Can Moderate Levels of Organic Selenium in Dairy Cow Feed Naturally Enrich Dairy Products?


 , ,
, ,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Yogurt Elaboration
2.3. Cheese Elaboration
2.4. Laboratory Procedures
2.4.1. Feed Analysis
2.4.2. Mineral Analysis (Ca, P, Zn, Cu, and Se)
2.4.3. GSH-Px, Hb and Metabolic Profile
2.4.4. Proximate Composition and pH of Milk and Dairy Products
2.4.5. Physical Parameters of Dairy Products
2.4.6. Sensorial Parameters of Dairy Products
2.5. Statistical Analyses
3. Results
3.1. Whole Blood and Plasma Analyses
3.2. Yield, Composition and pH of Milk
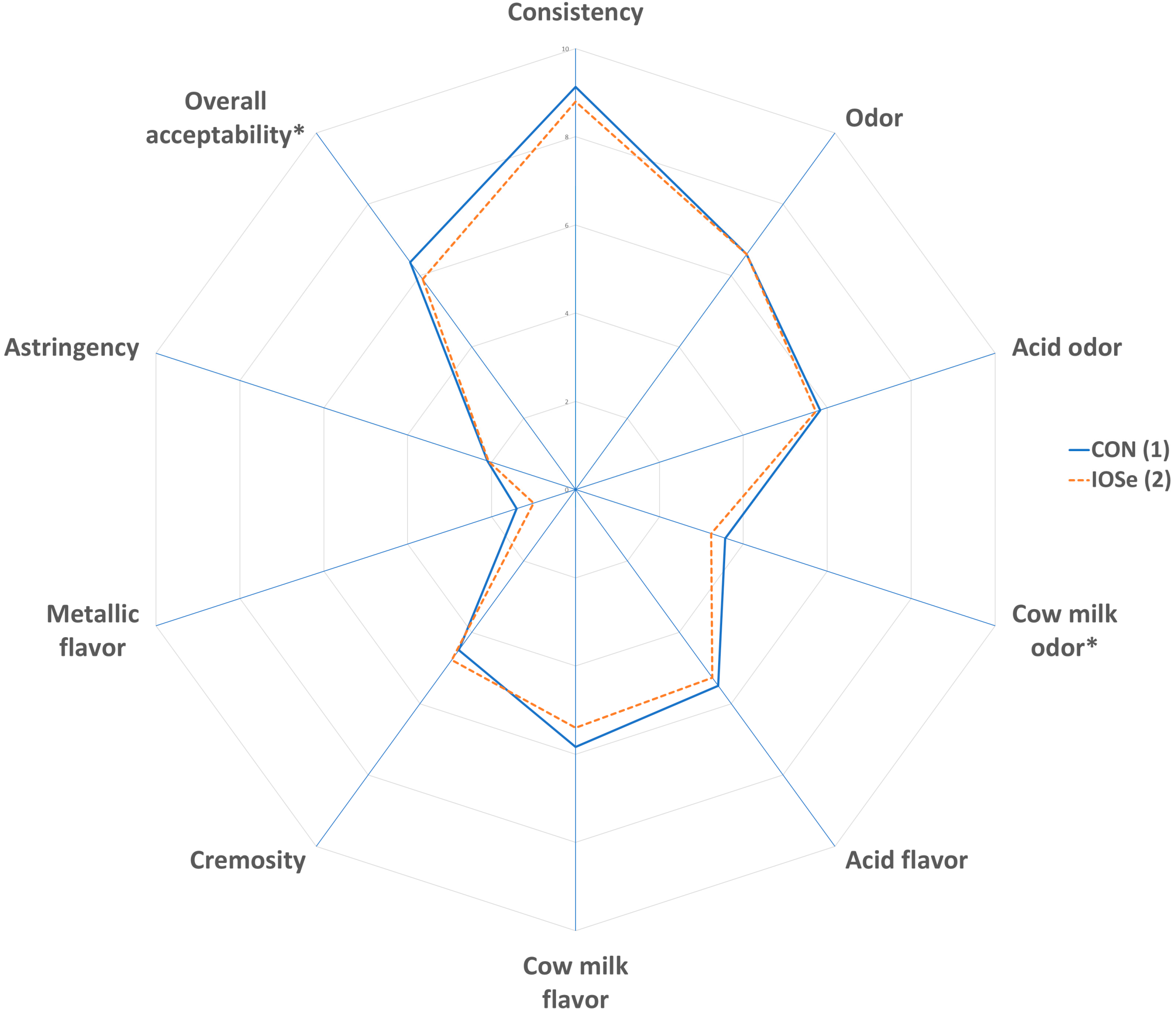
3.3. Physicochemical Composition and Sensory Parameters of Yogurt
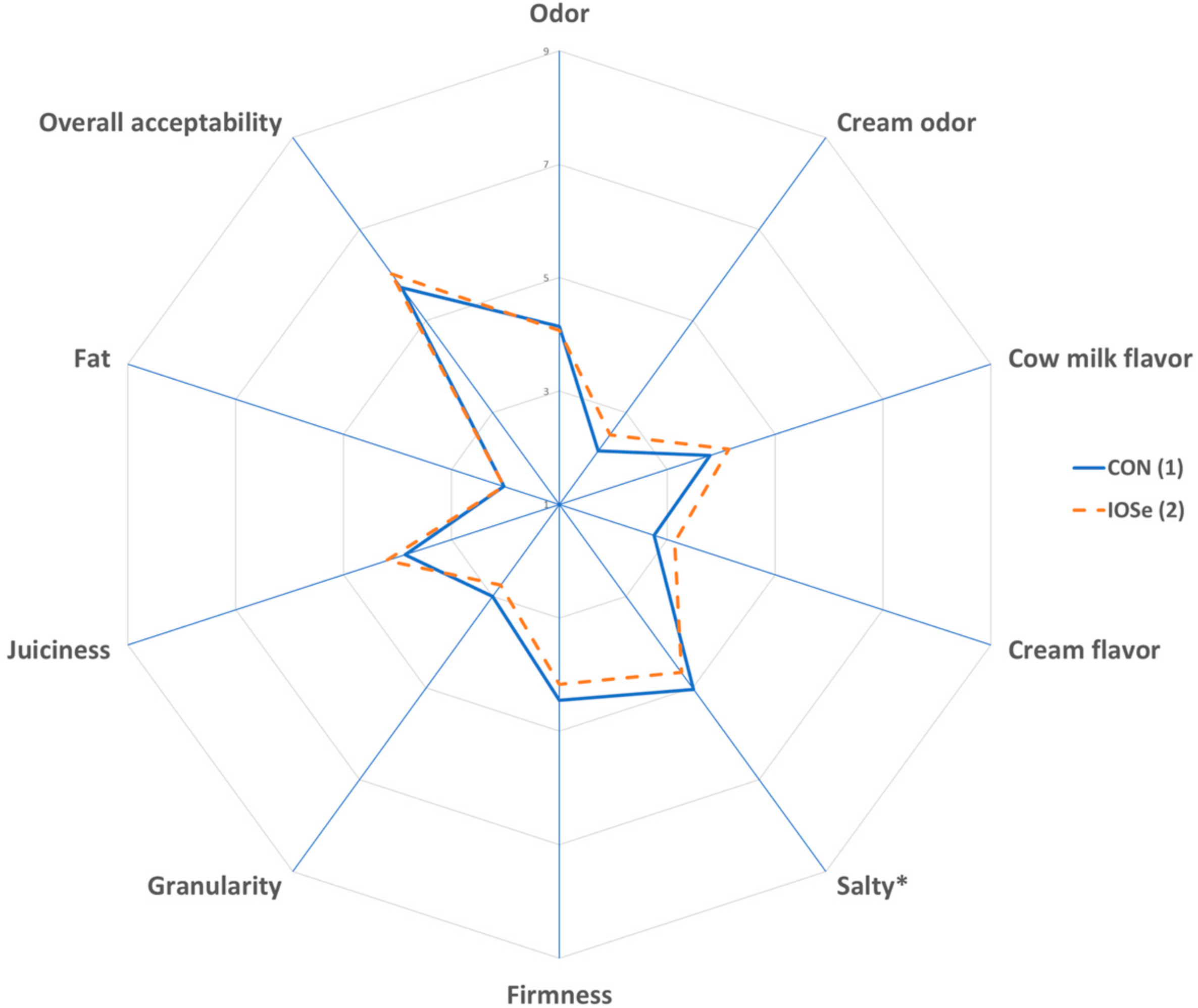
3.4. Physicochemical Composition and Sensory Parameters of Fresh Cheese
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kieliszek, M. Selenium—Fascinating microelement, properties and sources in food. Molecules 2019, 24, 1298. [Google Scholar] [CrossRef]
- Branco, V.; Canário, J.; Lu, J.; Holmgren, A.; Carvalho, C. Mercury and selenium interaction in vivo: Effects on thioredoxin reductase and glutathione peroxidase. Free Radic. Biol. Med. 2012, 52, 781–793. [Google Scholar] [CrossRef]
- Benstoem, C.; Goetzenich, A.; Kraemer, S.; Borosch, S.; Manzanares, W.; Hardy, G.; Stoppe, C. Selenium and its supplementation in cardiovascular disease—What do we know? Nutrients 2015, 7, 3094–3118. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Zhu, W.; Wang, W.; Li, R.; Hou, S.; Wang, D.; Yang, L. Selenium in soil and endemic diseases in China. Sci. Total Environ. 2002, 284, 227–235. [Google Scholar] [CrossRef]
- Fairweather-Tait, S.J.; Bao, Y.; Broadley, M.R.; Collings, R.; Ford, D.; Hesketh, J.E.; Hurst, R. Selenium in human health and disease. Antioxid. Redox Signal. 2011, 14, 1337–1383. [Google Scholar] [CrossRef] [PubMed]
- Thomson, C.D. Assessment of requirements for selenium and adequacy of selenium status: A review. Eur. J. Clin. Nutr. 2004, 58, 391–402. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Trace Elements in Human Nutrition and Health; World Health Organization: Geneva, Switzerland, 1996. [Google Scholar]
