Biofilm Formation by Shiga Toxin-Producing Escherichia coli on Stainless Steel Coupons as Affected by Temperature and Incubation Time

 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Cultivation
2.2. Biofilm Formation
2.3. Crystal Violet Staining
2.4. Enumeration of the Planktonic and Attached Cells
2.5. SEM Analysis
2.6. Statistical Analysis
3. Results
3.1. Growth of the Planktonic Cells in M9 Medium
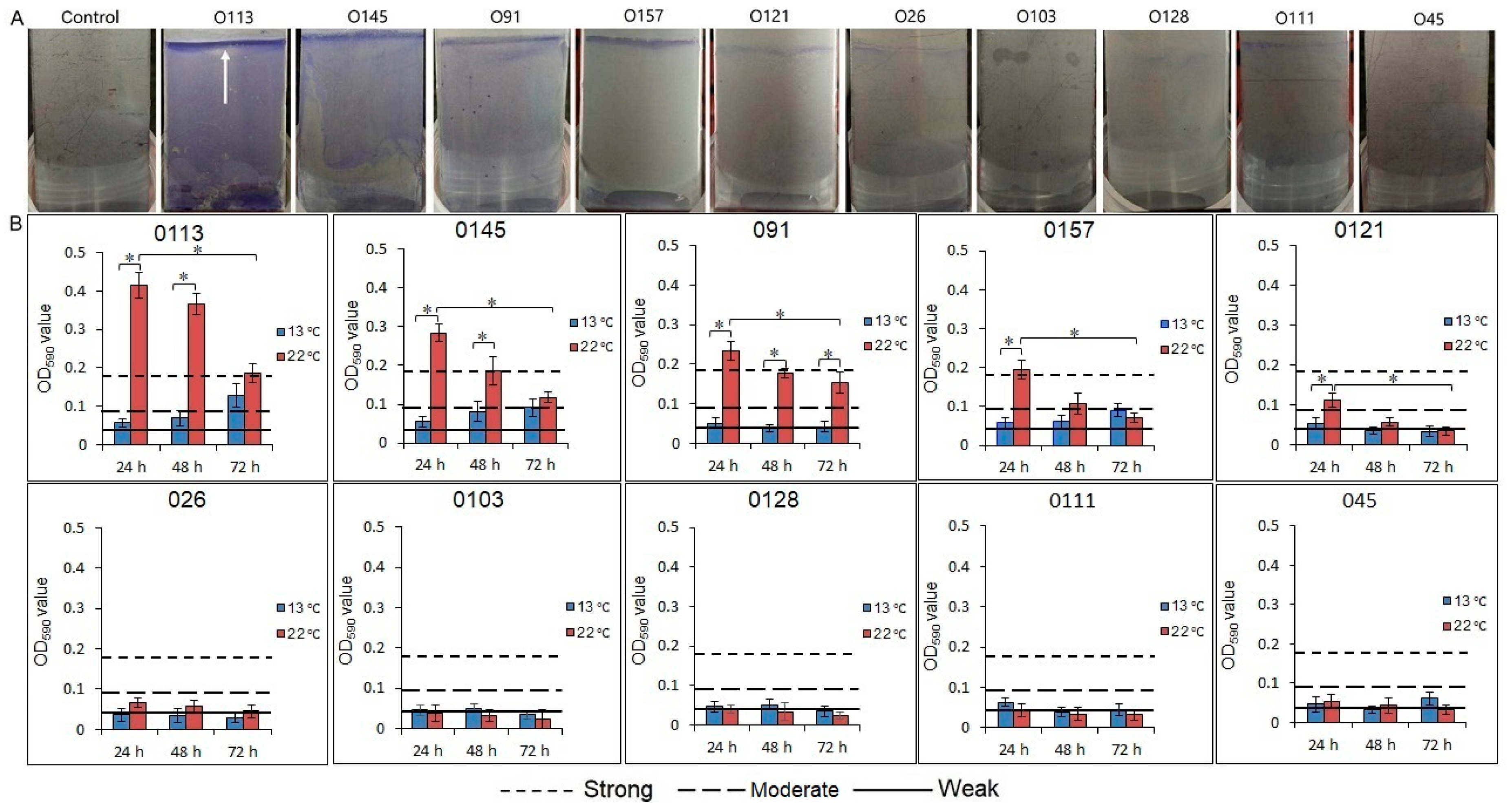
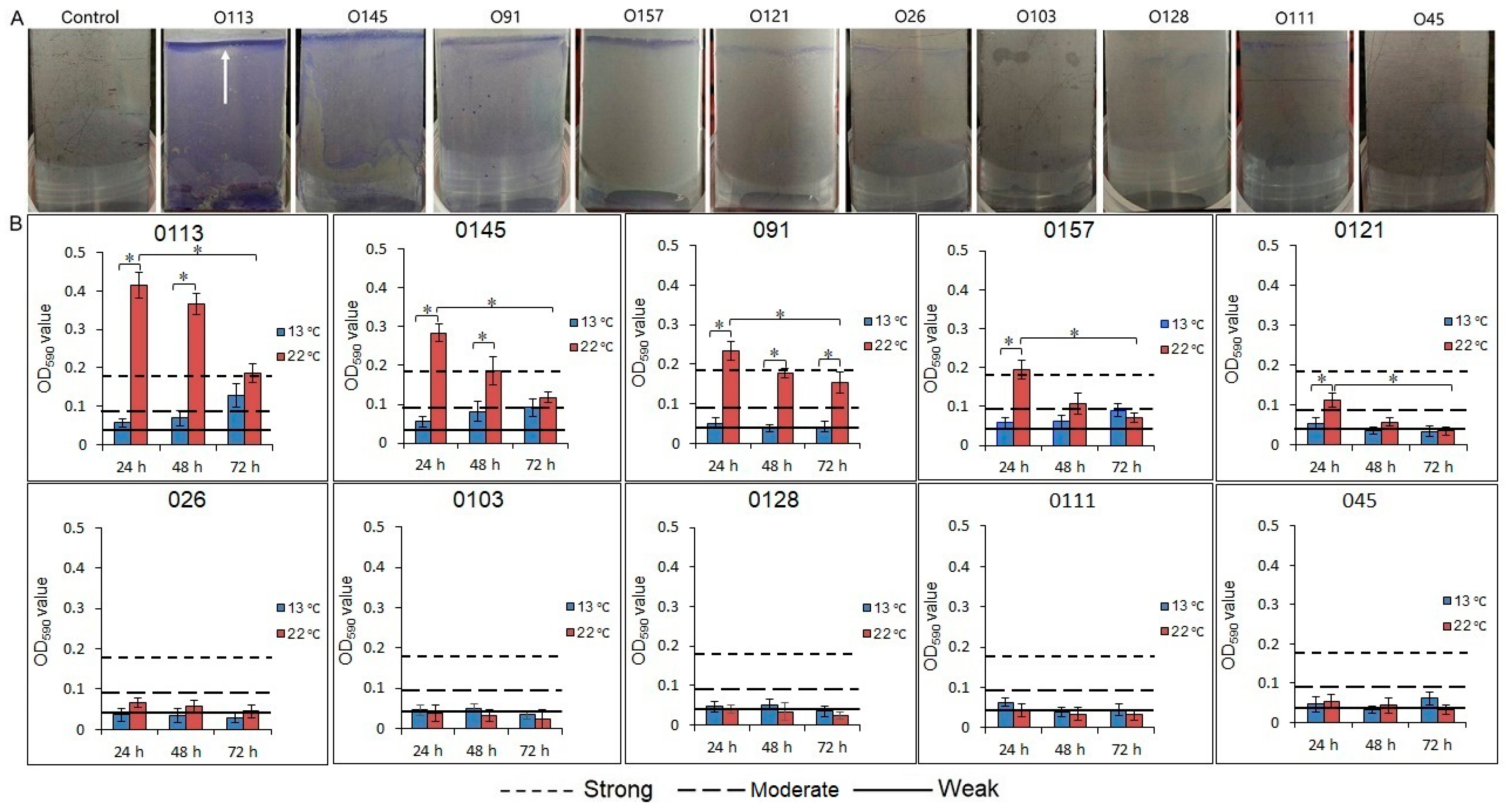
3.2. Biofilm Formation by STEC on a Stainless Steel Surface
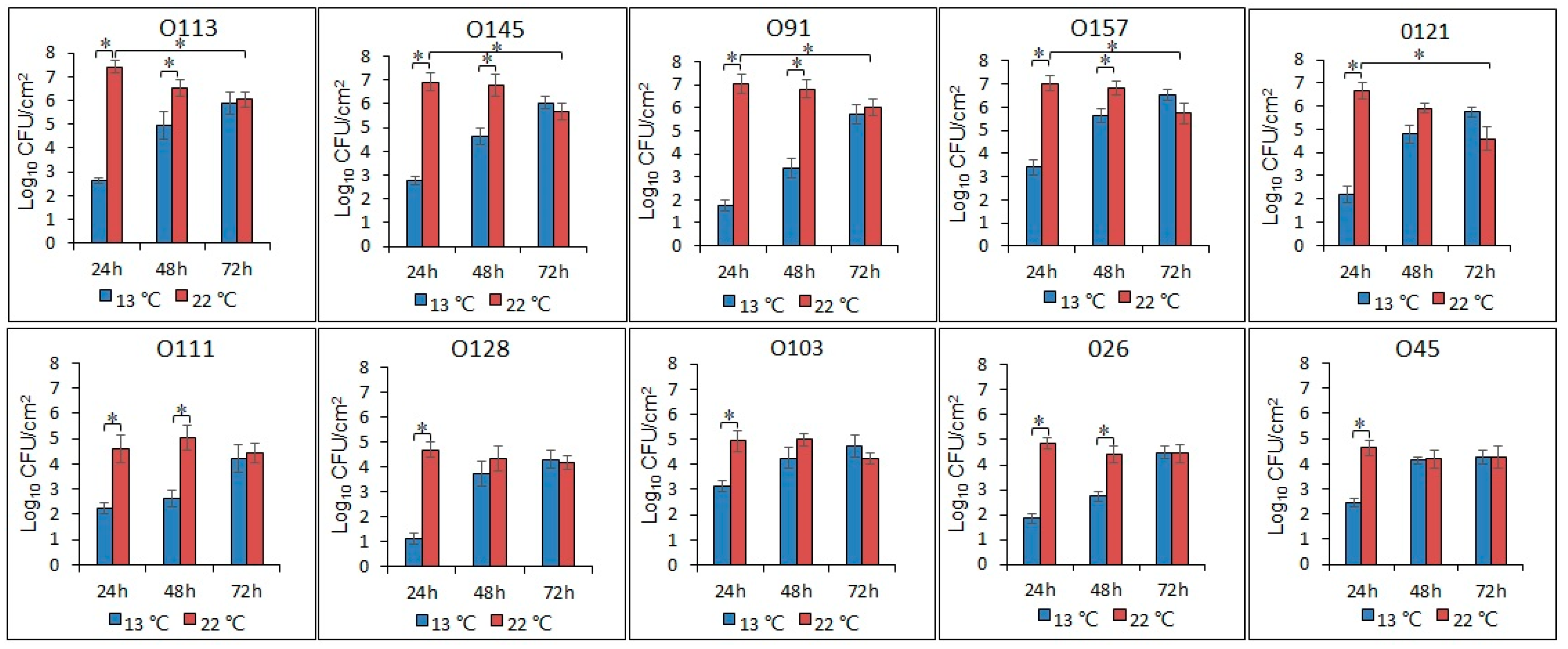
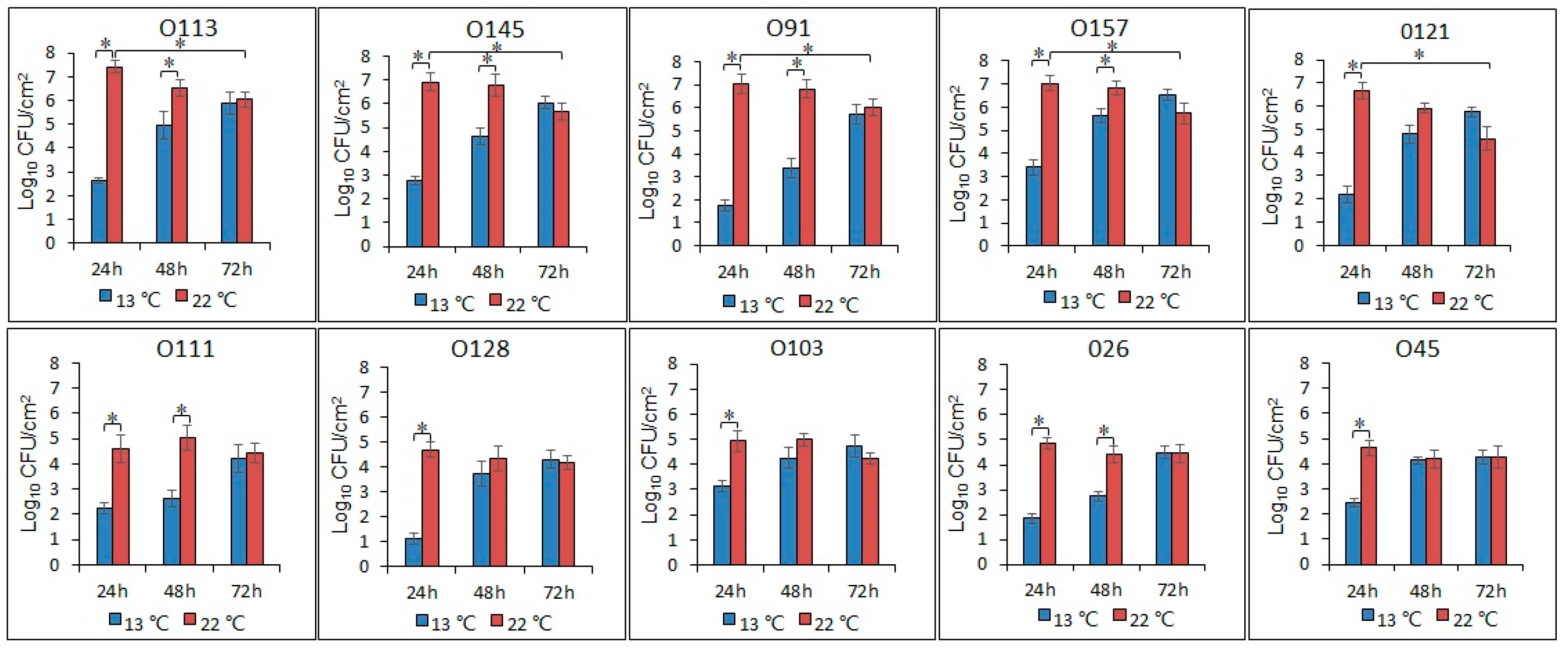
3.3. Enumeration of the Biofilm Cells
3.4. SEM Observation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dzieciol, M.; Schornsteiner, E.; Muhterem, M.; Stessl, B.; Wagner, M.; Schmitz-Esser, S. Bacterial diversity of floor drain biofilms and drain waters in a Listeria monocytogenes contaminated food processing environment. Int. J. Food Microbiol. 2016, 223, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Al-Adawi, A.S.; Gaylarde, C.C.; Sunner, J.; Beech, I.B. Transfer of bacteria between stainless steel and chicken meat: a CLSM and DGGE study of biofilms. AIMS Microbiol. 2016, 2, 340–358. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, X.; Zhang, Q.; Ye, K.; Xu, X.; Zhou, G. Comparison of microbial transfer rates from Salmonella spp. biofilm growth on stainless steel to selected processed and raw meat. Food Control 2015, 50, 574–580. [Google Scholar] [CrossRef]
- Gibson, H.; Taylor, J.; Hall, K.; Holah, J. Effectiveness of cleaning techniques used in the food industry in terms of the removal of bacterial biofilms. J. Appl. Microbiol. 1999, 87, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Vogeleer, P.; Tremblay, Y.D.; Jubelin, G.; Jacques, M.; Harel, J. Biofilm-forming abilities of Shiga toxin-producing Escherichia coli isolates associated with human infections. Appl. Environ. Microbiol. 