
Immunohistochemical Characterization of Tumor-Associated Macrophages in Canine Lymphomas

 , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Case Selection
2.2. Histopathology
2.3. Immunohistochemistry
2.4. Evaluation of Immunohistochemical Expression
2.5. Statistical Analysis
3. Results
3.1. Determination of HG and Subtype and IPT
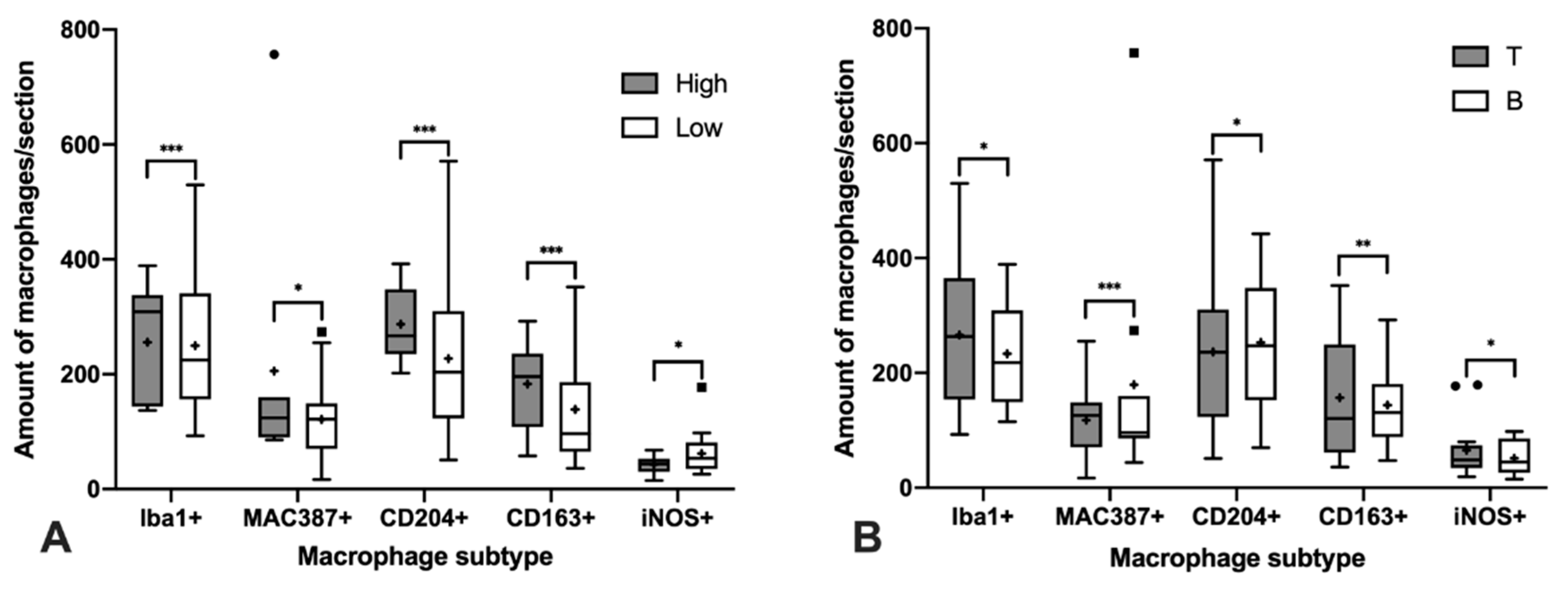
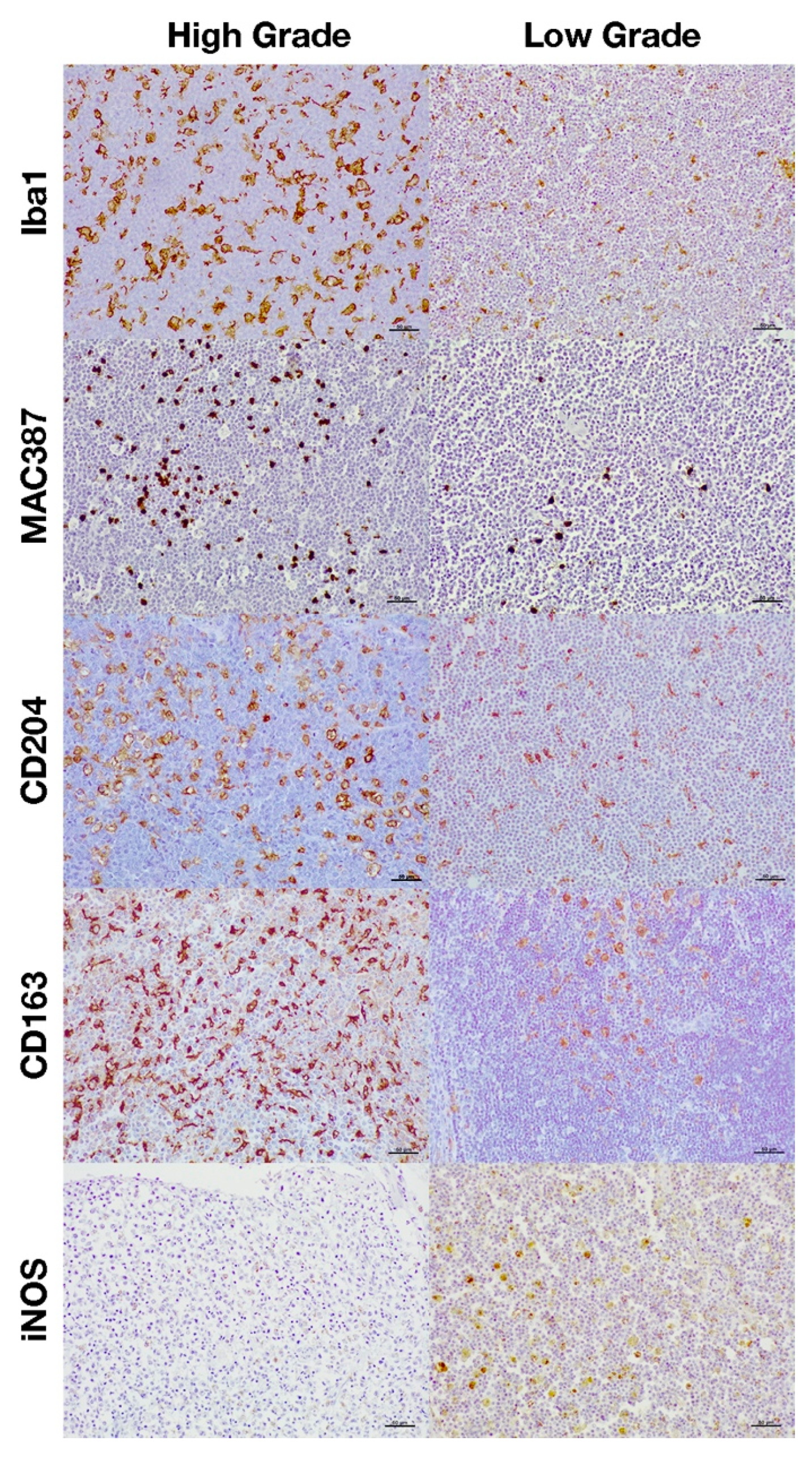
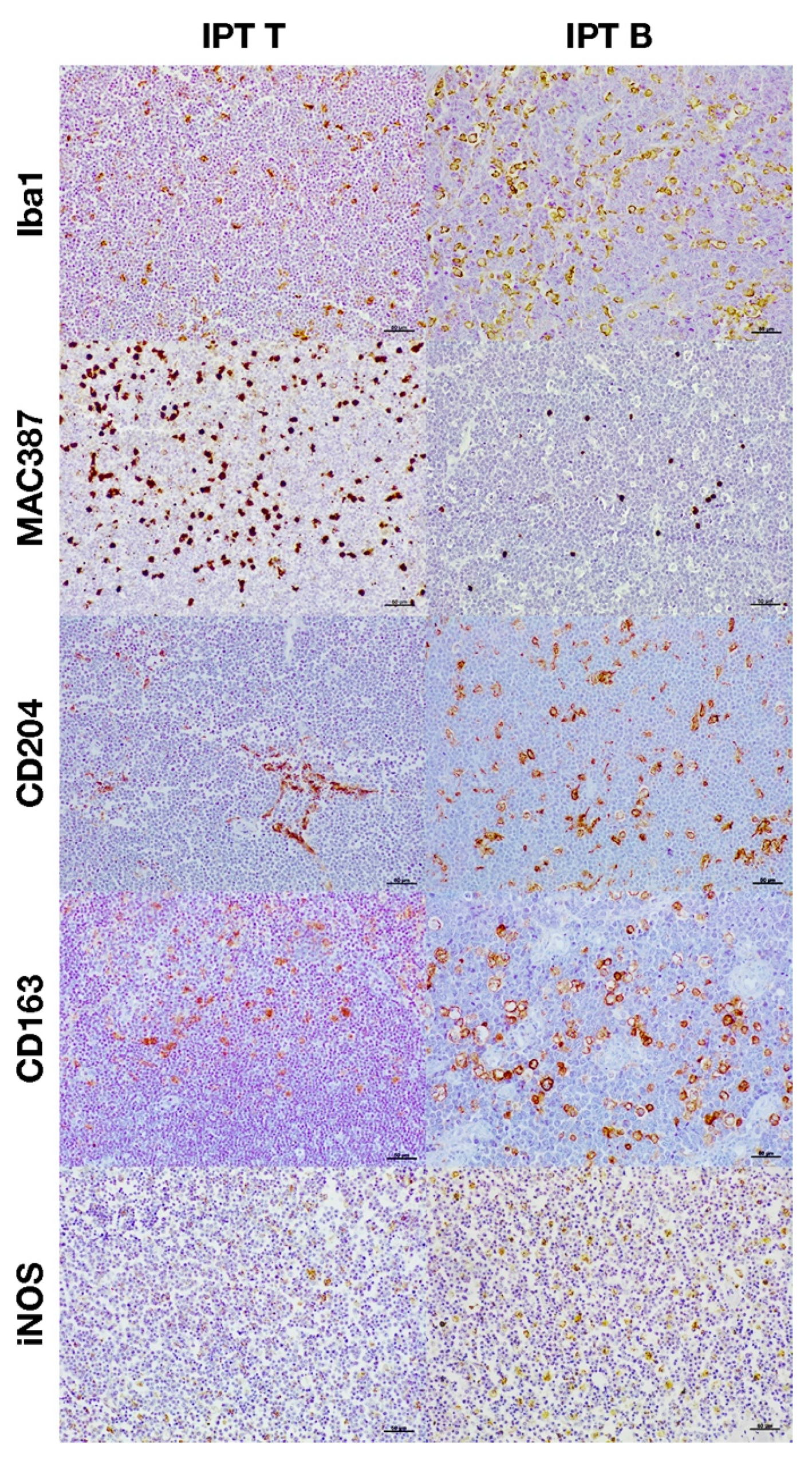
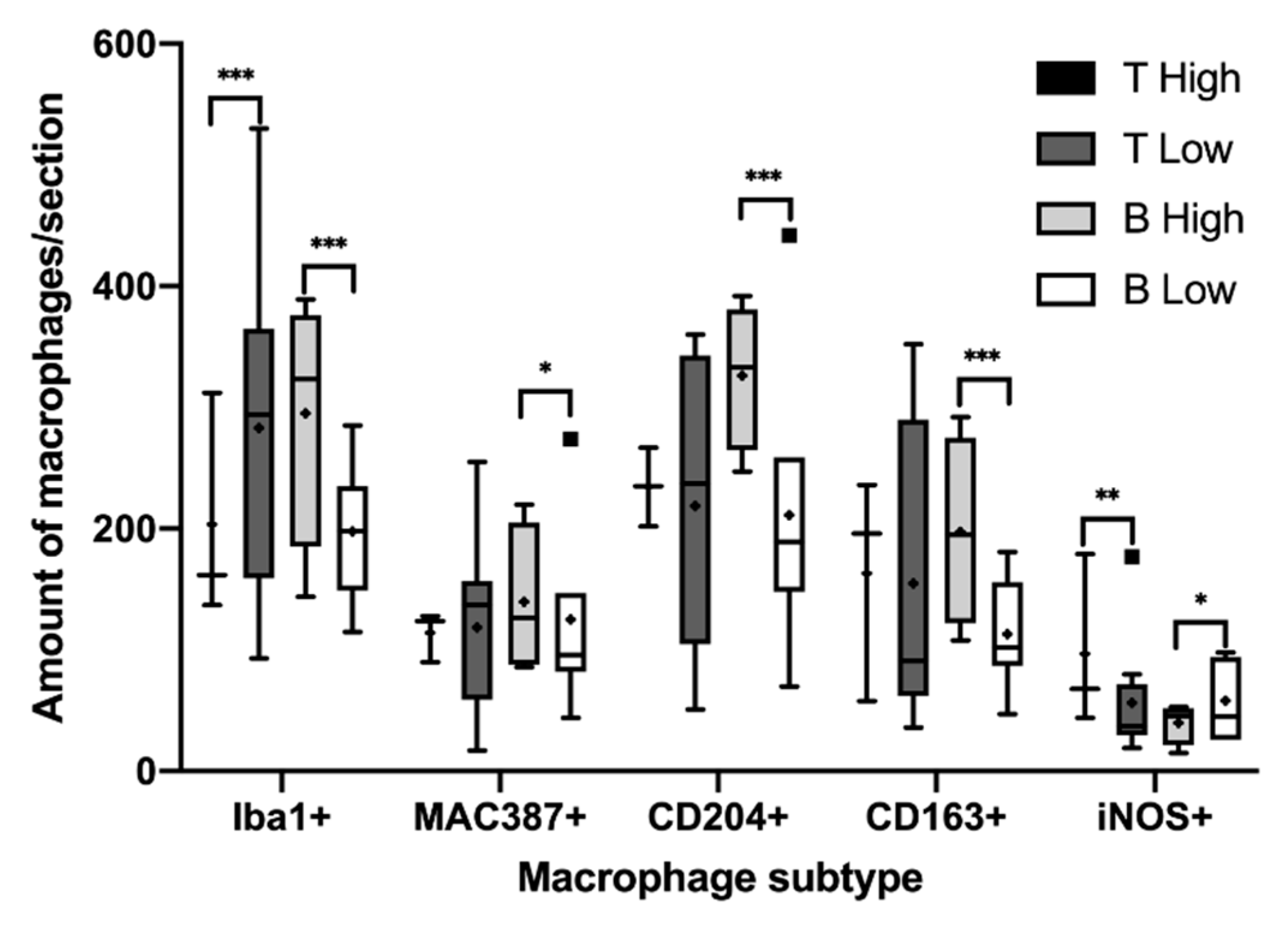
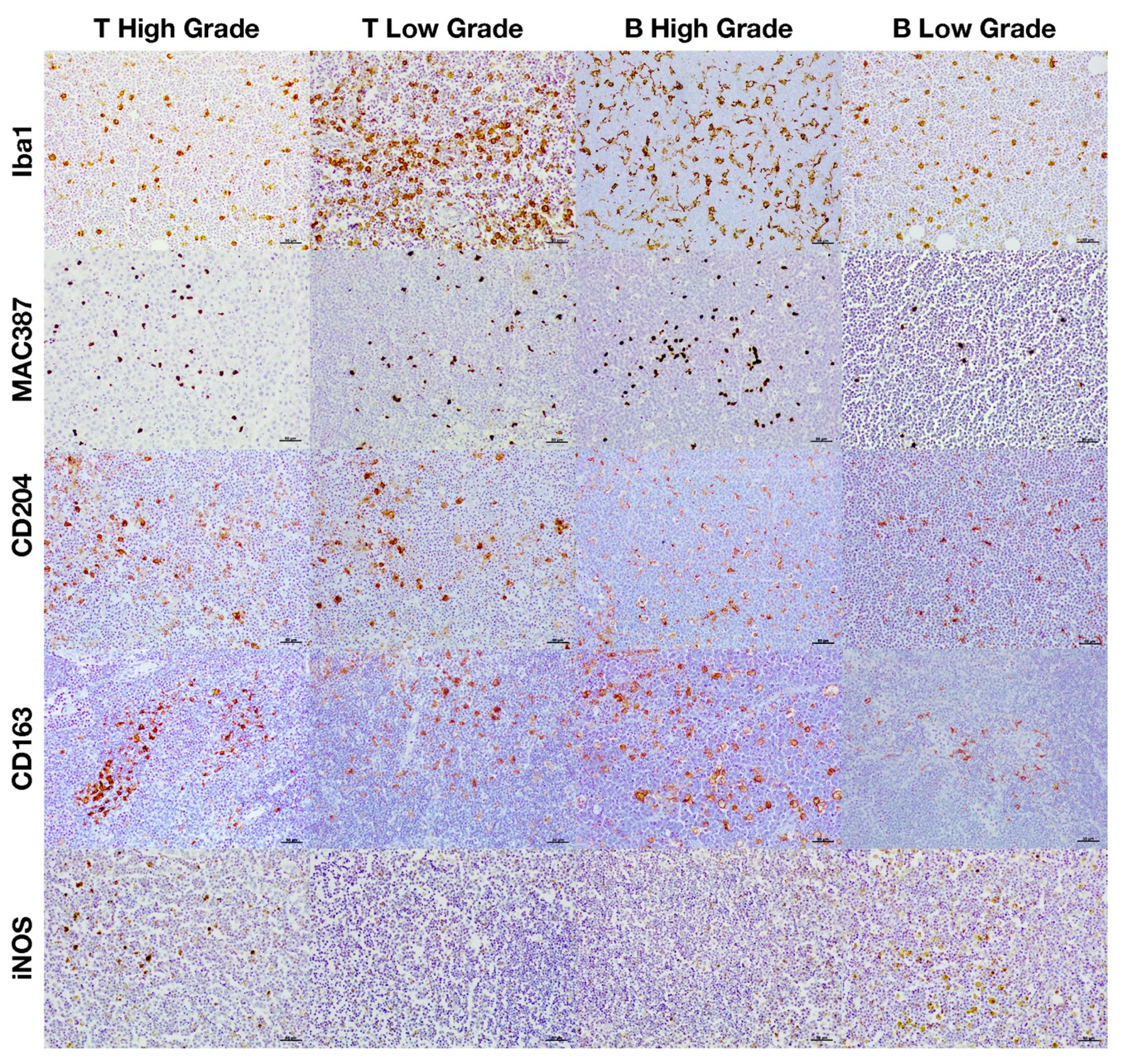
3.2. Immunohistochemical Characterization of Macrophages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Argyle, D.J.; Khann, C. Tumor biology and metastasis. In Withrow & MacEwen’s Small Animal Clinical Oncology; Withrow, S.J., Vail, D.M., Page, R.L., Eds.; Elsevier: St. Louis, MO, USA, 2020; pp. 36–60. [Google Scholar]
- Tse, E.; Kwong, Y.-L. T-Cell Lymphoma: Microenvironment-Related Biomarkers. Semin. Cancer Biol. 2015, 34, 46–51. [Google Scholar] [CrossRef]
- Regan, D.; Guth, A.; Coy, J.; Dow, S. Cancer Immunotherapy in Veterinary Medicine: Current Options and New Developments. Vet. J. 2016, 207, 20–28. [Google Scholar] [CrossRef]
- Guth, A.M.; Dow, S. Cancer immunotherapy. In Withrow & MacEwen’s Small Animal Clinical Oncology; Withrow, S.J., Vail, D.M., Page, R.L., Eds.; Elsevier: St. Louis, MO, USA, 2020; pp. 231–250. [Google Scholar]