- Bierta, K.; Vacchina, V.; Szpunar, J.; Bertin, G.; Lobiński, R. Simultaneous derivatization of selenocysteine and selenomethionine in animal blood prior to their specific determination by 2D size-exclusion ion-pairing reversed-phase HPLC-ICP MS. J. Anal. At. Spectrom. 2008, 23, 508–513. [Google Scholar] [CrossRef]
- Stoffaneller, R.; Morse, N.L. A review of dietary selenium intake and selenium status in Europe and the Middle East. Nutrients 2015, 7, 1494–1537. [Google Scholar] [CrossRef]
- Knowles, S.O.; Grace, N.D.; Knight, T.W.; McNabb, W.C.; Lee, J. Reasons and means for manipulating the micronutrient composition of milk from grazing dairy cattle. Anim. Feed Sci. Technol. 2006, 131, 154–167. [Google Scholar] [CrossRef]
- Górska-Warsewicz, H.; Rejman, K.; Laskowski, W.; Czeczotko, M. Milk and dairy products and their nutritional contribution to the average polish diet. Nutrients 2019, 11, 1771. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Juniper, D.T. Revisiting oxidative stress and the use of organic selenium in dairy cow nutrition. Animals 2019, 9, 462. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, Y.; Hornick, J.-L.; Istasse, L.; Dufrasne, I. Selenium in the environment, metabolism and involvement in body functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef] [PubMed]
- Conrad, H.R.; Moxon, A.L. Transfer of dietary selenium to milk. J. Dairy Sci. 1979, 62, 404–411. [Google Scholar] [CrossRef]
- Doucha, J.; Lívanskỳ, K.; Kotrbáček, V.; Zachleder, V. Production of Chlorella biomass enriched by selenium and its use in animal nutrition: A review. Appl. Microbiol. Biotechnol. 2009, 83, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, A.; Sánchez, J.; Stryhn, H.; Montgomery, J.B.; Barkema, H.W.; Wichtel, J.J. Meta-analysis of the effect of oral selenium supplementation on milk selenium concentration in cattle. J. Dairy Sci. 2009, 92, 324–342. [Google Scholar] [CrossRef]
- Walker, G.P.; Dunshea, F.R.; Heard, J.W.; Stockdale, C.R.; Doyle, P.T. Output of selenium in milk, urine, and feces is proportional to selenium intake in dairy cows fed a total mixed ration supplemented with selenium yeast. J. Dairy Sci. 2010, 93, 4644–4650. [Google Scholar] [CrossRef]