2016, 82, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Dourou, D.; Beauchamp, C.S.; Yoon, Y.; Geornaras, I.; Belk, K.E.; Smith, G.C.; Nychas, G.-J.E.; Sofos, J.N. Attachment and biofilm formation by Escherichia coli O157:H7 at different temperatures, on various food-contact surfaces encountered in beef processing. Int. J. Food Microbiol. 2011, 149, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Petrova, O.E.; Sauer, K. Escaping the biofilm in more than one way: desorption, detachment or dispersion. Curr. Opin. Microbiol. 2016, 30, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Beuchat, L.R. Biofilm formation by Escherichia coli O157:H7 on stainless steel: Effect of exopolysaccharide and curli production on its resistance to chlorine. Appl. Environ. Microbiol. 2005, 71, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Kim, H.; Beuchat, L.R. Attachment and biofilm formation by Escherichia coli O157:H7 on stainless steel as influenced by exopolysaccharide production, nutrient availability, and temperature. J. Food Prot. 2004, 67, 2123–2131. [Google Scholar] [CrossRef] [PubMed]
- Caprioli, A.; Scavia, G.; Morabito, S. Public health microbiology of Shiga toxin-producing Escherichia coli. In Enterohemorrhagic Escherichia coli and Other Shiga Toxin-Producing E. coli, 1st ed.; Vanessa, S., Carolyn, J.H., Eds.; American Society of Microbiology: Washington, DC, USA, 2015; pp. 263–295. [Google Scholar]
- Fakhouri, F.; Zuber, J.; Frémeaux-Bacchi, V.; Loirat, C. Haemolytic uraemic syndrome. Lancet 2017, 390, 681–696. [Google Scholar] [CrossRef]
- Public Health Agency of Canada. National Enteric Surveillance Program (NESP): Annual summary 2016; Public Health Agency of Canada: Ottawa, ON, Canada, 2016. [Google Scholar]
