2012, 15, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riggan, L.; Shah, S.; O’Sullivan, T.E. Arrested development: Suppression of NK cell function in the tumor microenvironment. Clin. Transl. Immunol. 2021, 10, e1238. [Google Scholar] [CrossRef]
- Hunter, M.J.; Tuschong, L.M.; Fowler, C.J.; Bauer, T.R.; Burkholder, T.H.; Hickstein, D.D. Gene Therapy of Canine Leukocyte Adhesion Deficiency Using Lentiviral Vectors With Human CD11b and CD18 Promoters Driving Canine CD18 Expression. Mol. Ther. 2011, 19, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 4th ed.; Alberts, B., Johnson, A., Eds.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Bendas, G.; Borsig, L. Cancer cell adhesion and metastasis: Selectins, integrins, and the inhibitory potential of heparins. Int. J. Cell Biol. 2012, 2012, 676731. [Google Scholar] [CrossRef]
- Ocana, A.; Nieto-Jiménez, C.; Pandiella, A.; Templeton, A.J. Neutrophils in cancer: Prognostic role and therapeutic strategies. Mol. Cancer 2017, 16, 137. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Zhao, Y.; Peng, L.; Chen, N.; Chen, W.; Lv, Y.P.; Mao, F.Y.; Zhang, J.Y.; Cheng, P.; Teng, Y.S.; et al. Tumour-activated neutrophils in gastric cancer foster immune suppression and disease progression through GM-CSF-PD-L1 pathway. Gut 2017, 66, 1900–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackey, J.B.G.; Coffelt, S.B.; Carlin, L.M. Neutrophil Maturity in Cancer. Front. Immunol. 2019, 10, 1912. [Google Scholar] [CrossRef] [Green Version]
- Stålhammar, M.E.; Douhan Håkansson, L.; Sindelar, R. Bacterial N-formyl Peptides Reduce PMA- and Escherichia coli-Induced Neutrophil Respiratory Burst in Term Neonates and Adults. Scand. J. Immunol. 2017, 85, 365–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burzynska-Pedziwiatr, I.; Bukowiecka-Matusiak, M.; Wojcik, M.; Machala, W.; Bienkiewicz, M.; Spolnik, G.; Danikiewicz, W.; Wozniak, L.A. Dual stimulus-dependent effect of Oenothera paradoxa extract on the respiratory burst in human leukocytes: Suppressing for Escherichia coli and phorbol myristate acetate and stimulating for formyl-methionyl-leucyl-phenylalanine. Oxid. Med. Cell Longev. 2014, 2014, 764367. [Google Scholar] [CrossRef] [Green Version]
- Weiskopf, K.; Anderson, K.L.; Ito, D.; Schnorr, P.J.; Hirotaka Tomiyasu, H.; Ring, A.M.; Bloink, K.; Efe, J.; Rue, S.; Lowery, D.; et al. Eradication of canine diffuse large b-cell lymphoma in a murine xenograft model with cd47 blockade and anti-cd20. Cancer Immunol. Res. 2016, 4, 1072–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group I—Bitches with Tumors of Epithelial Origin (No. of Dogs) | Group II—Bitches with Tumors of Mesenchymal Origin (No. of Dogs) |
|---|---|
| complex carcinoma G1 (5) | fibrosarcoma (1) |
| complex carcinoma G2 (4) | osteosarcoma (4) |
| simple carcinoma with fibrosis G1 (5) | |
| tubular carcinoma G2 (2) |
| Case | Breed | Age (Years) | Histological Diagnosis |
|---|---|---|---|
| 1 | German pomeranian | 8 | complex carcinoma Grade 1 |
| 2 | cross breed | 9 | complex carcinoma Grade 1 |
| 3 | spaniel cross | 9 | complex carcinoma Grade 1 |
| 4 | boxer | 8 | complex carcinoma Grade 1 |
| 5 | dachshund | 10 | complex carcinoma Grade 1 |
| 6 | cocker spaniel | 10 | complex carcinoma Grade 2 |
| 7 | cocker spaniel | 8 | complex carcinoma Grade 2 |
| 8 | cross breed | 10 | complex carcinoma Grade 2 |
| 9 | cross breed | 11 | complex carcinoma Grade 2 |
| 10 | Doberman | 11 | tubular carcinoma Grade 2 |
| 11 | cross breed | 9 | tubular carcinoma Grade 2 |
| 12 | spaniel | 10 | tubular carcinoma Grade 2 |
| 13 | German shepherd | 9 | tubular carcinoma Grade 2 |
| 14 | poodle | 12 | tubular carcinoma Grade 2 |
| 15 | dachshund | 12 | simple carcinoma with fibrosis Grade 1 |
| 16 | Maltese | 12 | simple carcinoma with fibrosis Grade 1 |
| 17 | poodle | 13 | fibrosarcoma |
| 18 | cocker spaniel | 11 | osteosarcoma |
| 19 | Rottweiler | 12 | osteosarcoma |
| 20 | Yorkshire terrier | 10 | osteosarcoma |
| 21 | boxer | 13 | osteosarcoma |
| Specificity | Clone | Isotype | Source |
|---|---|---|---|
| Canine CD5 FITC | YKIX322.3 | IgG2a | BIO-RAD |
| Canine CD11b | CA16.3E10 | IgG1 | BIO-RAD |
| goat anti-mouse IgG (Fc):FITC–STAR 120F secondary Ab | polyclonal Ab | polyclonal IgG | BIO-RAD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lisiecka, U.; Brodzki, P.; Śmiech, A.; Kocki, J.; Czop, M.; Adaszek, Ł.; Winiarczyk, S. Comparative Expression Analysis of Innate Immune Markers and Phagocytic Activity in Peripheral Blood of Dogs with Mammary Tumors. Animals 2021, 11, 2398. https://doi.org/10.3390/ani11082398
Lisiecka U, Brodzki P, Śmiech A, Kocki J, Czop M, Adaszek Ł, Winiarczyk S. Comparative Expression Analysis of Innate Immune Markers and Phagocytic Activity in Peripheral Blood of Dogs with Mammary Tumors. Animals. 2021; 11(8):2398. https://doi.org/10.3390/ani11082398
Chicago/Turabian StyleLisiecka, Urszula, Piotr Brodzki, Anna Śmiech, Janusz Kocki, Marcin Czop, Łukasz Adaszek, and Stanisław Winiarczyk. 2021. "Comparative Expression Analysis of Innate Immune Markers and Phagocytic Activity in Peripheral Blood of Dogs with Mammary Tumors" Animals 11, no. 8: 2398. https://doi.org/10.3390/ani11082398
APA StyleLisiecka, U., Brodzki, P., Śmiech, A., Kocki, J., Czop, M., Adaszek, Ł., & Winiarczyk, S. (2021). Comparative Expression Analysis of Innate Immune Markers and Phagocytic Activity in Peripheral Blood of Dogs with Mammary Tumors. Animals, 11(8), 2398. https://doi.org/10.3390/ani11082398