- Heard, J.W.; Stockdale, C.R.; Walker, G.P.; Leddin, C.M.; Dunshea, F.R.; McIntosh, G.H.; Shields, P.M.; McKenna, A.; Young, G.P.; Doyle, P.T. Increasing selenium concentration in milk: Effects of amount of selenium from yeast and cereal grain supplements. J. Dairy Sci. 2007, 90, 4117–4127. [Google Scholar] [CrossRef]
- Petrera, F.; Calamari, L.; Bertin, G. Effect of either sodium selenite or Se–yeast supplementation on selenium status and milk characteristics in dairy goats. Small Rumin. Res. 2009, 82, 130–138. [Google Scholar] [CrossRef]
- Calamari, L.; Petrera, F.; Bertin, G. Effects of either sodium selenite or Se yeast (Sc CNCM I-3060) supplementation on selenium status and milk characteristics in dairy cows. Livest. Sci. 2010, 128, 154–165. [Google Scholar] [CrossRef]
- Gong, J.; Ni, L.; Wang, D.; Shi, B.; Yan, S. Effect of dietary organic selenium on milk selenium concentration and antioxidant and immune status in midlactation dairy cows. Livest. Sci. 2014, 170, 84–90. [Google Scholar] [CrossRef]
- Pechova, A.; Janštová, B.; Mišurová, L.; Dračková, M.; Vorlova, L.; Pavlata, L. Impact of supplementation of various selenium forms in goats on quality and composition of milk, cheese and yoghurt. Acta Vet. Brno 2008, 77, 407–414. [Google Scholar] [CrossRef]
- Moschini, M.; Battaglia, M.; Beone, G.M.; Piva, G.; Masoero, F. Iodine and selenium carry over in milk and cheese in dairy cows: Effect of diet supplementation and milk yield. Animal 2010, 4, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Ling, K.; Henno, M.; Jõudu, I.; Püssa, T.; Jaakson, H.; Kass, M.; Anton, D.; Ots, M. Selenium supplementation of diets of dairy cows to produce Se-enriched cheese. Int. Dairy J. 2017, 71, 76–81. [Google Scholar] [CrossRef]
- EU Commission Implementing Regulation (EU) 2019/804 of 17 May 2019 Concerning the Renewal of the Authorisation of Organic form of Selenium Produced by Saccharomyces Cerevisiae CNCM I-3060 and of Selenomethionine Produced by Saccharomyces Cerevisiae NCYC R397 as Feed Additives for All Animal Species and Repealing Regulations (EC) No 1750/2006 and (EC) No 634/2007 (Text with EEA Relevance). Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=uriserv%3AOJ.L_.2019.132.01.0028.01.ENG&toc=OJ%3AL%3A2019%3A132%3ATOC (accessed on 14 July 2020).