- Sica, A.; Mantovani, A. Macrophage Plasticity and Polarization: In Vivo Veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Etzerodt, A.; Moestrup, S.K. CD163 and Inflammation: Biological, Diagnostic, and Therapeutic Aspects. Antioxid. Redox Signal. 2013, 18, 2352–2363. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, K.; Lei, W.; Dong, L.; Xuan, Q.; Feng, M.; Wang, J.; Ye, X.; Jin, T.; Zhang, Z.; et al. Lymphocyte-to-Monocyte Ratio Is Associated with Prognosis of Diffuse Large B-Cell Lymphoma: Correlation with CD163 Positive M2 Type Tumor-Associated Macrophages, Not PD-1 Positive Tumor-Infiltrating Lymphocytes. Oncotarget 2017, 8, 5414–5425. [Google Scholar] [CrossRef]
- Barros, M.H.M.; Hauck, F.; Dreyer, J.H.; Kempkes, B.; Niedobitek, G. Macrophage Polarisation: An Immunohistochemical Approach for Identifying M1 and M2 Macrophages. PLoS ONE 2013, 8, 1–11. [Google Scholar] [CrossRef]
- Soulas, C.; Conerly, C.; Kim, W.K.; Burdo, T.H.; Alvarez, X.; Lackner, A.A.; Williams, K.C. Recently Infiltrating MAC387+ Monocytes/ Macrophages: A Third Macrophage Population Involved in SIV and HIV Encephalitic Lesion Formation. Am. J. Pathol. 2011, 178, 2121–2135. [Google Scholar] [CrossRef] [PubMed]
- Raposo, T.P.; Pires, I.; Carvalho, M.I.; Prada, J.; Argyle, D.J.; Queiroga, F.L. Tumour-Associated Macrophages Are Associated with Vascular Endothelial Growth Factor Expression in Canine Mammary Tumours. Vet. Comp. Oncol. 2015, 13, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Ruffell, B.; Affara, N.I.; Coussens, L.M. Differential Macrophage Programming in the Tumor Microenvironment. Trends Immunol. 2012, 33, 119–126. [Google Scholar] [CrossRef]
- Zhang, M.; He, Y.; Sun, X.; Li, Q.; Wang, W.; Zhao, A.; Di, W. A High M1/M2 Ratio of Tumor-Associated Macrophages Is Associated with Extended Survival in Ovarian Cancer Patients. J. Ovarian Res. 2014, 7, 1–16. [Google Scholar] [CrossRef]