- Luna-Gierke, R.; Griffin, P.; Gould, L.; Herman, K.; Bopp, C.; Strockbine, N.; Mody, R. Outbreaks of non-O157 Shiga toxin-producing Escherichia coli infection: USA. Epidemiol. Infect. 2014, 142, 2270–2280. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, S.H.; Seo, J.J.; Kee, H.Y.; Kim, M.J.; Seo, K.W.; Lee, D.H.; Choi, Y.H.; Lim, D.J.; Hur, Y.J. An outbreak of inapparent non-O157 enterohemorrhagic Escherichia coli infection. Korean J. Med. 2006, 70, 495–504. [Google Scholar]
- Yatsuyanagi, J.; Saito, S.; Ito, I.J. A case of hemolytic-uremic syndrome associated with Shiga toxin 2-producing Escherichia coli O121 infection caused by drinking water contaminated with bovine feces. Jpn. J. Infect. Dis. 2002, 55, 174–176. [Google Scholar] [PubMed]
- Reid, T.M.S. A case study of cheese associated E. coli O157 outbreaks in Scotland. In Verocytotoxigenic Escherichia coli, 2nd ed.; Duffy, G., Garvey, P., McDowell, D., Eds.; Food and Nutrition: Hartford, CT, USA, 2001; pp. 201–212. [Google Scholar]
- Adator, E.H.; Cheng, M.; Holley, R.; McAllister, T.; Narvaez-Bravo, C. Ability of Shiga toxigenic Escherichia coli to survive within dry-surface biofilms and transfer to fresh lettuce. Int. J. Food Microbiol. 2018, 269, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Luedtke, B.E.; Bosilevac, J.M.; Schmidt, J.W.; Kalchayanand, N.; Arthur, T.M. Escherichia coli O157:H7 strains isolated from high-event period beef contamination have strong biofilm-forming ability and low sanitizer susceptibility, which are associated with high pO157 plasmid copy number. J. Food Prot. 2016, 79, 1875–1883. [Google Scholar] [CrossRef]
- Sharma, V.K.; Kudva, I.T.; Bearson, B.L.; Stasko, J.A. Contributions of EspA filaments and curli fimbriae in cellular adherence and biofilm formation of enterohemorrhagic Escherichia coli O157:H7. PLoS ONE 2016, 11, e0149745. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Bono, J.L.; Kalchayanand, N.; Shackelford, S.; Harhay, D.M. Biofilm formation by Shiga toxin–producing Escherichia coli O157:H7 and Non-O157 strains and their tolerance to sanitizers commonly used in the food processing environment. J. Food Prot. 2012, 75, 1418–1428. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Stanford, K.; McAllister, T.A.; Johnson, R.P.; Chen, J.; Hou, H.; Zhang, G.; Niu, Y.D. Biofilm formation, virulence gene profiles, and antimicrobial resistance of nine serogroups of non-O157 Shiga toxin–producing Escherichia coli. Foodborne Pathog. Dis. 2016, 13, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.D.; Groot, M.N.; Smid, E.J.; Hols, P.; Kleerebezem, M.; Abee, T. Role of cell surface composition and lysis in static biofilm formation by Lactobacillus plantarum WCFS1. Int. J. Food Microbiol. 2018, 271, 15–23. [Google Scholar] [CrossRef]
- Dhowlaghar, N.; Bansal, M.; Schilling, M.W.; Nannapaneni, R.J. Scanning electron microscopy of Salmonella biofilms on various food-contact surfaces in catfish mucus. Food Microbiol. 2018, 74, 143–150. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M.J. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Fuente-Núñez, C.; Reffuveille, F.; Fernández, L.; Hancock, R.E. Bacterial biofilm development as a multicellular adaptation: Antibiotic resistance and new therapeutic strategies. Curr. Opin. Microbiol. 2013, 16, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Prado, D.B.; Fernandes, M.S.; Anjos, M.M.; Tognim, M.C.; Nakamura, C.V.; Machinski Junior, M.; Mikcha, J.M.; Abreu Filho, B.A. Biofilm-forming ability of Alicyclobacillus spp. isolates from orange juice concentrate processing plant. J. Food Saf. 2018, e12466. [Google Scholar] [CrossRef]
- Medrano-Félix, J.A.; Chaidez, C.; Mena, K.D.; Socorro, M.; Castro, N. Characterization of biofilm formation by Salmonella enterica at the air-liquid interface in aquatic environments. Environ. Monit. Assess. 2018, 190, 221. [Google Scholar] [CrossRef] [PubMed]
- Hilbert, L.R.; Bagge-Ravn, D.; Kold, J.; Gram, L.J. Influence of surface roughness of stainless steel on microbial adhesion and corrosion resistance. Int. Biodeterior. Biodegr. 2003, 52, 175–185. [Google Scholar] [CrossRef]