- Ianni, A.; Martino, C.; Pomilio, F.; Di Luca, A.; Martino, G. Dietary selenium intake in lactating dairy cows modifies fatty acid composition and volatile profile of milk and 30-day-ripened caciotta cheese. Eur. Food Res. Technol. 2019, 245, 2113–2121. [Google Scholar] [CrossRef]
- Ianni, A.; Bennato, F.; Martino, C.; Innosa, D.; Grotta, L.; Martino, G. Effects of selenium supplementation on chemical composition and aromatic profiles of cow milk and its derived cheese. J. Dairy Sci. 2019, 102, 6853–6862. [Google Scholar] [CrossRef]
- Foster, L.H.; Chaplin, M.F.; Sumar, S. The effect of heat treatment on intrinsic and fortified selenium levels in cow’s milk. Food Chem. 1998, 62, 21–25. [Google Scholar] [CrossRef]
- Alzate, A.; Pérez-Conde, M.C.; Gutiérrez, A.M.; Cámara, C. Selenium-enriched fermented milk: A suitable dairy product to improve selenium intake in humans. Int. Dairy J. 2010, 20, 761–769. [Google Scholar] [CrossRef]
- EU Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protectionof Animals Used for Scientific PurposesText with EEA Relevance. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32010L0063 (accessed on 8 June 2020).
- Fundación Española para el Desarrollo de la Nutrición Animal (FEDNA). Necesidades Nutricionales para Rumiantes de Leche. Normas FEDNA; Fundación Española para el Desarrollo de la Nutrición Animal: Madrid, Spain, 2009. [Google Scholar]
- Fundación Española para el Desarrollo de la Nutrición Animal (FEDNA). Tablas FEDNA de Composición y valor Nutritivo de Alimentos para la Fabricación de Piensos Compuestos; Fundación Española para el Desarrollo de la Nutrición Animal: Madrid, Spain, 2010. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2006. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Weichselbaum, C.T. An accurate and rapid method for the determination of proteins in small amounts of blood serum and plasma. Am. J. Clin. Path. 1946, 16, 40–49. [Google Scholar] [CrossRef]
- Talke, H.; Schubert, G. Enzymatic urea determination in the blood and serum in the Warburg optical test. Klin. Wochenschr. 1965, 43, 174–175. [Google Scholar] [CrossRef]
- Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- IDF standard 141B Whole Milk: Determination of Milk Fat, Protein and Lactose Content. In Guide for the Operation of Mid-Infrared Instruments; International Dairy Federation: Brussels, Belgium, 1996.
- IDF Standard 4 Cheese and Processed Cheese. In Determination of Total Solids Content (Reference Method); International Dairy Federation: Brussels, Belgium, 2004.
- IDF Standard 5 Cheese and processed cheese products. In Determination of Fat Content. Gravimetric Method (Reference Method); International Dairy Federation: Brussels, Belgium, 2004.
- IDF Standard 25 Processed cheese products. In Determination of Nitrogen Content and Crude Protein Calculation. Kjeldahl Method; International Dairy Federation: Brussels, Belgium, 2008.