- Shen, L.; Li, H.; Shi, Y.; Wang, D.; Gong, J.; Xun, J.; Zhou, S.; Xiang, R.; Tan, X. M2 Tumour-Associated Macrophages Contribute to Tumour Progression via Legumain Remodelling the Extracellular Matrix in Diffuse Large B Cell Lymphoma. Sci. Rep. 2016, 6, 30347. [Google Scholar] [CrossRef]
- Seung, B.J.; Lim, H.Y.; Shin, J.I.; Kim, H.W.; Cho, S.H.; Kim, S.H.; Sur, J.H. CD204-Expressing Tumor-Associated Macrophages Are Associated with Malignant, High-Grade, and Hormone Receptor–Negative Canine Mammary Gland Tumors. Vet. Pathol. 2018, 55, 417–424. [Google Scholar] [CrossRef]
- Cora, R.; Gal, A.F.; Tabaran, F.; Taulescu, M.; Nagy, A. Epidemiological Data Concerning Canine Lymphoma over a Ten-Year Period (2005–2014), in Cluj-Napoca, Romania. Bull. UASVM Vet. Med. 2016, 73, 83–88. [Google Scholar] [CrossRef][Green Version]
- Vail, D.M.; Pinkerton, M.E.; Young, K.M. Hematopoietic tumors. In Withrow & MacEwen’s Small Animal Clinical Oncology; Withrow, S.J., Vail, D.M., Page, R.L., Eds.; Elsevier: Missouri, MO, USA, 2020; pp. 688–772. [Google Scholar]
- Pedersen, M.B.; Danielsen, A.V.; Hamilton-Dutoit, S.J.; Bendix, K.; Nørgaard, P.; Møller, M.B.; Steiniche, T.; D’Amore, F. High Intratumoral Macrophage Content Is an Adverse Prognostic Feature in Anaplastic Large Cell Lymphoma. Histopathology 2014, 65, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Zafra, R.; Jaber, J.R.; Pérez-Écija, R.A.; Barragán, A.; Martínez-Moreno, A.; Pérez, J. High INOS Expression in Macrophages in Canine Leishmaniasis Is Associated with Low Intracellular Parasite Burden. Vet. Immunol. Immunopathol. 2008, 123, 353–359. [Google Scholar] [CrossRef]