- Giaouris, E.; Chorianopoulos, N.; Nychas, G.J. Effect of temperature, pH, and water activity on biofilm formation by Salmonella enterica enteritidis PT4 on stainless steel surfaces as indicated by the bead vortexing method and conductance measurements. J. Food Prot. 2005, 68, 2149–2154. [Google Scholar] [CrossRef]
- Chia, T.; Goulter, R.; McMeekin, T.; Dykes, G.; Fegan, N.J. Attachment of different Salmonella serovars to materials commonly used in a poultry processing plant. Food Microbiol. 2009, 26, 853–859. [Google Scholar] [CrossRef]
- Biscola, F.T.; Abe, C.M.; Guth, B.E. Determination of adhesin gene sequences in, and biofilm formation by, O157 and Non-O157 Shiga toxin-producing Escherichia coli strains isolated from different sources. Appl. Environ. Microbiol. 2011, 77, 2201–2208. [Google Scholar] [CrossRef] [PubMed]
- Dewanti, R.; Wong, A.C. Influence of culture conditions on biofilm formation by Escherichia coli O157:H7. Int. J. Food Microbiol. 1995, 26, 147–164. [Google Scholar] [CrossRef]
- Nguyen, H.; Yang, Y.; Yuk, H. Biofilm formation of Salmonella Typhimurium on stainless steel and acrylic surfaces as affected by temperature and pH level. LWT-Food Sci. Technol. 2014, 55, 383–388. [Google Scholar] [CrossRef]
- Sinde, E.; Carballo, J.J. Attachment of Salmonella spp. and Listeria monocytogenes to stainless steel, rubber and polytetrafluorethylene: The influence of free energy and the effect of commercial sanitizers. Food Microbiol. 2000, 17, 439–447. [Google Scholar] [CrossRef]
- Bakterij, A.J. An overview of the influence of stainless-steel surface properties on bacterial adhesion. Mater. Tehnol. 2014, 48, 609–617. [Google Scholar]
- Hood, S.K.; Zottola, E.A. Adherence to stainless steel by foodborne microorganisms during growth in model food systems. Int. J. Food Microbiol. 1997, 37, 145–153. [Google Scholar] [CrossRef]