- Bourne, M.C. Texture profile analysis. Food Technol. 1978, 32, 62–66. [Google Scholar]
- Kincaid, R. Changes in the concentration of minerals in blood of peripartum cows. In Proceedings of the Mid-South Ruminant Nutrition Conference, Arlington, TX, USA, 7–8 April 2008; pp. 1–8. [Google Scholar]
- Stowe, H.D.; Herdt, T.H. Clinical assessment of selenium status of livestock. J. Animal Sci. 1992, 70, 3928–3933. [Google Scholar] [CrossRef]
- Smith, K.L.; Hogan, J.S.; Conrad, H.R. Selenium in dairy cattle: Its role in disease resistance. Vet. Med. 1988, 83, 72–78. [Google Scholar]
- Juniper, D.T.; Phipps, R.H.; Jones, A.K.; Bertin, G. Selenium supplementation of lactating dairy cows: Effect on selenium concentration in blood, milk, urine, and feces. J. Dairy Sci. 2006, 89, 3544–3551. [Google Scholar] [CrossRef]
- Weiss, W.P. Selenium nutrition of dairy cows: Comparing responses to organic and inorganic selenium forms. 33-343 in Nutritional Biotechnology in the Feed and Food Industries. In Proceedings of the Alltech’s 19th Annual Symposium, Lexington, KY, USA, 11–14 May 2003; Nottingham University Press: Nottingham, UK, 2003; pp. 333–343. [Google Scholar]
- Knowles, S.O.; Grace, N.D.; Wurms, K.; Lee, J. Significance of amount and form of dietary selenium on blood, milk, and casein selenium concentrations in grazing cows. J. Dairy Sci. 1999, 82, 429–437. [Google Scholar] [CrossRef]
- Thompson, K.G.; Fraser, A.J.; Harrop, B.M.; Kirk, J.A. Glutathione peroxidase activity in bovine serum and erythrocytes in relation to selenium concentrations of blood, serum and liver. Res. Vet. Sci. 1980, 28, 321–324. [Google Scholar] [CrossRef]
- Piccione, G.; Messina, V.; Marafioti, S.; Casella, S.; Giannetto, C.; Fazio, F. Changes of some haematochemical parameters in dairy cows during late gestation, post partum, lactation and dry periods. Vet. Med. Zoot. 2012, 58, 59–64. [Google Scholar]
- Sevinç, M.; Başoğlu, A.; Öztok, İ.; Sandikçi, M.; Birdane, F. The clinical-chemical parameters, serum lipoproteins and fatty infiltration of the liver in ketotic cows. Turk. J. Vet. Anim. Sci. 1998, 22, 443–447. [Google Scholar]
- Rayssiguier, Y.; Mazur, A.; Gueux, E.; Reid, I.M.; Roberts, C.J. Plasma lipoproteins and fatty liver in dairy cows. Res. Vet. Sci. 1988, 45, 389–393. [Google Scholar] [CrossRef]
- McArt, J.A.A.; Nydam, D.V.; Oetzel, G.R. Epidemiology of subclinical ketosis in early lactation dairy cattle. J. Dairy Sci. 2012, 95, 5056–5066. [Google Scholar] [CrossRef] [PubMed]
- Gunter, S.A.; Beck, P.A.; Phillips, J.M. Effects of supplementary selenium source on the performance and blood measurements in beef cows and their calves. J. Anim. Sci. 2003, 81, 856–864. [Google Scholar] [CrossRef]
- Stockdale, C.R.; Gill, H.S. Effect of duration and level of supplementation of diets of lactating dairy cows with selenized yeast on selenium concentrations in milk and blood after the withdrawal of supplementation. J. Dairy Sci. 2011, 94, 2351–2359. [Google Scholar] [CrossRef]
- Ortman, K.; Pehrson, B. Effect of selenate as a feed supplement to dairy cows in comparison to selenite and selenium yeast. J. Anim. Sci. 1999, 77, 3365–3370. [Google Scholar] [CrossRef]
- Pehrson, B. Selenium in nutrition with special reference to the biopotency of organic and inorganic selenium compounds. In Proceedings of the Alltech’s 9th Annual Symposium, Lexington, KY, USA, 5–9 September 1993; Nottingham University Press: Nottingham, UK, 1993; pp. 71–89. [Google Scholar]
- McIntosh, G.H.; Royle, P.J. Supplementation of Cows with Organic Selenium and the Identification of Selenium-Rich Protein Fractions in Milk. In Proceedings of the Alltech’s 18th Annual Symposium, Lexington, KY, USA, 13–15 May 2002; Nottingham University Press: Nottingham, UK, 2002; pp. 233–238. [Google Scholar]
- Heard, J.W.; Walker, G.P.; Royle, P.J.; McIntosh, G.H.; Doyle, P.T. Effects of short-term supplementation with selenised yeast on milk production and composition of lactating cows. Aust. J. Dairy Technol. 2004, 59, 199–203. [Google Scholar]
- EU Regulation (EU) No 1169/2011 of the European Parliament and of the Council of 25 October 2011 on the provision of Food Information to Consumers, Amending Regulations (EC) No 1924/2006 and (EC) No 1925/2006 of the European Parliament and of the Council, and repealing Commission Directive 87/250/EEC, Council Directive 90/496/EEC, Commission Directive 1999/10/EC, Directive 2000/13/EC of the European Parliament and of the Council, Commission Directives 2002/67/EC and 2008/5/EC and Commission Regulation (EC) No 608/2004 Text with EEA Relevance. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32011R1169 (accessed on 10 July 2020).