- Murray, P.J.; Wynn, T.A. Obstacles and Opportunities for Understanding Macrophage Polarization. J. Leukoc. Biol. 2011, 89, 557–563. [Google Scholar] [CrossRef]
- Jackute, J.; Zemaitis, M.; Pranys, D.; Sitkauskiene, B.; Miliauskas, S.; Vaitkiene, S.; Sakalauskas, R. Distribution of M1 and M2 Macrophages in Tumor Islets and Stroma in Relation to Prognosis of Non-Small Cell Lung Cancer. BMC Immunol. 2018, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Parisi, F.; Tesi, M.; Millanta, F.; Gnocchi, M.; Poli, A. M1 and M2 Tumour-Associated Macrophages Subsets in Canine Malignant Mammary Tumours: An Immunohistochemical Study. Res. Vet. Sci. 2021, 136, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Finotello, R.; Whybrow, K.; Scarin, G.; Ressel, L. Correlation between Tumour Associated Macrophage (TAM) Infiltration and Mitotic Activity in Canine Soft Tissue Sarcomas. Animals 2021, 11, 684. [Google Scholar] [CrossRef]
- Monteiro, L.N.; Rodrigues, M.A.; Gomes, D.A.; Salgado, B.S.; Cassali, G.D. Tumour-Associated Macrophages: Relation with Progression and Invasiveness, and Assessment of M1/M2 Macrophages in Canine Mammary Tumours. Vet. J. 2018, 234, 119–125. [Google Scholar] [CrossRef]
- Pham, L.V.; Pogue, E.; Ford, R.J. The Role of Macrophage/B-Cell Interactions in the Pathophysiology of B-Cell Lymphomas. Front. Oncol. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cioroianu, A.I.; Stinga, P.I.; Sticlaru, L.; Cioplea, M.D.; Nichita, L.; Popp, C.; Staniceanu, F. Tumor Microenvironment in Diffuse Large B-Cell Lymphoma: Role and Prognosis. Anal. Cell. Pathol. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Menter, T.; Tzankov, A. Lymphomas and Their Microenvironment: A Multifaceted Relationship. Pathobiology 2019, 86, 225–236. [Google Scholar] [CrossRef]
- Rybicka, A.; Eyileten, C.; Taciak, B.; Mucha, J.; Majchrzak, K.; Hellmen, E.; Krol, M. Tumour-Associated Macrophages Influence Canine Mammary Cancer Stem-like Cells Enhancing Their pro-Angiogenic Properties. J. Physiol. Pharmacol. 2016, 67, 491–500. [Google Scholar]
- Woldemeskel, M.; Hawkins, I.; Whittington, L. Ki-67 Protein Expression and Tumor Associated Inflammatory Cells (Macrophages and Mast Cells) in Canine Colorectal Carcinoma. BMC Vet. Res. 2017, 13, 1–6. [Google Scholar] [CrossRef]
- Perry, J.A.; Thamm, D.H.; Eickhoff, J.; Avery, A.C.; Dow, S.W. Increased Monocyte Chemotactic Protein-1 Concentration and Monocyte Count Independently Associate with a Poor Prognosis in Dogs with Lymphoma. Vet. Comp. Oncol. 2011, 9, 55–64. [Google Scholar] [CrossRef]
- Valli, V.E.; Kass, P.H.; Myint, M.S.; Scott, F. Canine Lymphomas: Association of Classification Type, Disease Stage, Tumor Subtype, Mitotic Rate, and Treatment with Survival. Vet. Pathol. 2013, 50, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Valli, V.E.; Myint, M.; Barthel, A.; Bienzle, D.; Caswell, J.; Colbatzky, F.; Durham, A.; Ehrhart, E.J.; Johnson, Y.; Jones, C.; et al. Classification of Canine Malignant Lymphomas According to the World Health Organization Criteria. Vet. Pathol. 