- Somers, E.B.; LEE, A.C. Efficacy of two cleaning and sanitizing combinations on Listeria monocytogenes biofilms formed at low temperature on a variety of materials in the presence of ready-to-eat meat residue. J. Food Prot. 2004, 67, 2218–2229. [Google Scholar] [CrossRef]
- Frank, J.F. Microbial attachment to food and food contact surfaces. In Advances in Food and Nutrition Research; Academic Press: Cambridge, MA, USA, 2001; Volume 43, pp. 319–370. [Google Scholar]
- Andersen, T.E.; Kingshott, P.; Palarasah, Y.; Benter, M.; Alei, M.; Kolmos, H.J. A flow chamber assay for quantitative evaluation of bacterial surface colonization used to investigate the influence of temperature and surface hydrophilicity on the biofilm forming capacity of uropathogenic Escherichia coli. J. Microbiol. Methods 2010, 81, 135–140. [Google Scholar] [CrossRef]
- Cookson, A.L.; Cooley, W.A.; Woodward, M.J. The role of type 1 and curli fimbriae of Shiga toxin-producing Escherichia coli in adherence to abiotic surfaces. Int. J. Food Microbiol. 2002, 292, 195–205. [Google Scholar] [CrossRef]
- Walker, S.L.; SOJKA, M.; Dibb-fuller, M.; Woodward, M.J. Effect of pH, temperature and surface contact on the elaboration of fimbriae and flagella by Salmonella serotype Enteritidis. J. Med. Microbiol. 1999, 48, 253–261. [Google Scholar] [CrossRef]
- Bonaventura, G.; Piccolomini, R.; Paludi, D.; Dorio, V.; Vergara, A.; Conter, M.; Ianieri, A.J. Influence of temperature on biofilm formation by Listeria monocytogenes on various food-contact surfaces: Relationship with motility and cell surface hydrophobicity. J. Appl. Microbiol. 2008, 104, 1552–1561. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, S.F.; Li, Y.; Xu, H.; Qin, L.; Tay, J.H. The influence of cell and substratum surface hydrophobicities on microbial attachment. J. Biotechnol. 2004, 110, 251–256. [Google Scholar] [CrossRef]