- Čejna, V.; Chládek, G. The importance of monitoring changes in milk fat to milk protein ratio in Holstein cows during lactation. J. Cent. Eur. Agric. 2005, 6, 539–546. [Google Scholar]
- Csapó, J.; Holló, G.; Holló, I.; Salamon, R.V.; Salamon, S.; Toró, S.; Csapóné Kiss, Z. Production of selenium-enriched milk and dairy products. Acta Univ. Sapientiae Aliment. 2015, 8, 5–29. [Google Scholar] [CrossRef]
- Yoshida, M.; Yasumoto, K.; Iwami, K.; Tashiro, H. Distribution of selenium in bovine milk and selenium deficiency in rats fed casein-based diets, monitored by lipid peroxide level and glutathione peroxidase activity. Agric. Biol. Chem. 1981, 45, 1681–1688. [Google Scholar] [CrossRef]
- Mathias, M.M.; Hogue, D.E.; Loosli, J.K. The biological value of selenium in bovine milk for the rat and chick. J. Nutr. 1967, 93, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Van Dael, P.; Vlaemynck, G.; Van Renterghem, R.; Deelstra, H. Selenium content of cow’s milk and its distribution in protein fractions. Z. Lebensm. Unters. Forch. 1991, 192, 422–426. [Google Scholar] [CrossRef]
- Netto, A.S.; Zanetti, M.A.; Del Claro, G.R.; Vilela, F.G.; de Melo, M.P.; Correa, L.B.; Pugine, S.M.P. Copper and selenium supplementation in the diet of Brangus steers on the nutritional characteristics of meat. R. Bras. Zootec. 2013, 42, 70–75. [Google Scholar] [CrossRef]
- Netto, A.S.; Zanetti, M.A.; Del Claro, G.R.; de Melo, M.P.; Vilela, F.G.; Correa, L.B. Effects of copper and selenium supplementation on performance and lipid metabolism in confined brangus bulls. Asian-Australas. J. Anim. Sci. 2014, 27, 488. [Google Scholar] [CrossRef]
- Maas, S.; Lucot, E.; Gimbert, F.; Crini, N.; Badot, P.-M. Trace metals in raw cows’ milk and assessment of transfer to Comté cheese. Food Chem. 2011, 129, 7–12. [Google Scholar] [CrossRef]
- Outinen, M.; Rantamaki, P. Whey protein isolate (WPI) gel hardness and structure influenced by the concentration of alpha-lactalbumin and kappa-casein macropeptide. Milchwissenschaft 2008, 63, 77–80. [Google Scholar]



{kind=link}
{kind=link}
| Item | Content |
|---|---|
| Ingredients, g/kg DM 1 | |
| Forage | |
| Alfalfa hay | 475.0 |
| Barley straw | 47.5 |
| Concentrate | |
| Corn grain | 177.0 |
| Barley grain | 99.5 |
| Dried beet pulp | 94.5 |
| Corn gluten feed | 36.7 |
| Soybean meal (47% Crude Protein) | 35.5 |
| Palm oil | 17.7 |
| Sodium bicarbonate | 7.8 |
| Monocalcium phosphate | 3.5 |
| Sodium chloride | 2.9 |
| Vitamin-mineral premix 2 | 2.4 |
| Calculated composition 3 | |
| Crude Protein (g/kg DM) | 156.0 |
| Starch (g/kg DM) | 232.7 |
| Neutral Detergent Fiber (g/kg DM) | 381.7 |
| Net Energy for Lactation (MJ/kg DM) | 6.52 |
| Ca (g/kg DM) | 11.8 |
| P (g/kg DM) | 3.9 |
| Se (mg/kg DM) | 0.240 |
| Item | Alfalfa Hay | Barley Straw | CON Concentrate 1 | IOSe Concentrate 2 | CON Ration 3 | IOSe Ration 4 |
|---|---|---|---|---|---|---|
| Analysed composition (g/kg DM 5) | ||||||
| Ash | 111.7 | 78.5 | 55.3 | 50.3 | 83.2 | 80.8 |
| Ether Extract | 22.1 | 17.4 | 22.8 | 23.0 | 22.2 | 22.3 |
| Crude Protein | 175 | 40.3 | 118 | 124 | 142 | 144 |
| Neutral Detergent Fiber | 546 | 785 | 196 | 199 | 389 | 390 |