2011, 48, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, F.R.; Vieira, P.M.A.; Correa-Oliveira, R.; Giunchetti, R.C.; Carneiro, C.M.; Reis, A.B.; Malaquias, L.C.C. Qualitative and Quantitative Immunohistochemical Evaluation of INOS Expression in the Spleen of Dogs Naturally Infected with Leishmania Chagasi. Parasitol. Res. 2011, 108, 1397–1403. [Google Scholar] [CrossRef]
- Stein, L.; Bacmeister, C.; Kiupel, M. Immunophenotypic Characterization of Canine Nodal T-Zone Lymphoma. Vet. Pathol. 2020, 20, 1–5. [Google Scholar] [CrossRef]
- Thongtharb, A.; Uchida, K.; Chambers, J.K.; Kagawa, Y.; Nakayama, H. Histological and Immunohistochemical Studies on Primary Intracranial Canine Histiocytic Sarcomas. J. Vet. Med. Sci. 2016, 78, 593–599. [Google Scholar] [CrossRef]
- Vanherberghen, M.; Day, M.J.; Delvaux, F.; Gabriel, A.; Clercx, C.; Peeters, D. An Immunohistochemical Study of the Inflammatory Infiltrate Associated with Nasal Carcinoma in Dogs and Cats. J. Comp. Pathol. 2009, 141, 17–26. [Google Scholar] [CrossRef]
- Ahrberg, C.D.; Lee, J.M.; Chung, B.G. Poisson Statistics-Mediated Particle/Cell Counting in Microwell Arrays. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model, Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A Protocol for Data Exploration to Avoid Common Statistical Problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Ullah, M.I.; Aslam, M.; Altaf, S. Lmridge: A Comprehensive R Package for Ridge Regression. R J. 2018, 10, 326–346. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biometrical. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- R Development Core Team R (Versión 3.5.3) [Software]. Available online: https://www.r-project.org (accessed on 14 May 2019).