- Jain, S.; Chen, J.J. Attachment and biofilm formation by various serotypes of Salmonella as influenced by cellulose production and thin aggregative fimbriae biosynthesis. J. Food Prot. 2007, 70, 2473–2479. [Google Scholar] [CrossRef]
- Cogan, T.A.; Rgensen, F.; Lappin-Scott, H.M.; Benson, C.E.; Woodward, M.J.; Humphrey, T.J. Flagella and curli fimbriae are important for the growth of Salmonella enterica serovars in hen eggs. Microbiology 2004, 150, 1063. [Google Scholar] [CrossRef] [PubMed]
- Paton, A.W.; Woodrow, M.C.; Doyle, R.M.; Lanser, J.A.; Paton, J.C. Molecular characterization of a Shiga toxigenic Escherichia coli O113: H21 strain lacking eae responsible for a cluster of cases of hemolytic-uremic syndrome. J. Clin. Microbiol. 1999, 37, 3357–3361. [Google Scholar] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Bumunang, E.W.; Stanford, K.; Bie, X.; Niu, Y.D.; McAllister, T.A. Biofilm Formation by Shiga Toxin-Producing Escherichia coli on Stainless Steel Coupons as Affected by Temperature and Incubation Time. Microorganisms 2019, 7, 95. https://doi.org/10.3390/microorganisms7040095
Ma Z, Bumunang EW, Stanford K, Bie X, Niu YD, McAllister TA. Biofilm Formation by Shiga Toxin-Producing Escherichia coli on Stainless Steel Coupons as Affected by Temperature and Incubation Time. Microorganisms. 2019; 7(4):95. https://doi.org/10.3390/microorganisms7040095
Chicago/Turabian StyleMa, Zhi, Emmanuel W. Bumunang, Kim Stanford, Xiaomei Bie, Yan D. Niu, and Tim A. McAllister. 2019. "Biofilm Formation by Shiga Toxin-Producing Escherichia coli on Stainless Steel Coupons as Affected by Temperature and Incubation Time" Microorganisms 7, no. 4: 95. https://doi.org/10.3390/microorganisms7040095
APA StyleMa, Z., Bumunang, E. W., Stanford, K., Bie, X., Niu, Y. D., & McAllister, T. A. (2019). Biofilm Formation by Shiga Toxin-Producing Escherichia coli on Stainless Steel Coupons as Affected by Temperature and Incubation Time. Microorganisms, 7(4), 95. https://doi.org/10.3390/microorganisms7040095