| Acid Detergent Fiber | 394 | 505 | 49.8 | 50.2 | 235 | 235 |
| Lignin Acid Detergent | 91.2 | 91.6 | 40.5 | 41.7 | 67.0 | 67.6 |
| P | 2.3 | 0.2 | 5.3 | 5.5 | 3.6 | 3.7 |
| Ca | 14.5 | 3.2 | 7.4 | 5.7 | 10.5 | 9.7 |
| Se (mg/kg) | 0.066 | 0.042 | 0.382 | 0.423 | 0.205 | 0.225 |
| Item | CON Ration 1 | IOSe Ration 2 | SEM 3 | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| T0 4 | T21 | T49 | T0 | T21 | T49 | Ration (R) | Day (D) | R × D | ||
| Se (µg/L) | 135.7 | 121.4 | 129.7 | 128.3 | 129.9 | 158.6 | 9.39 | 0.222 | 0.605 | 0.081 |
| P (mg/100 g) | 17.4 | 18.4 | 17.6 | 18.3 | 20.3 | 19.2 | 0.39 | 0.236 | 0.086 | 0.701 |
| Ca (mg/100 g) | 7.57 | 6.61 | 6.80 | 7.35 | 7.21 | 7.02 | 0.257 | 0.275 | 0.723 | 0.653 |
| Cu (mg/100 g) | 0.075 | 0.091 | 0.074 | 0.069 | 0.067 | 0.067 | 0.003 | 0.789 | 0.079 | 0.875 |
| Zn (mg/100 g) | 0.212 | 0.206 | 0.180 | 0.248 | 0.249 | 0.208 | 0.009 | 0.002 | 0.079 | 0.682 |
| Hb (g/dL) | 10.5 | 10.5 | 10.1 | 11.2 | 10.7 | 10.6 | 0.14 | 0.129 | 0.058 | 0.566 |
| GSH-Px (U/L) | 20,611 | 24,812 | 26,448 | 21,937 | 23,821 | 26,815 | 713.9 | 0.872 | <0.001 | 0.377 |
| GSH-Px (U/g Hb) | 196.7 | 236.3 | 259.3 | 195.7 | 221.1 | 252.7 | 5.96 | 0.534 | <0.001 | 0.547 |
| Item | CON Ration 1 | IOSe Ration 2 | SEM 3 | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| T0 4 | T21 | T49 | T0 | T21 | T49 | Ration (R) | Day (D) | R × D | ||
| Cholesterol (mg/dL) | 223.6 | 264.5 | 265.4 | 230.2 | 259.7 | 282.8 | 12.71 | 0.805 | <0.001 | 0.512 |
| TG (mg/dL) 5 | 10.6 | 9.5 | 38.0 | 11.8 | 10.9 | 17.0 | 3.96 | 0.449 | 0.118 | 0.365 |
| NEFA (mmol/L) 6 | 0.051 | 0.029 | 0.027 | 0.064 | 0.029 | 0.036 | 0.033 | 0.097 | 0.218 | 0.150 |
| β-BHA (mmol/L) 7 | 0.400 | 0.331 | 0.261 | 0.384 | 0.448 | 0.375 | 0.015 | 0.036 | 0.004 | 0.008 |
| Glucose (mg/dL) | 64.6 | 66.0 | 66.2 | 64.6 | 66.4 | 63.6 | 0.77 | 0.650 | 0.829 | 0.386 |
| Urea (mg/dL) | 28.2 | 24.4 | 27.8 | 26.9 | 27.6 | 29.7 | 0.76 | 0.411 | 0.254 | 0.130 |
| Total proteins (g/dL) | 7.29 | 7.42 | 7.38 | 7.22 | 7.00 | 7.38 | 0.105 | 0.458 | 0.151 | 0.686 |
| TOS (µmol/L) 8 | 24.6 | 17.8 | 25.3 | 15.2 | 18.6 | 15.1 | 1.187 | 0.018 | 0.932 | 0.921 |
| TAC (mmol/L) 9 | 0.498 | 0.503 | 0.475 | 0.428 | 0.461 | 0.487 | 0.023 | 0.472 | 0.579 | 0.221 |
| Item | CON Ration 1 | IOSe Ration 2 | SEM 3 | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| T0 4 | T21 | T49 | T0 | T21 | T49 | Ration (R) | Day (D) | R × D | ||
| Milk yield (L/d) | 30.9 | 33.4 | 29.4 | 27.1 | 31.4 | 29.4 | 1.771 | 0.583 | 0.770 | 0.216 |
| Milk composition | ||||||||||
| Fat (g/100 g) | 4.56 | 4.31 | 3.58 | 4.05 | 4.56 | 3.54 | 0.115 | 0.657 | 0.019 | 0.423 |
| Protein (g/100 g) | 3.25 | 3.02 | 3.08 | 3.41 | 3.18 | 3.05 | 0.035 | 0.199 | 0.027 | 0.395 |
| Lactose (g/100 g) | 4.77 | 4.72 | 4.80 | 4.95 | 4.68 | 4.81 | 0.019 | 0.244 | 0.026 | 0.130 |
| Ca (mg/100 g) | 118 | 113 | 119 | 122 | 113 | 119 | 3.27 | 0.838 | 0.809 | 0.460 |
| P (mg/100 g) | 85.4 | 81.3 | 91.1 | 86.3 | 83.6 | 87.7 | 1.69 | 0.981 | 0.132 | 0.360 |
| Zn (mg/100 g) | 0.447 | 0.449 | 0.418 | 0.456 | 0.442 | 0.412 | 0.013 | 0.958 | 0.047 | 0.655 |