- Valli, V.E.; Bienzle, D.; Meuten, D.J. Tumors of the hemolymphatic system. In Tumors in Domestic Animals; Meuten, D.J., Ed.; Wiley-Blackwell: Ames, IA, USA, 2017; pp. 203–321. [Google Scholar]
- Voss, J.J.L.P.; Ford, C.A.; Petrova, S.; Melville, L.; Paterson, M.; Pound, J.D.; Holland, P.; Giotti, B.; Freeman, T.C.; Gregory, C.D. Modulation of Macrophage Antitumor Potential by Apoptotic Lymphoma Cells. Cell Death Differ. 2017, 24, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Canioni, D.; Salles, G.; Mounier, N.; Brousse, N.; Keuppens, M.; Morchhauser, F.; Lamy, T.; Sonet, A.; Rousselet, M.C.; Foussard, C.; et al. High Numbers of Tumor-Associated Macrophages Have an Adverse Prognostic Value That Can Be Circumvented by Rituximab in Patients with Follicular Lymphoma Enrolled onto the GELA-GOELAMS FL-2000 Trial. J. Clin. Oncol. 2008, 26, 440–446. [Google Scholar] [CrossRef]
- Rőszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediat. Inflamm. 2015, 2015, 1–16. [Google Scholar] [CrossRef]
- Ross, J.; Auger, M. The biology of the macrophage. In The Macrophage; Burke, B., Lewis, C., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 1–72. [Google Scholar]
- Coupland, S.E. The Challenge of the Microenvironment in B-Cell Lymphomas. Histopathology 2011, 58, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Argyle, D.; Kitamura, T. Targeting Macrophage-Recruiting Chemokines as a Novel Therapeutic Strategy to Prevent the Progression of Solid Tumors. Front. Immunol. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Seimon, T.; Tabas, I. Mechanisms and Consequences of Macrophage Apoptosis in Atherosclerosis. J. Lipid Res. 2009, 50, S382–S387. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Abbas, A.K.; Aster, J.C. Diseases of white blood cells, lymph nodes, spleen and thymus. In Robbins & Cotran: Pathologic Basis of Disease; Kumar, V., Abbas, A.K., Aster, J.C., Turner, J.R., Eds.; Elsevier: Philadelphia, PA, USA, 2021; pp. 583–634. [Google Scholar]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/M-2 Macrophages and the Th1/Th2 Paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef] [PubMed]
- Farinha, P.; Masoudi, H.; Skinnider, B.F.; Shumansky, K.; Spinelli, J.J.; Gill, K.; Klasa, R.; Voss, N.; Connors, J.M.; Gascoyne, R.D. Analysis of Multiple Biomarkers Shows That Lymphoma-Associated Macrophage (LAM) Content Is an Independent Predictor of Survival in Follicular Lymphoma (FL). Blood 2005, 106, 2169–2174. [Google Scholar] [CrossRef]
- Gotti, M.; Nicola, M.; Lucioni, M.; Fiaccadori, V.; Ferretti, V.; Sciarra, R.; Costanza, M.; Bono, E.; Molo, S.; Maffi, A.; et al. Independent Prognostic Impact of Tumour-Infiltrating Macrophages in Early-Stage Hodgkin’s Lymphoma. Hematol. Oncol. 2017, 35, 296–302. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Ab | Origin | Targeted Cell | Dilution | Epitope Retrieval |
|---|---|---|---|---|
| CD20 ThermoFisher Scientific (Waltham, MA, USA) | Rabbit (polyclonal) | B-cells | 1:200 | No |
| CD3 Agilent (Glostrup, Denmark) | Rabbit (polyclonal) | T-cells | 1:300 | Heat-induced pH 6, 90 min |
| Iba1 Wako (Richmond, VA, USA) | Rabbit (polyclonal) | Macrophages | 1:2000 | Heat-induced pH 6, 90 min |