| Cu (mg/100 g) | 0.007 | 0.003 | 0.003 | 0.006 | 0.004 | 0.004 | 0.000 | 0.675 | <0.001 | 0.120 |
| Se (µg/100 g) | 1.54 | 1.33 | 1.26 | 1.64 | 1.94 | 1.80 | 0.047 | 0.001 | 0.789 | 0.167 |
| Milk pH | 6.61 | 6.79 | 6.65 | 6.64 | 6.67 | 6.62 | 0.023 | 0.418 | 0.611 | 0.177 |
| Item | CON Ration 1 | IOSe Ration 2 | SEM 3 | p-Value |
|---|---|---|---|---|
| Proximate composition and pH | ||||
| Protein (g/100 g) | 5.71 | 7.91 | 0.737 | 0.148 |
| Fat (g/100 g) | 2.15 | 1.49 | 0.269 | 0.261 |
| pH | 4.53 | 4.71 | 0.070 | 0.226 |
| Mineral composition | ||||
| Ca (mg/100 g) | 214.8 | 201.4 | 12.4 | 0.045 |
| P (mg/100 g) | 170.7 | 164.1 | 8.97 | 0.184 |
| Zn (mg/100 g) | 0.874 | 0.848 | 0.038 | 0.650 |
| Cu (mg/100 g) | 0.422 | 0.410 | 0.116 | 0.877 |
| Se (μg/100 g) | 3.86 | 3.56 | 0.418 | 0.538 |
| Physical characters | ||||
| Syneresis (g/100 g) | 11.50 | 14.10 | 1.779 | 0.527 |
| Colorimetric parameters 4 | ||||
| L* | 85.44 | 85.11 | 1.166 | 0.907 |
| a* | −2.97 | −3.23 | 0.104 | 0.252 |
| b* | 5.37 | 5.61 | 1.455 | 0.944 |
| Item | CON Ration 1 | IOSe Ration 2 | SEM 3 | p-Value |
|---|---|---|---|---|
| Proximate composition | ||||
| Dry extract (g/100 g) | 34.65 | 30.68 | 0.869 | 0.013 |
| Protein (g/100 g) | 15.36 | 14.74 | 0.539 | 0.588 |
| Fat (g/100 g) | 12.71 | 9.07 | 2.786 | 0.101 |
| Mineral composition | ||||
| Ca (mg/100 g) | 559.4 | 505.0 | 21.1 | 0.062 |
| P (mg/100 g) | 351.0 | 323.1 | 13.44 | 0.110 |
| Zn (mg/100 g) | 2.81 | 2.83 | 0.25 | 0.923 |
| Cu (mg/100 g) | 1.84 | 0.70 | 0.03 | <0.001 |
| Se (μg/100 g) | 6.00 | 8.29 | 0.83 | 0.001 |
| Physical characters | ||||
| Cheese yield | 17.1 | 20.2 | 1.15 | 0.209 |
| Colorimetric parameters 4 | ||||
| L* | 89.08 | 89.67 | 0.326 | 0.427 |
| a* | −2.62 | −2.60 | 0.046 | 0.857 |
| b* | 7.27 | 8.06 | 0.338 | 0.287 |
| Texture profile | ||||
| Hardness (N) | 10.01 | 8.48 | 0.638 | 0.273 |
| Cohesiveness (dimensionless) | 1.20 | 1.21 | 0.031 | 0.974 |
| Adhesiveness (N s) | −0.70 | −0.23 | 0.177 | 0.192 |
| Elasticity (mm) | 1.21 | 1.23 | 0.004 | 0.009 |
| Chewiness (N mm) | 12.28 | 10.25 | 0.851 | 0.241 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azorín, I.; Madrid, J.; Martínez, S.; López, M.; López, M.B.; López, M.J.; Hernández, F. Can Moderate Levels of Organic Selenium in Dairy Cow Feed Naturally Enrich Dairy Products? Animals 2020, 10, 2269. https://doi.org/10.3390/ani10122269
Azorín I, Madrid J, Martínez S, López M, López MB, López MJ, Hernández F. Can Moderate Levels of Organic Selenium in Dairy Cow Feed Naturally Enrich Dairy Products? Animals. 2020; 10(12):2269. https://doi.org/10.3390/ani10122269
Chicago/Turabian StyleAzorín, Irene, Josefa Madrid, Silvia Martínez, Marina López, María Belén López, Miguel José López, and Fuensanta Hernández. 2020. "Can Moderate Levels of Organic Selenium in Dairy Cow Feed Naturally Enrich Dairy Products?" Animals 10, no. 12: 2269. https://doi.org/10.3390/ani10122269
APA StyleAzorín, I., Madrid, J., Martínez, S., López, M., López, M. B., López, M. J., & Hernández, F. (2020). Can Moderate Levels of Organic Selenium in Dairy Cow Feed Naturally Enrich Dairy Products? Animals, 10(12), 2269. https://doi.org/10.3390/ani10122269