| MAC387 Agilent (Glostrup, Denmark) | Mouse (monoclonal) | Monocytes and macrophages | 1:200 | Heat-induced pH 9, 90 min |
| CD204 TransGenic (Kobe, Japan) | Mouse (monoclonal) | M2 macrophages | 1:400 | Heat-induced pH 6, 90 min |
| CD163 TransGenic (Kobe, Japan) | Mouse (monoclonal) | M2 macrophages | 1:300 | Heat-induced pH 6, 90 min |
| iNOSS anta Cruz Biotechnology (Santa Cruz, CA, USA) | Rabbit (polyclonal) | M1 macrophages | 1:100 | Heat-induced pH 6, 90 min |
| Marker | Linear Hypotheses | Estimate | SE | Z Value | p Value |
|---|---|---|---|---|---|
| Iba1 | IPT (T)—IPT (B) = 0 | −0.370 | 0.049 | −7.433 | <0.05 |
| HG (L)—HG (H) = 0 | −0.399 | 0.039 | −10.082 | <0.001 | |
| IPT (T) *HG (H)—IPT (T) *HG (L) = 0 | −0.330 | 0.044 | −7.449 | <0.001 | |
| IPT (B) *HG (H)—IPT (B) *HG (L) = 0 | 0.399 | 0.040 | 10.082 | <0.001 | |
| MAC387 | IPT (T)—IPT (B) = 0 | 0.877 | 0.619 | 14.160 | <0.001 |
| HG (L)—HG (H) = 0 | −0.782 | 0.045 | −17.270 | <0.05 | |
| IPT (T) *HG (H)—IPT (T) *HG (L) = 0 | −0.041 | 0.061 | −0.669 | 0.504 | |
| IPT (B) *HG (H)—IPT (B) *HG (L) = 0 | 0.783 | 0.045 | 17.272 | <0.05 | |
| CD204 | IPT (T)—IPT (B) = 0 | −0.329 | 0.047 | −7.046 | <0.05 |
| HG (L)—HG (H) = 0 | −0.435 | 0.038 | −11.440 | <0.001 | |
| IPT (T) *HG (H)—IPT (T) *HG (L) = 0 | −0.011 | 0.043 | −0.260 | 0.994 | |
| IPT (B) *HG (H)—IPT (B) *HG (L) = 0 | −0.434 | 0.038 | 11.439 | <0.001 | |
| CD163 | IPT (T)—IPT (B) = 0 | −0.190 | 0.057 | −3.303 | 0.001 |
| HG (L)—HG (H) = 0 | −0.557 | 0.050 | −11.079 | <0.001 | |
| IPT (T) *HG (H)—IPT (T) *HG (L) = 0 | 0.052 | 0.051 | 1.022 | 0.307 | |
| IPT (B) *HG (H)—IPT (B) *HG (L) = 0 | 0.557 | 0.050 | 11.079 | <0.001 | |
| iNOS | IPT (T)—IPT (B) = 0 | −0.892 | 0.099 | −9.046 | <0.05 |
| HG (L)—HG (H) = 0 | 0.383 | 0.093 | 4.094 | <0.05 | |
| IPT (T) *HG (H)—IPT (T) *HG (L) = 0 | 0.540 | 0.071 | 7.599 | <0.01 | |
| IPT (B) *HG (H)—IPT (B) *HG (L) = 0 | −0.383 | 0.093 | −4.094 | <0.05 |
| Marker | Iba1 | MAC387 | CD204 | CD163 | iNOS |
|---|---|---|---|---|---|
| Iba1 | 0.397 * | 0.725 *** | 0.668 *** | −0.205 | |
| MAC387 | 0.252 | 0.362 | 0.118 | ||
| CD204 | 0.572 ** | −0.449 * | |||
| CD163 | −0.250 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez, S.; Vallejo, R.; Espinosa, J.; Arteche, N.; Vega, J.A.; Pérez, V. Immunohistochemical Characterization of Tumor-Associated Macrophages in Canine Lymphomas. Animals 2021, 11, 2301. https://doi.org/10.3390/ani11082301
Vázquez S, Vallejo R, Espinosa J, Arteche N, Vega JA, Pérez V. Immunohistochemical Characterization of Tumor-Associated Macrophages in Canine Lymphomas. Animals. 2021; 11(8):2301. https://doi.org/10.3390/ani11082301
Chicago/Turabian StyleVázquez, Sergio, Raquel Vallejo, José Espinosa, Noive Arteche, José A. Vega, and Valentín Pérez. 2021. "Immunohistochemical Characterization of Tumor-Associated Macrophages in Canine Lymphomas" Animals 11, no. 8: 2301. https://doi.org/10.3390/ani11082301
APA StyleVázquez, S., Vallejo, R., Espinosa, J., Arteche, N., Vega, J. A., & Pérez, V. (2021). Immunohistochemical Characterization of Tumor-Associated Macrophages in Canine Lymphomas. Animals, 11(8), 2301. https://doi.org/10.3390/ani11082301